

Abstract
Triple negative breast cancer (TNBC), which does not express the progesterone, estrogen, or HER2/neu receptor, is aggressive and difficult to treat. Paclitaxel, a tubulin stabilizing agent, is one of the most frequently prescribed anticancer agents for breast cancers, including TNBC. Residual disease that occurs due to resistance or partial resistance of cancer cells in a tumor against anticancer agents is the most important issue in oncology. In the present study, when MDA-MB-231 cells, a TNBC cell line, were treated with 30 µM paclitaxel, a slightly higher concentration than its GI50 value, for 6 days, a small number of cells with different morphologies survived. Among the surviving cells, small round cells were isolated, cloned, and named MDA-MB-231-JYJ cells. MDA-MB-231-JYJ cells were observed to be highly proliferative and tumorigenic. In addition, signal transduction molecules involved in proliferation, survival, malignancy, or stemness of cancer cells, such as c-Src, c-Met, Notch 1, c-Myc, Sox2, Oct3/4, Nanog, and E-cadherin were highly expressed or activated. While further study is required, MDA-MB-231-JYJ cells appear to have some of the characteristics of cancer precursor cells. Although MDA-MB-231-JYJ cells were isolated from the cells that survived in the continuous presence of paclitaxel, they were not resistant to paclitaxel but developed resistance to dasatinib, a Bcr-Abl and Src kinase family inhibitor. The activated state of Src and Notch 1, and the expression levels of c-Myc and cyclins in MDA-MB-231-JYJ cells were less affected than MDA-MB-231 cells by the treatment of dasatinib, which may explain the resistance of MDA-MB-231-JYJ cells to dasatinib. These results suggest that cancer cells that become resistant to dasatinib during the process of paclitaxel therapy in patients may appear, and caution is required in the design of clinical trials using these two agents.
Keywords: breast cancer, dasatinib, paclitaxel, drug resistance
Introduction
Breast cancer is the leading type of cancer in women worldwide (1). Several therapeutic strategies including hormone blocking therapy, chemotherapy, and monoclonal antibodies, are used to treat breast cancer. Paclitaxel exerts anti-tubulin activity, and is one of a number of chemotherapeutic agents used in the treatment of patients with breast cancer (2,3). Paclitaxel inhibits mitotic progression by interfering with the homeostasis of microtubule assembly, and induces apoptotic cell death. As has been demonstrated previously with a number of other chemotherapeutic agents, the emergence of cancer resistance to paclitaxel is an important issue in the clinic (4). It has been suggested that resistance to paclitaxel is associated with the overexpression of multidrug transporters such as P-glycoprotein1 (also known as multidrug resistant protein 1) (5), and alterations in the tubulin/microtubule system (6). Cancer cells that are resistant to paclitaxel can be established in vitro by culturing them in the presence of increasing concentrations of paclitaxel for several months. The final concentration at the end of the establishment process of paclitaxel resistant cancer cells is far beyond its GI50 concentration. A recent study has shown that patients treated with 175 mg/m2 paclitaxel for 3 h had plasma concentrations ranging from 80–280 nM, and intratumoral concentrations of 1.1–9.0 µM at 20 h following administration of the agent (7). These high intratumoral concentrations are due to the intracellular accumulation of paclitaxel. In addition, the study showed that breast cancer cell lines treated with low nanomolar concentrations of paclitaxel (5–50 nM for MDA-MB-231 cells and 10–50 nM for Cal51 cells), had intracellular concentrations of paclitaxel in the range of 1–9 µM, which is a clinically relevant concentration range. This suggests that low nanomolar concentrations of paclitaxel can mimic intratumoral concentrations. The aim of the present study therefore, was to examine whether nanomolar concentrations of paclitaxel, which mimic intratumoral concentrations, are sufficient to induce death of the TNBC cell line MDA-MB-231 in vitro; and to isolate and characterize any surviving cells.
Materials and methods
Cell culture and cloning of MDA-MB-231-JYJ cells
The MDA-MB-231 human breast adenocarcinoma cell line was obtained from the American Type Culture Collection (Manassas, VA, USA) and maintained in RPMI 1640 medium supplemented with 10% heat-inactivated fetal bovine serum (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA) and 1% penicillin/streptomycin (100 µg/ml). Cells were incubated at 37°C in a humidified atmosphere containing 5% CO2. MDA-MB-231 cells were treated with paclitaxel (Sigma-Aldrich, St. Louis, MO, USA) at a concentration of 0.03 µM for 2 days. Any dead cells were removed by gentle washing with RPMI 1640 medium, and this step was repeated twice with the surviving cells. The remaining cells were briefly incubated in the presence of 0.025% trypsin to ensure that only dead, rounded cells detached from the bottom of the culture plate. The surviving adherent cells were then passaged again to remove the majority of cells with different morphologies. Finally, the remaining round cells were diluted in maintenance culture medium and seeded in 96-well culture plates. A clone was isolated and named MDA-MB-231-JYJ.
Cell proliferation assay
MDA-MB-231 and MDA-MB-231-JYJ cells were seeded in 100-mm dishes at a density of 1×105 cells/ml. The cells were trypsinized at 24, 48, and 72 h and stained with trypan blue. The number of viable cells was counted using a hemocytometer.
Human tumor xenograft in nude mice
6-week-old specific-pathogen-free BALB/c nude mice (Charles River Development, Inc., Tokyo, Japan) were maintained as previously described (8). The protocols regarding the use of animals were reviewed by the Korea University Institutional Animal Care & Use Committee (Seoul, Korea). MDA-MB-231 cells, suspended in 200 µl of a 1:1 mixture of phosphate-buffered saline (PBS) (pH 7.4) and BD Matrigel™ (BD Biosciences, Franklin Lakes, NJ, USA), and MDA-MB-231-JYJ cells suspended in 200 µl of PBS (pH 7.4), were injected subcutaneously at a concentration of 6×106 cells/mouse. Tumor volumes were measured three times a week for 3 weeks using a Vernier caliper, and subsequently calculated using the formula 0.5 × height × length × width.
Western blotting analysis
Protein extracts (20 µg) obtained from the cell lysates were resolved on 8–10% sodium dodecyl sulphate polyacrylamide gels and transferred to Immobilon-P transfer membranes as described previously (9). The membranes were incubated in Tris-buffered saline containing 0.2% Tween-20 and 5% nonfat dried milk, and probed with rabbit monoclonal antibodies against Src (36D10) (Cat No. 2110), E-cadherin (24E10) (Cat No. 3195), Notch 1 (C44H11) (Cat No. 3268), cleaved Notch 1 (V744) (D3B8) (Cat No. 4147), and c-Myc (D3N8F) (Cat No. 13987), and with rabbit polyclonal antibodies against phospho-Src (Y416) (D49G4) (Cat No. 6943), Akt (11E7) (Cat No. 4685), phospho-Akt (S473) (D9E) (Cat No. 4060), Erk1/2 (137F5) (Cat No. 4695), phospho-Erk1/2 (T202/Y204) (Cat No. 4370), c-Met (D1C2) (Cat No. 8198), phospho-c-Met (Y1234/1235) (Cat No. 3077), Sox2 (D6D9) (Cat No. 3579), Nanog (D73G4) (Cat No. 4903), Oct3/4A (C30A3) (Cat No. 2840) and β-actin (Cat No. A5441). After washing, membranes were probed with a horseradish peroxidase-conjugated secondary antibody. Detection was performed using an enhanced chemiluminescent protein detection system (Amersham Biosciences, Little Chalfont, United Kingdom) and exposure was carried out using X-ray film. All antibodies were obtained from Cell Signaling Technologies (Danvers, MA, USA) and diluted 1:1,000 to 1:2,000 before use except the antibodies against β-actin (1:10,000).
Cell cycle analysis
Cell cycle distribution was analyzed using a modification of a previously described protocol (10). Briefly, cells were plated at a density of 1×106 cells/100-mm dish. Cells were cultured for 48 h, collected, fixed in 70% ethanol, washed, and stained with Krishan buffer (0.1% sodium citrate, 20 µg/ml RNase A, 0.3% Triton X-100 and 50 µg/ml propidium iodide, pH 7.4). Samples were centrifuged, resuspended in 1 ml PBS (pH 7.4) and applied to flow cytometry using an LSRFortessa™ Cell Analyzer (BD Biosciences, San Jose, CA, USA). Data were collected for 10,000 events and analyzed with the FlowJo ver. 9.3.3 (Tree Star Inc., Ashland, OR, USA).
Cell growth inhibition assay
Inhibition of cell growth by anticancer agents was determined according to the sulforhodamime B (SRB) assay (11). Cells were seeded at a density of 3×104 cells/well in a 96-well plate and incubated for 24 h. Cells were further incubated for 48 h in the presence and absence of the compounds listed in Table I, fixed, and stained with SRB. The absorbance was measured at 565 nm.
Table I.
Comparison of the GI50 concentrations of anticancer agents against MDA-MB-231 and MDA-MB-231-JYJ cells.
| GI50 (µM) | ||
|---|---|---|
| Compounds | MDA-MB-231 | MDA-MB-231-JYJ |
| SN-38 | 0.034 | 0.018 |
| 5-FU | 2.202 | 2.339 |
| Docetaxel | 0.003 | 0.003 |
| Paclitaxel | 0.008 | 0.021 |
| Dasatinib | 0.014 | N/C |
| Doxorubicin | 0.447 | 0.103 |
GI50, growth inhibition of 50%; 5-FU, 5-fluorouracil; N/C, not calculatable (because MDA-MB-231-JYJ cells did not die much even at higher concentrations than 10 µM of dasatinib).
Statistical analysis
Graphpad Prism 5.03 (GraphPad Software, Inc., La Jolla, CA, USA) was used for statistical analysis. One-way ANOVA and Dunnett's t-test were used for multiple comparisons. P<0.05 was considered to indicate a statistically significant difference.
Results
Cloning and characterization of MDA-MB-231-JYJ cells
MDA-MB-231 cells cultured in culture medium containing 10% fetal bovine serum had a flat and spindle shape (Fig. 1A). After MDA-MB-231 cells were treated with 0.03 µM paclitaxel (the GI50 concentration) for 4 days, the majority of cells were dead, with the exception of a few cells with varying morphologies. Among the surviving cells, the small, round cells were isolated, cloned, and named MDA-MB-231-JYJ. These cells tended to grow together without spreading out evenly (Fig. 1B). Furthermore, MDA-MB-231-JYJ cells were highly proliferative compared with MDA-MB-231 cells. After 3 days, the number of MDA-MB-231 cells increased from 1×105 to 26.33±0.88, while the number of MDA-MB-231-JYJ cells increased to 86.83±1.59 (Fig. 1C), a statistically significant difference. MDA-MB-231 cells are tumorigenic when they are transplanted subcutaneously into BALB/c nude mice, producing small tumors (68.27±72.14 mm3) by day 15. However, the growth rate of the tumors formed by MDA-MB-231-JYJ cells was significantly higher, with a mean volume at day 15 of 2865.71±1091 mm3 (Fig. 1D).
Figure 1.
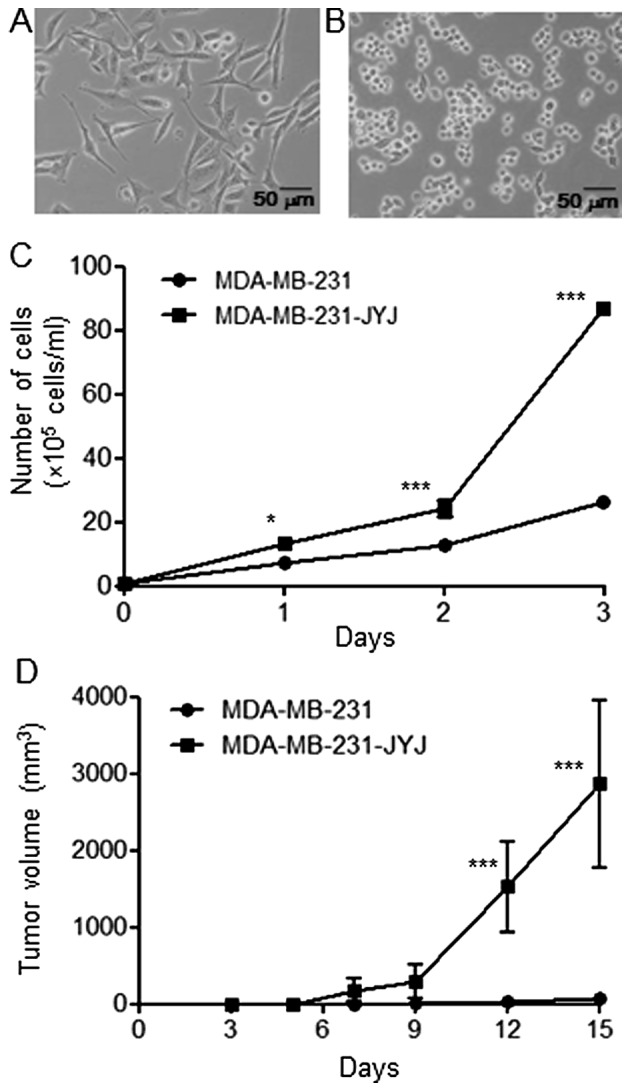
Morphology, proliferation rates, and tumorigenicity of MDA-MB-231 and MDA-MB-231-JYJ breast cancer cells. (A) MDA-MB-231 and (B) MDA-MB-231-JYJ cells were cultured in vitro and observed under an optical microscope. (C and D) The proliferation rates and tumorigenicity of these two cell lines were determined as described in the Materials and methods section. Results are presented as the mean ± SD of triplicate determinations. *P<0.05, ***P<0.001.
Since the rates of proliferation and tumor growth of MDA-MB-231-JYJ cells were significantly greater than those of MDA-MB-231 cells (Fig. 1C and D), the activation status of signal transduction molecules known to be involved in the regulation of cell survival, proliferation, and apoptosis was compared between the two cell types (Fig. 2B). Levels of phosphorylated c-Src and c-Met (also known as hepatocyte growth factor receptor) in MDA-MB-231-JYJ cells, which are involved in the invasive growth of cancer, were elevated compared to MDA-MB-231 cells. However, levels of Akt and phosphor-Erk1/2, which are involved in the regulation of cell survival, were lower in MDA-MB-231-JYJ cells than in MDA-MB-231 cells. The activation and expression of signal transduction molecules that increase the malignancy or stemness of cancer cells were also compared (Fig. 2B). While the expression and cleavage of Notch 1 was either barely detected or not detected at all in MDA-MB-231 cells, they were greatly increased in MDA-MB-231-JYJ cells. Similarly, expression of Sox2, Oct3/4, c-Myc, Nanog, and E-cadherin was absent or barely detectable in MDA-MB-231 cells, but the expression of these molecules was highly increased in MDA-MB-231-JYJ cells.
Figure 2.

Expression and phosphorylation of signal transduction molecules that regulate proliferation, survival, and pluripotency. Proteins were detected by western blot in MDA-MB-231 and MDA-MB-231-JYJ breast cancer cells. Cells in the exponential growth phase were lysed, and the phosphorylation, expression, and activation of signal transduction molecules involved in (A) proliferation and survival and (B) malignancy and pluripotency of cancer cells were examined.
Selective resistance of MDA-MB-231-JYJ cells to dasatinib
To examine whether the MDA-MB-231-JYJ cells could develop resistance to a number of anticancer agents, they were treated with SN-38 (an active metabolite of irinotecan), 5-fluorouracil (5-FU), docetaxel, paclitaxel, dasatinib, and doxorubicin, and their GI50 concentrations were calculated for both MDA-MB-231 and MDA-MB-231-JYJ cells (Table I). Although MDA-MB-231-JYJ cells were isolated from cells treated with paclitaxel, the GI50 concentrations of paclitaxel in these cells only slightly increased from 0.008 to 0.021 µM, showing they maintained susceptibility to the drug. By contrast, MDA-MB-231-JYJ cells were resistant only to dasatinib of all the anticancer agents tested. While the GI50 concentration of dasatinib for MDA-MB-231 cells was 0.014 mM, the concentration for MDA-MB-231-JYJ cells was >10 µM, indicating that these cells had become resistant.
Change in the response of signal transduction molecules to dasatinib
To understand the mechanism by which dasatinib caused the growth inhibition of MDA-MB-231 cells, the expression and/or activation status of signal transduction molecules known to regulate cell survival or proliferation were measured in the presence and absence of dasatinib. Phosphorylation of c-Src (p-Src) in MDA-MB-231 cells was almost completely inhibited by 0.01 and 0.03 µM dasatinib. However in MDA-MB-231-JYJ cells, phosphorylation of c-Src was only slightly inhibited at these concentrations, meaning the signal transduction pathway regulated by c-Src was activated at higher concentrations of dasatinib in MDA-MB-231-JYJ cells (Fig. 3). While the expression of c-Myc, a representative signal transduction molecule involved in cell proliferation, gradually decreased with increasing concentrations of dasatinib in MDA-MB-231 cells, it did not decrease at all in MDA-MB-231-JYJ cells, even at the highest concentration of dasatinib. Both the expression and cleavage of Notch 1, which has been reported to be activated in malignant tumors (12), were inhibited with increasing concentrations of dasatinib in MDA-MB-231 cells, but were unchanged in MDA-MB-231-JYJ cells (Fig. 3).
Figure 3.

Effect of dasatinib on the phosphorylation of c-Src, the expression of c-Myc and Notch 1, and the cleavage of Notch 1. MDA-MB-231 and MDA-MB-231-JYJ breast cancer cells were incubated for 24 h in the presence of dasatinib at the indicated concentrations. Cell lysates were subjected to Western blotting using antibodies specific to phospho-c-Src (Y416), c-Myc, Notch 1, and cleaved Notch 1.
In order to examine whether these changes in signal transduction regulatory proteins affected cell cycle progression, levels of cyclins D1, A2 and B were measured in both cell populations following treatment with dasatinib. As shown in Fig. 4, dasatinib gradually inhibited the expression of Cyclin D1 and A2 with increasing concentrations in MDA-MB-231 cells. It is possible that this inhibition contributed, at least in part, to the growth inhibition of MDA-MB-231 cells by dasatinib. However, dasatinib did not inhibit the expression of Cyclin D1, A2, or B in MDA-MB-231-JYJ cells. Flow cytometry analysis supports the results obtained with cyclins (Table II). While dasatinib decreased the proportion of MDA-MB-231 cells in S-phase and increased those in the G0/G1 phase, it did not significantly affect the proportion of MDA-MB-231-JYJ cells in any phase of the cell cycle.
Figure 4.
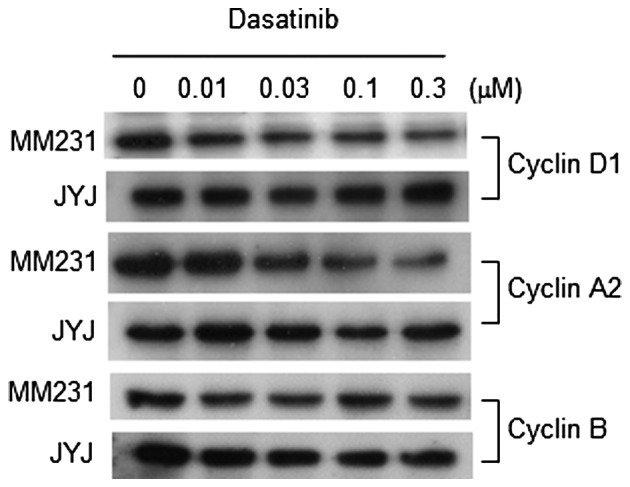
Expression of cyclin proteins following treatment with dasatinib in MDA-MB-231 and MDA-MB-231-JYJ breast cancer cells. Cells were incubated for 24 h in the presence of dasatinib at the indicated concentrations. Cell lysates were subjected to western blotting using antibodies specific to cyclins D1, A2, and B. MM231, MDA-MB-231 cells; JYJ, MDA-MB-231-JYJ cells.
Table II.
Proportion of MDA-MB-231 and MDA-MB-231-JYJ breast cancer cells at each phase of the cell cycle following treatment with dasatinib.
| MDA-MB-231 | MDA-MB-231-JYJ | |||||
|---|---|---|---|---|---|---|
| Dasatinib (µM) | G0/G1 | S | G2/M | G0/G1 | S | G2/M |
| 0 | 64.2 | 7.3 | 22.8 | 49.0 | 10.3 | 23.1 |
| 0.03 | 71.5 | 3.7 | 19.3 | 46.6 | 9.7 | 24.6 |
| 0.1 | 68.3 | 3.1 | 20.8 | 44.3 | 10.3 | 25.3 |
| 0.3 | 74.4 | 2.8 | 19.5 | 40.1 | 10.2 | 27.5 |
Cells were treated with the indicated concentrations of dasatinib for 24 h and then subjected to flow cytometry.
Discussion
It has been reported previously that a tumor contains a variety of cancer cells that are phenotypically, genetically, and functionally different from each other (13,14). Cancer cells in a cell line in vitro are more homogeneous compared with those in a tumor, but they still maintain a certain level of heterogeneity (15,16). This is one reason why a tumor and cancer cells in culture respond differently to an anticancer agent. Since breast cancer also contains many types of stromal and cancer cells, it is difficult to understand the underlying mechanism in a tumor treated with an anticancer agent. A breast cancer cell line may be an easy and simple model to investigate what happens in cancer cells when they are treated with an anticancer agent.
Although paclitaxel has long been used to treat patients with breast cancer, its use is limited due to several significant toxicities such as bone marrow suppression, peripheral neuropathy, and cardiac disturbances (17). Paclitaxel is usually administered by intravenous injection to inhibit a sudden increase in its plasma concentration and to maintain the plasma concentration at an appropriate range for a certain period of time. Since a tumor contains a heterogeneous population of cancer cells, there is a possibility that the tumors in patients treated with paclitaxel include partially resistant or resistant cancer cells. When MDA-MB-231 cells were treated with 30 nM paclitaxel for 6 days, the majority of the cells died, however, a few cells survived that had different morphologies to the parental cells. Among these, the majority were round cells and appeared highly proliferative (Fig. 1B). A small number of large and flat cells were also observed, but their proliferation rate seemed very slow. It is believed that this kind of event, evoked by paclitaxel, may occur more frequently in vivo due to the fact that a tumor is formed with a complex structure of different types of cells and tissues, which provides cancer cells with a protective environment. Although the concentration of paclitaxel treated in vitro was much lower than that of its plasma concentration in patients (80–280 nM), it is thought that the intracellular concentration of paclitaxel accumulated in MDA-MB-231 cells in vitro and in a tumor are similar, in view of a study conducted by Zasadil et al (7).
Since MDA-MB-231-JYJ cells, the small round cells that survived treatment with paclitaxel, were much more proliferative and tumorigenic than MDA-MB-231 cells (Fig. 1C and D), the possibility of that these cells had acquired resistance to paclitaxel was tested. In addition, there were great differences in signal transduction pathways between MDA-MB-231 and MDA-MB-231-JYJ cells (Fig. 2A and B). Signal transduction molecules involved in the proliferation, survival, angiogenesis, and metastasis of cancer, such as c-Src and c-Myc (18,19) were phosphorylated at greater levels in MDA-MB-231-JYJ cells compared to MDA-MB-231 cells. While the expression and cleavage of Notch 1, which are involved in the malignancy or stemness of cancer cells (12,20), were either absent or undetectable in MDA-MB-231 cells, their expression was elevated and they were activated in MDA-MB-231-JYJ cells. In addition to Notch 1, other intracellular molecules that participate in and regulate the stemness of cells, such as Sox2, Oct3/4, c-Myc, Nanog, and E-cadherin were highly expressed in MDA-MB-231-JYJ cells. These results suggest that MDA-MB-231-JYJ cells are more malignant than their parental cell line and appear to possess some characteristics of cancer stem/precursor cells. In contrast to the response of the signal transduction molecules described above, the level of phosphorylation of Akt and Erk1/2, which are known to promote cell survival and proliferation (21,22), were low in MDA-MB-231-JYJ cells compared with MDA-MB-231 cells. Further study is required, however over-activation of other signal transduction molecules involved in cell survival and proliferation such as c-Src, c-Myc, and Notch 1, may circumvent the effect of the lower level of activation of Akt and Erk1/2.
The GI50 concentrations of several anticancer agents including paclitaxel and dasatinib were unexpected (Table I). Although MDA-MB-231-JYJ cells were isolated and cloned from MDA-MB-231 cells that survived paclitaxel treatment, the GI50 concentration (0.021 µM) for paclitaxel was only slightly increased compared with that of the MDA-MB-231 cell line (0.008 µM). This may be because MDA-MB-231-JYJ cells are a subpopulation of MDA-MB-231 cells that were only partially resistant to paclitaxel. Recent studies regarding cancer stem cells have shown that cancer contains several subpopulations of cells, which express different surface and intracellular markers (23–25). Many cancer patients, if not all, eventually succumb to pan-resistant cancer (26). However, partial resistance is also a problem in the treatment of primary cancer, as a proportion of cancer cells that are slightly resistant to an anticancer agent may escape the activity of anticancer agents, survive, and proliferate in the body. There is a possibility that cells similar to MDA-MB-231-JYJ may arise from the treatment of paclitaxel, owing to their partial resistance. This type of phenomenon has previously been observed with other cancer cell lines. When LNCaP, a human prostatic adenocarcinoma cell line, was exposed to paclitaxel, round cells appeared together with large cells and multinucleated cells (27). The round cells were observed with optical and electron microscopy, but were not cloned and studied further for changes in transduction pathways.
Dasatinib is an oral dual Bcr-Abl and Src family inhibitor approved for use in patients with chronic myelogeneous leukemia and Philadelphia chromosome-positive acute lymphoblastic leukemia (28,29). MDA-MB-231-JYJ cells only exhibited resistance to dasatinib among the six anticancer agents tested in the present study. These results raised concern regarding the combined use of dasatinib and paclitaxel, or similar drugs, in patients with breast or other types of cancer. Since many clinical trials using dasatinib are currently being conducted or planned, elucidation of the mechanism by which MDA-MB-231-JYJ cells become resistant to dasatinib will help clinical trial researchers to design their studies to minimize the chances of resistance arising. As shown in Fig. 3, phosphorylation of c-Src in MDA-MB-231 cells was effectively inhibited at low concentrations of dasatinib, however, inhibition of phosphorylation of c-Src in MDA-MB-231-JYJ cells occurred at higher concentrations of dasatinib. The cleavage of Notch 1 and the expression of c-Myc did not decrease in the treated concentration range of dasatinib. This may explain, at least in part, why MDA-MB-231-JYJ cells were resistant to dasatinib.
In addition to the effects of dasatinib on the signal transduction molecules involved in the proliferation, survival, and stemness of cancer cells, the effect of dasatinib on the cell cycle itself or on the cellular molecules related to the cell cycle was also different between MDA-MB-231 and MDA-MB-231-JYJ cells (Table II and Fig. 4). MDA-MB-231-JYJ cells had a higher proportion of cells in the S- and G2/M-phases, meaning that they were more proliferative, and were not affected by dasatinib treatment, with the cell cycle continuously and actively proceeding even in its presence. The results regarding the cell cycle may, at least partly, be explained by the fact that the levels of cyclin D1, A2, and B in MDA-MB-231-JYJ cells were not affected by the presence of dasatinib.
In conclusion, MDA-MB-231-JYJ cells were round, highly proliferative, and tumorigenic. They were resistant to dasatinib but remained susceptible to paclitaxel. The resistance of certain signaling molecules and cyclins in MDA-MB-231-JYJ cells to the inhibitory activity of dasatinib may be the reason why these cells are resistant to dasatinib. Signal transduction molecules related to the malignancy or stemness of cancer cells were also more actively expressed and activated. Further study is required in order to elucidate whether MDA-MB-231-JYJ cells are genetically or epigenetically different from their parental line, and whether the changes in the response of signal transduction molecules are somehow manageable. However, cancer cells may become resistant to dasatinib during the process of paclitaxel therapy, and this must be taken into consideration by researchers and medical practitioners.
Acknowledgements
The present study was supported by the Basic Science Research Program through the National Research Foundation of Korea, funded by the Ministry of Education (grant no. 2014R1A1A2059237) and by a grant from the Korea Research Institute of Bioscience and Biology research initiative program.
References
- 1.Torre LA, Bray F, Siegel RL, Ferlay J, Lortet-Tieulent J, Jemal A. Global cancer statistics, 2012. CA Cancer J Clin. 2015;65:87–108. doi: 10.3322/caac.21262. [DOI] [PubMed] [Google Scholar]
- 2.Crown J, O'Leary M, Ooi WS. Docetaxel and paclitaxel in the treatment of breast cancer: A review of clinical experience. Oncologist. 2004;9(Suppl 2):S24–S32. doi: 10.1634/theoncologist.9-suppl_2-24. [DOI] [PubMed] [Google Scholar]
- 3.Gradishar WJ. Taxanes for the treatment of metastatic breast cancer. Breast Cancer (Auckl) 2012;6:159–171. doi: 10.4137/BCBCR.S8205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Galletti E, Magnani M, Renzulli ML, Botta M. Paclitaxel and docetaxel resistance: Molecular mechanisms and development of new generation taxanes. ChemMedChem. 2007;2:920–942. doi: 10.1002/cmdc.200600308. [DOI] [PubMed] [Google Scholar]
- 5.Krishna R, Mayer LD. Multidrug resistance (MDR) in cancer. Mechanisms, reversal using modulators of MDR and the role of MDR modulators in influencing the pharmacokinetics of anticancer drugs. Eur J Pharm Sci. 2000;11:265–283. doi: 10.1016/S0928-0987(00)00114-7. [DOI] [PubMed] [Google Scholar]
- 6.Orr GA, Verdier-Pinard P, McDaid H, Horwitz SB. Mechanisms of Taxol resistance related to microtubules. Oncogene. 2003;22:7280–7295. doi: 10.1038/sj.onc.1206934. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Zasadil LM, Andersen KA, Yeum D, Rocque GB, Wilke LG, Tevaarwerk AJ, Raines RT, Burkard ME, Weaver BA. Cytotoxicity of paclitaxel in breast cancer is due to chromosome missegregation on multipolar spindles. Sci Transl Med. 2014;6:229ra43. doi: 10.1126/scitranslmed.3007965. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Kim HM, Lim J, Park SK, Kang JS, Lee K, Lee CW, Lee KH, Yun MJ, Yang KH, Han G, et al. Antitumor activity of cytokine-induced killer cells against human lung cancer. Int Immunopharmacol. 2007;7:1802–1807. doi: 10.1016/j.intimp.2007.08.016. [DOI] [PubMed] [Google Scholar]
- 9.Kang MR, Park SK, Lee CW, Cho IJ, Jo YN, Yang JW, Kim JA, Yun J, Lee KH, Kwon HJ, et al. Widdrol induces apoptosis via activation of AMP-activated protein kinase in colon cancer cells. Oncol Rep. 2012;27:1407–1412. doi: 10.3892/or.2012.1644. [DOI] [PubMed] [Google Scholar]
- 10.Krishan A. Rapid flow cytofluorometric analysis of mammalian cell cycle by propidium iodide staining. J Cell Biol. 1975;66:188–193. doi: 10.1083/jcb.66.1.188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kang MR, Kang JS, Yang JW, Kim BG, Kim JA, Jo YN, Lee K, Lee CW, Lee KH, Yun J, et al. Gene expression profiling of KBH-A42, a novel histone deacetylase inhibitor, in human leukemia and bladder cancer cell lines. Oncol Lett. 2012;3:113–118. doi: 10.3892/ol.2011.430. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Li L, Zhao F, Lu J, Li T, Yang H, Wu C, Liu Y. Notch-1 signaling promotes the malignant features of human breast cancer through NF-kB activation. PLoS One. 2014;9:e95912. doi: 10.1371/journal.pone.0095912. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 13.Meacham CE, Morrison SJ. Tumour heterogeneity and cancer cell plasticity. Nature. 2013;501:328–337. doi: 10.1038/nature12624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Burrell RA, McGranahan N, Bartek J, Swanton C. The causes and consequences of genetic heterogeneity in cancer evolution. Nature. 2013;501:338–345. doi: 10.1038/nature12625. [DOI] [PubMed] [Google Scholar]
- 15.Croce MV, Colussi AG, Price MR, Segal-Eiras A. Identification and characterization of different subpopulations in a human lung adenocarcinoma cell line (A549) Pathol Oncol Res. 1999;5:197–204. doi: 10.1053/paor.1999.0212. [DOI] [PubMed] [Google Scholar]
- 16.Stockholm D, Benchaouir R, Picot J, Rameau P, Neildez TM, Landini G, Laplace-Builhé C, Paldi A. The origin of phenotypic heterogeneity in a clonal cell population in vitro. PLoS One. 2007;2:e394. doi: 10.1371/journal.pone.0000394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Marupudi NI, Han JE, Li KW, Renard VM, Tyler BM, Brem H. Paclitaxel: A review of adverse toxicities and novel delivery strategies. Expert Opin Drug Saf. 2007;6:609–621. doi: 10.1517/14740338.6.5.609. [DOI] [PubMed] [Google Scholar]
- 18.Gherardi E, Birchmeier W, Birchmeier C, Vande Woude G. Targeting MET in cancer: Rationale and progress. Nat Rev Cancer. 2012;12:89–103. doi: 10.1038/nrc3205. [DOI] [PubMed] [Google Scholar]
- 19.Zhang S, Yu D. Targeting Src family kinases in anti-cancer therapies: Turning promise into triumph. Trends Pharmacol Sci. 2012;33:122–128. doi: 10.1016/j.tips.2011.11.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Li D, Masiero M, Banham AH, Harris AL. The notch ligand JAGGED1 as a target for anti-tumor therapy. Front Oncol. 2014;4:254. doi: 10.3389/fonc.2014.00254. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Lu Z, Xu S. ERK1/2 MAP kinases in cell survival and apoptosis. IUBMB Life. 2006;58:621–631. doi: 10.1080/15216540600957438. [DOI] [PubMed] [Google Scholar]
- 22.Xu N, Lao Y, Zhang Y, Gillespie DA. Akt: A double-edged sword in cell proliferation and genome stability. J Oncol. 2012;2012:951724. doi: 10.1155/2012/951724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Tirino V, Desiderio V, Paino F, De Rosa A, Papaccio F, La Noce M, Laino L, De Francesco F, Papaccio G. Cancer stem cells in solid tumors: An overview and new approaches for their isolation and characterization. FASEB J. 2013;27:13–24. doi: 10.1096/fj.12-218222. [DOI] [PubMed] [Google Scholar]
- 24.Ling GQ, Chen DB, Wang BQ, Zhang LS. Expression of the pluripotency markers Oct3/4, Nanog and Sox2 in human breast cancer cell lines. Oncol Lett. 2012;4:1264–1268. doi: 10.3892/ol.2012.916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Klonisch T, Wiechec E, Hombach-Klonisch S, Ande SR, Wesselborg S, Schulze-Osthoff K, Los M. Cancer stem cell markers in common cancers-therapeutic implications. Trends Mol Med. 2008;14:450–460. doi: 10.1016/j.molmed.2008.08.003. [DOI] [PubMed] [Google Scholar]
- 26.Borst P. Cancer drug pan-resistance: Pumps, cancer stem cells, quiescence, epithelial to mesenchymal transition, blocked cell death pathways, persisters or what? Open Biol. 2012;2:120066. doi: 10.1098/rsob.120066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Röyttä M, Laine KM, Härkönen P. Morphological studies on the effect of taxol on cultured human prostatic cancer cells. Prostate. 1987;11:95–106. doi: 10.1002/pros.2990110112. [DOI] [PubMed] [Google Scholar]
- 28.Talpaz M, Shah NP, Kantarjian H, Donato N, Nicoll J, Paquette R, Cortes J, O'Brien S, Nicaise C, Bleickardt E, et al. Dasatinib in imatinib-resistant Philadelphia chromosome-positive leukemias. N Engl J Med. 2006;354:2531–2541. doi: 10.1056/NEJMoa055229. [DOI] [PubMed] [Google Scholar]
- 29.Steinberg M. Dasatinib: A tyrosine kinase inhibitor for the treatment of chronic myelogenous leukemia and philadelphia chromosome-positive acute lymphoblastic leukemia. Clin Ther. 2007;29:2289–2308. doi: 10.1016/j.clinthera.2007.11.005. [DOI] [PubMed] [Google Scholar]