

Summary
The incidence of Lyme disease has continued to rise despite attempts to control its spread. Vaccination of zoonotic reservoirs of human pathogens has been successfully used to decrease the incidence of rabies in raccoons and foxes. We have previously reported on the efficacy of a vaccinia virus vectored vaccine to reduce carriage of Borrelia burgdorferi in reservoir mice and ticks. One potential drawback to vaccinia virus vectored vaccines is the risk of accidental infection of humans. To reduce this risk, we developed a process to encapsulate vaccinia virus with a pH-sensitive polymer that inactivates the virus until it is ingested and dissolved by stomach acids. We demonstrate that the vaccine is inactive both in vitro and in vivo until it is released from the polymer. Once released from the polymer by contact with an acidic pH solution, the virus regains infectivity. Vaccination with coated vaccinia virus confers protection against B. burgdorferi infection and reduction in acquisition of the pathogen by naïve feeding ticks.
Keywords: Reservoir-targeted vaccine, Lyme disease, polymethylacrylate, Safety, Encapsulation, Borrelia burgdorferi
Introduction
Lyme disease is the most commonly reported vector-borne disease in the Northern Hemisphere. The disease is caused by the bacterium Borrelia burgdorferi, which is transmitted to humans via the bite of infected Ixodes ticks [1]. Its incidence and geographic distribution continue to grow, despite efforts to control its spread [2]. A human vaccine was previously available but has since been removed from the market [3]. Interrupting the transmission of the organism in its wild reservoirs is an alternative strategy for reducing human disease.
The lifecycle of B. burgdorferi is closely linked to the lifecycle of its tick vector. There is no transovarial transmission of B. burgdorferi, so larval ticks hatch from eggs uninfected. Larval Ixodes ticks acquire the bacteria by feeding on infected rodents and birds. Infected larvae maintain B. burgdorferi transstadially and molt into nymphal ticks, which infected new rodents and birds during their next feeding. Disruption of the enzootic cycle of the pathogen could reduce the incidence of Lyme disease. In the U.S. the white-footed mouse, Peromyscus leucopus, is the primary reservoir of B. burgdorferi [4–6]. By targeting the major reservoir, a vaccine could reduce the carriage in these animals, resulting in a reduction in the number of infected ticks and a subsequent decrease in the incidence of human disease [7,8].
Outer surface protein A (OspA) was the antigen used in the approved human vaccine [9]. OspA is expressed by B. burgdorferi during tick colonization and is rapidly downregulated during transmission to a mammalian host. Vaccination with OspA is thought to prevent infection through uptake of antibodies by a feeding tick during its blood meal. Anti-OspA antibodies kill B. burgdorferi in the tick before it is able to shut off OspA production and infect a vaccinated host. In animal studies, vaccination with OspA does not result in clearance of infection in animals already infected with B. burgdorferi due to lack of expression of the protein in the mammalian host [10]. However, uninfected ticks feeding on infected, vaccinated animals show reduced acquisition of B. burgdorferi [11,12]. The low level of OspA expression in the mouse host makes it unlikely that vaccination will select for resistant OspA mutants in vaccinated but infected animals. These characteristics make OspA an excellent antigen for a reservoir targeted vaccine of wildlife.
Two trials of OspA vaccination in wild mice to reduce B. burgdorferi infection have been reported. Vaccination of wild mice with OspA by injection has been shown to reduce the prevalence of B. burgdorferi infected ticks in the subsequent year, providing proof of principle for development of an ecologically targeted Lyme vaccine [13]. A recent field trial of an oral, recombinant protein vaccine showed increased OspA seropositivity in white-footed mice. Repeated trapping and feeding of wild mice with E. coli expressing recombinant OspA showed some efficacy in reducing carriage of B. burgdorferi in ticks in some but not all test sites [14]. Immunogenicity of the recombinant OspA was weak despite multiple doses administered to most mice (5–60 doses). Low titers of antibody were likely responsible for the variability of protection seen during this field trial. Despite mixed results, this study confirms the overall feasibility of oral vaccination in wildlife and suggests that more immunogenic vaccines are likely to show improved efficacy.
An alternative to recombinant protein oral vaccination is vaccination with virally vectored vaccines. A vaccinia virus vectored oral vaccine expressing the rabies virus glycoprotein is available commercially (Raboral) and has been highly effective in preventing rabies in raccoons and foxes [15,16]. Vaccinia virus expressing B. burgdorferi OspA (VV-ospA) has been shown to be highly immunogenic. VV-ospA was effective in protecting mice from B. burgdorferi infection after a single dose when administered orally [17,18]. Although much more immunogenic than recombinant protein vaccines, there are potential risks when using virally vectored vaccines. With over 80 million doses of the rabies-vaccinia virus vaccine distributed in the wild, the vaccine has not caused any environmental issues or adverse events in wildlife. However, there have been two cases of human vaccinia virus infections acquired from contact with the rabies vaccine. Both occurred when humans tried to remove a vaccine containing bait from the mouth of a domesticated dog [19,20]. The chance of human infection with the VV-ospA vaccine are less than with the rabies vaccine. Formulations of rabies vaccines for raccoons and foxes include a liquid center containing virus surrounded by a bait. Formulations of vaccine baits for mice do not use a liquid center. However, to further reduce the potential for human infection, we developed a vaccine encapsulated with pH sensitive polymers designed to inactivate the virus until contact with stomach fluids. Encapsulation has been widely used in human drug delivery to target drugs to specific locations in the gastrointestinal tract [21,22]. Eudragit® is a cationic, nontoxic and biocompatible family of polymer that has been widely tested in pharmaceutical applications. These polymethacrylate-based copolymers are designed to dissociate at specific pHs [23]. They have not previously been used to encapsulate live viruses due to permanent inactivation of the virus by solvents used in the coating process [24]. Here, we report a strategy for encapsulating virus in these polymers without permanent inactivation of the virus. We show that the vaccine is non-infectious in vitro and in vivo when coated. Release of the virus occurs only after exposure to fluids at pH less than 5. A single dose of encapsulated virus reduced infection of vaccinated mice with B. burgdorferi and decreased transmission of B. burgdorferi to uninfected ticks. Our encapsulation strategy increased the safety of the vaccine, while maintaining its protection against B. burgdorferi infection. This work lays the basis for improved targeted delivery of live virus vaccines.
Materials and Methods
Vaccinia virus, bacteria and mice strains
Vaccinia virus expressing OspA (VV-ospA) was previously constructed [17]. All strains of vaccinia virus (VV) were grown and maintained in Vero cells as previously described [25]. VV-vp37 which was also previously constructed [17] was used as a control.
B. burgdorferi sensu stricto (strain N40) was used for the experiments. The spirochetes were grown in Barbour–Stoenner–Kelly II (BSK) media at 37°C.
C3H/HeN and Balb/c mice were purchased from Charles River Laboratories (Boston, MA).
Ticks
Ixodes scapularis tick larvae were obtained from National Tick Research and Education Center, Oklahoma State University (Stillwater, OK). B. burgdorferi-infected nymphs were generated as previously described [17]. Briefly, uninfected larvae were allowed to feed to repletion on B. burgdorferi-infected SCID mice. The engorged larvae were collected and allowed to molt into nymphs in 4–6 weeks at room temperature and high relative humidity. Prevalence of B. burgdorferi infection in fed larvae was determined by culture of a portion of the recovered ticks from each batch.
Virus encapsulation
Eudragit® E100 (referred to EE100) was kindly provided by Evonik Industries, USA. Vaccinia virus was coated with EE100 by solvent evaporation. The polymer was dissolved in toluene with the addition of Span®60 as an emulsifier (20% w/v of EE100 in toluene supplemented with 5% w/v of Span®60). The virus was mixed into the organic phase at a ratio of 1:10 (virus:organic phase) by vigorous vortexing. This solution (200μL) was added dropwise into an external aqueous phase, consisting of 10mL of distilled water and 500μL of Tween 80 (5% v/v). The mixture was stirred for 24 hours at room temperature to allow the solvent to evaporate. During the evaporative process, the virus and polymer are co-precipitated in the droplets forming microparticles. The microparticles containing vaccinia virus were then collected and washed with phosphate buffer at a pH 7.0 and allowed to air dry.
Morphology and size analysis
The morphology of microparticles was evaluated by scanning electron microscopy. The particles were fixed in 2.5% gluteraldehyde, 3% paraformaldehyde. The samples were then coated with a thin layer of gold using a sputter coater (Quorum Technologies Ltd., West Sussex, UK), and imaged using SEM (Carl Zeiss, Thornwood, NY).
Preparation of oral baits
The bait formulation consisted of a proprietary mixture of natural food products developed by Foodsource Lures Corp. (Alabaster, AL) as previously described [18]. Peanut butter was added as an attractant. The ingredients were thoroughly mixed and allowed to cool to around 40°C. The desired dose of recombinant VV was added to the semi-solid bait mix and cooled at room temperature to make solid bait blocks.
Determination of virus titer
Viral titering was performed as previously described [25]. Briefly, serial dilutions of virus were plated on BSC-1 cells for 2 hours. The inoculum was then removed and replaced by fresh MEM-5% FBS in each well. After 2 days, the media was removed and cells were stained using 0.1% crystal violet to facilitate counting of the plaques.
Mouse vaccination
C3H/HeN mice were vaccinated with VV-ospA or a control (VV-vp37) vaccine with or without encapsulation, delivered by oral baits that were placed in cages for the mice to ingest. Only one mouse was kept per cage and each cage was supplied with approximately 2 g of bait containing 2 × 108 pfu of VV-ospA or control virus (VV-vp37). All mice consumed the baits within 48h.
Determination of antibody response
The antibody response in mice was determined by endpoint titer ELISA using a protocol similar to that described previously [18]. Briefly, 96-well plates (Costar, Corning, NY) were coated with 2 μg/ml of MBP-OspA diluted in PBS and incubated overnight at 4°C. The MBP-OspA was removed and the plates were blocked with 1% BSA dissolved in PBS. After removing the blocking reagent, the top well of each lane was loaded in duplicate with serum dilutions. The samples were serially diluted in equal volume of PBS. Following overnight incubation at 4°C, the samples were aspirated and the plates were washed 0.1% Tween 20 in PBS. The plates were incubated with anti-mouse IgG, HRP-linked secondary antibody (1:10,000 dilution in PBS) (Cell Signaling Technology) for 1 h at room temperature. The secondary antibody was aspirated, washed and incubated with Sure Blue TMB peroxidase substrate (KPL, Gaithersburg, MD) followed by addition of equal volume of stop solution (1N HCl solution). Absorbance was measured at 450 nm. Serum from unvaccinated mice was used as the negative control. The cutoff value was determined as three standard deviations above the average OD of negative controls. The reciprocal of highest dilution above the cut-off was defined as endpoint antibody titer of each serum sample.
Determination of VV-ospA efficacy
Efficacy of the VV-ospA vaccine was determined for both inhibition of larval tick infection and for inhibition of transmission to uninfected mice by infected nymphal ticks as described in [17].
To test the efficacy against transmission to uninfected mice, mice were provided VV-ospA baits or control (VV-vp37) baits. Antibody response was determined 4 weeks post vaccination. On week 5 post-vaccination, mice were challenged by allowing 3 to 4 B. burgdorferi infected nymphal ticks to feed to repletion as previously described. The fed nymphs were collected and assayed for B. burgdorferi infection by culture in BSK media. The mice were assayed for B. burgdorferi infection by culture of an ear punch biopsy in BSK media at week 2 and cultures of the ear, bladder and heart culture at week 4 post-challenge. Presence of B. burgdorferi was determined by observing the cultures by dark field microscopy. A mouse was defined as infected with B. burgdorferi if one or more organ cultures were found positive by darkfield microscopy. Negative cultures were also tested by PCR specific to B. burgdorferi.
To determine the vaccine efficacy in preventing B. burgdorferi acquisition by larvae, mice were infected with B. burgdorferi by allowing 3–4 infected nymphal ticks to feed on these mice. Infection was confirmed by culture of an ear punch in BSK at weeks 2 after tick infestation. The infected mice were vaccinated with VV-ospA or control by bait feeding and serum antibody response to OspA was determined 4 weeks post-vaccination by ELISA. Ixodes larvae were fed on VV-ospA and control vaccinated mice. The fed ticks were collected and observed for 5 days. After 5 days, the ticks were homogenized and cultured in BSK media. The presence of B. burgdorferi was determined by darkfield microscopy observation.
Tail scarification
Balb/c mice were anesthetized by inhalation of isoflurane gas. The tail cleaned with 70% ethanol and allowed to dry. Thirty scarifications were made with a 27 G syringe along the base of the tail as described [26,27]. A droplet of 10 μl of uncoated virus (VV-ospA), intact EE100 coated virus, dissolved EE100 coated virus or PBS was deposited on the dorsal base of the tail and allowed to air dry. 106 PFU of virus was used. A similar protocol was used with bait containing the virus with the exception that the bait was rubbed onto the tail.
Statistics
Fisher’s exact test was used for comparisons between groups in protection experiments. All statistics were generated using Prism (GraphPad)
Ethics Statement
All animal procedures were carried out in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health and were reviewed and approved by the Tufts University Institutional Animal Care and Use Committee.
Results
Vaccinia virus viability in different solvents
The majority of organic solvents used to solubilize polymer EE100 destroy the infectivity of vaccinia virus. To determine which solvents could be used in the preparation of polymer encapsulated vaccinia virus, we incubated the virus in common organic solvents used for polymer preparation. We titered the virus on BSC-1 cells after 15 minutes or 3 hours exposure at a 1:1 ratio of the viral preparation to solvent. As expected, we observed a complete inactivation of the virus in most solvents (Table 1). Exposure to chloroform and dichloromethane (DCM) resulted in a drastic decrease of infectious virus after 15 minutes and complete inactivation within 3 hours. Short exposure to toluene, within a timeframe for vaccine preparation, did not affect the virus’ ability to infect BSC-1 cells. Three hours incubation in toluene resulted in about 60% loss of viral infectivity compared with virus incubated in PBS (pH 7.2). We also tested exposure to higher ratios of toluene to vaccinia virus (10:1 and 5:1) for 15 minutes and neither ratio significantly affected the virus (data not shown).
Table 1.
Vaccinia virus viability in organic solvents
Vaccinia virus was exposed to different organic solvents at a 1:1 ratio for 15 minutes and 3 hours. The virus was titered on BSC-1 cells after exposure and the percent of infectious virus was calculated in comparison to the control virus incubated in MEM medium at 1:1 ratio.
| Buffer or solvent | % of infectious virus after 15 min exposure | % of infectious virus after 3 hours exposure | ||
|---|---|---|---|---|
|
| ||||
| average | SD | average | SD | |
|
| ||||
| PBS buffer pH 7.2 | 100 | +/− 3.2 | 100 | +/− 3.0 |
| Toluene | 94 | +/− 7.1 | 38.5 | +/− 2.6 |
| Chloroform | 0.4 | +/− 0.5 | 0.0 | N/a |
| DCM (dimethyl carbonate) | 0.8 | +/− 1.6 | 0.0 | N/a |
| DMF (dimethylformamide) | 0 | N/a | 0.0 | N/a |
| DMSO (dimethyl sulfoxide) | 0 | N/a | 0.0 | N/a |
| Isopropanol | 0 | N/a | N/a | N/a |
| Methanol | 0 | N/a | N/a | N/a |
Efficacy of the encapsulation of vaccinia virus with EE100
Toluene was used as a solvent to dissolve the pH sensitive polymer, EE100. It is a poly (methacylic acid)-based polymer soluble in fluids with pH below 5.0 (Figure 1). A single emulsion process combined with solvent evaporation was chosen to coat VV. Briefly, the virus was dispersed in an organic polymer solution, which is then emulsified in an external aqueous phase. The virus and polymer were precipitated in microparticles by evaporation. The EE100 microparticles containing the virus are shown in Figure 2. The size of the microparticles ranges from 5–100 μm.
Figure 1.

Eudragit E100 chemical structure
Figure 2. Microparticles photographs.

(A) Photograph of the EE100 microparticles encapsulated with VV-ospA in aqueous solution. (B, C) Optical and SEM image of the microparticles.
The efficiency of encapsulation was determined by measuring the amount of virus released from the microparticles dissolved at pH 5.0 and the amount of residual virus in the supernatant and in the washes by plaque assay. Approximately 75% of the total starting virus was encapsulated within the polymer and remained infectious (Table 2).
Table 2.
Vaccinia virus encapsulation efficiency in EE100
Vaccinia virus was encaspulated in EE100. The encapsulation efficiency was determined by plaque assay on BSC-1 cells in the supernatant collected after microspheres preparation, in the washes and after dissolution at pH 5.0. The percent of infectious virus was calculated relatively to the amount of starting material.
| % of virus
|
||
|---|---|---|
| average | SD | |
| Supernatant after microspheres collection | 8.9 | +/− 2.5 |
| Wash #1 in PBS (pH 7.2) | 1.2 | +/− 0.8 |
| Wash #2 in PBS (pH 7.2) | 0.9 | +/− 0.3 |
| After microspheres dissolution (pH 5.0) | 75.8 | +/− 11.3 |
We then investigated the targeted release in specific pH-buffered solutions. We measured the virus released over time by exposing the microparticles to PBS at pH 7.2 or to a simulated gastric fluid (0.01M acetic acid and 0.01M sodium citrate adjusted to pH 5.0) (Figure 3). At pH 7.2, the overall release was less than 1% of the virus over a 2-hour exposure period. Virus detected after exposure at pH 7.2 is likely due to virus attached to the surface of the microparticles, as the titer did not increase during the 2-hour exposure period suggesting that no release was occurring. In contrast, exposure to a pH 5.0 solution resulted in increasing viral titers, plateauing at 1 hour.
Figure 3. Release of infectious vaccinia virus from EE100 microspheres in different pH buffered solutions.
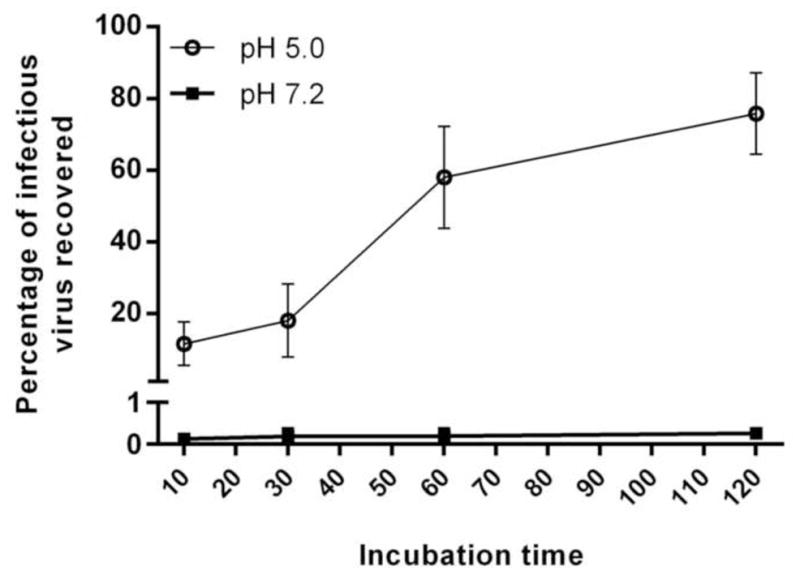
Encapsulated vaccinia virus was exposed to pH buffered solutions for up to 2 hours and the virus released was quantified by plaque assay on BSC-1 cells. The results are expressed in percent of virus recovered compared to the amount of virus before encapsulation. Statistical difference observed at all times points (P values < 0.05). P values generated by analyzing data with an unpaired t test corrected for multiple comparisons using the Holm-Sidak method (Prism, GraphPad Software).
Preparation and evaluation of bait containing the microparticles
To test the release characteristics of the encapsulated VV-ospA, we added encapsulated or unencapsulated vaccine to bait formulation that will be used to deliver vaccine to animals in the wild. Baits containing the VV-microparticles were exposed to a pH 5.0 or pH 7.2 solution and the viable virus released was measured by plaque assay. We selected an exposure time of one hour to mimic transit time through the mouse stomach [28]. At pH 5.0, 61% of the virus was released from the bait, whereas only 14% was released after exposure to pH 7.2 (Figure 4). The virus released from the bait is greater that the virus recovered after exposure to pH 7.2-buffered solution. This is most likely due to dissolution of the bait matrix, which lowers the pH. Buffering of the bait matrix is likely to reduce the release of the coated virus.
Figure 4. Infectious vaccinia virus released from bait containing microparticles.

Baits containing encapsulated virus were disaggregated in pH 5.0 or pH 7.2 buffer. Supernatant was tested for the presence of infectious vaccinia virus on BSC-1 cells. P value = 0.0016 obtained by analyzing data with a t test using Prism (GraphPad Software).
Protection against accidental infection with encapsulated virus
We then tested whether encapsulation prevents accidental infection during contact by scarification of Balb/c mice. Scarification is a traditional method for transmitting vaccinia virus and is used for administration of the human smallpox vaccine. Balb/c mice were chosen because their light skin color makes visualization of viral vesicles easier. Vaccinia virus, intact microparticles, dissolved microparticles or saline were applied to mouse tails using a scarification needle. The presence of a productive infection was monitored visually by inspection for the development of vesicles (Figure 5A–D). To check microparticle integrity during the bait preparation, we also applied baits by scarification on the mouse tails (Figure 5E–F). Mice scarified with virus, dissolved microparticles or bait containing the virus showed cutaneous lesions 8 days post scarification. Mice scarified with PBS, intact microparticles and bait containing microparticles showed no lesions.
Figure 5. Scarification lesion on mice tails.

Balb/c mice were scarified using a 26 gauge needle with the indicated encapsulated or unencapsulated vaccine or control. Mice were monitored every other day. Photographs show typical appearance one week post scarification. Pictures are representative of 3 independent experiments.
Efficacy of the coated virus in preventing the transmission of B. burgdorferi to uninfected mice
To determine the efficacy of the encapsulated virus in protecting uninfected mice against transmission of B. burgdorferi from infected ticks, C3H/HeN mice were vaccinated with oral baits containing coated or uncoated virus. After 4–5 weeks, we allowed Borrelia-infected nymphs to feed to repletion on the vaccinated mice. Mice were sacrificed 2 weeks post-challenge and tissues were tested by culture and PCR for the presence of B.burgdorferi. Serum was collected pre-vaccination, at 2 and 4 weeks after vaccination and at time of euthanasia (7–8 weeks after vaccination) and tested for anti-OspA antibodies. Six mice out of seven vaccinated with the coated virus developed a detectable antibody response against OspA. Protection against B. burgdorferi was correlated with the antibody titer, as the only mouse vaccinated with the coated virus that acquired B. burgdorferi infection upon tick feeding had no detectable anti-OspA antibody titers post-vaccination (Table 3).
Table 3.
Efficacy of coated VV-ospA bait vaccine in prevention of B. burgdorferi transmission to C3H/HeN mice.
Mice were fed with baits containing with 2×108 pfu VV-ospA or control (VV-vp37) encapsulated in EE100. On week 5 post-vaccination, the mice were challenged through infected nymph feeding. The presence of spirochetes in a mouse was tested by culturing ear punches, bladder, and heart two weeks post challenge.
| Total mice | # Mice infected | % Mice infecteda | |
|---|---|---|---|
| E100® coated VV-ospA | 7 | 1 | 14% |
| E100® coated VV-vp37 | 3 | 3 | 100% |
Statistical analyses (Fisher’s exact test) showed a significant degree of protection from spirochete infection in VV-ospA vaccinated mice compared to controls (p = 0.03).
Efficacy of the coated virus in preventing the acquisition of B. burgdorferi by larvae ticks from infected mice
C3H/HeN mice were infected by B. burgdorferi through feeding of infected ticks. These infected mice were then vaccinated with EE100 coated VV-OspA or control virus (VV-vp37). Naïve larvae were placed on these mice five weeks post vaccination and were allowed to fed to repletion. Fed larvae were tested 5 days after collection by culture for the presence of live spirochetes. We observed a significant reduction in B. burgdorferi transmission from vaccinated mice to tick larvae during feeding in the VV-ospA vaccinated group compared to the control group (Table 4). Those results were comparable to the results obtained previously with the uncoated virus, suggesting that polymer encapsulation does not reduce the efficacy of the vaccine [18]. A subset of the fed larva was allowed to molt into nymphal ticks. Infection rate of these nymphs was tested by crushing the ticks individually in BSK and culture them for two weeks. The presence of spirochetes was determined by dark-field microscopy. Spirochetes were observed in 14 out of 15 nymphs fed on control vaccinated mice (93%), whereas B. burgdorferi was detected in 14 out of 30 nymphs fed on VV-ospA vaccinated mice (47%). These results are comparable to the infection rates observed in the fed larvae.
Table 4.
Efficacy of coated VV-ospA vaccine to prevent acquisition of B. burgdorferi by naïve larvae.
B. burgdorferi-infected mice were vaccinated with EE100 encapsulated VV-ospA or control vaccine via bait-feeding. Five weeks post vaccination, uninfected tick larvae were allowed to feed on these mice to repletion. Presence of the spirochetes in fed larvae was tested by culturing crushed ticks in BSK media. Statistical analyses (Fisher’s exact test) showed a significant difference in acquisition of the infection by larvae feeding on VV-ospA vaccinated mice compared to control vaccinated mice.
| Total mice | Total fed larvae | # Larvae infected | % of infected larvaea | |
|---|---|---|---|---|
| VV-OspA coated with EE100 | 5 | 82 | 35 | 42.68% |
| VV-vp37 coated with EE100 | 5 | 61 | 66 | 92.42% |
The two-tailed P value is less than 0.0001.
Discussion
Many of the emerging infectious diseases worldwide are zoonotic. Previous work has shown that reservoir-targeted vaccination can be a successful approach to management of zoonotic diseases [29]. The vaccinia virus based vaccine to prevent rabies in foxes and raccoons has been very effective in eradicating rabies in areas where it has been deployed and in preventing the spread of rabies to new areas [15,16,30,31]. A raccoonpox vectored vaccine to prevent plague in prairie dogs is currently in advanced field trials. To date, there have been no reported adverse effects in prairie dogs and other small rodents that consumed baits containing the raccoon poxvirus [32,33]. While the distribution of vaccines containing live poxviruses has resulted in no discernable impact on the environment, release of live viruses does pose an exposure risk for humans [19,20,34]. Reducing human exposure to vaccinia infection while retaining the protective efficacy would facilitate the acceptance and widespread deployment of VV-OspA and similar reservoir targeted vaccines. Acrylic polymers have long been used in the pharmaceutical industry for targeted drug delivery to specific locations in the gastrointestinal tract. The Eudragit® family of polymers has been successfully used to deliver peptide and protein based vaccines [35–37]. Recently, these polymers have been used to encapsulate inactivated oral influenza virus vaccines [38]. The main hurdle to using polymers for encapsulation of live viral vaccines has been the toxicity of organic solvents used to manufacture the coatings for viruses. In fact, Eudragit® polymers have been used to capture and permanently inactivate enveloped viruses [24].
Herein we report the development of a safe and efficient polymer coating for a live virus based vaccine. Our goal was to encapsulate the virus in a pH-sensitive polymer known to dissolve in acidic pH. Thus, the virus should only be released and active after passing through the low pH environment in the stomach and not be released upon accidental contact with the skin. EE100 coating of vaccinia virus successfully decreased infectivity of the virus prior to ingestion without impairing its efficacy as an oral vaccine. Prevention of transmission of B. burgdorferi to vaccinated mice and prevention of acquisition of B. burgdorferi by uninfected ticks feeding on infected, vaccinated animals, and was similar to that seen with unencapsulated vaccine. We believe that the development of this technique to encapsulate live viruses with pH sensitive polymers that allow targeting of release to specific regions of the intestinal tract could be applied to other oral, virally vectored vaccines.
In addition to enhancing safety of virally vectored vaccines by reducing the chance of accidental infection, polymer encapsulation could also be used to enhance the potency of viral vaccines. Many viruses, including vaccina virus, are inactivated by exposure to acid. Using additional layers of polymers with different pH solubilities to target release of the virus beyond the stomach could potentially decrease the amount of virus needed to vaccinate an animal. Development of this technology could significantly improve the safety and cost of virally vectored oral vaccines.
Highlights.
Reservoir-targeted vaccination is a promising method for preventing Lyme disease
Virally vectored vaccines are highly immunogenic but pose an exposure risk for humans
Coating with a pH-sensitive polymer targets gastrointestinal release of the virus
Polymer coatings increase safety of vaccinia vaccines by preventing unwanted exposure
Encapsulation with a pH sensitive polymer does not reduce efficacy of the vaccine
Acknowledgments
The authors wish to thank Meghan Ramsey, Erin Troy and Tanja Petnicki-Ocwieja for their helpful discussions. The project was supported by R42AI078631 (L.T.H.; S.R.T), a pilot award from the Pacific Southwest Regional Center of Excellence, U54AI065359 (L.T.H.; Q.X.) and Tufts Innovation Institute funding program.
Footnotes
Conflicts of interest
Conflicts of interest and financial disclosure: none
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Radolf JD, Caimano MJ, Stevenson B, Hu LT. Of ticks, mice and men: understanding the dual-host lifestyle of Lyme disease spirochaetes. Nat Rev Microbiol. 2012;10:87–99. doi: 10.1038/nrmicro2714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Mead PS. Epidemiology of Lyme disease. Infect Dis Clin North Am. 2015;29:187–210. doi: 10.1016/j.idc.2015.02.010. [DOI] [PubMed] [Google Scholar]
- 3.Nigrovic LE, Thompson KM. The Lyme vaccine: a cautionary tale. Epidemiol Infect. 2007;135:1–8. doi: 10.1017/S0950268806007096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Mather TN, Wilson ML, Moore SI, Ribeiro JM, Spielman A. Comparing the relative potential of rodents as reservoirs of the Lyme disease spirochete (Borrelia burgdorferi) Am J Epidemiol. 1989;130:143–50. doi: 10.1093/oxfordjournals.aje.a115306. [DOI] [PubMed] [Google Scholar]
- 5.Donahue JG, Piesman J, Spielman A. Reservoir competence of white-footed mice for Lyme disease spirochetes. Am J Trop Med Hyg. 1987;36:92–6. doi: 10.4269/ajtmh.1987.36.92. [DOI] [PubMed] [Google Scholar]
- 6.Anderson JF. Ecology of Lyme disease. Conn Med. 1989;53:343–6. [PubMed] [Google Scholar]
- 7.Anderson RM. Rabies control. Vaccination of wildlife reservoirs. Nature. 1986;322:304–5. doi: 10.1038/322304a0. [DOI] [PubMed] [Google Scholar]
- 8.Gortazar C, Diez-Delgado I, Barasona JA, Vicente J, la Fuente de J, Boadella M. The Wild Side of Disease Control at the Wildlife-Livestock-Human Interface: A Review. Front Vet Sci. 2014;1:27. doi: 10.3389/fvets.2014.00027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Steere AC, Sikand VK, Meurice F, Parenti DL, Fikrig E, Schoen RT, et al. Vaccination against Lyme disease with recombinant Borrelia burgdorferi outer-surface lipoprotein A with adjuvant. Lyme Disease Vaccine Study Group. N Engl J Med. 1998;339:209–15. doi: 10.1056/NEJM199807233390401. [DOI] [PubMed] [Google Scholar]
- 10.Fikrig E, Barthold SW, Flavell RA. OspA vaccination of mice with established Borrelia burgdorferi infection alters disease but not infection. Infect Immun. 1993;61:2553–7. doi: 10.1128/iai.61.6.2553-2557.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.de Silva AM, Fish D, Burkot TR, Zhang Y, Fikrig E. OspA antibodies inhibit the acquisition of Borrelia burgdorferi by Ixodes ticks. Infect Immun. 1997;65:3146–50. doi: 10.1128/iai.65.8.3146-3150.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Tsao J, Barbour AG, Luke CJ, Fikrig E, Fish D. OspA immunization decreases transmission of Borrelia burgdorferi spirochetes from infected Peromyscus leucopus mice to larval Ixodes scapularis ticks. Vector Borne Zoonotic Dis. 2001;1:65–74. doi: 10.1089/153036601750137705. [DOI] [PubMed] [Google Scholar]
- 13.Tsao JI, Wootton JT, Bunikis J, Luna MG, Fish D, Barbour AG. An ecological approach to preventing human infection: vaccinating wild mouse reservoirs intervenes in the Lyme disease cycle. Proc Natl Acad Sci USa. 2004;101:18159–64. doi: 10.1073/pnas.0405763102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Meirelles Richer L, Aroso M, Contente-Cuomo T, Ivanova L, Gomes-Solecki M. Reservoir targeted vaccine for lyme borreliosis induces a yearlong, neutralizing antibody response to OspA in white-footed mice. Clin Vaccine Immunol. 2011;18:1809–16. doi: 10.1128/CVI.05226-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Cliquet F, Aubert M. Elimination of terrestrial rabies in Western European countries. Dev Biol (Basel) 2004;119:185–204. [PubMed] [Google Scholar]
- 16.Mackowiak M, Maki J, Motes-Kreimeyer L, Harbin T, Van Kampen K. Vaccination of wildlife against rabies: successful use of a vectored vaccine obtained by recombinant technology. Adv Vet Med. 1999;41:571–83. doi: 10.1016/s0065-3519(99)80043-3. [DOI] [PubMed] [Google Scholar]
- 17.Scheckelhoff MR, Telford SR, Hu LT. Protective efficacy of an oral vaccine to reduce carriage of Borrelia burgdorferi (strain N40) in mouse and tick reservoirs. Vaccine. 2006;24:1949–57. doi: 10.1016/j.vaccine.2005.10.044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Bhattacharya D, Bensaci M, Luker KE, Luker G, Wisdom S, Telford SR, et al. Development of a baited oral vaccine for use in reservoir-targeted strategies against Lyme disease. Vaccine. 2011;29:7818–25. doi: 10.1016/j.vaccine.2011.07.100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Rupprecht CE, Blass L, Smith K, Orciari LA, Niezgoda M, Whitfield SG, et al. Human infection due to recombinant vaccinia-rabies glycoprotein virus. N Engl J Med. 2001;345:582–6. doi: 10.1056/NEJMoa010560. [DOI] [PubMed] [Google Scholar]
- 20.Centers for Disease Control and Prevention (CDC) Human vaccinia infection after contact with a raccoon rabies vaccine bait - Pennsylvania, 2009. MMWR Morb Mortal Wkly Rep. 2009;58:1204–7. [PubMed] [Google Scholar]
- 21.Lavelle EC. Targeted delivery of drugs to the gastrointestinal tract. Crit Rev Ther Drug Carrier Syst. 2001;18:341–86. [PubMed] [Google Scholar]
- 22.Lavelle EC, O’Hagan DT. Delivery systems and adjuvants for oral vaccines. Expert Opin Drug Deliv. 2006;3:747–62. doi: 10.1517/17425247.3.6.747. [DOI] [PubMed] [Google Scholar]
- 23.Thakral S, Thakral NK, Majumdar DK. Eudragit: a technology evaluation. Expert Opin Drug Deliv. 2013;10:131–49. doi: 10.1517/17425247.2013.736962. [DOI] [PubMed] [Google Scholar]
- 24.Alasino RV, Bianco ID, Vitali MS, Zarzur JA, Beltramo DM. Characterization of the inhibition of enveloped virus infectivity by the cationic acrylate polymer eudragit E100. Macromol Biosci. 2007;7:1132–8. doi: 10.1002/mabi.200700102. [DOI] [PubMed] [Google Scholar]
- 25.Earl PL, Moss B, Wyatt LS, Carroll MW. Generation of recombinant vaccinia viruses. Curr Protoc Mol Biol. 2001;Chapter 16(Unit16):17. doi: 10.1002/0471142727.mb1617s43. [DOI] [PubMed] [Google Scholar]
- 26.Sharon M, Nir P, Lior K, David B-N, Tomer I, Paula S, et al. Tail scarification with Vaccinia virus Lister as a model for evaluation of smallpox vaccine potency in mice. Vaccine. 2007;25:7743–53. doi: 10.1016/j.vaccine.2007.09.023. [DOI] [PubMed] [Google Scholar]
- 27.Mota BEF, Gallardo-Romero N, Trindade G, Keckler MS, Karem K, Carroll D, et al. Adverse events post smallpox-vaccination: insights from tail scarification infection in mice with Vaccinia virus. PLoS ONE. 2011;6:e18924. doi: 10.1371/journal.pone.0018924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Schwarz R, Kaspar A, Seelig J, Künnecke B. Gastrointestinal transit times in mice and humans measured with 27Al and 19F nuclear magnetic resonance. Magnetic Resonance in Medicine. 2002;48:255–61. doi: 10.1002/mrm.10207. [DOI] [PubMed] [Google Scholar]
- 29.Cross ML, Buddle BM, Aldwell FE. The potential of oral vaccines for disease control in wildlife species. The Veterinary Journal. 2007;174:472–80. doi: 10.1016/j.tvjl.2006.10.005. [DOI] [PubMed] [Google Scholar]
- 30.Müller T, Bätza H-J, Freuling C, Kliemt A, Kliemt J, Heuser R, et al. Elimination of terrestrial rabies in Germany using oral vaccination of foxes. Berl Munch Tierarztl Wochenschr. 2012;125:178–90. [PubMed] [Google Scholar]
- 31.Brochier B, Aubert MF, Pastoret PP, Masson E, Schon J, Lombard M, et al. Field use of a vaccinia-rabies recombinant vaccine for the control of sylvatic rabies in Europe and North America. Rev - Off Int Epizoot. 1996;15:947–70. doi: 10.20506/rst.15.3.965. [DOI] [PubMed] [Google Scholar]
- 32.Tripp DW, Rocke TE, Streich SP, Abbott RC, Osorio JE, Miller MW. Apparent field safety of a Racoon poxvirus-vectored plague vaccine in free-ranging prairie dogs (Cynomys spp) Vol. 51. Colorado, USA: The Society for the Experimental Analysis of Behavior, Inc. Department of Psychology, Indiana University, Bloomington, Indiana 47405; 2015. [DOI] [PubMed] [Google Scholar]
- 33.Rocke TE, Pussini N, Smith SR, Williamson J, Powell B, Osorio JE. Consumption of Baits Containing Raccoon Pox-Based Plague Vaccines Protects Black-Tailed Prairie Dogs (Cynomys ludovicianus) 2010;10:53–8. doi: 10.1089/vbz.2009.0050. Http://DxDoiorg/101089/Vbz20090050. [DOI] [PubMed] [Google Scholar]
- 34.Rocke TE, Dein FJ, Fuchsberger M, Fox BC, Stinchcomb DT, Osorio JE. Limited infection upon human exposure to a recombinant raccoon pox vaccine vector. Vaccine. 2004;22:2757–60. doi: 10.1016/j.vaccine.2004.01.030. [DOI] [PubMed] [Google Scholar]
- 35.Hori M, Onishi H, Machida Y. Evaluation of Eudragit-coated chitosan microparticles as an oral immune delivery system. Int J Pharm. 2005;297:223–34. doi: 10.1016/j.ijpharm.2005.04.008. [DOI] [PubMed] [Google Scholar]
- 36.Delgado A, Lavelle EC, Hartshorne M, Davis SS. PLG microparticles stabilised using enteric coating polymers as oral vaccine delivery systems. Vaccine. 1999;17:2927–38. doi: 10.1016/s0264-410x(99)00140-1. [DOI] [PubMed] [Google Scholar]
- 37.Zhu Q, Talton J, Zhang G, Cunningham T, Wang Z, Waters RC, et al. Large intestine-targeted, nanoparticle-releasing oral vaccine to control genitorectal viral infection. Nat Med. 2012;18:1291–6. doi: 10.1038/nm.2866. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Shastri PN, Kim M-C, Quan F-S, D’Souza MJ, Kang S-M. Immunogenicity and protection of oral influenza vaccines formulated into microparticles. J Pharm Sci. 2012;101:3623–35. doi: 10.1002/jps.23220. [DOI] [PMC free article] [PubMed] [Google Scholar]