

Abstract
This study explores the conceptual history of systems biology and its impact on philosophical and scientific conceptions of reductionism, antireductionism and emergence. Development of systems biology at the beginning of 21st century transformed biological science. Systems biology is a new holistic approach or strategy how to research biological organisms, developed through three phases. The first phase was completed when molecular biology transformed into systems molecular biology. Prior to the second phase, convergence between applied general systems theory and nonlinear dynamics took place, hence allowing the formation of systems mathematical biology. The second phase happened when systems molecular biology and systems mathematical biology, together, were applied for analysis of biological data. Finally, after successful application in science, medicine and biotechnology, the process of the formation of modern systems biology was completed.
Systems and molecular reductionist views on organisms were completely opposed to each other. Implications of systems and molecular biology on reductionist–antireductionist debate were quite different. The analysis of reductionism, antireductionism and emergence issues, in the era of systems biology, revealed the hierarchy between methodological, epistemological and ontological antireductionism. Primarily, methodological antireductionism followed from the systems biology. Only after, epistemological and ontological antireductionism could be supported.
Keywords: Systems biology, Molecular biology, Reductionism, Antireductionism, Emergence, General systems theory, Nonlinear dynamics
1. Introduction
Systems biology is a strategy or a way of thinking how to investigate biological organism. Systems biology studies the organisms as integrated systems composed of dynamic and interrelated genetic, protein, metabolic and cellular components with the help of biology, mathematics, technology and computer science (Auffray et al., 2003, Friboulet and Thomas, 2005, Bruggeman and Westerhoff, 2006). O’Malley and Dupré (2005) classified systems biology on pragmatic systems biology, whose emphasis is on the analysis of molecular interactions at the general level and system–theoretical biology which considers the systemic principles. Both parts of systems biology insist on mathematical modeling (O’Malley and Dupré, 2005).
When and how does systems biology emerged? According to Westerhoff and Palsson (2004) systems biology has two historical roots. The first and the most frequently mentioned root, relates to the discovery of the structure and function of the genetic material, as well as on the methods of gene manipulation. Second root is related to the thermodynamic aspects of living organisms introduced in biology during the 40s of the 20th century.
This article discusses three phases of systems biology development. First phase included the transformation of molecular biology into systems molecular biology. This phase referred to the discovery of the structure and function of genes and genetic engineering (Westerhoff and Palsson, 2004). Since 1953, molecular biologists discovered the structure and function of genes and finally, at the beginning of 21st century, deciphered human genome. In postgenomic era, the goal of molecular biology has changed. The search for explanation how complex molecular pathways and networks supported biological structure and function has become a central issue of molecular biology. Shifting from single molecule to molecular network approach definitely marked the emergence of systems molecular biology. The second phase, the development of systems-mathematical biology, referred to the general systems theory (GSS) and nonlinear dynamics of living organisms. Third phase, followed after the convergence of molecular systems biology and systems mathematical biology, relates to the development of systems based medicine, biotechnology and drug discovery.
Systems biology influenced the longstanding reductionism–antireductionism debate. Systems biology view on biology, as being holistic and integrative, was quite opposed to molecular reductionist position (Strange, 2005, Ahn et al., 2006). Reductionists claimed that every biological phenomenon could be explained in terms of molecular biology and ultimately physics. The physics is supposed to be only fundamental science of natural world. Hence, theories and laws of other natural sciences should be explainable by fundamental theories and laws of physics. All sciences should be a part of one unified science. In the 20th century, supporters of the unity of science used theory reduction for the unification project (Bechtel and Hamilton, 2007). Systems biology changed the way we think about the concept of emergence. As being the most important concept discussed in reductionism–antireductionism debate, it deserves necessary reexamination in the light of modern systems biology.
This article deals with conceptual history of systems biology and its impact on important scientific and philosophical issues. First I will argue that systems mathematical biology has two historical roots, general systems theory and nonlinear dynamics. Their convergence leads to modern systems mathematical biology. Afterward, I argued that contemporary systems biology rested on the convergence between systems of molecular biology and systems mathematical biology. The above mentioned convergence enabled application of systems approach to science, medicine, and biotechnology. Only after the completion of all historical steps, the modern systems biology was born. Then, it will be examined the impact of systems biology on longstanding philosophical and scientific conceptions of reductionism, antireductionism and emergence.
2. The birth of systems biology: molecular biology, general systems theory, and nonlinear dynamics
What are the differences between molecular biologist and systems biologist view on biological systems? Ahn and colleagues have made some of the crucial distinction between reductionist and system-oriented view on the biology (Ahn et al., 2006). When it comes to the underlying principles, the two approaches differ in understanding how biological systems behave. In reductionist view, the behavior of biological systems can be explained by the properties of components. The system oriented approach insisted that biological systems have emergent properties that only can have a system as a whole and not its constituent parts. The metaphors used by these two approaches are quite different: machine/magic bullet versus network. Considering the approach, reductionism gives explanatory significance only to one factor, while the system biology considers a number of factors in order to describe the behavior of dynamic system. In reductionist approach the critical factors are directly determining while in the system view it depends on time, space and context. According to reductionist approach, the characteristics of models that explain the behavior of the system are linearity, predictability and determinism. Contrastingly, the systems approach insists on nonlinearity, sensitivity to initial conditions, stochasticity (probabilism) and chaotic behavior. Systems-oriented overview of the concept of health in medicine implies robustness, adaptability/plasticity and homeodynamic (dynamic understood homeostasis). The reductionist approach emphasized the normalcy, reducing exposure to risk and static homeostasis. Obviously reductionist and systems view on biology are quite different. When and how does this Copernican revolution in biology happen?
From the early days, molecular biology treated the components of a cell as static and isolated, neglecting the dynamical interaction between them. But, the methodological aspect of reductionist approaches had a good side. After all, this approach influenced the development of so many diverse molecular techniques. Without them scientists would not be able to identify genes, molecules and processes, and surely human genome project would have never happened. Hence, the systems molecular biology would not exist. The negative side of reductionist molecular approach referred to lack of holistic and integrative knowledge of biological processes. As molecular biologists acquire vast number of facts about DNA, RNA and proteins, it becomes more difficult to give explanation of what organism is. Thus, how to connect molecular processes with higher level biological phenomena becomes the issue. Therefore, instead of concentrating on the physiochemical and biological properties of single or limited number of molecules, the priority has become to find out how the molecular networks assembled and function. Finally, the completion of human genome project directed molecular biology toward systems molecular biology. By developing the systems molecular biology, molecular biologist made the first step toward contemporary systems biology.
During the second half of 20th century systems mathematical biology existed in parallel with molecular biology. The term systems biology was introduced by Mihajlo Mesarovic in 1968 (Mesarovic, 1968, Mesarovic et al., 2004), but there are opinions that the term may be introduced in the 1920s by Ludwig von Bertalanffy, the father of general systems theory (Drack, 2007). However, Mesarovic applied general systems theory in order to discover how biological objects relate, rather than what they are composed of (Mesarovic, 1968, Mesarovic et al., 2004). Before the pioneering work of Mihajlo Mesarovic, general systems theory was developed by Bertalanffy (Drack, 2009). Bertalanffy developed what he called a “system theory of life” contributing to the development of theoretical biology (Drack, 2009). His theory was an attempt to scientifically study the organism as “wholeness”. Unsatisfied by mechanists and vitalist doctrine that implies metaphysics, Bertalanffy found the solution in the mathematical modeling of biological organism (Drack, 2009). General systems theory that he developed, concentrated on the topics such as self-regulating systems, i.e. systems self-correcting through feedback, the concepts widely used in physiology and engineering science. Since it was introduced, general systems theory was successful in modeling the biochemical reaction networks, homeostasis, neural, immune, cardio and endocrine system function (Ederer et al., 2003). GSS theory found its important place in modern systems neurobiology as a part of neural networks research and modeling. Application of general systems theory in biology was the first step in formation of systems mathematical biology. The next step was the development and application of nonlinear dynamics.
Chaos theory is mathematical theory that yet has not been fully developed (Oestreicher, 2007). The beginning of chaos theory dates back to Henry Poincare and his studies of sensitivity to initial conditions of physical systems (Oestreicher, 2007). The important contributions to chaos theory gave Andrei Kolmogorov with the work on statistics of dynamical systems (Oestreicher, 2007). Another contribution to chaos theory comes from the work of Edward Lorenz, David Ruelle and many others researchers in the field (Oestreicher, 2007). A major advance in the development of nonlinear dynamics has been the introduction of the concepts of fractals by Benoit Mandelbrot in 1973. What are fractals? Simple process that is repeated endlessly becomes a very complex process, and this fact is the basis for the description of fractals in nature (Naik et al., 2011). Fractal is a shape that retains its structural detail despite scaling and this is the reason why complex object could be described by a single number: the fractal dimension (FD) (Arle and Simon, 1990). The next big step in nonlinear theory development was introduction of Higuchi fractal dimension (HFD) (Higuchi, 1988). The simplest definition of fractal dimension is: FD represents nonlinear measure of the complexity in time domain. After HFD was introduced, few other methods for FD calculation have been developed, but HFD provides the most accurate estimate of the FD (Esteller et al., 2001).
How nonlinear dynamics relates to systems mathematical biology? As mentioned, systemic view on biology includes nonlinearity, sensitivity to initial conditions, stochasticity (probabilism) and chaotic behavior (Ahn et al., 2006). These concepts come from the hearth of nonlinear dynamics. Nonlinear dynamics is the theory of nonlinear systems and processes (Klonowski, 2007). Linearity and nonlinearity, in mathematics, can be defined in the context of relations between cause and effect (linear and nonlinear causality) (Scott, 2002). Linearity implies the existence of proportionality or additivity. It means that system responds proportionately to the applied input (Higgins, 2002). Nonlinearity of the system, physical or biological, means that the properties of the constituent components are different than the properties of the system itself (Higgins, 2002). Nonlinear systems are characterized by deterministic (dynamic) chaoticity (Stam, 2005). Biological systems and processes are inherently complex, nonlinear and nonstationary (Paraschiv-Ionescu and Aminian, 2009). The behavior of nonlinear systems depends on the initial conditions and that is the reason why it is difficult to predict their behavior (Stam, 2005). The stochastic (random) behavior of nonlinear systems is ostensible, because their characteristics can be predicted in the short time series (Leith, 1996). In the long term, nonlinear systems are characterized by unpredictability as a consequence of sensitivity to initial conditions (Leith, 1996).
Biological processes characterize time variability (nonstationarity). Nonstationarity is the result of the interaction of internal and external factors and regulatory processes that maintain homeostasis of the organism (Eke et al., 2002). The nonlinear behavior, besides being characteristic of complex wholeness (cell, brain), also could be found in different subsystems of the cell. Thus, membrane potentials, spontaneous bioelectric activity of neurons, and ionic currents caused by movement of ions through the ion channels are also chaotic processes (Liebovitch and Todorov, 1996, Abarbanel et al., 1996, Schütt et al., 2002). Therefore, all levels of central nervous systems organization, from molecules to the brain, shows nonlinear behavior.
Successful application of nonlinear dynamics in analysis of biological hierarchies finished the formation of systems mathematical biology. The stage was set for convergence of systems molecular and systems mathematical biology. Conceptually, convergence took place when nonlinear dynamics and general systems theory was introduced for gene, protein and metabolic networks analyses. This was the moment when the systems molecular biology and systems mathematical biology converge, in the second step of systems biology development. Therefore, nonlinear modeling of gene regulatory networks, from time series gene expression data is one of the most difficult problems in systems biology (Hasty et al., 2001, Mazur et al., 2009, Meister et al., 2013). Biological structure and function depends on regulatory interactions between many genes (Meister et al., 2013). Description of gene regulatory network interference depends on efficient nonlinear models of vast number of genome-wide measurements.
Finally, after successful application in science research, medicine and biotechnology, systems biology was completely shaped. Thus, understanding the origin of neurodegenerative, cancer, inflammatory and genetic diseases is only possible by systems biological holistic approach. Identification of differences between cancerous and healthy cell is difficult because of many different molecules from parallel signal transduction pathways (Hornberg et al., 2006). Systems biology quantification and modeling of regulatory circuits, nonlinear reaction kinetics of biochemical processes and cross-talk between pathways determine the site at which oncologist should intervene (Hornberg et al., 2006). Similarly, the complex mechanism of neurodegenerative disease could only be deciphered by systems biology approach (Noorbakhsh et al., 2009). Surely, there are so many examples of successful application of contemporary systems biology that there is no enough space to mention all of them (Fig. 1).
Figure 1.

Schematic diagram that represents the three phases of contemporary systems biology development.
3. Reductionism and antireductionism in the era of systems biology
The appearances of molecular biology in the early 50s of the 20th century marked a major milestone in efforts to connect biology with the chemical and physical sciences. Scientists and philosophers believed that molecular biology supposed to be a bridge between biology and physics. Hence, if it was possible to reduce biological phenomena on corresponding molecular structures and processes, then the process of reduction on physics will be almost completed (Rosenberg, 1985). This reductionist reasoning was a part of larger logical-positivist project of science unification under the auspices of physics (Faye, 2014). From such a perspective, it was inevitable that reductionism as one of the key problems of philosophy of science poured on theoretical and practical field of biology.
What is reduction and reductionism in biology? The basic assumption underlying reduction is the possibility to prove that certain things are nothing more than another type of things. John Searle stated that this claim can simply be expressed by using the relation of identity, which could be called “nothing-but” (Searle, 1992). One of the essential classifications of reductionism, applied to the problems of living world, was introduced by the famous geneticist and evolutionary biologist Francis Ayala (Ayala, 1974). He made a distinction between the ontological, epistemological and methodological reductionism in biology. Ontological reductionism stated that physical–chemical entities and processes are essentially same for inanimate nature and living organisms. The difference between organic life and inanimate matter can only be explained by the specific organization of living systems. In the case of epistemological reductionism, laws, generalizations, explanations and theories of biology could be ultimately reduced, by certain formal rules, to more general and fundamental laws and theories of physics. Methodological reductionism is related to the acceptance of certain scientific-research strategies. In biology methodological reductionism are related to the implementation for example molecular, cellular or systemic approach to research.
The ultimate pretension of reductionist is to prove that an assumption of ontological reductionism is true. But to defend this view one must accept certain metaphysical position, particularly one form of substantial monism called materialistic monism. First mention of substantial monism could be found in presocratic philosophy and later in the works of Spinoza and Hegel. There are various forms of monism and the most basic classification is on idealistic and materialistic monism. Idealistic monism is the view that the basis of reality is mind or spirit. On the other hand materialist monism assumes that only physical reality exists. In contemporary philosophy of mind frequently mentioned position is materialistic monism (physicalism), the view according to which the only reality is physical and all mental phenomena could be reduced to the physical (Crane, 2000). If ontological reductionism exceeded the domain of scientific thinking, the next line of reductionist defense will be the acceptance of epistemological reductionism. But as we are going to see epistemological reductionism is also a fertile ground for the supporters of antireductionism.
What is epistemological reductionism and antireductionism? The most detailed classification of epistemological reductionism and antireductionism in genetics and biology was presented by the Russell Vance. His classification was based on those made by Philip Kitcher in 1984 (Vance, 1996). According to Vance and Kitcher strong (eliminative) reductionism, admits the existence of independent, but not autonomous levels of explanation in biology except in molecular biology. Therefore, the explanation of biological phenomena can always find an adequate explanation of molecular biology and, ultimately, physics. In contrast to the strong, weak reductionism recognizes the existence of autonomous levels of explanation, but their relationship is asymmetrical, thus reserving the possibility of reducing genetics and whole biology to molecular biology (Vance, 1996).
Besides the reductionist, Vance mentioned the following antireductionist positions: strong, weak and heroic antireductionism. Strong antireductionism admitted the existence of autonomous levels of explanation in biology, and the relationship between the levels are necessarily symmetric and neither level of explanation is privileged. Weak antireductionism proposed, that in addition to the autonomy of levels, the relationship between the levels, not necessarily, could be symmetrical.
Vance opposite to Kitcher stands out for heroic antireductionism, a position which is actualized in the era of systems biology. Heroic antireductionism stated that there are no independent levels of explanation and that reducing and reduced theories should be considered as complementary components of a unified whole. Heroic antireductionism supposed to defend the view that molecular and Mendelian genetics are fundamentally united with the help of two central practices of modern genetics, cloning and the human genome project (Vance, 1996).
In the second half of 20th century, methodological reductionism was a widely accepted view. For molecular biologist, only correct and meaningful approach to the study of life processes was those based on experiments and techniques of molecular biology. Molecular paradigm was supposed to be the foundation for all other subfields of biology. Therefore, physiology, ecology and genetics should be transformed into molecular physiology, molecular genetics and molecular ecology. After the development of systems biology and systems approach, molecular research has become an integral part of systems paradigm. I would say that there was no shifting between molecular and systems paradigm. In post-genomic era the molecular research have continued to take place. Today, it is the starting point for systems biology research. The best is to describe situation as research integration. Good example of research integration could be found in scientific endeavor to define gene. In pre-genomic era, gene was defined as a linear sequence of DNK that provides instructions for hereditary character. But, post-genomic definition of gene is far more complex and besides different segments of genomes (DNA and RNA), includes protein molecules. Even, in order to define gene additional information on the initial conditions (initial state interaction within the network) should be provided (Noble, 2010). These conditions only could be specified by applied nonlinear dynamic measures, an important part of systems-mathematical biology (Noble, 2010). In addition, gene could be understood only after the functional interpretation of the relationship of genotype and phenotype was done (Noble, 2010). This was only possible to achieve, if Mendel’s classic conception of the gene, which is used in population and evolutionary genetics, was accepted (Stotz, 2004, Griffiths and Stotz, 2006). The research practice in an era of systems biology includes both molecular and Mendelian approach, thus uniting molecular and classical Mendelian genetics.
At this point I would like to introduce the term heroic methodological antireductionism. The heroic methodological antireductionism could be defended by the contemporary systems biology. It is feasible considering the capacity of systems biology to unify the practice of genetics and molecular biology with mathematical and computational approaches. Vance heroic antireductionism is wrongly called epistemological because it was based on methodological approach. The more appropriate term should be heroic methodological antireductionism. If the objective of heroic antireductionism is to provide an explanation that reducing and reduced theories are complementary, then practice is not an appropriate way to achieve that. The search for explanation is in the domain of epistemology. The practice of contemporary systems biology first implies heroic methodological antireductionism. Epistemological consequence can be implemented only after methodological consideration. Therefore, systems biology implies the hierarchy of antireductionist position. The most directly supported is heroic methodological antireductionism, and only then, one can consider epistemological antireductionism. At the end of hierarchy is ontological antireductionism, that can be introduced only if it possible to defend the specific ontology implied by systems biology. Whatever the type of antireductionism is accepted, the independence of biology from physics, could be defended (Fig. 2).
Figure 2.
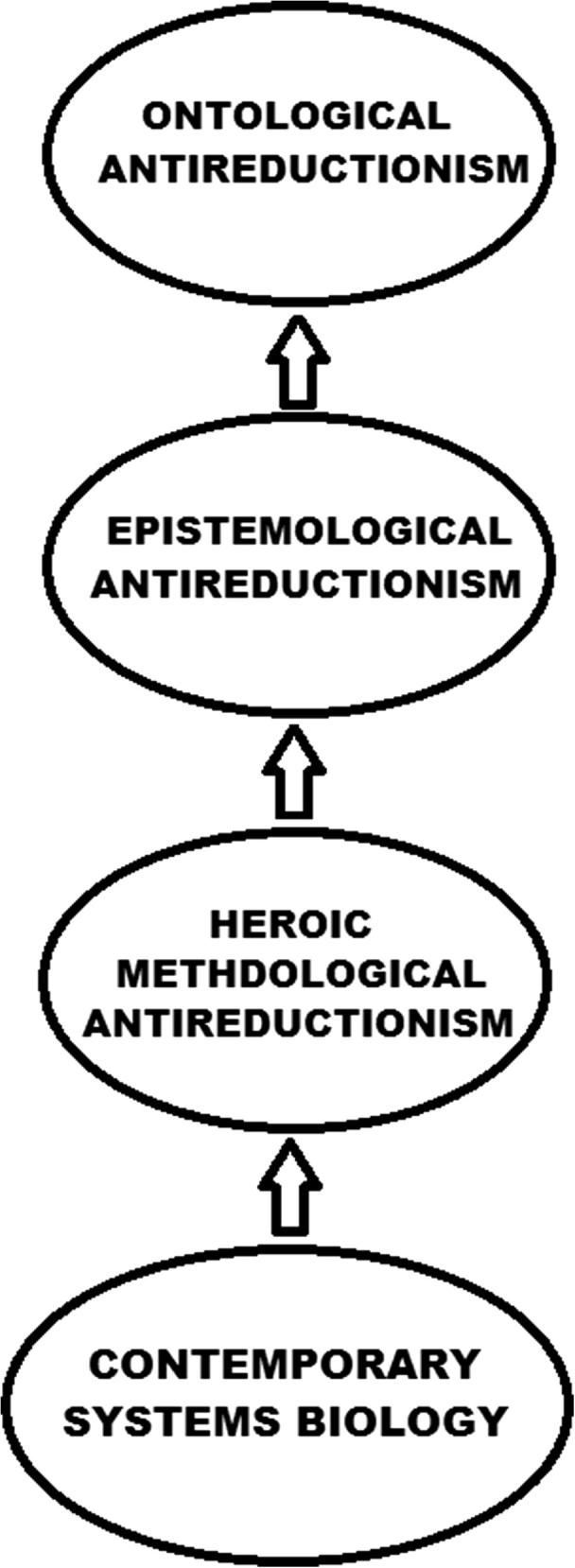
The proposed hierarchy established between the three types of antireductionism. Heroic methodological antireductionism is directly supported by the practice of systems biology. Only after that, epistemological and ontological antireductionisms are supported.
4. Emergence: a systems biology view
How does systems biology influence scientific and philosophical debate about emergence? Do concepts of emergence reinforce antireductionism in the era of systems biology? The basic assumption of emergence is that the natural world is organized hierarchically from the level of subatomic particles to the ecosystem, biosphere etc. (Mazzochi, 2008). Each of the proposed hierarchical level has certain emergent principles that do not appear in the lower level of the organization, and therefore it would be impossible to explain the functioning of a biological organism using only physicochemical principles.
Silberstein and McGeever (1999), distinguishes between two types of emergence, ontological and epistemological emergence. Ontological emergence refers to the properties of the system or a whole, having “causal capacities” irreducible to the intrinsic “causal capacity” of the system parts (Silberstein and McGeever, 1999). Ontological emergence is often present in the philosophy of mind and psychology debates. Today, the most actual view on emergence is emergent materialism, especially its sub-branches called as emergent property dualism (Gulick, 2001). Property dualist theories accept physical substance, but assert two different kinds of properties, mental and physical (Gulick, 2001). Mental properties supervene on lower levels constituted by fundamental physical principles. Specific organization of material substance is the cause of “emergence” of novel properties. The most puzzling question is how mental properties “emerge” from more fundamental levels. Well, we will not discuss that issue, but certainly emergence is poorly understood and it is one of the central problems in modern philosophy and science.
Another important issue is ontological emergence connections with vitalism. When vitalism was rejected as non-scientific, the only reasonable aspect of vitalism that survives was concept of emergence (Emmeche et al., 1997). Unlike vitalism, emergence postulates that the creation of new properties is invariant to the substance involved (Emmeche et al., 1997). Therefore, emergence is often defined as an ontological middle course, between monist materialist and dualist ontologies (Bitbol, 2007). Besides biology, this concept could be applied to inanimate world. But it is necessary to address differences between emergence and vitalism. In the heart of vitalism is assumption of substantial dualism. Vitalist believed that all living organism are driven by inexplicable “vital forces”. “Vital forces” could not be described by means of physical chemistry or any other scientific approach. The essence of life is immaterial and this is the reason why biology is quite different than physical sciences of non-living world. As not being scientific concept, vitalism could not have a significant role in scientific-based antireductionist thinking. Maybe it could find its place in metaphysical and history of philosophy discussions. Contrastingly to the vitalist worldview, emergent property dualist accepted the view that the world is only made by physical substance.
How can systems biology support for example emergent property dualism? The question is quite practical and it is based on the hypothesis that functioning of organism as a whole could be understood after discovering all possible interactions between constitutive elements of organism. Then, the aim of systems biology would not be the only quantification of interaction between molecules, but also to predict and describe (quantitatively and qualitatively) emergent properties on the higher level. Before that step systems biologist should recognize when structural and functional unity of constitutive parts produce something novel and quite different. In that case systems biology methods would allow predictions of emergent properties with high accuracy. Still, we are not able to recognize when and how the vast number of molecular networks produce highly organized and functional single cell, not to mention the whole human organism. However, if ontological emergence is true, then it would be possible to defend strong antireductionism. Looking from a today’s scientific perspective the best way to argue in favor of antireductionism is by accepting the epistemological emergence.
Epistemological emergence occurs in situations when we do not have satisfactory explanation of the higher-level phenomena by those that exist at a lower level (Silberstein and McGeever, 1999). These problems are the result of deficiencies in our cognitive powers but also in the methodological limitations of science. Epistemological emergence includes any explanation of the higher hierarchical levels with the help of modeling or some functional explanation (Silberstein and McGeever, 1999). Hence, accepting the systems biology as powerful methodological tool, researchers can overcome the limitations of empirical research. But still the question is how to compensate deficiencies of our cognitive powers? Systems biology studies often include large sets of data. For the purpose of analysis, researchers and mathematicians developed complex algorithms. However, to comprehend and connect all data in one coherent and functional whole, science must rely on human cognitive powers. For example, in order to find out how drugs mutually interact, a metric “S-score” that measures the strength of network connection between drug targets was developed (Huang et al., 2013). This algorithm is robust and based on Bayesian probabilistic model, thus achieving certain degree of accuracy. Similarly this situation is with other complex algorithms for drug design. To predict interactions between different molecules that constitute the networks of biochemical interactions scientists developed ClustRNet algorithm based on a Markov chain (Bansal et al., 2009). Considering that biological system contains huge number of components, interactions and variables, it becomes clear how difficult disentangling the various network properties can be. These findings suggest that systems biology solution to epistemological emergence is partial. The problem of limited capacity of human species to develop research hardware or software, and to acquire complete and meaningful explanation of how molecules give rise to organism, probably will be unresolved. The question is how epistemological emergence can support antireductionism?
Epistemological emergence goes hand to hand with weak antireductionism proposed by Kitcher and Vance. Accepting epistemological emergence antireductionist can argue in favor of autonomy of levels only relying on the current state of knowledge. The reason is that systems biology is in its early phase of development, and generally science method has its limitation especially in understanding the life phenomena. Reductionist could argue that epistemic and methodological shortcomings, which are temporary, would be overcome by future developments of systems biology. Also one could theorize that the problem of limited human cognitive powers can be solved by cognitive enhancement or even evolution of human brain. In such a perspective, antireductionist would not be able to count on epistemological emergence. Then, the only solution for them will be to reconsider ontological emergence as potential source of argument.
5. Conclusion
The present paper examines the necessary historical steps that shaped contemporary systems biology. First I have agreed with Westerhoff and Palsson (2004) account about the first historical root of systems biology. My objection to their account is that they did not use the term systems molecular biology, independently from systems biology. I proposed the use of term systems molecular biology as justified, especially when we talk about the interaction of molecular pathways or molecular networks, without the use of mathematical modeling. Systems molecular biology should be distinguished from O’Malley and Dupré understanding of pragmatic systems biology, where mathematical models have an important role. Systems molecular biology dates back to the era of beginning of molecular biology with subsequent discoveries that led to the completion of human genome project. Unlike Westerhoff and Palsson (2004), I have argued that the second phase of systems biology development included two roots. The first root dates back to the development of general systems theory and the second were related to the emergence of nonlinear dynamics. The convergence of general systems theory and nonlinear dynamics has enabled the appearance of systems mathematical biology. I used the term systems mathematical biology instead of systems-theoretical biology, because the word theoretical besides mathematical could have philosophical implications. Since systems biology is closely related to methodology, practice was the only correct way to prove its advantage over molecular reductionist approach. Hence, I proposed that successful application of systems biology in science, medicine and biotechnology was the final phase that shaped contemporary systems biology.
In the postgenomic era molecular approach still exists as an integral part of systems biology approach. As being a part of systems molecular biology, molecular approach provides necessary data that can be used for detailed systems analysis of organism. I have already discussed the unification of molecular and systems paradigm by presenting the postgenomic concept of gene. In addition, it can be concluded that systems biology integrates all existing biological, mathematical, computational and medical knowledge. Systems biology does not represent the revolution in the sense of Kuhnian paradigm shift. It is more of an integrative endeavor, where much of the biological and other sciences are accepted. Hence, systems biology may serve as a good example for other sciences, such as physics, where the problem of scientific revolution is omnipresent. Instead of changing the existing, the best way may be integration of old and new paradigm, thus ensuring the progress of science.
Discussion of Ayala’s (1974) concept of methodological reductionism and critic of Vance (1996) heroic antireductionism was necessary before I introduced the term heroic methodological antireductionism. This type of antireductionism anticipates practice of systems biology. Perhaps, it is the most convenient antireductionist position supported by systems biology. However, at least the weak epistemological antireductionism could be supported by systems biology. But, systems biology is a more research strategy than the subfield of biology, so it is logical to assume its support to methodological antireductionism. The best way to argue in favor of antireductionism is by accepting the ontological emergence. If one could argue in favor of ontological emergence, then strong antireductionism will be guaranteed. In order to defend variants of ontological emergence such as emergent property dualism, systems biology should transcend its current limitations. But in that case, systems biology should be capable to recognize when new emergent properties arise from molecular networks interaction. The transcendence of scientific domain is generally an problem of ontological emergence. Ontological emergence implies unlikely ontology which is why it is very difficult to defend it (Heard, 2006). Although, strengthened by the success of systems biology, epistemological emergence could not yet provide irrefutable antireductionist argumentation. The reason for that could be found in actual state of knowledge and lack of human cognitive powers. The systems biology is completed as a conceptual approach, but advancement in methods and successful application is far from over. The further development may allow humans to find out how all biological hierarchies function. But to do that, besides scientific and technological progress, in order to comprehend all facts, the higher capacity of human mind will be necessary. Systems biology is the field of inexhaustible questions. This paper asks some big historical and philosophical questions and gives modest solutions. Discussion on the subject is only open and scientist and philosophers worldwide should be involved in open-minded dialog.
Conflict of interest
None declared.
Acknowledgments
The author would like to thank the anonymous reviewers for their valuable comments. Suggestions they made contributed significantly to the final version of manuscript.
Footnotes
Peer review under responsibility of King Saud University.
References
- Abarbanel H., Huerta R., Rabinovich M., Rulkov N., Rowat P., Selverston A. Synchronized action of synaptically coupled chaotic model neurons. Neural Comput. 1996;8:1567–1602. doi: 10.1162/neco.1996.8.8.1567. [DOI] [PubMed] [Google Scholar]
- Ahn A.C., Tewari M., Poon C.S., Phillips R.S. The limits of reductionism in medicine: could systems biology offer an alternative? PLoS Med. 2006;3:e208. doi: 10.1371/journal.pmed.0030208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arle J.E., Simon R.H. An application of fractal dimension to the detection of transients in the electroencephalogram. Electroencephalogr. Clin. Neurophysiol. 1990;75:296–305. doi: 10.1016/0013-4694(90)90108-v. [DOI] [PubMed] [Google Scholar]
- Auffray C., Imbeaud S., Roux-Rouquie M., Hood L. From functional genomics to systems biology: concepts and practices. Comptes Rendus Biol. 2003;326:879–892. doi: 10.1016/j.crvi.2003.09.033. [DOI] [PubMed] [Google Scholar]
- Ayala F.L. Introduction. In: Ayala F.L., Dobzhansky T.G., editors. Studies in the Philosophy of Biology: Reduction and Related Problems. University of California Press; Los Angeles: 1974. [Google Scholar]
- Bansal S., Khandelwal S., Meyers L.A. Exploring biological network structure with clustered random networks. BMC Bioinformatics. 2009;10:405. doi: 10.1186/1471-2105-10-405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bechtel W., Hamilton A. Reduction, integration, and the unity of science. natural, behavioral, and social science and the humanities. In: Kuipers T., editor. North Holland – Elsevier; Oxford, UK: 2007. (General Philosophy of Science. Focal Issues). [Google Scholar]
- Bitbol M. Ontology, matter and emergence. Phenomenol. Cogn. Sci. 2007;6:293–307. [Google Scholar]
- Bruggeman F.J., Westerhoff H.V. The nature of systems biology. Trends Microbiol. 2006;15:45–50. doi: 10.1016/j.tim.2006.11.003. [DOI] [PubMed] [Google Scholar]
- Crane T. Dualism, Monism, Physicalism. Mind Soc. 2000;1:73–85. [Google Scholar]
- Drack M. On the making of a system theory of life: Paul A Weiss and Ludwig Von Bertalanffy’s conceptual connection. Q. Rev. Biol. 2007;82:349–373. doi: 10.1086/522810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drack M. Ludwig von Bertalanffy’s early system approach. Syst. Res. Behav. Sci. 2009;26:563–572. [Google Scholar]
- Ederer M., Sauter T., Bullinger E., Gilles E.D., Allgower F. An approach for dividing models of biological reaction networks into functional units. Simulation. 2003;79:703–716. [Google Scholar]
- Eke A., Herman P., Kocsis L., Kozak L. Fractal characterization of complexity in temporal physiological signals. Physiol. Meas. 2002;23:1–38. doi: 10.1088/0967-3334/23/1/201. [DOI] [PubMed] [Google Scholar]
- Emmeche C., Koppe S., Stjernfelt F. Explaining emergence: towards ontology of levels. J. Gen. Phil. Sci. 1997;28:83–119. [Google Scholar]
- Esteller R., Vachtsevanos G., Echauz J., Litt B. A comparison of waveform fractal dimension algorithms. IEEE Trans. Circuits Syst. 2001;48:177–183. [Google Scholar]
- Faye M. Does the unity of science have a future? In: Galavotti M.C., editor. European Philosophy of Science – Philosophy of Science in Europe and the Viennese Heritage. Springer International Publishing; Switzerland: 2014. [Google Scholar]
- Friboulet A., Thomas D. Systems biology – an interdisciplinary approach. Biosens. Bioelectron. 2005;20:2404–2407. doi: 10.1016/j.bios.2004.11.014. [DOI] [PubMed] [Google Scholar]
- Griffiths P.E., Stotz K. Genes in the postgenomic era. Theor. Med. Bioethics. 2006;27:499–521. doi: 10.1007/s11017-006-9020-y. [DOI] [PubMed] [Google Scholar]
- Gulick R.V. Reduction, emergence and other recent options on the mind/body problem. J. Conscious. Stud. 2001;8:1–34. [Google Scholar]
- Hasty J., McMillen D., Isaacs F., Collins J.J. Computational studies of gene regulatory networks: in numero molecular biology. Nat. Rev. Genet. 2001;2:268–279. doi: 10.1038/35066056. [DOI] [PubMed] [Google Scholar]
- Heard D. A new problem for ontological emergence. Philos. Quart. 2006;56:55–62. [Google Scholar]
- Higgins J.P. Nonlinear systems in medicine. Yale J. Biol. Med. 2002;75:247–260. [PMC free article] [PubMed] [Google Scholar]
- Higuchi T. Approach to an irregular time series on the basis of the fractal theory. Physica D. 1988;31:277–283. [Google Scholar]
- Hornberg J.J., Bruggeman F.J., Westerhof H.V., Lankelma J. Cancer: a systems biology disease. BioSystems. 2006;83:81–90. doi: 10.1016/j.biosystems.2005.05.014. [DOI] [PubMed] [Google Scholar]
- Huang J., Niu C., Green C.D., Yang L., Mei H., Han J.D.J. Systematic prediction of pharmacodynamic drug–drug interaction through protein–protein–interaction network. PLoS Comput. Biol. 2013;9:e1002998. doi: 10.1371/journal.pcbi.1002998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kitcher P. 1953 and all that. A tale of two sciences. Philos. Rev. 1984;93:335–373. [PubMed] [Google Scholar]
- Klonowski W. From conformons to human brains: an informal overview of nonlinear dynamics and its applications in biomedicine. Nonlinear Biomed. Phys. 2007;1:5. doi: 10.1186/1753-4631-1-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leith C.E. Stochastic models of chaotic systems. Physica D. 1996;98:481–491. [Google Scholar]
- Liebovitch L., Todorov A. Using fractals and nonlinear dynamics to determine the physical properties of ion channel protein. Crit. Rev. Neurobiol. 1996;10:169–187. [PubMed] [Google Scholar]
- Mazzochi F. Complexity in biology: exceeding the limits of reductionism and determinism using complexity theory. EMBO J. 2008;9:10–14. doi: 10.1038/sj.embor.7401147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mazur J., Ritter D., Renelt G., Kaderali L. Reconstructing nonlinear dynamic models of gene regulation using stochastic sampling. BMC Bioinformatics. 2009;10:448. doi: 10.1186/1471-2105-10-448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meister A., Li Y.H., Bokyung C., Wong W.H. Learning a nonlinear dynamical system model of gene regulation: a perturbed steady-state approach. Ann. Appl. Stat. 2013;7:1311–1333. [Google Scholar]
- Mesarovic M.D. General systems theory and biology – view of a theoretician. In: Mesarović D.M., editor. General Systems Theory and Biology. Springer; 1968. [Google Scholar]
- Mesarovic M.D., Sreenath S.N., Keene J.D. Search for organising principles: understanding in systems biology. Syst. Biol. 2004;1:19–27. doi: 10.1049/sb:20045010. [DOI] [PubMed] [Google Scholar]
- Naik G., Arjunan S., Kumar D. Application of ICA and fractal dimension in sEMG signal processing for subtle movement analysis: a review. Aust. Phys. Eng. Sci. Med. 2011;34:179–193. doi: 10.1007/s13246-011-0066-4. [DOI] [PubMed] [Google Scholar]
- Noble D. Differential and integral views of genetics in computational systems biology. Interface Focus. 2010;1:7–15. doi: 10.1098/rsfs.2010.0444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Noorbakhsh F., Overall C.M., Power C. Deciphering complex mechanisms in neurodegenerative diseases: the advent of systems biology. Trends Neurosci. 2009;32:88–100. doi: 10.1016/j.tins.2008.10.003. [DOI] [PubMed] [Google Scholar]
- O’Malley M.A., Dupré J. Fundamental issues in systems biology. BioEssays. 2005;27:1270–1276. doi: 10.1002/bies.20323. [DOI] [PubMed] [Google Scholar]
- Oestreicher C. A history of chaos theory. Dialog. Clin. Neurosci. 2007;9:279–289. doi: 10.31887/DCNS.2007.9.3/coestreicher. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paraschiv-Ionescu A., Aminian K. Nonlinear analysis of physiological time series. In: Nait-Ali A., editor. Advanced Biosignal Processing. Springer-Verlag; Berlin: 2009. [Google Scholar]
- Rosenberg A. Cambridge University Press; Cambridge, New York: 1985. The Structure of Biological Sciences. [Google Scholar]
- Schütt A., Ross A., Figliola A. A discovery of new features of gastropod local field potentials by application of wavelet tools. J. Neurosci. Methods. 2002;119:89–104. doi: 10.1016/s0165-0270(02)00184-x. [DOI] [PubMed] [Google Scholar]
- Scott A. Springer-Verlag; New York: 2002. Neuroscience: A Mathematical Primer. [Google Scholar]
- Searle J.R. MIT Press; Cambridge, MA: 1992. The Rediscovery of the Mind. [Google Scholar]
- Silberstein M., McGeever J. The Search for Ontological Emergence. Philos. Quart. 1999;49:201–214. [Google Scholar]
- Stam C.J. Nonlinear dynamical analysis of EEG and MEG: review of an emerging field. Clin. Neurophysiol. 2005;116:2266–2301. doi: 10.1016/j.clinph.2005.06.011. [DOI] [PubMed] [Google Scholar]
- Stotz K. How biologist conceptualize genes: an empirical study. Stud. Hist. Philos. Biol. Biomed. Sci. 2004;35:647–673. [Google Scholar]
- Strange K. The end of naive reductionism: rise of systems biology or renaissance of physiology? Am. J. Physiol. Cell Physiol. 2005;288:C968–C974. doi: 10.1152/ajpcell.00598.2004. [DOI] [PubMed] [Google Scholar]
- Vance R.E. Heroic antireductionism and genetics: a tale of one science. Philos. Sci. 1996;63:S36–S45. [Google Scholar]
- Westerhoff H.V., Palsson B.O. The evolution of molecular biology into systems biology. Nat. Biotechnol. 2004;22:1249–1252. doi: 10.1038/nbt1020. [DOI] [PubMed] [Google Scholar]