

Abstract
Voltage responses of insect photoreceptors and visual interneurons can be accurately recorded with conventional sharp microelectrodes. The method described here enables the investigator to measure long-lasting (from minutes to hours) high-quality intracellular responses from single Drosophila R1-R6 photoreceptors and Large Monopolar Cells (LMCs) to light stimuli. Because the recording system has low noise, it can be used to study variability among individual cells in the fly eye, and how their outputs reflect the physical properties of the visual environment. We outline all key steps in performing this technique. The basic steps in constructing an appropriate electrophysiology set-up for recording, such as design and selection of the experimental equipment are described. We also explain how to prepare for recording by making appropriate (sharp) recording and (blunt) reference electrodes. Details are given on how to fix an intact fly in a bespoke fly-holder, prepare a small window in its eye and insert a recording electrode through this hole with minimal damage. We explain how to localize the center of a cell's receptive field, dark- or light-adapt the studied cell, and to record its voltage responses to dynamic light stimuli. Finally, we describe the criteria for stable normal recordings, show characteristic high-quality voltage responses of individual cells to different light stimuli, and briefly define how to quantify their signaling performance. Many aspects of the method are technically challenging and require practice and patience to master. But once learned and optimized for the investigator's experimental objectives, it grants outstanding in vivo neurophysiological data.
Keywords: Neuroscience, Issue 112, Sharp microelectrode, current-clamp, electrophysiology, signaling performance, neural response, insect vision, signal-to-noise ratio, neuroethology, naturalistic light stimulation
Introduction
The fruit fly (Drosophila melanogaster) compound eye is a great model system to investigate the functional organization of photoreceptor and interneuron arrays for neural image sampling and processing, and for animal vision. The system has the most complete wiring diagram1,2 and is amiable to genetic manipulations and accurate neural activity monitoring (of high signal-to-noise ratio and time-resolution)3-10.
The Drosophila eye is modular, containing ~750 seemingly regular lens-capped structures called ommatidia, which together provide the fly a panoramic visual field that covers almost every direction around its head. The eye's primary information sampling units are its rhabdomeric photoreceptors7,8,11. Each ommatidium contains eight photoreceptor cells (R1-R8), which share the same facet lens but are aligned to seven different directions. Whilst the outer photoreceptors R1-R6 are most sensitive to blue-green light, spectral sensitivities of the inner cells R7 and R8, which lie on top of each other and point to the same direction, exhibit three distinctive subtypes: pale, yellow and dorsal rim area (DRA)12-15.
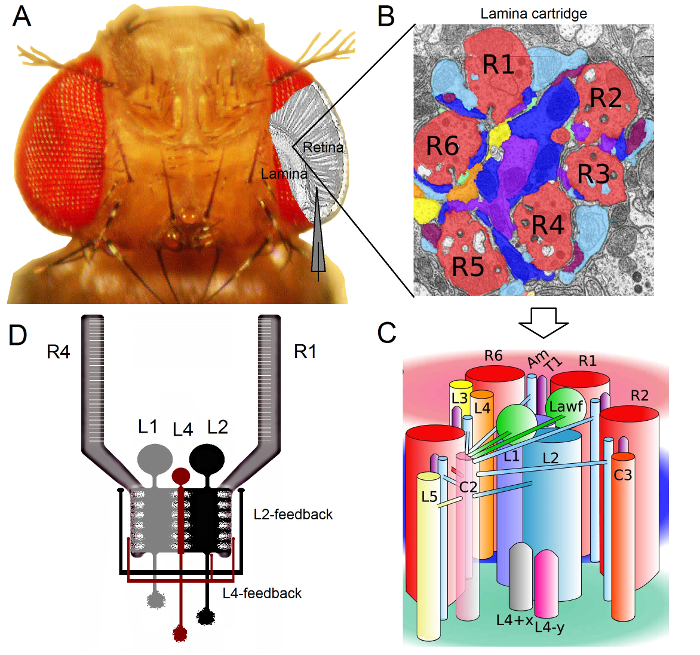 Figure 1.Functional Organization of theDrosophila Eye. (A) The two first optic ganglia, retina and lamina, are highlighted in gray inside the fly eye. Retina R1-R6 photoreceptors and lamina Large Monopolar Cells (LMCs: L1-L3) are readily accessible in vivo to conventional sharp microelectrode recordings. The schematic electrode highlights the normal path to record from R1-R6 in the retina. One path to record from LMCs in the lamina is to shift in parallel the electrode to left. (B) Lamina is a matrix of retinotopically organized cartridges, each of which is packed with neurons that processes information from a specific small area in the visual space. Due to neural superposition, six photoreceptors from different neighboring ommatidia send their axons (R1-R6) to the same lamina cartridge, forming histaminergic output synapses to L1-L3 and an amacrine cell (Am). (C) The spread of neural information between R1-R6 axon terminals and the visual interneurons (including L4, L5, Lawf, C2, C3 and T1), inside a lamina cartridge is complex. (D) R1-R6 photoreceptor axons receive synaptic feedbacks from L2 and L4 monopolar cells. (B) and (C) modified from Rivera-alba et al2. Please click here to view a larger version of this figure.
Figure 1.Functional Organization of theDrosophila Eye. (A) The two first optic ganglia, retina and lamina, are highlighted in gray inside the fly eye. Retina R1-R6 photoreceptors and lamina Large Monopolar Cells (LMCs: L1-L3) are readily accessible in vivo to conventional sharp microelectrode recordings. The schematic electrode highlights the normal path to record from R1-R6 in the retina. One path to record from LMCs in the lamina is to shift in parallel the electrode to left. (B) Lamina is a matrix of retinotopically organized cartridges, each of which is packed with neurons that processes information from a specific small area in the visual space. Due to neural superposition, six photoreceptors from different neighboring ommatidia send their axons (R1-R6) to the same lamina cartridge, forming histaminergic output synapses to L1-L3 and an amacrine cell (Am). (C) The spread of neural information between R1-R6 axon terminals and the visual interneurons (including L4, L5, Lawf, C2, C3 and T1), inside a lamina cartridge is complex. (D) R1-R6 photoreceptor axons receive synaptic feedbacks from L2 and L4 monopolar cells. (B) and (C) modified from Rivera-alba et al2. Please click here to view a larger version of this figure.
The Drosophila eye is of the neural superposition type16. This means that the neural signals of eight photoreceptors belonging to seven neighboring ommatidia, which look at the same point in space, are pooled together at one neural cartridge in the next two neuropils: the lamina and medulla. While the six outer photoreceptors R1-R6 project their axon terminals to neural columns in the lamina (Figure 1), R7 and R8 cells bypass this layer and make synaptic contacts with their corresponding medulla column17-19. These exact wirings produce the neural substrate for the retinotopic mapping of fly early vision, whereupon every lamina (Figures 1A-C) and medulla column (cartridge) represents a single point in space.
Direct inputs from R1-R6 photoreceptors are received by the Large Monopolar Cells (LMCs: L1, L2 and L3) and the Amacrine Cell (Am) in the lamina1,2,20. Out of these, L1 and L2 are the largest cells, mediating major information pathways (Figure 1D), which respond to On- and Off-moving edges, and thus form the computational basis of the motion detector21,22. Behavioral experiments suggests that at intermediate contrast, the two pathways facilitate motion perception of opposite directions: back-to-front in L1 and front-to-back in L2 cells23,24. Connectivity further implies that L4 neurons may play critical role in the lateral communication between neighboring cartridges25,26. Reciprocal synapses were found between L2 and L4 cells located in the same and two adjacent cartridges. Downstream, each L2 cell and its three associated L4 cells project their axons to a common target, the Tm2 neuron in the medulla, where inputs from neighboring cartridges are believed to be integrated for processing of front-to-back motion27. Although L1 neurons receive input from same-cartridge L2s via both gap junctions and synapses, they are not directly connected to L4s and hence adjacent lamina cartridges.
Synaptic feedbacks to R1-R6 photoreceptor axons are provided only by neurons belonging to the L2/L4 circuits but not the L1 pathway1,2 (Figure 1D). Whilst same-cartridge connections are selectively from L2 to R1 and R2 and from L4 to R5, all R1-R6 photoreceptors receive synaptic feedback from L4 of either or both neighboring cartridges. Furthermore, there are strong synaptic connections from Am to R1, R2, R4 and R5, and glia cells are also synaptically connected to the network and may thus participate in neural image processing6. Finally, axonal gap-junctions, linking neighboring R1-R6 and between R6 and R7/R8 photoreceptors in the lamina, contribute to the asymmetric information representation and processing in each cartridge14,20,28.
Intracellular voltage recordings from individual photoreceptors and visual interneurons in nearly intact Drosophila provide high signal-to-noise ratio data at sub-millisecond resolution3,5,7-10,29, which is necessary for making sense of the fast neural computations between the connected neurons. This level of precision is impossible by current optical imaging techniques, which are significantly noisier and typically operate at 10 - 100 msec resolution. Furthermore, because the electrodes have very small and sharp tips, the method is not restricted to cell bodies, but can provide direct recordings from small active neural structures; such as the LMCs' dendritic trees or photoreceptor axons, which cannot be accessed by much larger tips of patch-clamp electrodes. Importantly, the method is also structurally less invasive and damaging than most patch-clamp applications, and so affects less the studied cells' intracellular milieu and information sampling. Thus, conventional sharp microelectrode techniques have contributed, and keep on contributing, fundamental discoveries and original insight into neural information processing at the appropriate time scale; improving our mechanistic understanding of vision3-10.
This article explains how in vivo intracellular recordings from Drosophila R1-R6 photoreceptors and LMCs are performed in the Juusola laboratory. This protocol will describe how to construct a suitable electrophysiology rig, prepare the fly, and perform the recordings. Some representative data is presented, and some common issues and potential solutions are discussed that may be encountered when using this method.
Protocol
The following protocol complies with all the animal care guidelines of The University of Sheffield and Beijing Normal University.
1. Reagents and Equipment Preparation
- Recording and Light Stimulation Equipment Setup
- Choose at least a 2.5 x 2.5 m recording area for performing electrophysiological experiments in a room that has air conditioning with regulated humidity and means to provide dark recording conditions. Ensure that this area is large enough to comfortably fit a : (i) 1 x 1 m vibration-isolation table that houses the rig [fly stimulation and recording apparatus], stereomicroscope and a cold light source with two goose necks, all enclosed within a large >180 cm tall Faraday cage; (ii) a 38U equipment rack for housing a personal computer with a flat LCD monitor, microelectrode amplifier, LED drivers, filters, temperature control units, oscilloscopes and other required electrical instruments; and (iii) a small desk and a chair for the investigator.
- Place the rig far away from electrical and mechanical noise sources, such as refrigerators, centrifuges and elevators. Use separate surge protectors to protect the rig's electrical devices from voltage spikes occurring in the mains. Ideally, connect the rig to its own uninterruptible power supply (UPS battery) to minimize noise.
- Construct a conical fly-holder out of brass and black plastic (Figure 2). Drill a small tapering hole through the brass unit with its external rim narrowing to ~0.8 mm diameter (matching a typical fly's thorax width). Note: This hole needs to taper toward the tip of the fly-holder so that a larger than average Drosophila, which is projected from below by airflow, would get stuck shoulder-deep at the top rim.
- Design and build a mechanically-robust, yet precise, fly stimulation and recording apparatus (Figure 3). Construct out of aluminum or brass (high conductivity metals) a fly preparation platform pole and around it a Cardan-arm system, with embedded ball bearings, to provide smooth and accurate x,y-positioning and locking of the light stimulus. Note: This integrated composite design minimizes mechanical vibrations, which could otherwise dislodge the recording electrode tip out from the studied cell. It can further incorporate a Peltier-element-based close-loop temperature control system, enabling investigators to use temperature-sensitive genetic constructs, such as shibireTS, for assessing synaptic circuit computations9,30. Anodize the apparatus or paint it black to minimize light stimulus scatter.
- Fix the fly stimulation and recording apparatus on the anti-vibration table's breadboard; for example by M6-bolts, using its metric screw holes. Use a black breadboard or cover it with black fabric to minimize light scatter during experiments.
- Position and lock (using a locking screw) a vertically adjustable fly preparation platform pole at center of a Cardan-arm system. Place the fly-preparation (within the fly-holder, see step 2) on the platform pole so that the light source attached to the Cardan-arm radially points to the fly's head. Ensure that the center of the fly eyes is exactly at the intersection point (0, 0) of the Cardan-arm's x- and y-axes, as this enables accurate x,y-positioning of the light stimulus to any point within the fly's visual field. Note: This functionality is necessary for mapping the response properties of individual cells to specific eye locations; e.g., when searching for electrophysiological evidence for structural adaptations, such as bright or acute zones, which would show increasing sensitivity or resolution, respectively31.
- Mount the stereomicroscope behind the fly stimulation and recording apparatus on the anti-vibration table so that it provides comfortable high magnification viewing of the fly eye.
- Mount the cold light source on the top of the microscope with the light source's dual head semi-rigid gooseneck light guides pointing down toward the fly preparation holder. Freely movable two beam illumination makes it easier to visualize the recording electrode tip when driving it through a small opening into the fly eye.
- Attach an appropriate x,y,z-micromanipulator set (coarse & fine) for the recording electrode and the head-stage on the anti-vibration table, at the right side of the fly stimulation and recording apparatus, using M6-bolts or magnetic stands. Note: In the Juusola laboratory, different rigs are equipped with different manipulators; for details see the Table of Materials and Reagents. These all provide high-quality intracellular recordings.
- Mount a small manual 3-axis micromanipulator for the reference electrode holder on the vertically adjustable fly preparation platform pole. Orient the reference electrode so that it is pointing toward the fly preparation.
- Construct a free-standing light-shielded Faraday cage out of steel-panels around the anti-vibration table, surrounding the fly stimulation and recording apparatus, to prevent outside electromagnetic interference. Leave the front of the cage open, providing access to transport the fly preparation for the experiments. Attach black fabric curtains (having copper- or aluminum-mesh implanted inside them for grounding) at the front to shield out noise and light. Paint the interior of the cage black to minimize light scatter and bolt the feet of the cage on the floor to prevent vibrations.
- Connect the voltage and current outputs of the high-impedance intracellular microelectrode amplifier to the inputs of two separate low-pass filters (Bessel or similar) using BNC-cables. Likewise, connect the filter outputs into the appropriate channels of the AD-connector blocks/boards of the data acquisition system (DA/AD cards). Connect the DA/AD card(s) into a personal computer by specialized cables, according to supplier manuals.
- Install appropriate acquisition software for the data acquisition system of choice on the personal computer. Ensure that the data acquisition drivers are compatible with the operating system on the personal computer.
- Ground electrically the fly stimulation and recording apparatus, Faraday cage, copper mesh (within the curtains), microscope, micromanipulators, cold light source, 38U equipment rack with all its instruments (the intracellular amplifier, filters, temperature control unit, PC and LCD monitor etc.) to a single central ground point by using equipment grounding wire and M6 crimp ring grounding ends. Use an electric multimeter to test that all the parts are in the same ground. Note: To achieve the best possible low-noise recording conditions, the grounding configurations typically vary from one set-up to another.
- If needed, connect the central ground point further to the building ground, and/or the microelectrode amplifier's central ground. After testing the fully functioning system during real electrophysiological experiments, be prepared to change the grounding configuration as needed to minimize noise in the recordings.
- Configure software amplification (1 - 10X), signal filtering (typically low-pass filters set at 500 Hz, which is suitable for both R1-R6 and LMC data), and sampling rate (at least 1 KHz). Ensure that the settings obey Nyquist-Shannon sampling theorem32; for example, when acquiring data that is low-pass-filtered at 500 Hz, use a sampling frequency of 1 kHz or higher to minimize aliasing effects.
- As characteristic voltage responses of R1-R6 photoreceptors are 40 - 65 mV, and those of LMCs 20 - 45 mV, set the amplification and display scales accordingly to enable high-resolution sampling and data visualization.
 Figure 2. Conical Fly-holder. The fly-holder is made out of two pieces: the central brass unit and its conical black plastic coat. The central hole inside the brass unit tapers to a small diameter that barely lets the fly through. Please click here to view a larger version of this figure.
Figure 2. Conical Fly-holder. The fly-holder is made out of two pieces: the central brass unit and its conical black plastic coat. The central hole inside the brass unit tapers to a small diameter that barely lets the fly through. Please click here to view a larger version of this figure.
 Figure 3. Overview of the Electrophysiological Rig. The set-up contains a free-standing light-shielded Faraday cage, the anti-vibration table, the fly stimulation and recording apparatus, and black fabric curtains with copper- or aluminum-mesh inside for grounding. The instrument rack is electrically connected to the same central ground with all the equipment inside the Faraday cage. Please click here to view a larger version of this figure.
Figure 3. Overview of the Electrophysiological Rig. The set-up contains a free-standing light-shielded Faraday cage, the anti-vibration table, the fly stimulation and recording apparatus, and black fabric curtains with copper- or aluminum-mesh inside for grounding. The instrument rack is electrically connected to the same central ground with all the equipment inside the Faraday cage. Please click here to view a larger version of this figure.
- Fabricating Microelectrodes
- Pull the reference microelectrode from filamented borosilicate (outer diameter: 1.0 mm; inner diameter 0.6 mm) or quartz glass (outer diameter: 1.0 mm; inner diameter 0.5, 0.6 or 0.7 mm) tubing using a pipette puller instrument. Try to achieve a short gradual taper. Note: The exact settings of the pipette puller program vary from instrument to instrument; more details in the Table of Materials and regents. The pore size at the tip is not crucial because the tip of the reference electrode will be broken before being inserted into the fly preparation.
- Pull the recording microelectrode from filamented borosilicate (outer diameter: 1.0 mm; inner diameter 0.6 mm) or quartz glass (outer diameter: 1.0 mm; inner diameter 0.5, 0.6 or 0.7 mm) tubing using a pipette puller instrument. Try to achieve a long (10 - 15 mm) fine gradual taper.
- Inspect with a light microscope that the recording electrodes show correct tapering. Mount the electrode on a glass slide with moldable adhesive and use 40X air objective to inspect its tip. Note: A good electrode tapers smoothly until its invisibly small tip, around which continuous parallel darker and lighter interference patterns can be seen. Some puller settings generate high resistance electrodes, which cannot yield successful cell penetrations because their tips resemble "trumpets". Thus, visual inspection of the electrodes is important.
- Attach the electrodes horizontally on a large Petri-dish with modelling clay (or similar moldable adhesive) for safe-keeping and transportation to the electrophysiology rig. Ensure that the electrode tips are always in the air and not accidentally touching anything.
- Back-fill the recording and reference electrodes just before the experiment with the appropriate salt solution. Use a small 5 ml syringe connected to a small particle filter with a fine plastic tip (such as a microloader).
- For photoreceptors experiments, fill the recording electrode until full (a droplet forms in its large end) with 3 M KCl as this solution minimizes the effect of liquid junction potential to the recorded voltage.
- For investigating the histaminergic LMCs, which respond to synaptic input from R1-R6 photoreceptors by chloride-conductance changes, fill the recording electrodes with 3 M potassium acetate and 0.5 mM KCl, as this solution has less effect on the cell's chloride battery. Fill the reference electrode with fly Ringer, containing in mM: 120 NaCl, 5 KCl, 10 TES (C6H15NO6S), 1.5 CaCl2, 4 MgCl2, and 30 sucrose5.
- Test the resistance of a newly pulled recording electrode in the recording system.
- Ensure that the silver wires inside the electrode-holders are evenly coated with silver chloride (appearing purple-grey - not shiny silvery) to minimize recording artefacts (such as drift in the junction potential). If not, replace them with properly chloridized wires.
- If needed, chloridize new silver wires. Carefully clean the wires (by quickly passing them through a flame) so that these appear bright silver in color. Avoid touching them with the fingers, in order to deposit on an even layer of AgCl. Soak the wires in full strength household bleach for 15 - 30 min until they appear purple-gray color. Alternatively, electroplate each wire (by making it positive with respect to a solution containing 3 M KCl and passing a current through it at a rate of 1 mA/cm2 of surface area) for 10 - 15 sec until adequately coated.
- Connect the back-filled recording and reference electrodes to their electrode-holders. Place a small Ringer's solution bath on the vertically adjustable fly preparation platform pole. Drive the electrode tips into the Ringer's solution and measure the recording electrode's tip resistance. Note: This step is only needed when testing the resistive properties of electrodes, which are pulled from a new batch of glass tubing, or when optimizing the microelectrode puller instrument programs through iteration.
- Before performing the resistance measurements, read the instructions in the amplifier manufacture's user manual for the appropriate measurement settings. For a good recording electrode, have a tip resistance of ~100 - 220 MΩ.
2. Drosophila Preparation
Collect 5 - 10 days old flies (after eclosion) and place them in a clean fly tube containing standard food. It is possible to achieve good recordings from younger flies too, even from the "newborns"; but because of their softer eyes, cutting a corneal opening for the recording electrode is more difficult.
- Construct a fly catching tube and a fly preparation stand (Figure 4). See Figure 4 for the general idea of how these self-made tools were put together.
- To make a fly catching tube, cut off the tip of 50 ml plastic centrifuge tube's conical bottom. Then, insert and glue the large end of 1 ml pipette tip on this new opening.
- Finally, cut the small end of pipette to a size that readily lets a fly to walk through. Consult a mechanical workshop to assemble a small fly preparation stage that enables 2-axes rotation and locking of the fly-holder to different positions.
 Figure 4. Tools and Devises Needed for Making the Fly Preparation. Fly catching tube is made by gluing a 1 ml plastic pipette tip to a 50 ml plastic centrifuge tube. Bespoke fly preparation stand enables free-rotation and locking of the fly-holder in a preferred position for preparing the fly. The fly is fixed by beeswax, using the electric wax-heater. Petroleum jelly is applied by a small applicator made by connecting a thick sort hair on a handle. Please click here to view a larger version of this figure.
Figure 4. Tools and Devises Needed for Making the Fly Preparation. Fly catching tube is made by gluing a 1 ml plastic pipette tip to a 50 ml plastic centrifuge tube. Bespoke fly preparation stand enables free-rotation and locking of the fly-holder in a preferred position for preparing the fly. The fly is fixed by beeswax, using the electric wax-heater. Petroleum jelly is applied by a small applicator made by connecting a thick sort hair on a handle. Please click here to view a larger version of this figure.
Collect a fly for the experiment in a 1 ml pipette tip, which barely enables a fly to fit through. Attach the fly catching tube, with the pipette tip on it, to the fly tube. In catching a fly, take advantage of their inherent tendency to climb upwards (antigravitaxis) into the pipette tip. Preferably, select the biggest female, as size matters in electrophysiology. Note: The bigger the fly, the bigger its cells and the better the chances for high quality intracellular recordings. Smaller flies (both females and males) can also provide excellent recordings, but the preparation is more difficult to make. Once the fly is trapped in a large pipette tip, remember to close the fly tube to stop other flies from escaping.
Connect a 100 ml syringe with a flexible plastic hose to the larger opening of the pipette tip - with the fly still in it.
- Place the narrow end on the large pipette tip, which is enlarged just to let a Drosophila through, to the opening in the bottom of the fly-holder and squeeze a small volume of air from the syringe to eject the fly into the fly-holder.
- Look through the stereomicroscope and gently administer more air until the fly's head is protruding from the conical end of the fly-holder. Ensure that the fly is firmly trapped from its thorax to the small opening on the top of the fly-holder.
Use a wax heater to fasten the fly with beeswax from its "shoulders" to the fly-holder. Adjust the temperature of the wax heater to be as low as possible yet cleanly melting the wax. Note: When the temperature is correct, the wax appears transparent. Too high of a temperature makes the wax "burn away"; too low keeps the wax stiff. When fixing the fly, be accurate and brief as prolonged heat exposure may damage it. Using light-cured dental glue is not recommended here as its application is too slow.
 Figure 5. Preparing the Fly for In Vivo Experiments. Left, A Drosophila's head is positioned straight in the fly-holder and fixed from its proboscis, right eye and shoulders to the fly-holder with heated beeswax. Right, A small opening is cut in the thickest part of the eye, just above the equator and only a few ommatidia away from the back cuticle, using a sharp razor edge. A piece of cornea is gently removed and the hole is sealed with petroleum jelly to prevent the eye from drying up. Please click here to view a larger version of this figure.
Figure 5. Preparing the Fly for In Vivo Experiments. Left, A Drosophila's head is positioned straight in the fly-holder and fixed from its proboscis, right eye and shoulders to the fly-holder with heated beeswax. Right, A small opening is cut in the thickest part of the eye, just above the equator and only a few ommatidia away from the back cuticle, using a sharp razor edge. A piece of cornea is gently removed and the hole is sealed with petroleum jelly to prevent the eye from drying up. Please click here to view a larger version of this figure.
Immobilize the fly's head. Apply beeswax to the proboscis (Figure 5) and the corner of the right eye, avoiding the cornea, and fix the head from these points to the fly-holder.
Produce a micro-knife. Clamp a non-stainless-steel razorblade with two blade-holders/breakers (both with flat grip) and crack a small strip of its sharp edge. For health and safety, use goggles for eye protection (even though it is highly unlikely that pieces would ricochet when the razor is cracked). Ideally, produce a sharp razor edge that resembles a spire. Ensure that this "spire" is firmly attached to the blade-holder, but be careful to avoid any self-injury!
Using the micro-knife, prepare a small opening size of few ommatidia in the fly's left eye - at about 4 - 5 ommatidia from the dorsal cuticle just above the eye's equator to provide the passage for the recording microelectrode. Perform under a stereomicroscope, viewing the preparation with high magnification. Note: Because the fly eye feels elastic and resistive to probing, the hole is best cut with a "spire"-knife. The cutting technique is quite challenging, so pay close attention to the video demonstration. Keeping the fly-holder in certain orientations (in the fly preparation stand) can make the dissection easier. Initially, the microsurgery may feel difficult to learn, but once committed, neural adaptation gradually improves the investigator's 3D-perception and dexterity.
Remove carefully the small piece of cornea from the opening that was just cut, exposing the retina underneath. Swiftly cover the hole in the eye with a tiny blob of petroleum jelly using the fine hair of the petroleum jelly applicator. Note: Petroleum jelly serves multiple roles here. It prevents tissue dehydration and coagulation of the hemolymph that would break the inserted recording electrode. It also incidentally coats the microelectrode, reducing its intramural capacitance. This can improve the frequency response of the recording system, and so the temporal resolution of the recorded neural signals. Avoid smearing petroleum jelly over the rest of the eye as this blurs the optics.
3. Recording from R1-R6 Photoreceptors or LMCs
Always be grounded when operating the microelectrode amplifier (for example by touching the metal surface of the Faraday cage or anti-vibration table), as this precludes one from accidentally delivering a static charge to the head-stage, which could damage the circuitry.
Illuminate the fly preparation platform pole from above by two goose-neck light guides (Figure 6A) (with the cold light source inside the Faraday cage) so that the fly-holder can be placed on the pole in the preferred position under close visual control.
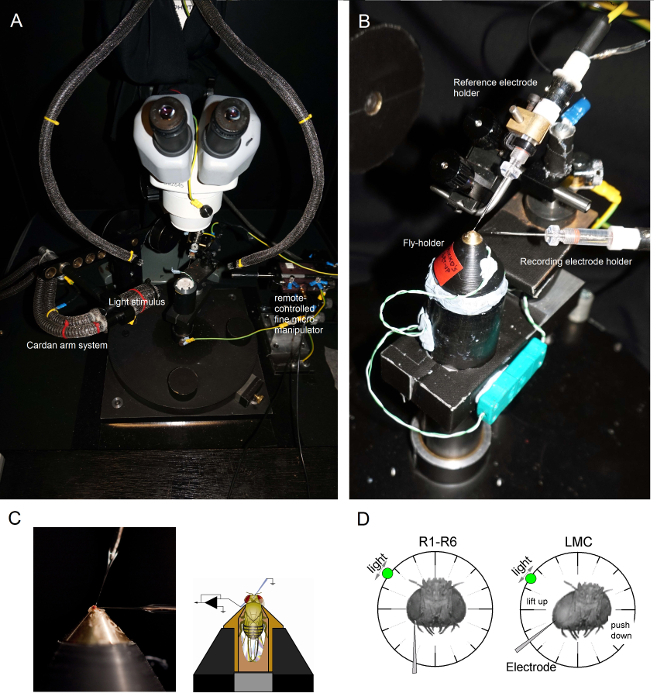 Figure 6.Positioning the Fly-holder and the Electrodes for the Experiments. (A-B) The fly-holder is placed on the recording platform that also provides temperature control via a Peltier element (A: white round platform in the center). The Cardan-arm enables exact positioning of the light stimulus at an equal distance (via x,y-rotation) around the fly, with the light source (a liquid or quartz fiber-optic bundle end) directly pointing to its eye. In many of our rigs, light stimulation is generated by LEDs (with linear current-drivers) or by a monochromator. Thus, their stimuli carry a specific (band-passed) spectral content, selected between 300 - 740 nm and cover 4 - 6 log intensity unit range (as attenuated by separate neutral density filters). (C) Two microelectrodes, controlled by separate micromanipulators, are positioned in the fly head: the reference electrode (above) through the ocelli; the recording electrode (left) through the small opening in the left eye. (D) For obtaining a maximum number of photoreceptor recordings, the recording microelectrode is driven into the hole, parallel to the proboscis-ocellus axis. When the electrode tip penetrates and seals to a photoreceptor, the freely rotatable light source is fixed to the position where the cell produces the maximum voltage response to a given light stimulus. This point in space lies in the center of the cell's receptive field. If the hole is close to the cuticle, LMC penetrations can further be achieved with this same electrode angle (left). If the hole is further from the cuticle, another useful electrode approach angle to obtain LMC recordings is also shown (right). Please click here to view a larger version of this figure.
Figure 6.Positioning the Fly-holder and the Electrodes for the Experiments. (A-B) The fly-holder is placed on the recording platform that also provides temperature control via a Peltier element (A: white round platform in the center). The Cardan-arm enables exact positioning of the light stimulus at an equal distance (via x,y-rotation) around the fly, with the light source (a liquid or quartz fiber-optic bundle end) directly pointing to its eye. In many of our rigs, light stimulation is generated by LEDs (with linear current-drivers) or by a monochromator. Thus, their stimuli carry a specific (band-passed) spectral content, selected between 300 - 740 nm and cover 4 - 6 log intensity unit range (as attenuated by separate neutral density filters). (C) Two microelectrodes, controlled by separate micromanipulators, are positioned in the fly head: the reference electrode (above) through the ocelli; the recording electrode (left) through the small opening in the left eye. (D) For obtaining a maximum number of photoreceptor recordings, the recording microelectrode is driven into the hole, parallel to the proboscis-ocellus axis. When the electrode tip penetrates and seals to a photoreceptor, the freely rotatable light source is fixed to the position where the cell produces the maximum voltage response to a given light stimulus. This point in space lies in the center of the cell's receptive field. If the hole is close to the cuticle, LMC penetrations can further be achieved with this same electrode angle (left). If the hole is further from the cuticle, another useful electrode approach angle to obtain LMC recordings is also shown (right). Please click here to view a larger version of this figure.
Mount the fly-holder (with the fly in it!) on the fly preparation platform pole. Rotate the fly-holder so that the fly's left eye is directly facing the investigator (Figure 6B).
- Insert the blunt reference electrode gently through the fly's occelli into the head capsule using a small coarse micromanipulator while observing the preparation through the stereomicroscope ( Figure 6C). Do not push the electrode too deep, as this can damage the fly brain.
- Alternatively, insert the reference electrode into the back of the thorax. Always, ensure that the fly appears heathy (moves its antennae) and its eyes are intact; not accidentally damaged. If the preparation looks less than immaculate, prepare a new fly for the experiments.
Drive the sharp recording microelectrode into the left eye through the petroleum jelly covered small opening prepared earlier. Use high magnification in the stereomicroscope and move the light guides and the focal plane so that the electrode tip location becomes apparent in 3D by its reflectance patterns. Note: Figure 6D shows how the fly's head should be positioned slightly differently (in respect to the angle the recording microelectrode enters the eye) for photoreceptor and LMC recordings. Driving the electrode into the eye without breaking it is the most difficult phase of the experiment. If the electrode tip misses the small opening in the eye, hitting the cornea, it typically breaks.
Turn on the microelectrode amplifier once both electrodes are firmly inside the preparation, in electrical contact with the fly's body fluids.
Turn off the cold-light source (inside the Faraday cage) and unplug it from the mains. Connect its plug to the central ground to minimize ground-loop induced electrical noise, and move the goose-neck light guides away so that the Cardan-arm system can be freely moved around the fly. Switch off the room lights to ensure that the fly preparation is now in relative darkness.
- Measure the resistance of the recording electrode in the eye (as instructed in the amplifier's user manual). Use only recording electrodes in which resistance is 100 - 250 MΩ. Note: It is virtually impossible to achieve high quality intracellular recordings by <70 MΩ electrode. If the resistance is <80 MΩ, it is likely that the electrode tip is broken. In this case, switch off the amplifier and change the recording electrode.
- Once the electrode is replaced and in the eye, switch on the amplifier to measure its resistance. Sometimes, the electrode tip can become blocked by some detritus as it enters the tissue. This can be remedied by using the amplifier's capacitive buzz and current pulse functions that typically clear it by rapid resonation or repulsion.
Set the amplifier to current-clamp (CC) or bridge recording mode. Cancel out any arbitrary voltage difference between the recording and reference electrodes, as both of them are now resting in the electrically interconnected extracellular space, by setting the signal offset (recording voltage) to zero. Follow the signal offset changes using the amplifier's display readout or an oscilloscope screen.
Wait 2-3 min for the fly eye to dark-adapt.
Drive the recording electrode tip gradually deeper into the eye with small 0.1 to 1 micron steps. Do this with an x-axis piezo-stepper of a remote-controlled micromanipulator or by gently rotating the fine resolution knob of a manual manipulator.
Stimulate the fly eye with brief (1 - 10 msec) light flashes as the recording electrode is being advanced in the tissue. Note: If the recording electrode is positioned in the retina and the eye functions normally, each light flash will cause a brief and small drop in the voltage (0.2 - 5 mV hyperpolarization), called the electroretinogram (ERG). This change in the field potential of the extracellular space is caused by the retinal cells' collective response to light. However, once the electrode tip enters the lamina, closing on the LMCs, the ERG reverses, showing depolarizing responses.
Move the light source around the fly eye by using the Cardan-arm system and find the position where the light evokes the largest ERG response. Note: This position marks the small area in the visual space where the photoreceptors (or LMCs), which are located next to the tip of the recording electrode, sample their light input.
Penetrate a cell with the recording electrode. Note: The penetration can occur spontaneously, or when the electrode is micro-stepped forward. It can be further facilitated by gently tapping the micromanipulator system or by using the buzz-function of the amplifier; these actions resonate the electrode tip in the tissue. When the electrode impales the photoreceptor membrane, entering its intracellular space, the voltage difference between the recording and reference electrode drops suddenly from 0 mV to ~-65 mV (between -55 and -75 mV); whereas during LMC penetrations, this drop is typically less (between -30 and -50 mV). These voltage differences represent the negative resting potentials of the given cells. Depending on the quality of the recording electrode (its sharpness) and the cellular process it penetrated, the voltage reading from the recording electrode can stabilize rapidly or gradually to the resting potential, as the cell membrane seals to the outer layer of the electrode. But if the penetration is only partial or poor, the electrode typically slips out of the cell with the recorded potential climbing back towards zero.
Localize the center of the penetrated cell's receptive field when the electrode appears properly sealed, showing stable membrane potential (dark resting potential). Move the flashing light stimulus around the fly eye, using the Cardan-arm system, to find the point in visual space, where the light flash evokes the cell's maximum voltage response. Lock the Cardan-arm when the light stimulus directly faces (points at) the receptive field center. Note: In darkness, Drosophila photoreceptors respond to bright light pulses with 40 - 65 mV depolarizing voltage responses4,5, while stable LMC recordings show 20 to 45 mV hyperpolarizing responses9,10,14. Glia penetrations may happen rarely, indicated by <-80 mV resting potentials and much slower and smaller (~5 mV) saturated light-induced depolarizations. Photoreceptors in Drosophila with different eye pigmentations, such as white-eye7 and cinnabar, show comparable response sizes to wild-type.
Using the amplifier's current-clamp (CC) mode, compensate the recording electrode's capacitance by injecting small 0.1 nA and brief (100 - 200 msec) current pulses into the studied cell to minimize recording artefacts during its membrane charging. Note: This important procedure is explained in detail in the amplifier's user manual, and should be practiced with an electrical cell model before the actual experiments.
Record voltage responses to light pulses and other stimuli of interest, having varying statistical or physical qualities (such as naturalistic light intensity time series or random contrast patterns). Test, for example, how the recorded responses change with light- or dark-adaptation. Note: One can accurately light-adapt the studied cell by continuous light of preselected intensity by adding neutral density filters on the light path4,5. Alternatively, for prolonged dark-adaptation switch off the light stimulus for a preset time. Because of the mechanical stability of the recording system, the high quality of the recording electrodes and the intactness of the preparation, stable recording conditions can sometimes last for many hours. Thus, on a good day, it is possible to collect a large amount of data at different adapting conditions from a single cell. When the electrode slips out of the cell, the recorded responses diminish and the mean voltage begins to approach zero.
Advance carefully the recording electrode with the fine x-axis control of the micromanipulator until the electrode makes contact and penetrates the next cell (this is typically the nearest neural neighbor). Do not move the electrode along y- or z-axis as these maneuvers would make the electrode to "plough the tissue" sideways, damaging eye structures! Note: With a good electrode and a healthy preparation, one can record high quality responses from many photoreceptors (but rarely from many LMCs) in the same fly over a period of several hours; occasionally, over the whole working day (>8 hr) without clear signal deterioration.
Save data files periodically with identifying information, such as date, genotype, and the recorded cell type. Because of the large amount of data that can be collected in a successful recording session, keep good written records in a lab-book for future data analysis.
Representative Results
The sharp microelectrode recording method, as adapted here for the Drosophila eye, can be used to reliably quantify neural information sampling and processing in the retina and lamina cells, and communication between them4,5,7,8,10,33. By using it to study encoding in different wild-type stocks, mutants or genetically engineered fly strains, the method has proven its value; not only in quantifying the effects of a mutation, temperature, diet or selected expression3,4,6,9,10,14,30,34, but also in revealing mechanistic reasons for altered visual behaviors14,34. The method is also readily applicable to other insect preparations35,36, empowering neuroethological vision studies. Next we showcase a few examples of its successful applications.
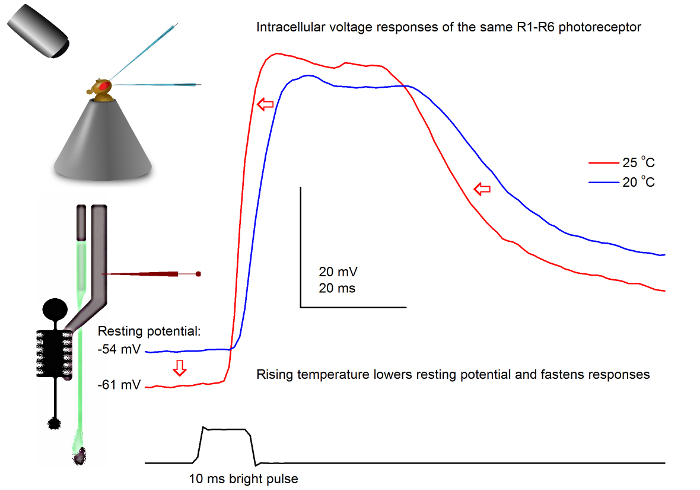 Figure 7.Voltage Responses of a Fruit Fly R1-R6 Photoreceptor to a Light Pulse at 20 and 25 oC. Because the sharp microelectrode penetrations are often very stable, it is possible to record voltage responses of the same R1-R6 photoreceptor to a given light stimulus at different ambient temperatures by warming or cooling the fly. In our set-ups, the fly-holder is placed on a close-loop Peltier-element-based temperature-control system. This enables us to change the fly's head temperature in seconds. Higher temperature accelerates the voltage responses and characteristically lowers the resting potential of R1-R6 photoreceptors (as indicated by red arrows). Please click here to view a larger version of this figure.
Figure 7.Voltage Responses of a Fruit Fly R1-R6 Photoreceptor to a Light Pulse at 20 and 25 oC. Because the sharp microelectrode penetrations are often very stable, it is possible to record voltage responses of the same R1-R6 photoreceptor to a given light stimulus at different ambient temperatures by warming or cooling the fly. In our set-ups, the fly-holder is placed on a close-loop Peltier-element-based temperature-control system. This enables us to change the fly's head temperature in seconds. Higher temperature accelerates the voltage responses and characteristically lowers the resting potential of R1-R6 photoreceptors (as indicated by red arrows). Please click here to view a larger version of this figure.
Studying the Effect of Temperature on Photoreceptor Output
With a well-designed and vibration-isolated recording system, the method can be used for measuring the effect of temperature on an individual cell's neural output by warming or cooling the fly. The given example shows voltage responses to a bright 10 msec long pulse, recorded in the same R1-R6 photoreceptor at 20 and 25 oC (Figure 7). As quantified before4,9, warming lowers a photoreceptor's resting potential in darkness, and accelerates its voltage responses.
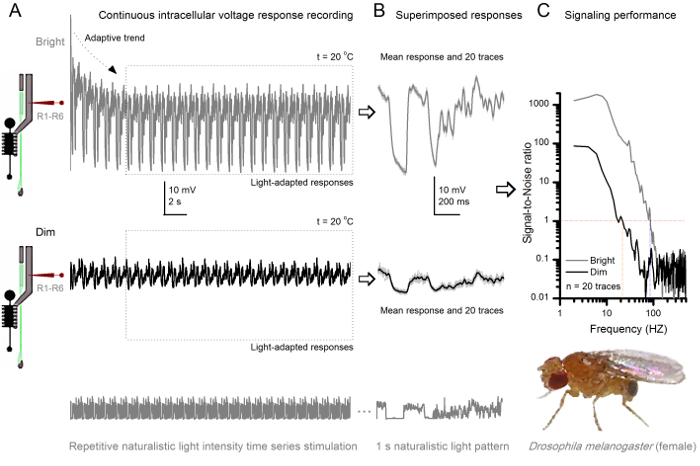 Figure 8.Signaling Performance a Fruit Fly R1-R6 Photoreceptor Improves with Light Intensity. (A) Photoreceptor output to dim (below) and bright (above; 10,000-times brighter light) repeated naturalistic light intensity time series recorded by the same microelectrode in the same cell at 20 oC. Responses to the bright stimulus are larger, because they integrate more samples, elementary responses (bumps) to single photons4,5,7,8. (B) 20 consecutive one-second-long voltage responses are superimposed. Individual responses (light gray) were taken after the adaptive trends (arrow in A) had receded (dotted box in A). The corresponding response means (the signals) are the darker traces. The difference between the signal and the individual responses is the noise. (C) The cells' signaling performance was quantified by the recordings' Signal-to-Noise Ratios (SNR) using the standard methods4,5,7,8. Photoreceptor output has about 64 Hz broader range of reliable signaling at the bright stimulation (SNR'Bright ≥1, up to 84 Hz ) than at the dim (SNR'Dim ≥1, up to 20 Hz), with signal-to-noise ratio improving greatly; from SNRDimMAX = 87 to SNRBrightMAX = 1,868. Please click here to view a larger version of this figure.
Figure 8.Signaling Performance a Fruit Fly R1-R6 Photoreceptor Improves with Light Intensity. (A) Photoreceptor output to dim (below) and bright (above; 10,000-times brighter light) repeated naturalistic light intensity time series recorded by the same microelectrode in the same cell at 20 oC. Responses to the bright stimulus are larger, because they integrate more samples, elementary responses (bumps) to single photons4,5,7,8. (B) 20 consecutive one-second-long voltage responses are superimposed. Individual responses (light gray) were taken after the adaptive trends (arrow in A) had receded (dotted box in A). The corresponding response means (the signals) are the darker traces. The difference between the signal and the individual responses is the noise. (C) The cells' signaling performance was quantified by the recordings' Signal-to-Noise Ratios (SNR) using the standard methods4,5,7,8. Photoreceptor output has about 64 Hz broader range of reliable signaling at the bright stimulation (SNR'Bright ≥1, up to 84 Hz ) than at the dim (SNR'Dim ≥1, up to 20 Hz), with signal-to-noise ratio improving greatly; from SNRDimMAX = 87 to SNRBrightMAX = 1,868. Please click here to view a larger version of this figure.
Studying Adaptation and Neural Encoding by Repetitive Stimulation
The noninvasiveness of the method, causing relatively little damage in the retina and lamina structures, makes it ideal for studying the signaling performance of individual cells to different light stimuli in their near natural physiological state in vivo. Figure 8 shows voltage responses of a R1-R6 photoreceptor to a dim and bright repeated naturalistic light intensity time series stimulus at 20 oC, whereas Figure 9 shows responses of another R1-R6 photoreceptor and a LMC to a different naturalistic stimulus at 25 oC. The pre- and postsynaptic recordings were performed separately from two different flies because simultaneous intracellular recordings by two sharp microelectrodes in the same fly, one in the retina and the other in the lamina, are too difficult to be viable30.
 Figure 9.Voltage Responses of a Fruit Fly R1-R6 Photoreceptor and LMC to Repeated Naturalistic Stimulation at 25 oC. (A) R1-R6 (gray) and LMC (black) outputs recorded by different microelectrodes from different flies. (B) Fully light-adapted 20 consecutive pre- (above) and postsynaptic (below) responses to the same naturalistic stimulus pattern with individual responses, shown in light gray and the corresponding response means (the signals) as the darker traces. The difference between the signal and the individual responses is the noise. (C) The cells' signaling performance was quantified by the recordings' Signal-to-Noise Ratios (SNR). LMC output has about 10 Hz broader range of reliable signaling (SNR'LMC ≥1, up to 104 Hz ) than R1-R6 output (SNR'R ≥1, up to 94 Hz). Both signal-to-noise ratios are high (SNRLMCMAX = 142, SNRRMAX = 752), and as the recording noise was low, their differences reflect real encoding differences between the cells. Please click here to view a larger version of this figure.
Figure 9.Voltage Responses of a Fruit Fly R1-R6 Photoreceptor and LMC to Repeated Naturalistic Stimulation at 25 oC. (A) R1-R6 (gray) and LMC (black) outputs recorded by different microelectrodes from different flies. (B) Fully light-adapted 20 consecutive pre- (above) and postsynaptic (below) responses to the same naturalistic stimulus pattern with individual responses, shown in light gray and the corresponding response means (the signals) as the darker traces. The difference between the signal and the individual responses is the noise. (C) The cells' signaling performance was quantified by the recordings' Signal-to-Noise Ratios (SNR). LMC output has about 10 Hz broader range of reliable signaling (SNR'LMC ≥1, up to 104 Hz ) than R1-R6 output (SNR'R ≥1, up to 94 Hz). Both signal-to-noise ratios are high (SNRLMCMAX = 142, SNRRMAX = 752), and as the recording noise was low, their differences reflect real encoding differences between the cells. Please click here to view a larger version of this figure.
After the stimulus onset, the recordings typically show fast adapting trends that largely subside within 5-6 sec. From then on, the cells produce highly consistent responses to each 1 sec long stimulus presentation (each dotted box encloses 20 of these). The repeatability of the responses becomes obvious when these are superimposed (Figure 8B and Figure 9B). Individual responses are the thin gray traces, and their mean the thicker darker trace. The mean response is taken as the neural signal, whereas the neural noise is the difference between the mean and each individual response4,5,9,37,38. The respective signal-to-noise ratios in frequency domain (Figure 8C and Figure 9C) were obtained by Fourier-transforming the signal and noise data chunks into power spectra, and dividing the mean signal power spectrum with the corresponding mean noise power spectrum4,5,9,37,38. Characteristically, the maximum signal-to-noise ratios of the recorded neural outputs to naturalistic stimulation are high (100 - 1,000), and in the most stable preparations with very low recording noise can reach values >>1,000 (e.g., Figure 8C). Notice also that warming expands the cells' bandwidth of reliable signaling4 (SNR'Bright≥ 1); for example, the relative difference between the two R1-R6s in Figures 8 and 9, respectively, is 10 Hz (84 at 20 oC and 94 Hz at 25 oC).
One can further estimate each cell's rate of information transfer from its signal-to-noise ratio by using the Shannon equation32, or by calculating the difference between the responses' entropy and noise entropy rates through the triple extrapolation method39. More details about the information theoretical analyses, and their use and limitations specifically with this method are given in the previous publications7,8,39.
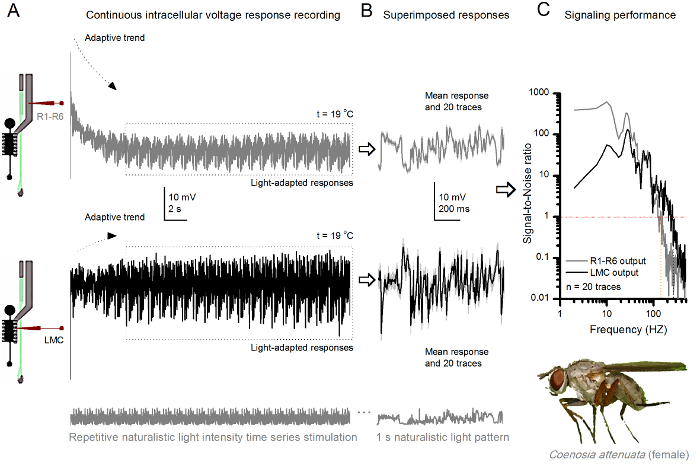 Figure 10. Voltage Responses of a Killer Fly R1-R6 Photoreceptor and LMC to Repeated Naturalistic Stimulation at 19 oC. (A) R1-R6 (gray) and LMC (black) outputs recorded by the same microelectrode from the same fly; first postsynaptically and later presynaptically, as the electrode was advanced in the eye. (B) 20 consecutive pre- (above) and postsynaptic (below) responses (light gray traces) to the same naturalistic stimulus pattern were captured after initial adaptation (dotted box in A). Their means are the signals (the darker traces on top), while their respective differences to the individual responses give the noise. (C) The corresponding Signal-to-Noise Ratios (SNR) were calculated as in Figures 8C and 9C. LMC output has about a 100 Hz broader range of reliable signaling (SNR'LMCMAX ≥1, up to 234 Hz) than R1-R6 output (SNR'RMAX ≥1, up to 134 Hz). Both signal-to-noise ratios are high (SNRLMCMAX = 137, SNRRMAX = 627), and as the same microelectrode was used in the recordings, their differences reflect real differences in the pre- and postsynaptic neural outputs. These results imply that the recording system had low noise, and its influence on the analyses was marginal. Please click here to view a larger version of this figure.
Figure 10. Voltage Responses of a Killer Fly R1-R6 Photoreceptor and LMC to Repeated Naturalistic Stimulation at 19 oC. (A) R1-R6 (gray) and LMC (black) outputs recorded by the same microelectrode from the same fly; first postsynaptically and later presynaptically, as the electrode was advanced in the eye. (B) 20 consecutive pre- (above) and postsynaptic (below) responses (light gray traces) to the same naturalistic stimulus pattern were captured after initial adaptation (dotted box in A). Their means are the signals (the darker traces on top), while their respective differences to the individual responses give the noise. (C) The corresponding Signal-to-Noise Ratios (SNR) were calculated as in Figures 8C and 9C. LMC output has about a 100 Hz broader range of reliable signaling (SNR'LMCMAX ≥1, up to 234 Hz) than R1-R6 output (SNR'RMAX ≥1, up to 134 Hz). Both signal-to-noise ratios are high (SNRLMCMAX = 137, SNRRMAX = 627), and as the same microelectrode was used in the recordings, their differences reflect real differences in the pre- and postsynaptic neural outputs. These results imply that the recording system had low noise, and its influence on the analyses was marginal. Please click here to view a larger version of this figure.
Neuroethological Vision Studies
The method can also be used to record pre- and postsynaptic voltage responses from the compound eyes of different insect species7,8,35,36 (Figure 10), permitting comparative neuroethological studies of visual information processing. For the presented recording system, the only required adaptation is new preparation-holders, each with a suitably-sized opening for the studied species. These exemplary recordings are from of a female killer fly (Coenosia attenuata). They show intracellular voltage responses of a R1-R6 photoreceptor and LMC to identical repetitive light stimulation, as used for the Drosophila counterparts in Figure 9, but at 19 oC. In this case, both the pre- and postsynaptic data were recorded from the same fly; one after the other, with the same recording electrode (filled with 3 M KCl) first advancing through the lateral lamina before entering the frontal retina. In comparison to the Drosophila data at 25 oC, the Coenosia data - even at the cooler temperature - shows responses with faster dynamics; expanding the range of reliable signaling (signal-to-noise ratio >>1) over a broader frequency range. Such functional adaptations in neural encoding of naturalistic stimuli are consistent with Coenosia's predatory lifestyle36, which require high-precision spatiotemporal information to attain fast aerial hunting behaviors.
Discussion
We have presented the basic key steps of how to use sharp conventional microelectrodes to record intracellular responses of R1-R6 photoreceptors and LMCs in intact fly eyes. This method has been optimized, together with bespoke hardware and software tools, over the last 18 years to provide high-quality long-lasting recordings to answer a wide range of experimental questions. By investing time and resources to construct robust and precise experimental set-ups, and to produce microelectrodes with favorable electrical properties, high-quality recordings can become the norm in any laboratory working on Drosophila visual neurophysiology. Whilst well-designed recording and light stimulation systems are important for swift execution of different experimental paradigms, there are three procedural steps that are even more critical to achieving successful recordings: (i) to make the fly preparation with minimal eye damage, (ii) to pull microelectrodes with the right electrical properties, and (iii) to drive the recording electrode into the eye without breaking its tip. Ultimately, to record meaningful data, the investigator has to understand the physical basis of electrophysiology and how to fabricate suitable microelectrodes for the targeted cell-types.
Therefore, the limitations of this technique are primarily set by the patience, experience and technical ability of the investigator. Because this technique can take a long time to master for small Drosophila cells, it is advisable for trainee electrophysiologists to first practice with larger insect eyes, such as the blowfly36 or locust35, using the same rig. Once performing high-quality intracellular recordings from the larger photoreceptors and interneurons becomes routine, it is time to move on to the Drosophila eye. Another limitation of the technique concerns cellular identification. Penetrated Drosophila cells can be loaded electrophoretically with dyes, including Lucifer yellow or neurobiotin. However, because of the small tip size of the microelectrodes, electrophoresis works less efficiently than with lower resistance electrodes, such as patch-electrodes. Furthermore, the dye-filled microelectrodes characteristically have less favorable electrical properties, making it much harder to record high-quality responses with them from Drosophila photoreceptors and LMCs.
A technical problem that occurs sometimes is unstable input signal, or a complete lack of it. This is often associated with the voltage signal being either constantly drifting or higher/lower than the amplifier's recording range. On most occasions, this behavior is caused by the recording electrode being blocked (or its tip being too fine - having too high a resistance or intramural capacitance - to properly conduct fast signal changes). Although one can try to unblock the tip by buzzing the electrode capacitance, which sometimes works, often the situation is best resolved by simply changing the recording electrode. This may further require parameter adjustments in the microelectrode puller instrument to lower the tip resistance of the new electrodes. The electrode tip can also become blocked in preparations, for which it took too much time to cover the corneal hole by petroleum jelly. Prolonged air-contact can dry up the freshly exposed retinal tissue, turning its surface layer into a glue-like substance. If this is the case, the investigator typically sees a red blob of tissue stuck on the recording electrode when pulling it out of the eye. The only solution here is to make a new preparation. Petroleum jelly may provide many benefits for electrophysiological recordings: (i) it prevents the coagulation of the hemolymph that could break the electrode tip; (ii) it coats the electrode tip reducing its intramural capacitance, which lowers the electrode's time constant, and thus has the potential to improve the temporal resolution of the recorded neural signals40,41; (iii) it keeps the electrode tip clean, facilitating penetrations; and after penetration, (iv) it may even help to seal the electrode tip to the cell membrane42.
The signal can further be unstable or lost when the silver-chloride wire of the electrode-holder is broken or dechloridized; in which case just replace or rechloridize the old wire. The missing signal can also result from one (or both) of the electrode-holders not being securely connected to their jacks. However, it is extremely unusual that a piece of equipment would be malfunctioning. If signal is undetectable and all other possibilities have been exhausted, test that each part of the recording apparatus, including the headstage, amplifier, low-pass filters and AD/DA-converters, are connected properly and functioning normally. One way to achieve this is to replace each instrument with another from a rig that is known to operate normally. Alternatively, use a signal generator to check the performance of the electronic components one by one.
But perhaps the most common technical problem facing the electrophysiologist is that of recording noise. Broadly, recording noise is the observed electrical activity other than the direct neuronal response to a given stimulus. Because the fly preparation, when properly done, is very stable, the observed noise (beyond the natural variably of the responses) most often results from ground-loops in the recording equipment, or is picked up from nearby electrical devices. Such noise is typically 50/60 Hz mains hum and its harmonics; but sometimes composed of more complex waveforms. To work out the origin of the noise, remove the fly preparation holder from the set-up, connect the recording and reference electrodes through a drop of fly Ringer (or place them in a small Ringer's solution bath; see step 1.2.6) and record the signal in CC- or bridge-mode. If noise is observable on the recorded signal, this likely means that the noise is external to the fly preparation.
Another good test for identifying the origin of noise is to replace the electrode-holders with an electric cell model connected to the amplifier. In an ideally configured and grounded set-up, the recorded signal should now be practically noise-free, showing only stochastic bit-noise from the AD-converter (in the best case not even that!). If noise is still present, then recheck that all rig equipment is properly grounded. A convenient approach to detect ground-loops is to: (i) disconnect all the grounding wires from all the parts within the rig; (ii) ensure that, after doing this, every single part is actually isolated from ground, by means of an ohm-meter; (iii) connect the parts, one by one, to the central ground directly, not through any other part of the rig. Try also changing the equipment configurations. For example, sometimes moving the computer and monitor further away from the rig can reduce noise; yet at other times, moving the computer inside the equipment rack reduces noise. It is also worth unplugging nearby equipment to see if noise is reduced, or shield additional components. Furthermore, try unplugging or replacing different components of the recording equipment, especially BNC cables (which can have faulty ground connections). If only bit-noise is observed when using the cell model, the initial noise source is either the electrodes or the fly preparation itself. For example, it could be that the reference electrode is inadvertently touching a motor nerve or active muscle fibers inside the head capsule (or disturbing flight muscles in the thorax - if placed there). It is usually simplest to prepare a new fly for recording, taking care to minimize damage to the fly. But if the noise persists and is broadband, it is likely that the electrodes are suboptimal for the experiments; too sharp/fine (hence too noisy) or just wrong for the purpose; we have even seen quartz-electrodes acting as antennas - picking up faint broadcasting signals! Although iteration of the puller-instrument parameter settings to generate the just right microelectrodes for consistent high-quality recordings from specific cell-types can take a lot of effort, it is worth it. Once the recording electrodes are well-tailored for the experiments, they can provide long-lasting recordings of outstanding quality.
Sharp microelectrode recording techniques can be similarly applied to study neural information processing in multitude of preparations, including different processing layers in the insect eyes and brain43,44. Because the microelectrode tips can be made very fine, these typically damage the studied cells less than most patch-clamp applications. Importantly, the modern sample-and-hold microelectrode amplifiers enable good control of the tips' electrical properties40,45-47. Thus, when correctly applied, this technique can provide reliable data from both in vivo3,5,7-10,44 or in vitro48 preparations with high signal-to-noise ratio at sub-millisecond resolution. Such precision would be impossible with today's optical imaging techniques, which are noisier and slower. Moreover, the method can be used to characterize small cells' electrical membrane properties both in current- and voltage-clamp configurations5,29,33,36,40-42,49, providing valuable data for biophysical and empirical modeling approaches7,8,11,33,49-54 that link experiments to theory.
Disclosures
The authors have nothing to disclose
Acknowledgments
The authors thank Mick Swann, Chris Askham and Martin Gautrey for their important contributions in designing and building many electrical and mechanical components of the rigs. MJ's current research is supported by the Biotechnology and Biological Sciences Research Council (BBSRC Grant: BB/M009564/1), the State Key Laboratory of Cognitive Neuroscience and Learning open research fund (China), High-End Foreign Expert Grant (China), Jane and Aatos Erkko Foundation Fellowship (Finland), and the Leverhulme Trust grant (RPG-2012-567).
References
- Meinertzhagen IA, O'Neil SD. Synaptic Organization of Columnar Elements in the Lamina of the Wild-Type in Drosophila-Melanogaster. J Comp Neurol. 1991;305:232–263. doi: 10.1002/cne.903050206. [DOI] [PubMed] [Google Scholar]
- Rivera-Alba M, et al. Wiring Economy and Volume Exclusion Determine Neuronal Placement in the Drosophila Brain. Curr Biol. 2011;21:2000–2005. doi: 10.1016/j.cub.2011.10.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Abou Tayoun AN, et al. The Drosophila SK Channel (dSK) Contributes to Photoreceptor Performance by Mediating Sensitivity Control at the First Visual Network. J Neurosci. 2011;31:13897–13910. doi: 10.1523/JNEUROSCI.3134-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Juusola M, Hardie RC. Light adaptation in Drosphila photoreceptors: II. Rising temperature increases the bandwidth of reliable signaling. J Gen Physiol. 2001;117:27–41. doi: 10.1085/jgp.117.1.27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Juusola M, Hardie RC. Light adaptation in Drosophila photoreceptors: I. Response dynamics and signaling efficiency at 25 degrees. C. J Gen Physiol. 2001;117:3–25. doi: 10.1085/jgp.117.1.3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pantazis A, et al. Distinct roles for two histamine receptors (hclA and hclB) at the Drosophila photoreceptor synapse. J Neurosci. 2008;28:7250–7259. doi: 10.1523/JNEUROSCI.1654-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Song Z, Juusola M. Refractory sampling links efficiency and costs of sensory encoding to stimulus statistics. J Neurosci. 2014;34:7216–7237. doi: 10.1523/JNEUROSCI.4463-13.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Song Z, et al. Stochastic, Adaptive Sampling of Information by Microvilli in Fly Photoreceptors. Curr Biol. 2012;22:1371–1380. doi: 10.1016/j.cub.2012.05.047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zheng L, et al. Feedback network controls photoreceptor output at the layer of first visual synapses in Drosophila. J Gen Physiol. 2006;127:495–510. doi: 10.1085/jgp.200509470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zheng L, et al. Network Adaptation Improves Temporal Representation of Naturalistic Stimuli in Drosophila Eye I Dynamics. Plos One. 2009;4 doi: 10.1371/journal.pone.0004307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hardie RC, Juusola M. Phototransduction in Drosophila. Curr opin neurobiol. 2015;34:37–45. doi: 10.1016/j.conb.2015.01.008. [DOI] [PubMed] [Google Scholar]
- Clandinin TR, Zipursky SL. Making connections in the fly visual system. Neuron. 2002;35:827–841. doi: 10.1016/s0896-6273(02)00876-0. [DOI] [PubMed] [Google Scholar]
- Wernet MF, et al. Homothorax switches function of Drosophila photoreceptors from color to polarized light sensors. Cell. 2003;115:267–279. doi: 10.1016/s0092-8674(03)00848-1. [DOI] [PubMed] [Google Scholar]
- Wardill TJ, et al. Multiple Spectral Inputs Improve Motion Discrimination in the Drosophila Visual System. Science. 2012;336:925–931. doi: 10.1126/science.1215317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hardie RC. In: Progress in Sensory Physiology. Ottoson D, editor. Vol. 5. Springer; 1985. pp. 1–79. [Google Scholar]
- Borst A. Drosophila's View on Insect Vision. Curr Biol. 2009;19:36–47. doi: 10.1016/j.cub.2008.11.001. [DOI] [PubMed] [Google Scholar]
- Kirschfeld K. Die Projektion Der Optischen Umwelt Auf Das Raster Der Rhabdomere Im Komplexauge Von Musca. Exp Brain Res. 1967;3:248–270. doi: 10.1007/BF00235588. [DOI] [PubMed] [Google Scholar]
- Morante J, Desplan C. Photoreceptor axons play hide and seek. Nat Neurosci. 2005;8:401–402. doi: 10.1038/nn0405-401. [DOI] [PubMed] [Google Scholar]
- Fischbach KF, Hiesinger PR. Optic lobe development. Adv Exp Med Biol. 2008;628:115–136. doi: 10.1007/978-0-387-78261-4_8. [DOI] [PubMed] [Google Scholar]
- Shaw SR. Early Visual Processing in Insects. J Exp Biol. 1984;112:225–251. doi: 10.1242/jeb.112.1.225. [DOI] [PubMed] [Google Scholar]
- Joesch M, Schnell B, Raghu SV, Reiff DF, Borst A. ON and OFF pathways in Drosophila motion vision. Nature. 2010;468:300–304. doi: 10.1038/nature09545. [DOI] [PubMed] [Google Scholar]
- Clark DA, Bursztyn L, Horowitz MA, Schnitzer MJ, Clandinin TR. Defining the Computational Structure of the Motion Detector in Drosophila. Neuron. 2011;70:1165–1177. doi: 10.1016/j.neuron.2011.05.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rister J, et al. Dissection of the peripheral motion channel in the visual system of Drosophila melanogaster. Neuron. 2007;56:155–170. doi: 10.1016/j.neuron.2007.09.014. [DOI] [PubMed] [Google Scholar]
- Vogt N, Desplan C. The first steps in Drosophila motion detection. Neuron. 2007;56:5–7. doi: 10.1016/j.neuron.2007.09.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strausfeld NJ, Braitenberg V. Compound Eye of Fly (Musca-Domestica) - Connections between Cartridges of Lamina Ganglionaris. Z Vergl Physiol. 1970;70:95–104. [Google Scholar]
- Strausfeld NJ, Campos-Ortega J. L4-Monopolar Neuron - Substrate for Lateral Interaction in Visual System of Fly Musca-Domestica (L) Brain Res. 1973;59:97–117. doi: 10.1016/0006-8993(73)90254-0. [DOI] [PubMed] [Google Scholar]
- Takemura SY, et al. Cholinergic Circuits Integrate Neighboring Visual Signals in a Drosophila Motion Detection Pathway. Curr Biol. 2011;21:2077–2084. doi: 10.1016/j.cub.2011.10.053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shaw SR, Frohlich A, Meinertzhagen IA. Direct Connections between the R7/8 and R1-6 Photoreceptor Subsystems in the Dipteran Visual-System. Cell Tissue Res. 1989;257:295–302. doi: 10.1007/BF00261833. [DOI] [PubMed] [Google Scholar]
- Wolfram V, Juusola M. Impact of rearing conditions and short-term light exposure on signaling performance in Drosophila photoreceptors. J Neurophysiol. 2004;92:1918–1927. doi: 10.1152/jn.00201.2004. [DOI] [PubMed] [Google Scholar]
- Nikolaev A, et al. Network Adaptation Improves Temporal Representation of Naturalistic Stimuli in Drosophila Eye II Mechanisms. Plos One. 2009;4 doi: 10.1371/journal.pone.0004306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Land MF. Compound eye structure: Matching eye to environment. Adaptive Mechanisms in the Ecology of Vision. Netherlands: Springer; 1999. [Google Scholar]
- Shannon CE. A Mathematical Theory of Communication. At&T Tech J. 1948;27:379–423. [Google Scholar]
- Niven JE, et al. The contribution of Shaker K+ channels to the information capacity of Drosophila photoreceptors. Nature. 2003;421:630–634. doi: 10.1038/nature01384. [DOI] [PubMed] [Google Scholar]
- Randall AS, et al. Speed and sensitivity of phototransduction in Drosophila depend on degree of saturation of membrane phospholipids. J Neurosci. 2015;35:2731–2746. doi: 10.1523/JNEUROSCI.1150-14.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Faivre O, Juusola M. Visual Coding in Locust Photoreceptors. Plos One. 2008;3:e2173. doi: 10.1371/journal.pone.0002173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gonzalez-Bellido PT, Wardill TJ, Juusola M. Compound eyes and retinal information processing in miniature dipteran species match their specific ecological demands. P Natl Acad Sci USA. 2011;108:4224–4229. doi: 10.1073/pnas.1014438108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Juusola M, Kouvalainen E, Järvilehto M, Weckström M. Contrast Gain, Signal-to-Noise Ratio, and Linearity in Light-Adapted Blowfly Photoreceptors. J Gen Physiol. 1994;104:593–621. doi: 10.1085/jgp.104.3.593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Juusola M, Uusitalo RO, Weckstrom M. Transfer of Graded Potentials at the Photoreceptor Interneuron Synapse. J Gen Physiol. 1995;105:117–148. doi: 10.1085/jgp.105.1.117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Juusola M, de Polavieja GG. The rate of information transfer of naturalistic stimulation by graded potentials. J Gen Physiol. 2003;122:191–206. doi: 10.1085/jgp.200308824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weckstrom M, Kouvalainen E, Juusola M. Measurement of Cell Impedance in Frequency-Domain Using Discontinuous Current Clamp and White-Noise-Modulated Current Injection. Pflug Arch Eur J Phy. 1992;421:469–472. doi: 10.1007/BF00370258. [DOI] [PubMed] [Google Scholar]
- Juusola M. Measuring Complex Admittance and Receptor Current by Single Electrode Voltage-Clamp. J Neurosci Meth. 1994;53:1–6. doi: 10.1016/0165-0270(94)90137-6. [DOI] [PubMed] [Google Scholar]
- Juusola M, Seyfarth EA, French AS. Rapid coating of glass-capillary microelectrodes for single-electrode voltage-clamp. J Neurosci Meth. 1997;71:199–204. doi: 10.1016/s0165-0270(96)00145-8. [DOI] [PubMed] [Google Scholar]
- Haag J, Borst A. Neural mechanism underlying complex receptive field properties of motion-sensitive interneurons. Nat Neurosci. 2004;7:628–634. doi: 10.1038/nn1245. [DOI] [PubMed] [Google Scholar]
- Rien D, Kern R, Kurtz R. Synaptic transmission of graded membrane potential changes and spikes between identified visual interneurons. Eur J Neurosci. 2011;34:705–716. doi: 10.1111/j.1460-9568.2011.07801.x. [DOI] [PubMed] [Google Scholar]
- Muller A, et al. Switched single-electrode voltage-clamp amplifiers allow precise measurement of gap junction conductance. Am J Physiol-Cell Ph. 1999;276:C980–C987. doi: 10.1152/ajpcell.1999.276.4.C980. [DOI] [PubMed] [Google Scholar]
- Polder HR, Swandulla D, Konnerth A, Lux HD. An Improved, High-Current Single Electrode Current Voltage Clamp System. Pflug Arch Eur J Phy. 1984;402:R35–R35. [Google Scholar]
- Richter DW, Pierrefiche O, Lalley PM, Polder HR. Voltage-clamp analysis of neurons within deep layers of the brain. J Neurosci Meth. 1996;67:121–131. [PubMed] [Google Scholar]
- Juusola M, French AS. The efficiency of sensory information coding by mechanoreceptor neurons. Neuron. 1997;18:959–968. doi: 10.1016/s0896-6273(00)80335-9. [DOI] [PubMed] [Google Scholar]
- Vähäsöyrinki M, Niven JE, Hardie RC, Weckström M, Juusola M. Robustness of neural coding in Drosophila photoreceptors in the absence of slow delayed rectifier K+ channels. J Neurosci. 2006;26:2652–2660. doi: 10.1523/JNEUROSCI.3316-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Friederich U, Coca D, Billings S, Juusola M. Data Modelling for Analysis of Adaptive Changes in Fly Photoreceptors. Lect Notes Comput Sc. 2009;5863:34–48. [Google Scholar]
- Asyali MH, Juusola M. Use of Meixner functions in estimation of Volterra kernels of nonlinear systems with delay. Ieee T Bio-Med Eng. 2005;52:229–237. doi: 10.1109/TBME.2004.840187. [DOI] [PubMed] [Google Scholar]
- French AS, et al. The Dynamic Nonlinear Behavior of Fly Photoreceptors Evoked by a Wide-Range of Light Intensities. Biophys J. 1993;65:832–839. doi: 10.1016/S0006-3495(93)81116-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Juusola M, French AS. Visual acuity for moving objects in first- and second-order neurons of the fly compound eye. J Neurophysiol. 1997;77:1487–1495. doi: 10.1152/jn.1997.77.3.1487. [DOI] [PubMed] [Google Scholar]
- Juusola M, Weckstrom M, Uusitalo RO, Korenberg MJ, French AS. Nonlinear models of the first synapse in the light-adapted fly retina. J Neurophysiol. 1995;74:2538–2547. doi: 10.1152/jn.1995.74.6.2538. [DOI] [PubMed] [Google Scholar]