

Abstract
Pseudomonas moraviensis is a predominant member of soil environments. We here report on the genomic analysis of Pseudomonas moraviensis strain Devor that was isolated from a gate at Oklahoma State University, Stillwater, OK, USA. The partial genome of Pseudomonas moraviensis strain Devor consists of 6016489 bp of DNA with 5290 protein-coding genes and 66 RNA genes. This is the first detailed analysis of a P. moraviensis genome. Genomic analysis revealed metabolic versatility with genes involved in the metabolism and transport of fructose, xylose, mannose and all amino acids with the exception of tryptophan and valine, implying that the organism is a versatile heterotroph. The genome of P. moraviensis strain Devor was rich in transporters and, based on COG analysis, did not cluster closely with P. moraviensis R28-S genome, the only previous report of a P. moraviensis genome with a native mercury resistance plasmid.
Keywords: Pseudomonas moraviensis, Draft genome sequence, Detailed analysis, Student Initiated Microbial Discovery (SIMD) project, Metabolic versatility
1. Introduction
The strain Devor was isolated from an outdoor gate located on Oklahoma State University (OSU) campus in Stillwater, OK. The isolation process was part of the Student Initiated Microbial Discovery (SIMD) project at OSU (introduced in [1]). The organism was isolated by an undergraduate student (RCD) during an introductory microbiology lab course, and analyzed by a team of undergraduate (MB and PB) and graduate students (NTM and DF). The genus Pseudomonas is known to be phylogenetically and physiologically diverse. Members are ubiquitously found in soil and water systems, with the potential to utilize a broad range of organic compounds [2], [3], [4], [5], [6], [7], [8], [9], [10], [11], [12]. Resistance for heavy metal has also been shown for some strains [13]. Genomic analysis of members belonging to the genus Pseudomonas can contribute to our understanding of the molecular mechanisms of the biodegradation of organic compounds, including environmental pollutants, and could potentially contribute to bioremediation efforts in multiple environments. Here we report on the draft genomic sequence, and the first detailed analysis of a Pseudomonas moraviensis strain.
2. Materials and methods
2.1. Genome project history
Pseudomonas moraviensis strain Devor was isolated with the aim of sequencing its genome as part of an undergraduate project at OSU. The project is funded by the Howard Hughes Medical Institute and aims at improving undergraduate student involvement in authentic research. Isolation of the strain, and analysis of the sequenced genome were performed by undergraduate students in an introductory microbiology course, and an upper division microbial genomics class, respectively. The quality draft assembly and annotation were completed in 2015–2016. Table 1 shows the genome project information.
Table 1.
Project information.
| MIGS ID | Property | Term |
|---|---|---|
| MIGS 31 | Finishing quality | Draft |
| MIGS-28 | Libraries used | 2 × 300 paired end chemistry |
| MIGS 29 | Sequencing platforms | Illumina |
| MIGS 31.2 | Fold coverage | 300 × |
| MIGS 30 | Assemblers | Velvet |
| MIGS 32 | Gene calling method | Prodigal |
| Genbank ID | MAYQ00000000 | |
| GenBank date of release | July 2016 | |
| GOLD ID | Gp0126756 | |
| BIOPROJECT | PRJNA327387 | |
| MIGS 13 | Project relevance | Environmental |
2.2. Growth conditions and genomic DNA preparation
Pseudomonas moraviensis strain Devor was grown overnight at 30 °C on tryptic soy agar plates, and genomic DNA was isolated using the MPBio PowerSoil® DNA extraction kit according to manufacturer's instructions. Negative stain TEM micrographs were obtained using the services of the Oklahoma State University Microscopy Lab. Briefly, the sample was placed on a carbon film TEM grid and allowed to incubate for 2 min, after which the excess liquid was wicked off. Phosphotungstic acid (PTA; 2% w/v) was then added to the grid followed by a 45-s incubation. Excess PTA was wicked off and the grid was allowed to dry before it was visualized using JOEL JEM-2100 transmission electron microscope.
2.3. Genome sequencing and assembly
The genome of Pseudomonas moraviensis strain Devor was sequenced at the University of Georgia Genomics Facility using the Illumina MiSeq platform 2 × 300 paired end chemistry. Average library insert size was 700 bp. The short read de Brujin graph assembly program Velvet [14] was used to assemble quality filtered sequence data using a kmer value of 101 bp and a minimum contig coverage value of 7 ×. The genome project is deposited in GOLD (Genomes On-Line Database) and this Whole Genome Shotgun (WGS) project has been deposited in GenBank under the accession MAYQ00000000. The version described in this paper is version MAYQ01000000.
2.4. Genome annotation
Gene models were created using the prokaryotic gene calling software package Prodigal [15]. A total of 5473 gene models were predicted with an average gene size of 1002 bp. Predicted protein sequences were annotated using a combination of NCBI Blast C ++ homology search, and HMMER 3.0 [16] hmmscan against the PFAM [17] 26.0 database. Additional functional annotations were carried out through the Integrated Microbial Genomes Expert Review (IMG-ER) platform.
2.5. Comparative genomics
We compared the genome of Pseudomonas moraviensis strain Devor to 13 closely related genomes (IMG genome IDs: 2619619019, 2654587541, 2603880217, 2561511156, 2639762619, 2636416187, 2639762638, 2639762633, 2597489942, 2639762506, 2517572232, 2639762618, 2556921015) using the “Genome clustering” function on the IMG-ER analysis platform based on the COG profile. We also used principal component analysis to compare the genomes based on several genomic features including the genome size, the number of genes, the number of transporters identified, the GC content, the number of non-coding bases, the number of genes belonging to COG categories, as well as the number of genes belonging to each COG category. The PCA analysis was conducted using the “princomp” function in the labdsv library of R [18]. The results were visualized using a biplot, where genomes were represented by stars and genomic features or COG categories used for comparison were represented by arrows.
3. Results and discussion
3.1. Classification and features
Cells of P. moraviensis strain Devor are Gram negative, motile, aerobic rods that are arranged as singles (Fig. 1). Colonies on TSA agar were orange.
Fig. 1.
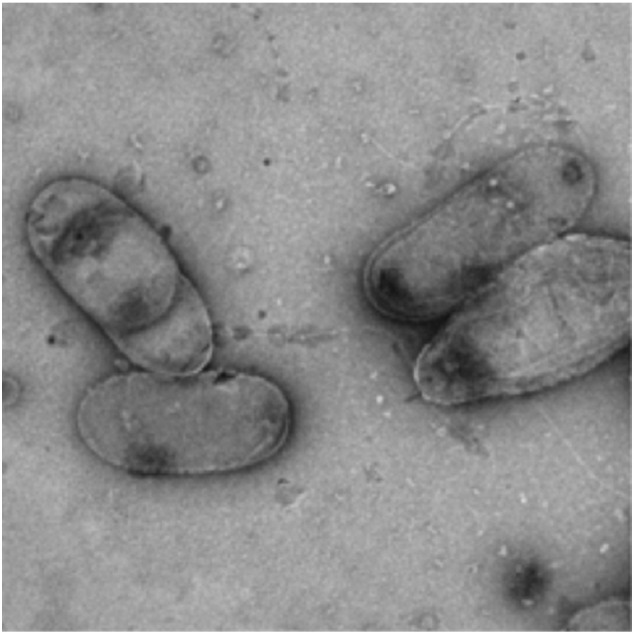
Negative stain TEM micrograph of Pseudomonas moraviensis strain Devor.
Within the genus Pseudomonas, 163 species are described with validly published names. Strain Devor shares 77.8%–99.8% 16S rRNA gene identities with other species in the Pseudomonas genus as follows: P. abietaniphilia (97.7%) type strain ATCC 700689T, P. aeruginosa (94.7%) type strain ATCC 10145T, P. aestusnigri (95.5%) VGXO14T, P. agarici (98%) type strain ATCC 25941T, P. alcaligenes (95.8%) type strain ATCC 14909T, P. alcalophilia (96.5%) type strain AL15-21T, P. amygdali (96.6%) type strain ATCC 33614T, P. anguilliseptica (94.7%) type strain ATCC 33660T, P. antarctica (97.3%) type strain DSM 15318T, P. argentinensis (97.6%) type strain CH01T, P. arsenicoxydans (97.7%) type strain VC-1T, P. asplenii (97.7%) type strain ATCC 23835T, P. asturiensis (97.1%) type strain LPPA 221T, P. asuensis (95.8%) type strain CP 155-2T, P. azotifigens (93.9%) type strain ATCC BAA-1049T, P. azotoformans (98.1%) type strain CCUG 12536T, P. baetica (99.2%) type strain a390T, P. balearica (95%) type strain DSM 6083T, P. bauzanensis (95%) type strain DSM 22558T, P. benzenivorans (96.9%) type strain DSM 8628T, P. borbori (96.1%) type strain DSM 17834, P. brassicacearum (97.1%) type strain DSM 13227T, P. brenneri (97.8%) type strain DSM 15294, P. caeni (93.8%) type strain HY-14T, P. cannabina (97.8%) type strain CFBP 2341T, P. carboxydohydrogena (77.8%) type strain ATCC 29978T, P. caricapapayae (97.7%) type strain ATCC 33615T, P. cedrina (98.2%) type strain DSM 17516T, P. chengduensis (96.6%) type strain DSM 26382, P. chloritidismutans (96.6%) type strain ATCC BAA-443T, P. chlororaphis (97.9%) type strain ATCC 9446T, P. cichorii (96.8%) type strain ATCC 10857T, P. citronellosis (94.4%) type strain ATCC 13674T, P. coleopterorum (97.8%) type strain LMG 28558T, P. composti (96.7%) type strain C2T, P. congealans (97.7%) type strain DSM 14939T, P. corrugata (97.7%) type strain ATCC 29736T, P. constantinii (97.4%) type strain PS 3aT, P. cremoricolorata (97.1%) type strain DSM 17059T, P. cuatrocienegasensis (96.5%) type strain LMG 24676T, P. deceptionensis (97.2%) type strain M1T, P. delhiensis (94.3%) type strain RLD-1T, P. duriflava (94.7%) type strain HR2T, P. elongata (89.3%) type strain ATCC 10144T, P. endophytica (97.3%) type strain BSTT44T, P. entomophila (97.7%) type strain L48T, P. extremaustralis (97.3%) type strain DSM 17835T, P. extremorientalis (97.5%) type strain KMM 3447T, P. ficuserectae (97.6%) type strain ATCC 35104T, P. flavescens (96.5%) type strain ATCC 51555T, P. fluorescens (96.8%) type strain ATCC 13525T, P. formosensis (94.1%) type strain CC-CY503T, P. fragi (97.2%) type strain ATCC 4973T, P. frederiksbergensis (97.3%) type strain ATCC BAA-257T, P. fulva (98.6%) type strain ATCC 31418T, P. fuscovaginae (97.5%) type strain DSM 7231T, P. gessardii (98.2%) type strain CIP 105469T, P. graminis (98%) type strain DSM 11363T, P. granadensis (99.4%) type strain DSM 28040T, P. grimontii (97.4%) type strain ATCC BAA-140T, P. guangdongensis (95.1%) type strain SgZ-6T, P. guariconensis (97.2%) type strain PCAVU11T, P. guineae (95.5%) type strain M8T, P. helmanticensis (98.8%) type strain OHA11T, P. hussainii (93.8%) type strain CC-AMH-11T, P. indica (94.1%) type strain DSM 14015T, P. japonica (96.9%) type strain WLT, P. jessennii (99.3%) type strain ATCC 700870T, P. jinjuensis (95%) type strain Pss 26T, P. kilonensis (97.7%) type strain DSM 13647T, P. knackmussii (94.5%) type strain DSM 6978T, P. koreensis (99.8%) type strain Ps 9-14T, P. kunmingensis (96.3%) type strain DSM 25974T, P. kuykendallii (97.7%) type strain H2T, P. libanensis (98.2%) type strain CCUG 43190T, P. lini (97.7%) type strain DLE411JT, P. linyingensis (95%) type strain LYBRD3-7T, P. litoralis (94.8%) type strain 2SM5T, P. lundensis (97.5%) type strain ATCC 49968T, P. lurida (97.4%) type strain DSM 15835T, P. lutea (98.2%) type strain OK2T, P. luteola (95.6%) type strain ATCC 43273T, P. mandelii (97.8%) type strain ATCC 700871T, P. marginalis (97.2%) type strain ATCC 10844T, P. marincola (96%) type strain JCM 14761T, P. matsuisoli (93.8%) type strain CC-MHH0089, P. mediterranea (97%) type strain CFBP 5447T, P. mendocina (96.2%) type strain ATCC 25411T, P. meridiana (97.1%) type strain DSM 15319T, P. migulae (98.1%) type strain CCUG 43165T, P. mohnii (99.2%) type strain DSM 18327T, P. monteilii (97.8%) type strain ATCC 700476T, P. moorei (99.3%) type strain DSM 12647T, P. moraviensis (99.4%) type strain DSM 16007T, P. mosselii (97.6%) type strain ATCC BAA-99T, P. mucidolens (98.2%) type strain ATCC 4685T, P. multiresinivorans (94.8%) type strain ATCC 700690T, P. nitritireducens (94.8%) type strain WZBFD3-5A2T, P. nitroreducens (94.9%) type strain ATCC 33634T, P. oleovorans (94.4%) type strain ATCC 8062T, P. orientalis (97.1%) type strain DSM 17489T, P. oryzihabitans (97.3%) type strain ATCC 43272T, P. otitidis (95%) type strain ATCC BAA-1130T, P. pachastrellae (95.2%) type strain CCUG 46540T, P. palleroniana (97.4%) type strain CFBP 4389T, P. panacis (97.7%) type strain CG20106T, P. panipatensis (94.5%) type strain Esp-1, P. parafulva (98.6%) type strain DSM 17004T, P. pelagia (94.3%) CL-AP6T, P. peli (95.5%) type strain DSM 17833T, P. pertucinogena (93.3%) type strain ATCC 190T, P. pictorum (84.4%) type strain ATCC 23328T, P. plecoglossicida (97.7%) type strain ATCC 700383T, P. poae (97.2%) type strain DSM 14936T, P. pohangensis (95.7%) type strain DSM 17875T, P. prosekii (97.8%) type strain AN/28/1T, P. protegens (97%) type strain DSM 19095T, P. proteolytica (97.4%) type strain DSM 15321T, P. pseudoalcaligenes (96.9%) type strain ATCC 17440T, P. psychrophilia (97.1%) type strain E-3T, P. psychrotolerans (94.7%) type strain DSM 15758T, P. punonensis (97.7%) type strain LMT03T, P. putida (97.2%) type strain ATCC 12633T, P. reinekei (99.5%) type strain DSM 18361T, P. resinovorans (93.6%) type strain ATCC 14235T, P. rhizosphaerae (97.8%) type strain IH5T, P. rhodesiae (97.4%) type strain DSM 14020T, P. sabulinigri (94.9%) type strain J64T, P. sagittaria (94.8%) type strain CC-OPY-1T, P. salegens (94%) type strain GBPy5T, P. salina (94.3%) type strain XCD-X85T, P. salomonii (96.9%) type strain CFBP 2022T, P. saponiphilia (96.5%) type strain DSM 9751T, P. savastanoi (97.6%) type strain ATCC 13522T, P. segetis (96.3%) type strain FR1439T, P. seleniipraecipitans (97%) type strain CA5T, P. simiae (97.5%) type strain OLiT, P. soli (97.8%) type strain DSM 28043T, P. straminea (97%) type strain ATCC 33636T, P. stutzeri (96.4%) type strain ATCC 17588T, P. synxantha (98.1%) type strain ATCC 9890T, P. syringae (97.6%) type strain ATCC 19310T, P. taenensis (96.9%) type strain MS-3T, P. taetrolans (97.1%) type strain ATCC 4683T, P. taiwanensis (97.8%) type strain DSM 21245T, P. testosteroni (80.8%) type strain ATCC 11996T, P. thivervalensis (97.6%) type strain DSM 13194T, P. tolaasii (97.4%) type strain ATCC 33618T, P. toyotomiensis (96.5%) type strain HT-3T, P. tremae (97.8%) type strain CFBP 3229T, P. trivialis (97.1%) type strain DSM 14937T, P. tuomuerensis (95.2%) type strain 78-123, P. usmongensis (98.9%) type strain Ps 3-10T, P. vancouverensis (99.3%) type strain ATCC 700688T, P. veronii (97.6%) type strain ATCC 700272/700474T, P. viridiflava (96.2%) type strain ATCC 13223T, P. vranovensis (96.7%) type strain DSM 16006T, P. xanthomarina (96.5%) type strain CCUG 46543T, P. xiamenensis (93.5%) type strain C10-2T, P. xinjiangensis (94.5%) type strain S3-3T, P. yamanorum (98%) type strain DSM 26522T, P. zeshuii (95.4%) type strain BY-1T, P. zhaodongensis (96.6%) type strain DSM 27559T.
Phylogenetic analysis based on the 16S rRNA gene identified strain WW1 to be the closest taxonomic relative of Pseudomonas moraviensis strain Devor (Table 2, and Fig. 2).
Table 2.
Classification and general features of Pseudomonas moraviensis strain Devor [28].
| MIGS ID | Property | Term | Evidence codea |
|---|---|---|---|
| Classification | Domain Bacteria | TAS [9] | |
| Phylum Proteobacteria | TAS [9] | ||
| Class Gammaproteobacteria | TAS [9] | ||
| Order Pseudomonadales | TAS [9] | ||
| Family Pseudomonadaceae | TAS [9] | ||
| Genus Pseudomonas | TAS [9] | ||
| Species moraviensis | TAS [9] | ||
| (Type) strain: Devor | |||
| Gram stain | Negative | TAS [9] | |
| Cell shape | Rod | TAS [9] | |
| Motility | Motile | TAS [9] | |
| Sporulation | Non-spore forming | TAS [9] | |
| Temperature range | 4–35 °C | TAS [9] | |
| Optimum temperature | 28–35 °C | TAS [9] | |
| pH range; optimum | Unknown | ||
| Carbon source | glycerol, l-arabinose, d-glucose, d-fructose, d-trehalose, d-lyxose, gluconate and 2-ketogluconate | TAS [9] | |
| MIGS-6 | Habitat | Soil/Gate | IDA |
| MIGS-6.3 | Salinity | Unknown | |
| MIGS-22 | Oxygen requirement | Aerobic | IDA |
| MIGS-15 | Biotic relationship | Free living | IDA |
| MIGS-14 | Pathogenicity | Non-pathogenic | NAS |
| MIGS-4 | Geographic location | Stillwater, OK, USA | IDA |
| MIGS-5 | Sample collection | March 2016 | IDA |
| MIGS-4.1 | Latitude | 36.1157 | IDA |
| MIGS-4.2 | Longitude | − 97.0586 | IDA |
| MIGS-4.4 | Altitude | 1 M |
Evidence codes - IDA: Inferred from Direct Assay; TAS: Traceable Author Statement (i.e., a direct report exists in the literature); NAS: Non-traceable Author Statement (i.e., not directly observed for the living, isolated sample, but based on a generally accepted property for the species, or anecdotal evidence). These evidence codes are from the Gene Ontology project [29].
Fig. 2.

A maximum likelihood phylogenetic tree constructed using multiple sequence alignments of 16S rRNA genes. “Pseudomonas moraviensis strain Devor” sequence is shown with in bold. Reference sequences are also shown and Genbank accession numbers are given in parentheses. The tree was obtained under “K2 + G” model with a variable site γ shape parameter of 0.05. “Pseudomonas aeruginosa 57 16S ribosomal RNA gene partial sequence” was used as the outgroup. Bootstrap values, in percent, are based on 100 replicates and are shown for branches with > 50% bootstrap support. Multiple sequence alignment, model selection, and maximum likelihood analysis were carried out in Mega [30].
Comparison to other Pseudomonas moraviensis sequenced genomes revealed that strain Devor shares 99% 16S rRNA sequence similarity to strain R28-S (Genbank accession number AYMZ00000000) [5].
3.2. Genome properties
The genome assembly process produced a contig N50 of 3,923,117 bp and a total genome size of 6,016,489 bp. The GC content was 60.0%. One hundred and eighty three RNA genes were identified including 3 ribosomal RNA and 63 tRNA genes. The ribosomal RNA operon showed an atypical organization, where 2 copies of 5S rRNA were present along with one copy of the 23S rRNA gene. Two copies each of the tRNAAla and the tRNAIle were present in the operon. The operon lacked a 16S rRNA gene. Of the 5473 genes detected, 5290 were protein-coding, of which 79.8% had a function prediction, 72.7% represented a COG functional category, and 11.3% were predicted to have a signal peptide. Using Psort [19], proteins were classified as 44.12% cytoplasmic, 1.06% extracellular, and 25.25% associated with the membrane. Based on the presence of 139 single copy genes [20], the genome is predicted to be 83.45% complete. Genome statistics are shown in Table 3. The distribution of genes into COG functional categories is shown in Table 4.
Table 3.
Genome statistics.
| Attribute | Value | % of Total |
|---|---|---|
| Genome size (bp) | 6,016,489 | 83.4 |
| DNA coding (bp) | 5,314,299 | 88.3 |
| DNA G + C (bp) | 3,609,956 | 60.0 |
| DNA scaffolds | 7 | 100 |
| Total genes | 5473 | 100 |
| Protein coding genes | 5290 | 96.6 |
| RNA genes | 183 | 3.3 |
| Pseudo genes | 0 | 0 |
| Genes in internal clusters | 1440 | 26.3 |
| Genes with function prediction | 4220 | 77.1 |
| Genes assigned to COGs | 3846 | 70.3 |
| Genes with Pfam domains | 4512 | 82.4 |
| Genes with signal peptides | 598 | 10.9 |
| Genes with transmembrane helices | 1208 | 22.1 |
| CRISPR repeats | 0 | 0 |
Table 4.
Number of genes associated with general COG functional categories.
| Code | Value | % age | Description |
|---|---|---|---|
| J | 236 | 5.40 | Translation, ribosomal structure and biogenesis |
| A | 2 | 0.05 | RNA processing and modification |
| K | 393 | 9.00 | Transcription |
| L | 124 | 2.84 | Replication, recombination and repair |
| B | 2 | 0.05 | Chromatin structure and dynamics |
| D | 40 | 0.92 | Cell cycle control, cell division, chromosome partitioning |
| V | 96 | 2.20 | Defense mechanisms |
| T | 286 | 6.55 | Signal transduction mechanisms |
| M | 270 | 6.18 | Cell wall/membrane biogenesis |
| N | 123 | 2.82 | Cell motility |
| U | 89 | 2.04 | Intracellular trafficking and secretion |
| O | 157 | 3.60 | Posttranslational modification, protein turnover, chaperones |
| C | 276 | 6.32 | Energy production and conversion |
| G | 230 | 5.27 | Carbohydrate transport and metabolism |
| E | 447 | 10.24 | Amino acid transport and metabolism |
| F | 96 | 2.20 | Nucleotide transport and metabolism |
| H | 206 | 4.72 | Coenzyme transport and metabolism |
| I | 213 | 4.88 | Lipid transport and metabolism |
| P | 253 | 5.79 | Inorganic ion transport and metabolism |
| Q | 121 | 2.77 | Secondary metabolites biosynthesis, transport and catabolism |
| R | 387 | 8.86 | General function prediction only |
| S | 263 | 6.02 | Function unknown |
| – | 1640 | 29.97 | Not in COGs |
The total is based on the total number of protein coding genes in the genome.
3.3. Insights from the genome sequence
Genome analysis of Pseudomonas moraviensis strain Devor identified a Gram negative microorganism with typical cell wall structure, a peptidoglycan layer lacking pentaglycine bridges with diaminopimelic acid as the second amino acid in the peptide linkage. Genes encoding enzymes for the biosynthesis of the glycerophospholipids phosphatidylcholine, phosphatidyl-glycerophosphate, phosphatidyl-serine, and phosphatidyl ethanolamine were identified in the genome. The analysis also revealed the presence of genes encoding a complete flagellar assembly in line with the evidence for the presence of flagella in electron micrographs of the organism (Fig. 1). Extracellular structures including type IV pili and the Flp system, and some structural genes for the production of exopolysaccharides including a partial Yjb operon [21], as well as evidence for the biosynthesis of multiple nucleotide sugars were also found in the genome.
Further analysis suggested the capability to utilize xylose, fructose, mannose, glucose, all amino acids except valine and tryptophan, and fatty acids as carbon and energy sources. Almost complete to complete catabolic KEGG pathways were identified in the genome for each of the above carbon sources. A complete TCA cycle and electron transport chain with F-type ATPase subunits confirmed the aerobic nature of the microorganism. Facultative fermentation capability was also identified in the genome where genes for lactate and acetate fermentation were present. Comparison of the protein-coding genes against the transporter database [22] identified ABC and secondary transporters for several amino acids.
When compared against the virulence factor database [23], the genome of Pseudomonas moraviensis strain showed 1733 virulence factor hits (32.8% of the protein-coding genes). These included secretion systems (type I, type II, and type IV secretion systems) among others.
The Devor genome also encoded several proteins that suggest the potential for using this strain in bioremediation efforts. These included genes encoding for 4-hydroxybenzoate degradation to succinate, as well as genes encoding for the detoxification of the plant-secreted toxin nitronate and the industry-applied chelating agent nitriloacetate [24], [25], [26], [27]. Evidence for the capability of salvaging S from organo-S-compounds (e.g. taurine and alkanesulfonates) in cases of limiting inorganic S were also identified in the genome.
3.4. Insights from comparative genomics
We used the COG profile to compare the genome of Pseudomonas moraviensis strain Devor to 13 closely related genomes. The genome clustered with Pseudomonas moraviensis strain BS3668 (Fig. 3A), which was in agreement with the phylogenetic position of the isolate as Pseudomonas moraviensis (Fig. 2). We used genomic features including the genome size, the number of genes, the number of transporters identified, the GC content, the number of non-coding bases, the number of genes belonging to COG categories, as well as the number of genes belonging to each COG category to cluster Pseudomonas moraviensis strain Devor's genome in comparison to the 13 other closely related genomes. Results are shown in Fig. 3B. The genome Pseudomonas moraviensis strain Devor clustered close to the two P. moraviensis genomes, and the P. koorensis genome as well. This is consistent with its phylogeny as a P. moraviensis and as a member of the P. koorensis clade [9]. These genomes appear to be enriched in the number of transporters identified in the genome.
Fig. 3.

Comparative genomics of Pseudomonas moraviensis strain Devor and 13 closely related genomes. (A) COG profile clustering of the genomes compared in this study. (B) Principal component analysis biplot of the genomic features and COG category distribution in the genomes compared. Genomes are represented by stars. Arrows represent genomic features or COG categories used for comparison. The arrow directions follow the maximal abundance, and their lengths are proportional to the maximal rate of change between genomes. The first two components explained 75% of variation. Color coding: P. jessenei, brown; P. baetica, orange; P. umsongenesis, purple; P. morviensis, red; P. reineke, grey; P. fluorescens, green; P. koorensis, black; P. moorei, blue.
4. Conclusions
This study presents the genome sequence and annotation of Pseudomonas moraviensis strain Devor. The genome revealed metabolic versatility with suggested capability to degrade fatty acids, most amino acids, fructose, mannose, and xylose. Comparison to the virulence factor database identified 1733 genes in the genome with potential virulence-associated function including type I, type II, and type IV secretion systems. The genome also suggests the potential use in bioremediation efforts with evidence for degradation of 4-hydroxybenzoate, nitronate, and nitriloacetate. Comparative genomics using general genomic features and the COG function profile are in line with the phylogenetic position predicted based on the 16S rRNA gene sequence placing strain Devor with other P. moraviensis representatives.
Competing interests
All authors declare no competing interests.
Funding
This is Draft Genome #7 in the SIMD project supported in part by a grant from the Howard Hughes Medical Institute (1554854) through the Science Education Program. WDH acknowledges support by NSF grants MCB-1051590, MRI-1338097, and CHE-1412500.
Authors' contributions
NTM, DF, MB, PB, MBC, and NY contributed to the analysis. NTM, DF, WDH, DPF, and NY wrote the manuscript. RCD, CB, AR, and RAH performed the lab experiments.
Transparency document
Transparency document
Footnotes
The Transparency document associated with this article can be found, in the online version.
References
- 1.Couger M.B., Hurlbut A., Murphy C.L., Budd C., French D.P., Hoff W.D., Elshahed M.S., Youssef N.H. Draft genome sequence of the environmental isolate Chryseobacterium sp. Hurlbut01. Genome Announc. 2015;3(5) doi: 10.1128/genomeA.01071-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Ash R.J., Mauck B., Morgan M. Antibiotic resistance of gram-negative bacteria in rivers, United States. Emerg. Infect. Dis. 2002;8(7):713–716. doi: 10.3201/eid0807.010264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Cho J.C., Tiedje J.M. Biogeography and degree of endemicity of fluorescent Pseudomonas strains in soil. Appl. Environ. Microbiol. 2000;66(12):5448–5456. doi: 10.1128/aem.66.12.5448-5456.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Grobe S., Wingender J., Truper H.G. Characterization of mucoid Pseudomonas aeruginosa strains isolated from technical water systems. J. Appl. Bacteriol. 1995;79(1):94–102. doi: 10.1111/j.1365-2672.1995.tb03129.x. [DOI] [PubMed] [Google Scholar]
- 5.Hunter S.S., Yano H., Loftie-Eaton W., Hughes J., De Gelder L., Stragier P., De Vos P., Settles M.L., Top E.M. Draft genome sequence of Pseudomonas moraviensis R28-S. Genome Announc. 2014;2(1) doi: 10.1128/genomeA.00035-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Kumar A., Kumar A. Isolation of a Pseudomonas aeruginosa strain capable of degrading acrylamide. J. Microbiol. Biotechnol. 1998;8(4):347–352. [Google Scholar]
- 7.Pandey A., Palni L.M.S. Isolation of Pseudomonas corrugata from Sikkim Himalaya. World J. Microbiol. Biotechnol. 1998;14(3):411–413. [Google Scholar]
- 8.Pandey A., Trivedi P., Kumar B., Palni L.M. Characterization of a phosphate solubilizing and antagonistic strain of Pseudomonas putida (B0) isolated from a sub-alpine location in the Indian Central Himalaya. Curr. Microbiol. 2006;53(2):102–107. doi: 10.1007/s00284-006-4590-5. [DOI] [PubMed] [Google Scholar]
- 9.Tvrzova L., Schumann P., Sproer C., Sedlacek I., Pacova Z., Sedo O., Zdrahal Z., Steffen M., Lang E. Pseudomonas moraviensis sp. nov. and Pseudomonas vranovensis sp. nov., soil bacteria isolated on nitroaromatic compounds, and emended description of Pseudomonas asplenii. Int. J. Syst. Evol. Microbiol. 2006;56(Pt 11):2657–2663. doi: 10.1099/ijs.0.63988-0. [DOI] [PubMed] [Google Scholar]
- 10.Verhille S., Baida N., Dabboussi F., Izard D., Leclerc H. Taxonomic study of bacteria isolated from natural mineral waters: proposal of Pseudomonas jessenii sp. nov. and Pseudomonas mandelii sp. nov. Syst. Appl. Microbiol. 1999;22(1):45–58. doi: 10.1016/S0723-2020(99)80027-7. [DOI] [PubMed] [Google Scholar]
- 11.Yang W., Cao H., Xu L., Zhang H., Yan Y. A novel eurythermic and thermostable lipase LipM from Pseudomonas moraviensis M9 and its application in the partial hydrolysis of algal oil. BMC Biotechnol. 2015;15:94. doi: 10.1186/s12896-015-0214-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Yoon K.-P. Isolation and characterization of Pseudomonas sp. KM10, a cadmium- and mercury-resistant, and phenol-degrading bacterium. J. Microbiol. Biotechnol. 1998;8(4):388–398. [Google Scholar]
- 13.Staicu L.C., Ackerson C.J., Cornelis P., Ye L., Berendsen R.L., Hunter W.J., Noblitt S.D., Henry C.S., Cappa J.J., Montenieri R.L. Pseudomonas moraviensis subsp. stanleyae, a bacterial endophyte of hyperaccumulator Stanleya pinnata, is capable of efficient selenite reduction to elemental selenium under aerobic conditions. J. Appl. Microbiol. 2015;119(2):400–410. doi: 10.1111/jam.12842. [DOI] [PubMed] [Google Scholar]
- 14.Zerbino D.R., Birney E. Velvet: algorithms for de novo short read assembly using de Bruijn graphs. Genome Res. 2008;18(5):821–829. doi: 10.1101/gr.074492.107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Hyatt D., Chen G.L., Locascio P.F., Land M.L., Larimer F.W., Hauser L.J. Prodigal: prokaryotic gene recognition and translation initiation site identification. BMC Bioinf. 2010;11:119. doi: 10.1186/1471-2105-11-119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Mistry J., Finn R.D., Eddy S.R., Bateman A., Punta M. Challenges in homology search: HMMER3 and convergent evolution of coiled-coil regions. Nucleic Acids Res. 2013;41(12) doi: 10.1093/nar/gkt263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Finn R.D., Coggill P., Eberhardt R.Y., Eddy S.R., Mistry J., Mitchell A.L., Potter S.C., Punta M., Qureshi M., Sangrador-Vegas A. The Pfam protein families database: towards a more sustainable future. Nucleic Acids Res. 2016;44(D1):D279–D285. doi: 10.1093/nar/gkv1344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Roberts D. labdsv: ordination and multivariate analysis for ecology. 2007.
- 19.Horton P., Park K.-J., Obayashi T., Fujita N., Harada H., Adams-Collier C.J., Nakai K. WoLF PSORT: protein localization predictor. Nucleic Acids Res. 2007;35:W585–W587. doi: 10.1093/nar/gkm259. Web Server issue. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Rinke C., Schwientek P., Sczyrba A., Ivanova N.N., Anderson I.J., Cheng J.F., Darling A., Malfatti S., Swan B.K., Gies E.A. Insights into the phylogeny and coding potential of microbial dark matter. Nature. 2013;499(7459):431–437. doi: 10.1038/nature12352. [DOI] [PubMed] [Google Scholar]
- 21.Ferrieres L., Aslam S.N., Cooper R.M., Clarke D.J. The yjbEFGH locus in Escherichia coli K-12 is an operon encoding proteins involved in exopolysaccharide production. Microbiology. 2007;153(Pt 4):1070–1080. doi: 10.1099/mic.0.2006/002907-0. Reading, England. [DOI] [PubMed] [Google Scholar]
- 22.Saier M.H., Jr., Reddy V.S., Tamang D.G., Vastermark A. The transporter classification database. Nucleic Acids Res. 2014;42(Database issue):D251–D258. doi: 10.1093/nar/gkt1097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Chen L., Yang J., Yu J., Yao Z., Sun L., Shen Y., Jin Q. VFDB: a reference database for bacterial virulence factors. Nucleic Acids Res. 2005;33:D325–D328. doi: 10.1093/nar/gki008. Database issue. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Francis K., Smitherman C., Nishino S.F., Spain J.C., Gadda G. The biochemistry of the metabolic poison propionate 3-nitronate and its conjugate acid, 3-nitropropionate. IUBMB life. 2013;65(9):759–768. doi: 10.1002/iub.1195. [DOI] [PubMed] [Google Scholar]
- 25.Salvi F., Agniswamy J., Yuan H., Vercammen K., Pelicaen R., Cornelis P., Spain J.C., Weber I.T., Gadda G. The combined structural and kinetic characterization of a bacterial nitronate monooxygenase from Pseudomonas aeruginosa PAO1 establishes NMO class I and II. J. Biol. Chem. 2014;289(34):23764–23775. doi: 10.1074/jbc.M114.577791. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Xu Y., Mortimer M.W., Fisher T.S., Kahn M.L., Brockman F.J., Xun L. Cloning, sequencing, and analysis of a gene cluster from Chelatobacter heintzii ATCC 29600 encoding nitrilotriacetate monooxygenase and NADH:flavin mononucleotide oxidoreductase. J. Bacteriol. 1997;179(4):1112–1116. doi: 10.1128/jb.179.4.1112-1116.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Zhang Y., Edwards T.E., Begley D.W., Abramov A., Thompkins K.B., Ferrell M., Guo W.J., Phan I., Olsen C., Napuli A. Structure of nitrilotriacetate monooxygenase component B from Mycobacterium thermoresistibile. Acta Crystallogr. Sect. F: Struct. Biol. Cryst. Commun. 2011;67(Pt 9):1100–1105. doi: 10.1107/S1744309111012541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Field D., Garrity G., Gray T., Morrison N., Selengut J., Sterk P., Tatusova T., Thomson N., Allen M.J., Angiuoli S.V. The minimum information about a genome sequence (MIGS) specification. Nat. Biotechnol. 2008;26(5):541–547. doi: 10.1038/nbt1360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Ashburner M., Ball C.A., Blake J.A., Botstein D., Butler H., Cherry J.M., Davis A.P., Dolinski K., Dwight S.S., Eppig J.T. Gene ontology: tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet. 2000;25(1):25–29. doi: 10.1038/75556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Kumar S., Stecher G., Tamura K. MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016;33(7):1870–1874. doi: 10.1093/molbev/msw054. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Transparency document