

Abstract
Thymus involution, associated with aging or pathological insults, results in diminished output of mature T-cells. Restoring the function of a failing thymus is crucial to maintain effective T cell-mediated acquired immune response against invading pathogens. However, thymus regeneration and revitalization proved to be challenging, largely due to the difficulties of reproducing the unique 3D microenvironment of the thymic stroma that is critical for the survival and function of thymic epithelial cells (TECs). We developed a novel hydrogel system to promote the formation of TEC aggregates, based on the self-assembling property of the amphiphilic EAK16-II oligopeptides and its histidinylated analogue EAKIIH6. TECs were enriched from isolated thymic cells with density-gradient, sorted with fluorescence-activated cell sorting (FACS), and labeled with anti-epithelial cell adhesion molecule (EpCAM) antibodies that were anchored, together with anti-His IgGs, on the protein A/G adaptor complexes. Formation of cell aggregates was promoted by incubating TECs with EAKIIH6 and EAK16-II oligopeptides, and then by increasing the ionic concentration of the medium to initiate gelation. TEC aggregates embedded in EAK hydrogel can effectively promote the development of functional T cells in vivo when transplanted into the athymic nude mice.
Keywords: Immunology, Issue 112, Thymic epithelial cells, hydrogel, density gradient, self-assembly, EAK16, thymus reconstruction
Introduction
The thymus is the primary lymphoid organ responsible for the generation of a diverse population of pathogen-reactive, self-tolerant T cells that is essential to the function of the acquired immune system. It is a dynamic organ where developing thymocytes, immigrated from the bone marrow as lymphocyte progenitor cells, migrate through the sponge-like three-dimensional (3-D) matrix of the thymic stroma, undergo lineage-specification and differentiation, and eventually emigrate as mature T-cells. The success of this well programmed process depends largely on the cross-talk between the migrating thymocytes and the residential thymic epithelial cells (TECs), the predominant population of the thymic stroma that are essential for establishing and maintaining the integrity of the thymus microenvironment.
Based on their anatomical location and unique function, TECs can be divided into two subsets: the TECs in the cortex (cTECs) that are responsible for selecting self-MHC (major histocompatibility complex) restricted T-cells (positive selection), and the TECs in the medulla (mTECs) that are essential for eliminating autoreactive T-cells (negative selection) 1,2. Many factors (e.g., aging, infection, irradiation, drug treatments) can cause irreversible damages to the thymic epithelium, resulting in compromised adaptive immunity. Despite numerous attempts, restoring the thymic function has been challenging due to the difficulty to reproduce the thymic microenvironment. Notably, thymic three-dimensional (3-D) configuration is critical to the survival and function of TECs, whereas TECs cultured in a 2-D environment rapidly decrease the expression of genes critical for thymopoiesis 3,4.
EAK16-II (AEAEKAKAEAEAKAK) and its C-terminal histidinylated analogue EAKIIH6 (AEAEKAKAEAEAKAKHHHHHH) are low-molecular weight, amphiphilic oligopeptides that are soluble in deionized water, but undergo gelation to form β-fibrils when exposed to ionic strength higher than 20 mM NaCl (normal salt concentration in human body fluid is 154 mM). This environmental responsive property makes them versatile building blocks to form 3D structures. The His-tag on EAKII-H6 provides a docking mechanism, by which the anti-His IgGs/Fc-binding recombinant protein A/G (αH6:pA/G) complexes can serve as an adaptor to anchor protein drugs and other biomolecules (e.g., antibodies) on the hydrogel composite 5-8.
We have previously demonstrated that fluorescent-labeled IgG molecules anchored on the hydrogel can be retained at the injection site for up to 13 days 9. Furthermore, when the αH6:pA/G adaptors anchored with anti-CD4 IgGs were added to the EAK16-II/EAKIIH6 (EAK) hydrogel, CD4+ T cells were specifically captured 10. Using similar technique, we have recently demonstrated that 3-D aggregation of TECs could be promoted in EAK hydrogel with adaptor complexes carrying TEC-specific anti-EpCAM antibodies (αH6:pA/G:αEpCAM). When transplanted underneath the kidney capsules of nude mice, the TEC clusters embedded in EAK hydrogel can effectively support the development of functional T cells in vivo11,12.
Here we illustrate our method to quickly and effectively purify TECs with fluorescence-activated cell sorting (FACS) and generate 3-D TEC aggregates with the EAK hydrogel system.
Protocol
All the animals used in the experiments were housed in the animal facility at Allegheny-Singer Research Institute under the protocol reviewed and approved by the Institutional Animal Care and Use Committee of the Allegheny Health Network/Allegheny Singer Research Institute.
1. Digesting the Thymus with Collagenase
- Harvest thymus glands.
- Euthanize 3-5 week-old C57BL6/J mice in a carbon dioxide (CO2) chamber to harvest thymus glands. In detail, place the mice in the chamber under CO2 induction for 3-5 min and confirm death by verifying cardiac or respiratory arrest.
- Lay the carcass on its back and wet the body with 70% ethanol. Make a small horizontal incision with a sharp, straight dissecting scissors on its abdomen through the skin. Pull the skin on both ends (head and legs) so that it is turned inside out.
- Make a horizontal cut with the dissecting scissors just under the lowest rib and cut through the diaphragm sideways. Hold the xiphoid process with a forceps and cut the middle of the ribs proceeding up on both sides. Pull up the ribs/sternum so that the thymus is clearly visible. Note: Thymus lies directly on top of the heart, consists of 2 lobes and is of a white translucent color.
- Using a curved forceps, gently scoop and pull the thymus lobes off of the connective tissue and transfer in washing solution (see List of Materials).
Prepare digestion solution by mixing 9 ml RPMI-1640 (Roswell Park Memorial Institute medium-1640), 0.025 mg/ml purified collagenase, 10 mM HEPES [4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid], and 0.2 mg/ml DNase I in a 50 ml centrifuge tube. Keep the digestion solution in 37 °C water bath until use. Note: The amount of the digestion solution prepared is sufficient for up to five adult mouse thymi, which can be incubated together in one 5 ml polystyrene round-bottom tube.
Transfer the thymus to a 60 mm tissue culture dish containing 5-6 ml RPMI-1640.
Dissect the thymus with 28 G insulin syringe needle into small pieces (about 1 mm3 square) to let the lymphocytes flow out.
Rinse the thymic pieces with RPMI-1640 that is in the petri dish with glass Pasteur pipette. Tilt the dish and let the thymic fragments settle to the bottom. Slowly aspirate and discard the RPMI-1640 supernatant using a glass pipette. Repeat this rinsing step with 5-6 ml RPMI-1640 for 4-5 times with glass pipette until the solution is less opaque. Note: Glass pipettes are recommended at this step, since thymic fragments tend to stick to plastic, resulting in excessive loss of thymic tissues. For the rest of the procedure, use plastic pipettes.
After the final rinse, discard the supernatant as described in step 1.5. Add 3 ml digestion solution and transfer the thymus pieces and the solution into a 5 ml round-bottom polystyrene tube.
Place the tube on a tube rotator and incubate at 37 °C for 6 min at a rotation speed of 12 rpm.
After incubation, let the thymic fragments settle to the bottom of the tube by gravity. Collect the supernatant with Pasteur pipette and transfer into a 50 ml centrifuge tube with 10 ml of washing solution. Centrifuge at 300 x g for 5 min, discard the supernatant and resuspend the cells in 10 ml washing solution. Keep on ice.
Repeat steps 1.6-1.8 twice on the thymic fragments in the 5 ml round-bottom polystyrene tube. Pool the supernatant in the same 50 ml tube. Note: Centrifugation is not necessary for the second and the third wash.
After the final digestion, pipet up and down with 2 ml plastic serological pipette to break down any remaining fragments in the tube and transfer to the 50 ml centrifuge tube.
Centrifuge the 50 ml tube at 300 x g for 5 min, aspirate the supernatant, resuspend in 10 ml of washing buffer and filter through 100 µm cell strainer. Keep on ice.
2. Enrichment of TECs with Discontinuous Density Gradient
Prepare 21% density gradient solution by mixing 2.4 ml density gradient medium (60% w/v of iodixanol in water) and 9.6 ml washing solution.
Centrifuge the dissociated thymic cells in the 50 ml collection tube from step 1.11 at 300 x g for 5 min and resuspend in 2.5 ml washing buffer.
Add 20 ml RPMI-1640 medium to the cell suspension.
- Fill 12 ml of 21% density gradient solution in a 10 ml serological pipet with an electronic pipettor. Place the tip of the serological pipet at the bottom of the 50 ml centrifuge tube containing cells, and allow the density gradient solution to drain to the bottom of the tube via gravity.
- Gently expel the density gradient solution from the pipet to finish generating the two layers of different densities.
Centrifuge at 600 x g for 20 min at RT in a swinging-bucket rotor. Set the deceleration rate at the slowest level.
Transfer the top layer and the interface to a new 50 ml tube. Note: Most of the TECs are retained in the interface. Lymphocytes will precipitate to the bottom of the tube after centrifugation. If these cells are to be used for other purposes, rinse the pellet with 20 ml washing solution for three times. Resuspend the cells in 10 ml of washing buffer.
Add washing solution up to 40 ml to the tube in step 2.6 and centrifuge at 400 x g for 6 min at 4 °C. To remove the supernatant, aspirate with a pipet.
Repeat the washing and the aspiration 2 more times as described in step 2.7.
Resuspend in 2 ml of washing solution and count the cells with a hemocytometer.
3. Isolating TECs with FACS
Adjust cells from step 2.9 at the concentration of 1x106 cells/100 µl with washing solution and transfer the cells to a 5 ml round-bottom tube (polypropylene or polystyrene, as recommended by the instructions of the flow sorter).
Add 2 µl anti-Fc receptor IgG per 1x106 cells and incubate at 4 °C for 10 min.
Add anti-CD45 (1:150 dilution, 10 µl per sample) and anti-EpCAM (1:100 dilution, 10 µl per sample) antibodies and incubate at 4 °C for more than 20 min.
Wash the cells with 2 ml washing solution and centrifuge at 400 x g for 6 min at 4 °C. Aspirate the supernatant with a pipette.
Resuspend the cells in 1 ml 1x phosphate-buffered saline (PBS).
Filter through 100 µm strainer.
Apply the cells to the sorting instrument. Select the lymphocyte and TEC population excluding the smallest cells on side-scattered light (SSC) / forward-scattered light (FSC) panel (dead cells), then gate on the selected population for CD45- EpCAM+ for sorting13.
4. Generation of TEC/EAK Aggregates
Note: Perform reagent preparation and steps involving cell mixing under the laminar hood.
Prepare pA/G adaptor complexes by mixing 4 µl of anti-His-tag IgG, 8 µl of anti-EpCAM IgG and 2.6 µl of pA/G in a sterile 1.5 ml microcentrifuge tube, and incubate at RT for 5 min.
Adjust the sorted TECs from step 3.7 to 5x104 cells/ml with washing solution, and pipet 100 µl to one well of a low evaporative, 96-well round-bottom tissue culture plate. Add 14.6 µl pA/G adaptor complexes to the cells. Place the plate on a rocking platform for 20 min at 4 °C.
Wash once with 200 µl washing solution. Centrifuge the plate at 400 x g for 6 min. Discard the supernatant with a micropipette.
Wash once with 200 µl 10% sucrose solution. Centrifuge at 400 x g for 6 min. Discard the supernatant with a micropipette.
Resuspend cells in 30 µl 10% sterile sucrose solution per well.
Add 10 µl of EAKIIH6 (7.5 mg/ml) and incubate for 5 min at RT.
Add 20 µl of EAK16-II (10 mg/ml) to the TEC mixture.
Add 100 µl of complete medium to the system to initiate gelation. Place the plate on a platform shaker for 15-20 min at RT.
Place the plate in the incubator at 37 °C and 5% CO2. After 2-4 hr, add another 100 µl of complete medium.
- Change medium every other day until use.
- Aspirate 100 µl of the medium from the surface without disturbing the gel using a micropipette and gently add 100 µl of pre-warmed medium by placing the pipette tip to the wall of the well.
5. Characterization of the TEC/EAK Aggregates
To examine the function of the TEC/EAK aggregates in vivo, collect the TEC/EAK hydrogel after O/N culture and transplant under the kidney capsule of an athymic nude mouse, using previously described procedure 11,14.
Monitor the development of T cells in the periphery by harvesting 50-100 µl blood samples into the EDTA-containing blood collection tube from the nude mice 4-6 weeks post transplantation and stain with a cocktail of anti-CD4, -CD8, -CD45, and -CD3 antibodies12.
Analyze the percentages of T-lymphocytes in the peripheral blood leukocytes with flow cytometry (FCM), using single cell analysis software12.
Representative Results
To examine the effectiveness of using the density gradient separation protocol to enrich the CD45- stromal cells, cells harvested from both the interface and the precipitated lymphocyte pellets were stained with anti-CD45 and anti-EpCAM antibodies. Both anti-Ulex Europaeus Agglutinin 1 (UEA1) and anti-MHC Class II antibodies were also included in the staining cocktail to further identify the cTEC and mTEC subsets. As shown in Figure 1, we were able to routinely achieve close to 15-20 times enrichment of the TECs.
To examine the aggregation of the TECs in the EAK hydrogel, TECs were isolated from 4-week old B6.ROSAmT/mG mice, which ubiquitously express the membrane-bound, red fluorescent tdTomato molecules. TEC aggregates, prepared as described above, were cultured for two days in vitro and examined under a confocal microscope (Figure 2).
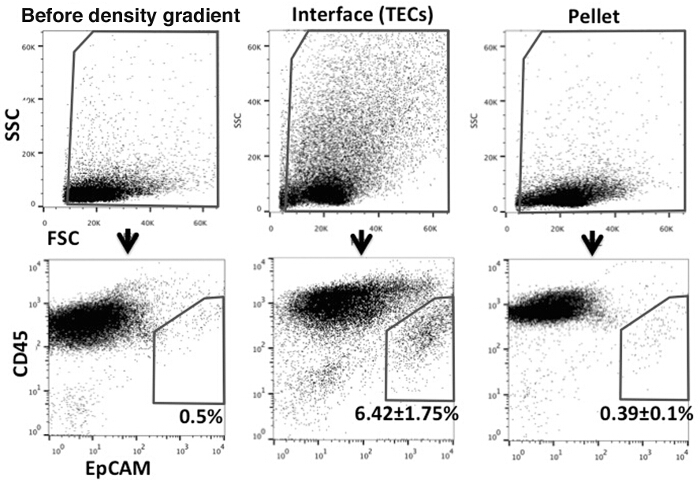 Figure 1.Flow cytometric analysis of TECs enriched with the density gradient solution. Cells were stained with anti-CD45 and anti-EpCAM antibodies to distinguish TEC population (CD45- EpCAM+). Left panels: Thymic cells after enzyme digestion, before the density gradient procedure. Middle panels: Cells in the top layer and the interface after separation with the density gradient solution. Right panels: Pelleted cells (lymphocyte pellet) after separation with the density gradient solution. Please click here to view a larger version of this figure.
Figure 1.Flow cytometric analysis of TECs enriched with the density gradient solution. Cells were stained with anti-CD45 and anti-EpCAM antibodies to distinguish TEC population (CD45- EpCAM+). Left panels: Thymic cells after enzyme digestion, before the density gradient procedure. Middle panels: Cells in the top layer and the interface after separation with the density gradient solution. Right panels: Pelleted cells (lymphocyte pellet) after separation with the density gradient solution. Please click here to view a larger version of this figure.
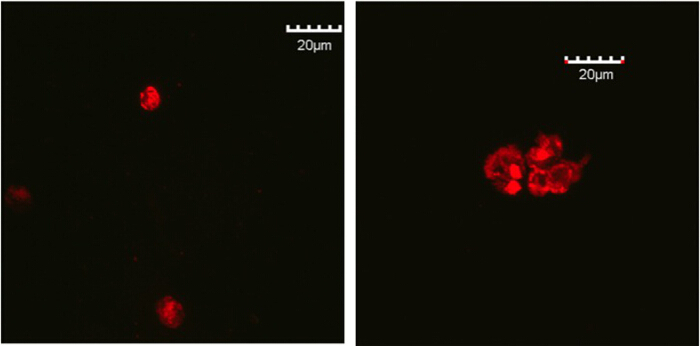 Figure 2.Confocal microscopic images of TECs clusters in EAK hydrogel. Red fluorescent TECs harvested from the B6.ROSAmT/mG mice were isolated with FACS. TECs clusters were embedded in EAK hydrogel, cultured in 96-well plate and were harvested at day 2. Left panel, TECs were labeled with IgG antibody; Right panel, TECs were labeled with anti-EpCAM antibody. Please click here to view a larger version of this figure.
Figure 2.Confocal microscopic images of TECs clusters in EAK hydrogel. Red fluorescent TECs harvested from the B6.ROSAmT/mG mice were isolated with FACS. TECs clusters were embedded in EAK hydrogel, cultured in 96-well plate and were harvested at day 2. Left panel, TECs were labeled with IgG antibody; Right panel, TECs were labeled with anti-EpCAM antibody. Please click here to view a larger version of this figure.
Discussion
While TECs are the predominant population of the thymic stroma and play essential roles for the structure and function of the thymus glands, they represent only about 0.1-0.5% of the total thymic cellularity. They are also fragile cells as high percentages of cell death are occasionally observed following collagenase digestion, i.e., the treatment to dissociate TECs from the extracellular matrix (ECM). Their rarity (~200,000 per mouse thymus) and fragility made it a challenging task to isolate TECs. The recent utilization of highly purified collagenase in TEC isolation have significantly increased the numbers of live cells following the enzymatic digestion 15-19.
However, the very small number of TECs remains a major challenge for their analysis and isolation, as large quantities of antibodies are needed for cell surface labeling and excessive numbers of events need to be collected by FACS. While antibody-conjugated magnetic bead technology can be used to deplete CD45+ thymocytes, their large quantity renders it economically prohibitive in the long run. We were able to effectively enrich TECs 15-20 times with a simple, one step density gradient centrifugation. The iso-osmotic property of the iodixanol density gradient medium allows the separation of cells based on their shapes, sizes and densities 20-22. It should be noted that the purity and yield of TECs are affected by the percentage of the density gradient medium used. Lower percentage of the density gradient medium (18-20% v/v) will result in higher purity of TECs in the interface but lower yield. In contrast, when higher iodixanol concentration (22% v/v) is used, larger quantity of thymic cells can be collected with lower percentage of TECs. The concentration of iodixanol should be adjusted based on the purpose of the experiment. While the purity of TECs is largely improved with the utility of density gradient solution, it should be emphasized that the cells collected from the top layer and the interface still contain a relatively high fraction of CD45+ cells (Figure 1). Therefore, when only the TEC population is required as in this experiment, cell sorting will be crucial, although this extra procedure may cause some loss of cells and requires more time.
Unlike most of the epithelial cells arranged in a single or multiple layers lining the visceral organs, TECs are organized in 3-D configuration in the thymus stroma, which is essential for their survival and function. The self-assembling EAK hydrogel system facilitates the formation of 3-D TEC aggregates that are mediated by the tri-molecular αH6:pA/G:αEpCAM adaptor complexes. Of note, by fine-tuning the concentrations and ratios of the EAK16-II/EAKIIH6 oligopeptides, the porosity and density of the hydrogel can be adjusted to allow the flow of nutrients and the migration of thymocytes. Indeed, functional T-cells were generated in athymic nude mice transplanted with the TEC/EAK composites.
The drawback in this EAK hydrogel system is that with this current protocol, specific cell composition cannot be localized to a designated area. In a physical environment a thymus is comprised of two distinct compartments, in which specific types of TECs reside, cTECs in the cortex and mTECs in medulla. With our system described here, both cTECs and mTECs are labeled with EpCAM, a general epithelial marker. Therefore it is speculated that the aggregates in the formed hydrogel includes both subtypes of TECs. Although cTECs and mTECs derive from common bipotent TEC progenitors, their roles in the development of T cells are clearly distinctive. Cortex is the first region in the thymus where the immature thymic progenitor cells enter, and the positive selection takes place 23. Then the selected cells move into the medullary region and interact with mTECs, through which the self-reactive cells are eliminated by negative selection 24. Moreover, it has been demonstrated that 25% of the genes are differentially expressed between cTECs and mTECs 25. These findings underline the importance of promoting aggregation of TEC subsets to increase the efficacy of T cell development. One possible approach is to use different antibodies targeting surface molecules specific to either cTECs or mTECs, such as anti-cytokeratin 8 for cTECs, and anti-cytokeratin 5 for mTECs.
From the perspective of future clinical application, the TEC clusters embedded in the biodegradable hydrogel might function as injectable "mini thymus units" to modulate the adaptive immune system that could be important for various medical conditions, such as rejuvenating the adaptive immune system in aged individuals, or inducing donor-specific tolerance in solid organ transplantation.
Disclosures
Authors declare no conflict of interest.
Acknowledgments
This work was supported in part by the National Institutes of Health grants R21 AI113000 (W.S.M) and R01 AI123392 (Y.F.).
References
- Anderson G, Jenkinson EJ, Moore NC, Owen JJ. MHC class II-positive epithelium and mesenchyme cells are both required for T-cell development in the thymus. Nature. 1993;362:70–73. doi: 10.1038/362070a0. [DOI] [PubMed] [Google Scholar]
- Fan Y, et al. Thymus-specific deletion of insulin induces autoimmune diabetes. The EMBO J. 2009;28:2812–2824. doi: 10.1038/emboj.2009.212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mohtashami M, Zuniga-Pflucker JC. Three-dimensional architecture of the thymus is required to maintain delta-like expression necessary for inducing T cell development. J Immunol. 2006;176:730–734. doi: 10.4049/jimmunol.176.2.730. [DOI] [PubMed] [Google Scholar]
- Pinto S, et al. An organotypic coculture model supporting proliferation and differentiation of medullary thymic epithelial cells and promiscuous gene expression. J Immunol. 2013;190:1085–1093. doi: 10.4049/jimmunol.1201843. [DOI] [PubMed] [Google Scholar]
- Fung SY, Yang H, Chen P. Formation of colloidal suspension of hydrophobic compounds with an amphiphilic self-assembling peptide. Colloids Surf B Biointerfaces. 2007;55:200–211. doi: 10.1016/j.colsurfb.2006.12.002. [DOI] [PubMed] [Google Scholar]
- Jun S, et al. Self-assembly of the ionic peptide EAK16: the effect of charge distributions on self-assembly. Biophys J. 2004;87:1249–1259. doi: 10.1529/biophysj.103.038166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saunders MJ, et al. Engineering fluorogen activating proteins into self-assembling materials. Bioconjug Chem. 2013;24:803–810. doi: 10.1021/bc300613h. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang S, Holmes T, Lockshin C, Rich A. Spontaneous assembly of a self-complementary oligopeptide to form a stable macroscopic membrane. Proc of the Natl Acad Sci U S A. 1993;90:3334–3338. doi: 10.1073/pnas.90.8.3334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wen Y, et al. Coassembly of amphiphilic peptide EAK16-II with histidinylated analogues and implications for functionalization of beta-sheet fibrils in vivo. Biomaterials. 2014;35:5196–5205. doi: 10.1016/j.biomaterials.2014.03.009. [DOI] [PubMed] [Google Scholar]
- Zheng Y, et al. A peptide-based material platform for displaying antibodies to engage T cells. Biomaterials. 2011;32:249–257. doi: 10.1016/j.biomaterials.2010.08.083. [DOI] [PubMed] [Google Scholar]
- Tajima A, et al. Bioengineering mini functional thymic units with EAK16-II/EAKIIH6 self-assembling hydrogel. Clin Immunol. 2015;160:82–89. doi: 10.1016/j.clim.2015.03.010. [DOI] [PubMed] [Google Scholar]
- Fan Y, et al. Bioengineering Thymus Organoids to Restore Thymic Function and Induce Donor-Specific Immune Tolerance to Allografts. Mol Ther. 2015;23:1262–1277. doi: 10.1038/mt.2015.77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fan Y, et al. Thymus-specific deletion of insulin induces autoimmune diabetes. The EMBO J. 2009;28:2812–2824. doi: 10.1038/emboj.2009.212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fan Y, et al. Compromised central tolerance of ICA69 induces multiple organ autoimmunity. J Autoimmun. 2014;53:10–25. doi: 10.1016/j.jaut.2014.07.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Palumbo MO, Levi D, Chentoufi AA, Polychronakos C. Isolation and characterization of proinsulin-producing medullary thymic epithelial cell clones. Diabetes. 2006;55:2595–2601. doi: 10.2337/db05-1651. [DOI] [PubMed] [Google Scholar]
- Williams KM, et al. Single cell analysis of complex thymus stromal cell populations: rapid thymic epithelia preparation characterizes radiation injury. Clin Transl Sci. 2009;2:279–285. doi: 10.1111/j.1752-8062.2009.00128.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McLelland BT, Gravano D, Castillo J, Montoy S, Manilay JO. Enhanced isolation of adult thymic epithelial cell subsets for multiparameter flow cytometry and gene expression analysis. J Immunol Methods. 2011;367:85–94. doi: 10.1016/j.jim.2011.02.008. [DOI] [PubMed] [Google Scholar]
- Seach N, Wong K, Hammett M, Boyd RL, Chidgey AP. Purified enzymes improve isolation and characterization of the adult thymic epithelium. J Immunol methods. 2012;385:23–34. doi: 10.1016/j.jim.2012.07.023. [DOI] [PubMed] [Google Scholar]
- Xing Y, Hogquist KA. Isolation, identification, and purification of murine thymic epithelial cells. J Vis Exp. 2014. p. e51780. [DOI] [PMC free article] [PubMed]
- Graham JM. Separation of monocytes from whole human blood. ScientificWorldJournal. 2002;2:1540–1543. doi: 10.1100/tsw.2002.842. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li X, Donowitz M. Fractionation of subcellular membrane vesicles of epithelial and nonepithelial cells by OptiPrep density gradient ultracentrifugation. Methods Mol Biol. 2008;440:97–110. doi: 10.1007/978-1-59745-178-9_8. [DOI] [PubMed] [Google Scholar]
- Mita A, et al. Purification method using iodixanol (OptiPrep)-based density gradient significantly reduces cytokine chemokine production from human islet preparations, leading to prolonged beta-cell survival during pretransplantation culture. Transplant Proc. 2009;41:314–315. doi: 10.1016/j.transproceed.2008.10.059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson G, Takahama Y. Thymic epithelial cells: working class heroes for T cell development and repertoire selection. Trends Immunol. 2012;33:256–263. doi: 10.1016/j.it.2012.03.005. [DOI] [PubMed] [Google Scholar]
- Kyewski B, Klein L. A central role for central tolerance. Annu Rev Immunol. 2006;24:571–606. doi: 10.1146/annurev.immunol.23.021704.115601. [DOI] [PubMed] [Google Scholar]
- St-Pierre C, et al. Transcriptome sequencing of neonatal thymic epithelial cells. Sci Rep. 2013;3:1860. doi: 10.1038/srep01860. [DOI] [PMC free article] [PubMed] [Google Scholar]