

Abstract
Neutrophil granulocytes are the most abundant leukocytes in the human blood. Neutrophils are the first to arrive at the site of infection. Neutrophils developed several antimicrobial mechanisms including phagocytosis, degranulation and formation of neutrophil extracellular traps (NETs). NETs consist of a DNA scaffold decorated with histones and several granule markers including myeloperoxidase (MPO) and human neutrophil elastase (HNE). NET release is an active process involving characteristic morphological changes of neutrophils leading to expulsion of their DNA into the extracellular space. NETs are essential to fight microbes, but uncontrolled release of NETs has been associated with several disorders. To learn more about the clinical relevance and the mechanism of NET formation, there is a need to have reliable tools capable of NET quantitation.
Here three methods are presented that can assess NET release from human neutrophils in vitro. The first one is a high throughput assay to measure extracellular DNA release from human neutrophils using a membrane impermeable DNA-binding dye. In addition, two other methods are described capable of quantitating NET formation by measuring levels of NET-specific MPO-DNA and HNE-DNA complexes. These microplate-based methods in combination provide great tools to efficiently study the mechanism and regulation of NET formation of human neutrophils.
Keywords: Immunology, Issue 112, neutrophil extracellular traps, NET, granulocytes, neutrophil, extracellular DNA, myeloperoxidase, neutrophil elastase, quantitation, ELISA, high throughput
Introduction
NET formation is a novel mechanism by which neutrophils fight pathogens.1 The core of NETs is nuclear DNA.1 This DNA network is associated with neutrophil granule proteins and histones.1 The main form of NET formation requires the death of neutrophils characterized by chromatin decondensation, disappearance of granular and nuclear membranes, translocation of neutrophils elastase to the nucleus, citrullination of histones and finally the spill of DNA-based NETs.2 NETs entrap and kill a wide variety of microbes and are an essential part of the innate immune weapon repertoire. Uncontrolled NET formation has, however, been linked to numerous autoinflammatory diseases.3,4 Despite their increasingly established relevance, little is known about the mechanism and regulation of NET release.
Neutrophils dying by releasing NETs are different from apoptotic or necrotic neutrophils.3,5 NET-releasing neutrophils show several features that are characteristic for NET formation. Granule components are associated with DNA in NETs.1 Myeloperoxidase (MPO) and human neutrophil elastase (HNE) are both found in primary granules in resting cells but are translocated to the nucleus to bind to DNA in NETs.1 MPO-DNA and HNE-DNA complexes are specific for NETs, do not occur in apoptotic or necrotic neutrophils.1,3,5 Chromatin decondensation is another feature typical for NETosis.2 NET release also requires citrullination of histones by peptidyl aminidase 4 (PAD4).6 Citrullinated histones are hallmarks of neutrophils that underwent NET release.6
Here three methods are introduced that in combination provide excellent tools to quantitate NETs on a high-throughput scale. The first assay has been used on the field with different changes and quantitates extracellular DNA release in a microplate format. The second and third assays provide confirmation of NETs by measuring NET-specific MPO-DNA and HNE-DNA complexes.
Protocol
The Institutional Review Board of the University of Georgia approved the human subject study to collect peripheral blood from healthy volunteers (UGA# 2012-10769-06).5,7,8 Volunteers signed the required informed consent form before blood draw. The research performed in this article is in compliance with the ethical guidelines for medical research involving human subjects of the Declaration of Helsinki.
1. Isolation of Neutrophils from Peripheral Human Blood (Figure 1)
Note: There are several ways to isolate neutrophils from peripheral blood. The following protocol provides one possibility. This protocol yields large numbers of non-activated human neutrophils capable of releasing NETs upon external stimulation. Use 40 ml whole blood obtained from healthy volunteers by venipuncture.7,9
Aliquot 20 ml blood into two 50 ml conical tubes. Add 10 ml 6% Dextran. Mix gently.
After 20 min transfer the leukocyte-rich upper phase without taking any pelleted red blood cells into clean 50 ml conical tubes, and fill it up with sterile PBS.
Centrifuge at 400 x g 10 min. Resuspend pellet in 4 ml sterile PBS.
Prepare a 5-step Percoll gradient of 85%-80%-75%-70%-65% in two 15 ml conical tubes and layer 2 ml leukocyte suspension on top of each gradient.
Centrifuge 800 x g for 30 min with brakes off.
Collect all the cells (neutrophils) in the layers at the 75%/80% and 70%/75% interface.
Wash cells twice in PBS (350 x g, 10 min, RT) and count cells using a hemocytometer.
2. Measurement of Kinetics of Extracellular DNA Release Using a Microplate Fluorimeter
Note: This method enables measurement of changes in fluorescence of a membrane-impermeable DNA-binding dye indicative of DNA release on a 96-well microplate format (Figure 2).
Prepare a suspension of neutrophils in assay medium (HBSS + 1% (v/v) autologous serum + 5 mM glucose) at a concentration of 2 x 106 cells/ml.
Add 5 µmol/L of the membrane impermeable DNA-binding dye. Mix gently.
Aliquot human neutrophils (50 µl/well) onto 96-well black, transparent bottom microplates. Make sure the assay medium covers the entire bottom of the well. If not, tap the plate gently on the side.
Warm up the plate containing neutrophils for 10 min at 37 °C.
In the meantime prepare solutions of stimuli at double of their final concentrations to be used (in 37 °C warm assay medium). As a positive NET control use human neutrophils with 100 nM phorbol-myristate-acetate (PMA) for 4 hr to trigger maximal NET release.5,8 PMA stimulation for 4 hr ensures maximal NET release while spontaneous NET formation remains low5,8.
Prepare the microplate fluorimeter for the measurement (4 hr, 37 °C, 530 nm for excitation and 590 nm for emission).
Stimulate neutrophils by adding 50 µl stimulus solution to 50 µl neutrophil suspensions. Use 0.5 µg/ml saponin in one well to measure maximal DNA release signal.
Place plate in reader and measure changes in fluorescence for 4 hr at every 2 min with no shaking.
- For data analysis, calculate normalized increases in fluorescence over time.
- Subtract baseline fluorescence from endpoint values for all wells including saponin (Figure 2A, B). Rise in saponin fluorescence should be the highest signal. It is referred to as "maximal DNA release" (Figure 2A).
- Calculate percentages of DNA release in unknown samples by dividing increases in fluorescence of unknown samples by maximal DNA release.
- Average results of replicates and present them as "% of maximal DNA release" (Figure 2C).
3. Quantitation of NET Formation by MPO-DNA and HNE-DNA ELISA Assays
Note: These assays modified (MPO-DNA) or established (HNE-DNA) in our laboratory quantitate NET formation by measuring levels of MPO-DNA and HNE-DNA complexes.5
Coat 96-well high-binding capacity ELISA plates overnight with capture antibodies (50 μl/well): anti-MPO (1:2,000 dilution) or anti-HNE (1:2,000).
Wash ELISA plates three times with PBS (200 μl/well) and block them with 5% BSA and 0.1% human serum albumin for 2 hr at RT (200 μl/well). Wash three times with PBS.
Seed neutrophils on 96-well microplates at a density of 100,000 cells/well in 100 µl assay medium, and stimulate NET formation. Incubate cells: 4 hr for 37 °C.
Perform a limited DNase digestion by adding 2 U/ml DNase to neutrophil supernatants, mix them well. Keep them at RT.
Stop the DNase after 15 min by adding 11 µl 25 mM EGTA (final concentration 2.5 mM). Mix well. Collect supernatants in clean microfuge tubes. Spin samples to get rid of cell debris at 300 x g for 5 min at RT. Transfer their supernatants into clean microfuge tubes. Keep them on ice until ready.
- Use an aliquot of a previously prepared "NET standard". Note: The NET-standard is a mix of DNase-digested supernatants of PMA-stimulated neutrophils obtained from at least 5 different, independent human donors.
- To use the same standard for a long time, collect and aliquot a large volume of neutrophil supernatants from each individual donor. Collecting for example 2 ml of DNase-digested supernatants of PMA-stimulated neutrophils will result in 200 aliquots (10 µl each) enough for 200 ELISA plates. Data characterizing the NET standard are found in Figure 5.
- Stimulate human neutrophils with 100 nM PMA for 4 hr and apply DNase digest to their supernatants according steps 3.4-3.5.
- Collect, pool, aliquot (10 µl) and freeze (-20 °C) neutrophil supernatants.
- To prepare the "NET-standard", thaw one aliquot/donor obtained from at least five separate donors, mix them and keep them on ice.
- Discard thawed samples and use fresh ones for the next experiment.
- Prepare a 1:2 serial dilution of the NET-standard in PBS+EGTA.
Dilute DNase1-digested samples (standard and unknown supernatants) 20-fold in PBS+EGTA and aliquot them on ELISA plates coated with capture antibodies.
Incubate ELISA plates over night at 4 °C and wash them three times with PBS.
Apply detection antibody solution (100 µl/well): horse radish peroxidase conjugated anti-DNA antibody (1:500, mouse). Incubate for 1 hr in dark.
After four washes with PBS add TMB peroxidase substrate: 100 µl/well, 30 min. Blue coloration is an indication for peroxidase activity (NETs present).
Stop reaction by adding 100 μl/well 1 M HCl. The blue solutions will turn yellow (Figure 5A).
Read absorbance at 450 nm using a microplate photometer.
- For data analysis, calculate the amount of MPO-DNA or HNE-DNA complexes compared to the NET standard.
- Subtract the background optical density value of the assay medium from all samples.
- Plot the absorbance values (X-axis) of the diluted standard samples against their relative NET content (Y-axis) (Figure 5A).
- Using the nonsaturated range of this standard curve establish a trend line (use best exponential fit from the software used) (Figure 5A). The equation of this trend line determines the conversion of OD values to quantitative amounts of NETs.
- Insert the measured OD values of the unknowns into the trend-line equation from 3.13.3 that will give the amount of NETs in the unknown sample as percentage of the NET-standard (Figure 5A).
- Calculate averages of replicates (triplicates) and present the final data as "amount of MPO-DNA or HNE-DNA complexes (% of standard)" (Figure 5C).
Representative Results
The figures in this manuscript describe the method of neutrophil isolation, experimental procedures, and present representative results with explanation of data analysis. Figure 1 shows the sequential steps of human neutrophil preparation. This protocol represents only one possible way of neutrophil isolation. It yields large amounts of resting neutrophils capable of releasing NETs upon stimulation. Figure 2 shows how the fluorescence-based DNA release assay works. Using a fluorescence microplate reader kinetics of DNA release in human neutrophils are followed in 96-well microplates. Figure 3 explains each step of the data analysis. First, calculate maximal DNA signal in neutrophils treated with saponin (Figure 3A). Increase in fluorescence is always calculated as the difference between the highest fluorescence value and the background fluorescence level (Figure 3A). This is calculated for each well. The example shows representative data of human neutrophils left non-stimulated or activated with PMA or P. aeruginosa (PAO1) (Figure 3B). P. aeruginosa triggers NET formation in human PMNs.5,8,10 It has been shown with several P. aeruginosa strains including PAO1 that bacteria alone (without neutrophils) do not bind the used DNA-detecting dye.8 PAO1 P. aeruginosa was obtained from ATCC, cultured in Luria-Bertani medium overnight, harvested in late-exponential phase, washed in assay medium and used to stimulate neutrophils at a multiplicity of infection (MOI) of 10:1 (Figure 3B, C, 4B, C). Averages of increase in fluorescence of replicate wells are calculated, normalized on maximal fluorescence (saponin) and are expressed as "DNA release (% of max)" (Figure 3C). Figure 3D shows representative fluorescence images of untreated or PMA-stimulated neutrophils.
Figure 4A depicts the scheme explaining the principles of the MPO-DNA and HNE-DNA ELISA assays. Figure 4B shows representative immunofluorescence images of nonstimulated, PMA- and PAO1-activated human neutrophils. Extracellular DNA shows typical NET morphology and co-localizes with MPO and citrullinated histone H4, a marker of NET formation.6 Figure 4C shows MPO-DNA and HNE-DNA release in human neutrophils under the same, previous conditions (like Figure 3B, 3C and 4B). IThe "NET" ELISA methods have been previously characterized.5 Briefly, 2 U/ml DNase treatment (equivalent of 1 µg/ml concentration of the DNase used) for 15 min at RT resulted in the highest MPO-DNA and HNE-DNA signals.5 Running samples (100 ng/lane) on an agarose gel revealed that DNA length in the lower kb range yields optimal ELISA signal.5 DNA overdigestion (DNase dose higher than 20 U/ml) or omitting either the capture or the detection antibody abolished the ELISA signals.5
In addition, here the assays and the "NET- standard" are further characterized (Figure 5A). The NET-standard contains on average 19.5 µg/ml extracellular DNA (measured by using a standard with known DNA concentrations), 399 ng/ml MPO and 103.4 ng/ml HNE (commercial ELISAkits) (n = 8). Based on the total molecular weights of MPO (84 kDa) and HNE (29.5 kDa) and the fact that 1 µg DNA contains 9.91 x 1014 nucleotides, the NET-standard contains 109 HNE molecules and 147 MPO molecules per 1,000 nucleotides (kb DNA). Two-fold serial dilution experiments of the NET-standard revealed that the dynamic ranges of both the MPO-DNA and HNE-DNA ELISA assays are between 19.0 - 609.0 ng/ml DNA concentrations (Figure 5B). Dilutions of lower than 32-fold of the NET standard resulted in saturation whereas dilutions of higher than 1,024-fold were not different from the background (Figure 5B). Treatment of the NET standard with protease inhibitors did not change its results obtained by the MPO-DNA and HNE-DNA assays indicating that proteases do not interfere with these assays (Figure 5C). Thus, results obtained in the same NET-standard sample by the MPO-DNA or HNE-DNA ELISA assays strongly correlate with each other (Figure 5D). To show that frozen PMN supernatants can also be used in these assays without losing efficiency, some aliquots of the NET-standard were left untreated or others were exposed to one or two freeze/thaw cycles performed at -20 °C or -80 °C (1 hr interval). None of the freeze/thaw treatments affected the MPO-DNA or HNE-DNA results (Figure 5E). In summary, the NET-standard provides a reliable reference point in these NET-quantifying assays enabling the quantitative comparison of NET results between independent experiments.
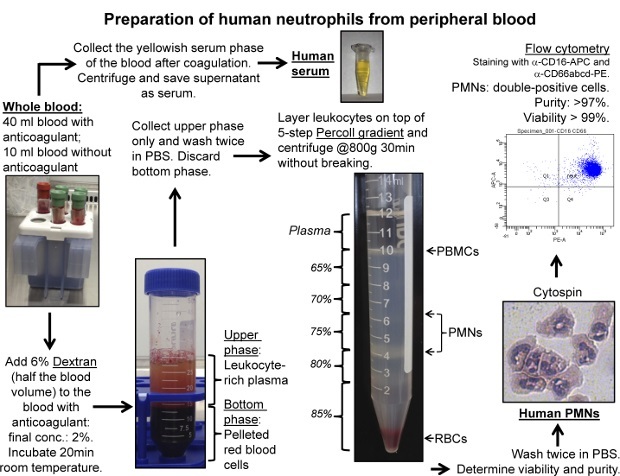 Figure 1:Isolation of Human Neutrophils. Neutrophil granulocytes were isolated from peripheral blood of volunteers according to the described protocol. Blood containing anticoagulant was used to isolate neutrophils by Dextran sedimentation and gradient centrifugation. Blood without anticoagulant was used to prepare serum. Viability of the neutrophil preparations is assessed by Trypan Blue exclusion assay. Neutrophil purity is assessed by cytospin or flow cytometry (CD16, CD66abcd double-positive cells). Please click here to view a larger version of this figure.
Figure 1:Isolation of Human Neutrophils. Neutrophil granulocytes were isolated from peripheral blood of volunteers according to the described protocol. Blood containing anticoagulant was used to isolate neutrophils by Dextran sedimentation and gradient centrifugation. Blood without anticoagulant was used to prepare serum. Viability of the neutrophil preparations is assessed by Trypan Blue exclusion assay. Neutrophil purity is assessed by cytospin or flow cytometry (CD16, CD66abcd double-positive cells). Please click here to view a larger version of this figure.
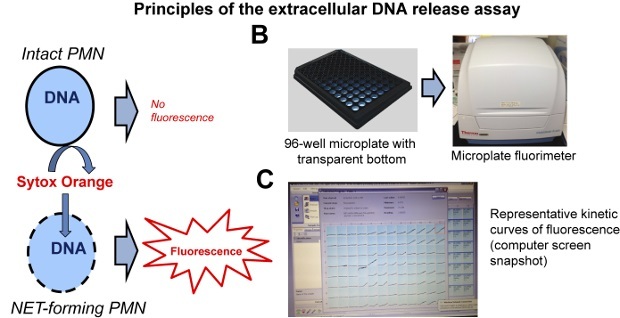 Figure 2:Principles of the Extracellular DNA Release Assay. (A) Fluorescence is only obtained in neutrophils whose plasma and nuclear membranes have been compromised and the membrane-impermeable DNA-binding dye can bind to their DNA. (B) The assay is performed on black 96-well microplates using a microplate fluorimeter. (C) The fluorimeter records real-time fluorescence curves in each well. One representative results in shown. Please click here to view a larger version of this figure.
Figure 2:Principles of the Extracellular DNA Release Assay. (A) Fluorescence is only obtained in neutrophils whose plasma and nuclear membranes have been compromised and the membrane-impermeable DNA-binding dye can bind to their DNA. (B) The assay is performed on black 96-well microplates using a microplate fluorimeter. (C) The fluorimeter records real-time fluorescence curves in each well. One representative results in shown. Please click here to view a larger version of this figure.
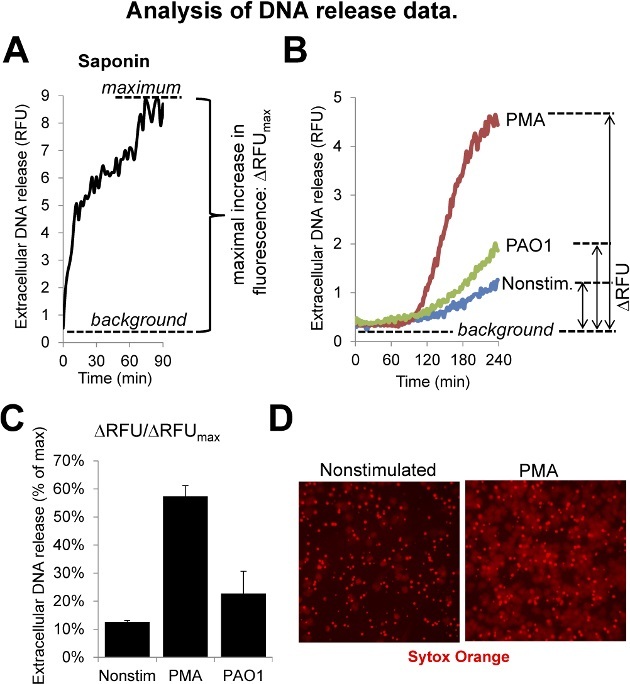 Figure 3: Analysis of the DNA Release Data. (A) Saponin is used to create the maximal value since it entirely lyses neutrophils and exposes their DNA. The kinetics of saponin-induced fluorescence is shown. (B) Kinetics of neutrophil DNA release induced by PMA (100 nM) and bacteria (Pseudomonas aeruginosa PAO1, 10 MOI). (C) Normalized summarized DNA release data obtained in one experiments using quadriplicates. (D) Representative fluorescence images of untreated and PMA-stimulated human neutrophils (Sytox Orange, 4 hr incubation). Please click here to view a larger version of this figure.
Figure 3: Analysis of the DNA Release Data. (A) Saponin is used to create the maximal value since it entirely lyses neutrophils and exposes their DNA. The kinetics of saponin-induced fluorescence is shown. (B) Kinetics of neutrophil DNA release induced by PMA (100 nM) and bacteria (Pseudomonas aeruginosa PAO1, 10 MOI). (C) Normalized summarized DNA release data obtained in one experiments using quadriplicates. (D) Representative fluorescence images of untreated and PMA-stimulated human neutrophils (Sytox Orange, 4 hr incubation). Please click here to view a larger version of this figure.
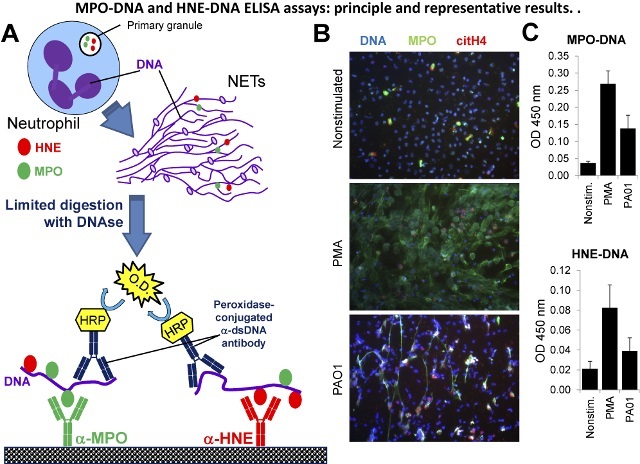 Figure 4:MPO-DNA and HNE-DNA ELISA Assays: Principle and Representative Results. (A) Scheme explaining how the MPO-DNA and HNE-DNA ELISA assays work. (B) Representative results of immunofluorescence images prepared on human neutrophils exposed to 100 nM PMA or P. aeruginosa PAO1 strain (10 MOI) for 4 hr. Extracellular DNA (DAPI, blue) co-localizes in NETs with MPO (green) and citrullinated histone H4 (red). One representative result, n = 3. (C) NET release was measured in human neutrophils exposed to the same stimuli as above by the MPO-DNA and HNE-DNA ELISA assays. Mean ± S.E.M., n = 3. Please click here to view a larger version of this figure.
Figure 4:MPO-DNA and HNE-DNA ELISA Assays: Principle and Representative Results. (A) Scheme explaining how the MPO-DNA and HNE-DNA ELISA assays work. (B) Representative results of immunofluorescence images prepared on human neutrophils exposed to 100 nM PMA or P. aeruginosa PAO1 strain (10 MOI) for 4 hr. Extracellular DNA (DAPI, blue) co-localizes in NETs with MPO (green) and citrullinated histone H4 (red). One representative result, n = 3. (C) NET release was measured in human neutrophils exposed to the same stimuli as above by the MPO-DNA and HNE-DNA ELISA assays. Mean ± S.E.M., n = 3. Please click here to view a larger version of this figure.
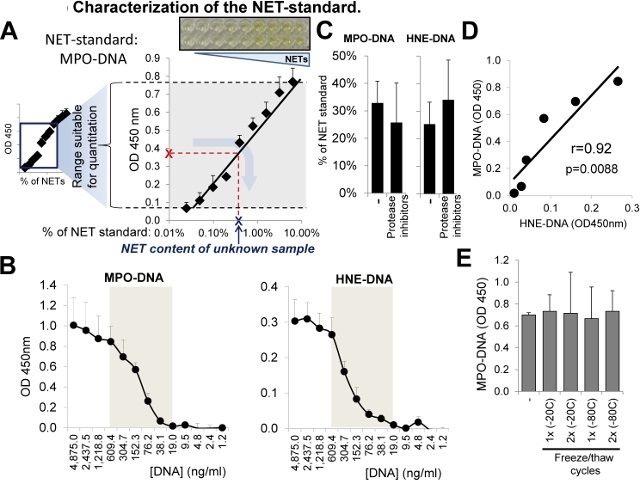 Figure 5:Characterization of the "NET-standard". (A) Serial dilutions (1:2) of the NET standard were prepared and subjected to MPO-DNA ELISA. Measured OD values are plotted against % of NET-standard content. Red "X" indicates a measured OD value of an unknown sample. Gray arrow indicates how the "NET concentration" (blue "X") of the unknown sample is to be determined using the central range of the fitted standard curve. (B) DNA concentrations in serially diluted NET-standard samples were determined and plotted against the measured OD values of the MPO-DNA or HNE-DNA ELISA assays. Gray areas indicate the dynamic ranges of the assays. (C) Protease inhibitor treatment (protease inhibitor cocktail, 1%) does not affect the outcome of the MPO-DNA or HNE-DNA assays (NET-standard). Mean ± S.E.M., n = 2. (D) Correlation of MPO-DNA and HNE-DNA results (OD 450 nm) of serially diluted "NET-standard" samples in their dynamic ranges (See 5B). (E) Freezing and thawing cycles (-20 °C or -80 °C; one or two) do not affect the results of MPO-DNA and HNE-DNA assays (NET-standard, mean ± S.E.M., n = 2). Please click here to view a larger version of this figure.
Figure 5:Characterization of the "NET-standard". (A) Serial dilutions (1:2) of the NET standard were prepared and subjected to MPO-DNA ELISA. Measured OD values are plotted against % of NET-standard content. Red "X" indicates a measured OD value of an unknown sample. Gray arrow indicates how the "NET concentration" (blue "X") of the unknown sample is to be determined using the central range of the fitted standard curve. (B) DNA concentrations in serially diluted NET-standard samples were determined and plotted against the measured OD values of the MPO-DNA or HNE-DNA ELISA assays. Gray areas indicate the dynamic ranges of the assays. (C) Protease inhibitor treatment (protease inhibitor cocktail, 1%) does not affect the outcome of the MPO-DNA or HNE-DNA assays (NET-standard). Mean ± S.E.M., n = 2. (D) Correlation of MPO-DNA and HNE-DNA results (OD 450 nm) of serially diluted "NET-standard" samples in their dynamic ranges (See 5B). (E) Freezing and thawing cycles (-20 °C or -80 °C; one or two) do not affect the results of MPO-DNA and HNE-DNA assays (NET-standard, mean ± S.E.M., n = 2). Please click here to view a larger version of this figure.
Discussion
NETs represent a fascinating novel mechanism by which neutrophils kill pathogens.1 Although the literature of NETs has been continuously expanding over the last ten years since their discovery, several important questions related to their role in biology, mechanism and regulation remain unclear. Appropriate methodology has to be developed to measure NETs, this very unique antimicrobial mechanism. This article describes methods that can be used to quantitate NETs in a high throughput manner. The first assay follows kinetics of fluorescence of a membrane-impermeable DNA-binding dye. This assay provides large amount of information on the slope and velocity of DNA release from neutrophils. This information is crucial in investigations studying early signaling events leading to NET formation. All this information is lost in endpoint assays measuring DNA release at the end of the incubation. The extracellular DNA dye-based assay is not specific for NETs. It only measures DNA release. To confirm NETs, it is widely accepted to perform immunofluorescence on NET-forming neutrophils and show co-localization of DNA with either granule markers (MPO, HNE) or histones. Quantitation of immunofluorescence data is though very tedious and subjective.
Here, a modification of a previously developed ELISA assay detecting MPO-DNA complexes is presented. A similar ELISA assay measuring HNE-DNA complexes that represent specific NET measures is also described.3,5 Both assays have been characterized in detailed.5 The MPO-DNA and HNE-DNA ELISA assays are quantitative and NET-specific at the same time.5 These features are not fulfilled by any other current method. The assays are easy to perform and provide reproducible data.5 A limited DNase digestion step is included that ensures prompt and standardized handling of NETs directly on neutrophils.5 These assays are suitable to compare several samples at the same time. In new, control measurements it is shown that freeze/thaw cycles and proteases do not affect the outcome of the MPO-DNA and HNE-DNA assays. It is unknown, however, whether proteins known to bind to DNA in NETs (including histones and LL-37)1,11 would interfere with these assays.
Critical steps of the presented protocols are the following. It is important to harvest neutrophils that are in a resting, quiescent state and do not release large amount of NETs spontaneously. Lack of a hypotonic lysis step to remove red blood cells in the presented neutrophil isolation protocol is crucial to obtain non-activated neutrophils. To inhibit neutrophil activation after the cells had been purified, it is also important to resuspend neutrophils in a medium containing their donor's own serum. To obtain quiescent cells it is also crucial to use pyrogen- and contamination-free reagents during neutrophil preparation. Spontaneous NET release of neutrophils during the extracellular DNA release assay could also happen. Adding 1% autologous serum to the assay medium is important to prevent it. Higher or lower doses of the autologous serum could be tested to get optimal results. It is critical during the MPO-DNA and HNE-DNA ELISA assays that released DNA should not be over-digested by the DNase because it will result in complete loss of the signal. Therefore, the optimal dose should be determined for every new DNase batch. The parameters of the DNA digestion could be slightly changed to obtain the highest ELISA signals possible. It is also important to mix the DNase and NETs thoroughly to ensure that the DNase will have clear access to the DNA. When harvesting the digested NETs, wash the well extensively to collect all the NETs.
The DNA detection limit of the MPO-DNA and HNE-DNA assays is around 10-20 ng/ml DNA (Figure 5B). Serum and plasma free DNA levels of several hundred to thousand ng/ml were reported in diseases (cancer, systemic lupus erythematosus, myocardial infection) that have been associated with abnormal NET formation.12-14 This suggests that the MPO-DNA and HNE-DNA ELISA assays also have the potential to be able to quantitate NETs in human clinical samples. This future application of the MPO-DNA and HNE-DNA ELISA assays will enable to quantitatively link NETs to disease pathogenesis.
Disclosures
The authors have nothing to disclose.
Acknowledgments
Special thanks to the personnel of the University of Georgia Health Center laboratory for their continuous support of our work on isolating human neutrophils. This work was supported by the start-up fund of Dr. Rada provided by UGA Office of Vice President for Research.
References
- Brinkmann V, et al. Neutrophil extracellular traps kill bacteria. Science. 2004;303:1532–1535. doi: 10.1126/science.1092385. [DOI] [PubMed] [Google Scholar]
- Fuchs TA, et al. Novel cell death program leads to neutrophil extracellular traps. J Cell Biol. 2007;176:231–241. doi: 10.1083/jcb.200606027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kessenbrock K, et al. Netting neutrophils in autoimmune small-vessel vasculitis. Nat Med. 2009;15:623–625. doi: 10.1038/nm.1959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leffler J, et al. Degradation of neutrophil extracellular traps co-varies with disease activity in patients with systemic lupus erythematosus. Arthritis Res Ther. 2013;15:R34. doi: 10.1186/ar4264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoo DG, Floyd M, Winn M, Moskowitz SM, Rada B. NET formation induced by Pseudomonas aeruginosa cystic fibrosis isolates measured as release of myeloperoxidase-DNA and neutrophil elastase-DNA complexes. Immunol Lett. 2014;160:186–194. doi: 10.1016/j.imlet.2014.03.003. [DOI] [PubMed] [Google Scholar]
- Wang Y, et al. Histone hypercitrullination mediates chromatin decondensation and neutrophil extracellular trap formation. J Cell Biol. 2009;184:205–213. doi: 10.1083/jcb.200806072. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pang L, Hayes CP, Buac K, Yoo DG, Rada B. Pseudogout-associated inflammatory calcium pyrophosphate dihydrate microcrystals induce formation of neutrophil extracellular traps. J Immunol. 2013;190:6488–6500. doi: 10.4049/jimmunol.1203215. [DOI] [PubMed] [Google Scholar]
- Yoo DG, et al. Release of cystic fibrosis airway inflammatory markers from Pseudomonas aeruginosa-stimulated human neutrophils involves NADPH oxidase-dependent extracellular DNA trap formation. J. Immunol. 2014;192:4728–4738. doi: 10.4049/jimmunol.1301589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brinkmann V, Laube B, Abu Abed , Goosmann U, Zychlinsky C, A Neutrophil extracellular traps, how to generate and visualize them. J. Vis. Exp. 2010. [DOI] [PMC free article] [PubMed]
- Rada B, et al. Pyocyanin-enhanced neutrophil extracellular trap formation requires the NADPH oxidase. PLoS One. 2013;8:e54205. doi: 10.1371/journal.pone.0054205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neumann A, et al. The antimicrobial peptide LL-37 facilitates the formation of neutrophil extracellular traps. Biochem J. 2014;464:3–11. doi: 10.1042/BJ20140778. [DOI] [PubMed] [Google Scholar]
- Leon SA, Shapiro B, Sklaroff DM, Yaros MJ. Free DNA in the serum of cancer patients and the effect of therapy. Cancer Res. 1977;37:646–650. [PubMed] [Google Scholar]
- Raptis L, Menard HA. Quantitation and characterization of plasma DNA in normals and patients with systemic lupus erythematosus. J Clin Invest. 1980;66:1391–1399. doi: 10.1172/JCI109992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chang CP, et al. Elevated cell-free serum DNA detected in patients with myocardial infarction. Clin Chim Acta. 2003;327:95–101. doi: 10.1016/s0009-8981(02)00337-6. [DOI] [PubMed] [Google Scholar]