

To the Editor: Recently, Yi-Yun Liu et al. reported on the discovery of mcr-1, a plasmidborne resistance gene mediating resistance to colistin, in isolates obtained from humans and animals (1). Since the original publication, mcr-1 with or without the insertion element ISApl1 has been detected on plasmids of different incompatibility groups, including IncI2, IncHI2, and IncX4, and in many different countries (1–3). Because colistin is a last-resort parenteral antimicrobial drug, the transfer of mcr-1 by conjugation or through mobilizable plasmids raises concern about the emergence of pan-resistant Enterobacteriaceae.
We previously described extended-spectrum β-lactamase (ESBL)–producing and carbapenemase-producing isolates obtained from livestock and a human in Germany that harbored the mcr-1 gene (2). Because the transfer of mcr-1 through the food chain is highly likely, we looked for its presence in 62 whole-genome sequenced ESBL-producing Escherichia coli isolates obtained during 2012–2013 from food products sampled in Germany (Technical Appendix). We detected 4 isolates harboring the mcr-1 gene (E. coli RL138, RL145, RL158, and RL465) that displayed a colistin MIC of 4 mg/L (Technical Appendix Table 1). The raw sequencing reads and the assembled contigs of the mcr-1–positive isolates were deposited in the European Nucleotide Archive under project accession no. PRJEB13470. We conducted conjugation experiments to analyze the transferability of mcr-1 (Technical Appendix). For all isolates except RL465, mcr-1 was transferable to E. coli J53 Azr. For isolates RL138, RL145, and RL158, the mcr-1 gene was present on IncX4 and IncHI2 plasmids (Figure, panel A; Technical Appendix Table 2). The sequence type (ST) 410 E. coli isolate RL465 was detected in a turkey hen meat sample from 2013 and harbored blaCTX-M-15 and mcr-1, a gene combination hitherto identified only in travelers from the Netherlands and children from China (4). Both the blaCTX-M-15 and mcr-1 genes were not transferable, indicating that neither gene was plasmid-encoded. Examination of the genetic environment of mcr-1 in the assembled gapped genome showed a chromosomal location for the mcr-1 transposition unit that included an ISApl1 element (Figure 1, panel A; Technical Appendix Figure 1, panel A) flanked by the inverted repeats (IR-R1, IR-R2, and IR-L1). We verified the chromosomal location for the mcr-1 gene by sequencing the genome to completion, using long-read single-molecule real-time sequencing (Pacific Biosciences, Menlo Park, CA, USA; Technical Appendix Figure 2); the resulting contigs of E. coli RL465 Were deposited in the European Nucleotide Archive under accession no. PRJEB14095. One copy of the ISApl1-mcr-1 transposition unit was located in the region between a predicted 4Fe-4S ferredoxin-type protein (ydhY) and ldtE (L,D-transpeptidase) (bp 2652307–2665241), and flanked on either side by a 2-bp direct repeat (CA). We observed a similar situation for the ISEcp1-blaCTX-M-15–orf477 transposition unit (Technical Appendix Figure 1, panel B). However, this insertion mapped to a different chromosomal location in a region encoding a defective lambdoid prophage inserted between the molybdate ABC transporter operon (modABC) and the biotin biosynthesis operon (bioABCDF) (bp 1662140–1716472). It was flanked by direct repeats (TGGTT).
Figure.
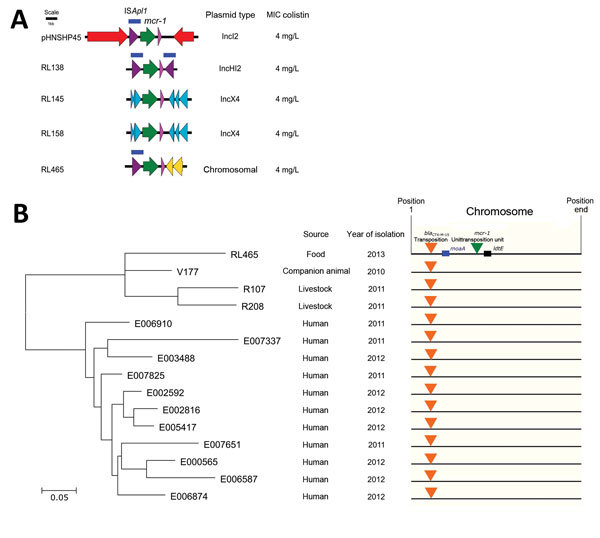
A) Genetic environment of mcr-1 in extended-spectrum β–lactamase (ESBL)–producing food isolates including the original environment found in pHNSHP45. Colors represent gene functions and insertion sequences: purple, transposase of ISApl1; dark blue, ISApl1 element; green, mcr-1; pink, hypothetical protein within the mcr-1 transposition unit; red, light blue, yellow, flanking genes specific for the respective isolate. B) Phylogenetic tree (unrooted) of the Escherichia coli sequence type 410 isolates harboring the blaCTX-M-15 transposition unit in the chromosome and a schematic depiction of the location of the mcr-1 and blaCTX-M-15 transposition unit. Isolates prefixed with the letter E represent consecutive isolates from 1 patient. Scale bar indicates nucleotide substitutions per site.
We reexamined our collection of 424 genome sequenced ESBL- and carbapenemase-encoding E. coli isolates, obtained during 2010–2014 (2), for isolates that harbored blaCTX-M-15 at a chromosomal location identical to that found in E. coli RL465. We detected 3 such isolates from 2010–2011 from companion animals and livestock (R107, sock swab dairy cattle farm, 2011; R208, sock swab pig fattening farm, 2011; V177, sick dog, 2010), and 11 consecutive isolates from a hemato-oncologic patient (5), obtained within an 11-month period during 2011–2012 (E006910, E007337, E007651, E007825, E000565, E002592, E002816, E003488, E005417, E006587, E006874) (Figure, panel B). All of these isolates were ST410 and negative for the mcr-1 gene. Phylogenetic analysis of the core genome of these isolates with E. coli RL465 using the program Harvest Suite (6) indicated they were highly related and separated from E. coli V177 (the oldest isolate) by 66 (E006910, E007651) to 110 (E007337) single-nucleotide polymorphisms (core genome size 94%, representing 4.58 Mbp). Thus, our results suggest that transposition of the ISApl1-mcr-1 unit to the chromosome in E. coli RL465 is a later event and probably occurred after transfer of the blaCTX-M-15 allele to the distinct chromosomal location into this E. coli ST410 subclone.
These findings highlight 2 independent points. First, our results extend data on the mobility of ISApl1-mcr-1 to a chromosomal location and reveal a new dimension in the transmissible nature of mcr-1 in colistin-resistant Enterobacteriaceae isolates and their ecology. Second, clonal isolates of ST410 have been isolated from diverse environments, livestock, companion animals, and humans and, as we demonstrate here, in turkey hen meat (7,8). Thus, the simultaneous spread of the mcr-1 and blaCTX-M-15 genes mediated by a single bacterial clone is real and suggests that mcr-1 is already present in the diverse reservoirs inhabited by these isolates.
Collection of isolates; antimicrobial susceptibility testing; whole-genome sequencing; in silico analyses; conjugation experiments; depiction of the minimal inhibitory concentration of the mcr-1–encoding and extended-spectrum β-lactamase (ESBL)–producing isolates from retail food; characteristics of the mcr-1–encoding ESBL-producing Escherichia coli isolates from retail meat; genetic environments of the chromosomally located antimicrobial resistance genes; schematic depiction of the chromosome, its IncFII/FIB plasmid, and the 2 phage elements in E. coli RL465.
Acknowledgments
We thank Christina Gerstmann, Natalia Lest, Simone Severitt, and Nicole Mrotzek for excellent technical assistance and Geovanna Brenner Michael, Yvonne Pfeifer, Axel Hamprecht, Lena Biehl, and Martin Kaase for providing the nonfood isolates.
This study was supported by grants to the German Center of Infection Research (DZIF), and the Zoonoses Network “ESBL and fluoroquinolone resistance in Enterobacteriaceae (RESET)” Consortium through the German Federal Ministry of Education and Research (BMBF; grant numbers 8000 701–3 [HZI], 01KI1013G and 01KI1313G to T.C. and C.I., and TI06.001 and 8032808811 to T.C.).
L.F., C.I., and T.C. conceived the study; S.E.W., K.G., H.G., S.D., B.B., C.S., and J.O. performed experiments; A.I., J.F., H.S., B.G., and A.K. contributed isolates and reagents; L.F., B.B., C.I., and T.C. analyzed the data; and T.C. and L.F. wrote the manuscript, which all authors approved.
B.G. is currently employed with the European Food Safety Authority (EFSA) in its BIOCONTAM Unit that provides scientific and administrative support to EFSA’s scientific activities in the area of Microbial Risk Assessment. The positions and opinions presented in this article are those of the authors alone and are not intended to represent the views or scientific works of EFSA. The other authors have nothing to proclaim.
Footnotes
Suggested citation for this article: Falgenhauer L, Waezsada SE, Gwozdzinski K, Ghosh H, Doijad S, Bunk B, et al. Chromosomal locations of mcr-1 and blaCTX-M-15 in fluoroquinolone-resistant Escherichia coli ST410 [letter]. Emerg Infect Dis. 2016 Sep [date cited]. http://dx.doi.org/10.3201/eid2209.160692
Current address: European Food Safety Authority, Parma, Italy.
References
- 1.Liu Y-Y, Wang Y, Walsh TR, Yi L-X, Zhang R, Spencer J, et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: a microbiological and molecular biological study. Lancet Infect Dis. 2016;16:161–8. 10.1016/S1473-3099(15)00424-7 [DOI] [PubMed] [Google Scholar]
- 2.Falgenhauer L, Waezsada S-E, Yao Y, Imirzalioglu C, Käsbohrer A, Roesler U, et al. RESET Consortium. Colistin resistance gene mcr-1 in extended-spectrum β-lactamase-producing and carbapenemase-producing gram-negative bacteria in Germany. Lancet Infect Dis. 2016;16:282–3. 10.1016/S1473-3099(16)00009-8 [DOI] [PubMed] [Google Scholar]
- 3.Hasman H, Hammerum AM, Hansen F, Hendriksen RS, Olesen B, Agersø Y, et al. Detection of mcr-1 encoding plasmid-mediated colistin-resistant Escherichia coli isolates from human bloodstream infection and imported chicken meat, Denmark 2015. Euro Surveill. 2015;20:30085. 10.2807/1560-7917.ES.2015.20.49.30085 [DOI] [PubMed] [Google Scholar]
- 4.Skov RL, Monnet DL. Plasmid-mediated colistin resistance (mcr-1 gene): three months later, the story unfolds. Euro Surveill. 2016;21:1–6. 10.2807/1560-7917.ES.2016.21.9.30155 [DOI] [PubMed] [Google Scholar]
- 5.Vehreschild MJGT, Hamprecht A, Peterson L, Schubert S, Häntschel M, Peter S, et al. A multicentre cohort study on colonization and infection with ESBL-producing Enterobacteriaceae in high-risk patients with haematological malignancies. J Antimicrob Chemother. 2014;69:3387–92 . 10.1093/jac/dku305 [DOI] [PubMed] [Google Scholar]
- 6.Treangen TJ, Ondov BD, Koren S, Phillippy AM. The Harvest suite for rapid core-genome alignment and visualization of thousands of intraspecific microbial genomes. Genome Biol. 2014;15:524. 10.1186/s13059-014-0524-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Falgenhauer L, Imirzalioglu C, Ghosh H, Gwozdzinski K, Schmiedel J, Gentil K, et al. Circulation of clonal populations of fluoroquinolone-resistant CTX-M-15-producing Escherichia coli ST410 in humans and animals in Germany. Int J Antimicrob Agents. 2016;47:457–65. 10.1016/j.ijantimicag.2016.03.019 [DOI] [PubMed] [Google Scholar]
- 8.Schaufler K, Semmler T, Wieler LH, Wöhrmann M, Baddam R, Ahmed N, et al. Clonal spread and interspecies transmission of clinically relevant ESBL-producing Escherichia coli of ST410—another successful pandemic clone? FEMS Microbiol Ecol. 2016;92:fiv155. 10.1093/femsec/fiv155 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Collection of isolates; antimicrobial susceptibility testing; whole-genome sequencing; in silico analyses; conjugation experiments; depiction of the minimal inhibitory concentration of the mcr-1–encoding and extended-spectrum β-lactamase (ESBL)–producing isolates from retail food; characteristics of the mcr-1–encoding ESBL-producing Escherichia coli isolates from retail meat; genetic environments of the chromosomally located antimicrobial resistance genes; schematic depiction of the chromosome, its IncFII/FIB plasmid, and the 2 phage elements in E. coli RL465.