

Abstract
Heparin binding proteins (HBPs) are produced by accessory glands. These are secreted into the seminal fluid, bind to the spermatozoa at the time of ejaculation, favour capacitation, acrosome reaction, and alter the immune system response toward the sperm. The present study was conducted with an objective to assess the effect of purified seminal plasma-HBPs (SP-HBPs) on cross bred cattle bull sperm attributes during two phases of cryopreservation: Pre freezing and freezing-thawing. SP-HBPs were purified from pooled seminal plasma by heparin affinity chromatography. Three doses of SP-HBPs i.e. 10, 20, 40 μg/mL semen were standardized to find out the optimum dose and 20 μg/mL was found to be an optimum dose. Semen as such and treated with SP-HBPs was diluted with sodium citrate-egg yolk diluter and cryopreserved as per the standard protocol. Sperm parameters i.e. motility, viability, Hypo-osmotic swelling test (HOST), acrosome damage, in vitro capacitation and lipid peroxidation were evaluated in SP-HBP treated and untreated (control) semen at both phases of cryopreservation. A considerable variation in percent sperm motility, viability, membrane integrity (HOST), acrosome damage, acrosome reaction and lipid peroxidation was observed at both phases among the bulls irrespective of the treatment. Incubation of neat semen with 20 μg/mL SP-HBP before processing for cryopreservation enhanced the average motility, viability, membrane integrity by 7.2%, 1.5%, 7.9%, and 5.6%, 6.6%, 7.4% in pre-frozen and frozen-thawed semen in comparison to control. There was also an average increase of 4.1%/3.9% in in vitro capacitation and acrosome reaction in SP-HBPs-treated frozen-thawed semen as compared to control. However, binding of SP-HBPs to the sperm declined acrosome damage and lipid peroxidation by 1.3%/4.1% and 22.1/32.7 μM/109 spermatozoa in SP-HBP treated pre-frozen/frozen-thawed semen as compared to control, respectively. Significant (p<0.05) effects were observed only in motility, HOST and in vitro acrosome reaction. It can be concluded that treatment of neat semen with SP-HBPs before cryopreservation minimized the cryoinjury by decreasing the generation of reactive oxygen species.
Keywords: Seminal Plasma, Heparin Binding Protein [HBP], Cryopreservation, Function Tests, Sperm, Cattle Bulls
INTRODUCTION
Sperm cryopreservation is an important tool for assisted reproductive techniques and is the most efficient way for storing germ plasm (Olaciregui et al., 2014). Cryopreservation has been applied to various species, including humans, swine, cattle, cats, and dogs (Mota Filho et al., 2014). Although sperm cryopreservation preserves sperm motility, metabolic functions, and fertility, but, still the freeze-thawing process causes irreversible damage to spermatozoa and reduces fertility (Ardon and Saurez, 2013). Even though cryopreservation helps in prolonged storage of semen, but cryo-capacitation is one of the major factors associated with reduced longevity and poor survivability of cryopreserved spermatozoa in female reproductive tract (Watson, 2000), resulting in reduced fertility of frozen-thawed semen. Even with optimized protocols, 40% to 50% of the sperms do not survive after cryopreservation due to irreversible damage (Watson, 2000). Several studies have focused on identifying damages during freezing and thawing, tests to screen sperm quality of frozen-thawed semen, evaluation of alternative cryoprotectants and other additives, and freezing procedures to improve sperm viability and fertility. Recently, Yoon et al. (2015) proposed that freezing/thawing steps are the most critical in cryopreservation and may provide a logical ground to understand the process of cryo-damage.
Currently, many diverse proteins are being tested that may increase the fertilization capability of sperm and heparin binding proteins (HBPs) are among these proteins. HBPs have been viewed as favouring capacitation, acrosome reaction, and altering the immune system response toward the sperm. The HBPs are produced by the accessory glands of the male under the control of androgens (Miller et al., 1990) and secreted within the seminal fluid (Nass et al., 1990). The HBPs bind to the epididymal sperm and increase the ability of the acrosome reaction in response to the heparin and other proteins of the zona pellucida (Ax et al., 2002). Absorption of isolated proteins from the seminal plasma can reduce the damage that cold shock can have on the sperm membrane (Barrios et al., 2000). Mogielnicka-Brzozowska and Kordan (2011) mentioned the possibility of adding specific plasma proteins to sperm for the retention of features responsible for the efficient fertilization after storage. HBPs protect sperm from stress of freezing and thawing and maintained intracellular protein homeostasis (Shi et al., 1998). HBPs also protect sperm from lipid peroxidation during cryopreservation (Kumar et al., 2008). Preliminary studies were done by Alvarez-Gallardo et al. (2013) with the recombinant proteins indicated that addition of rFAA and rTIMP-2 to semen before cryopreservation help stabilize the acrosome membrane of the bovine sperm by decreasing post-thaw acrosome damage. Hence the present study was conducted with objective to assess the effect of HBP on sperm function tests at pre-freeze and frozen-thawed phases of cryopreservation.
MATERIALS AND METHODS
Procurement of semen sample
Three ejaculates from each of six healthy cross cattle bulls (Holstein-Fresian×Red Dane×Sahiwal) were procured from GADVASU, Dairy Farm and Semen bank, Bhattian, Khanna, Ludhiana, India. The bulls were kept under loose housing system (covered area, 3.6 m×3.0 m and uncovered area, 7.5 m×3.0 m) and standard feeding schedule along with ad libitum green fodder and water availability and standard management conditions. Semen was collected twice a week through artificial vagina method. None of the bulls selected for this study had any preceding physical abnormalities.
A good quality fresh ejaculates (1 to 2 mL) with minimum initial motility of ≥60%, mass activity of 3+ to 4+, abnormal morphology of ≤25% and sperm concentration of >750×106 sperms/mL of each bull were processed for cryopreservation. Seminal plasma was separated by centrifuging pooled semen of six bulls at 3,000 rpm for 5 minutes and was stored at −20°C till use.
Purification of HBPs by affinity chromatography
Sepharose (Sigma-Aldrich) was activated and heparin was bound as per the protocol of Farooqui (1980). The chromatography glass column (28 mm×70 mm) was packed with heparin-sepharose media up to the mark at the top of the column and allowed to settle for 3 to 4 h. Thereafter, the column was equilibrated for 1 h with 10 mM Tris HCl (pH 7.4). About 1.0 mL of pooled seminal plasma (filtered through 0.22 μ filter) was loaded and circulated through the column for 15 minutes for absorption of HBPs to the heparin bound resins. The non-HBPs were washed out with 10 mM Tris HCl and the HBPs were eluted with 1 M NaCl at a flow rate of 1 mL/min in the tubes racked in a fraction collector. The recovered HBPs fractions were pooled in agreement with the observed curve, obtained from optical density (280 nm), detected by UV monitor, attached to the fraction collector (Manaskova et al., 2002). The pooled fractions of HBPs were concentrated by spinning through protein concentrators (Millipore, Billerica, MA, USA, 3 kDa) and analyzed for protein content (Lowry et al., 1951).
Conjugation of fluorescein isothiocyanate fluorochrome with purified HBP and confirmation of incorporation of SP-HBP to semen
Conjugation of fluorescein isothiocyanate (FITC) fluorochrome to SP-HBPs was done by using a kit (Life technology, Carisbad, CA, USA) as per the protocol provided with the kit. About 20 μg of purified HBPs was added to 1.0 mL of neat semen and incubated at 37°C for 20 minutes. Semen was washed thrice with phosphate buffered saline (PBS), pH 7.4 by centrifugation at 3,000 rpm for 5 min. Pellet was suspended in 1.0 mL of PBS and a drop of semen on a glass slide covered with coverslip was observed at 400× under fluorescence microscope (Olympus, Shinjuku, Tokyo, Japan) using blue filter at 480 nm.
Dilution and cryopreservation of semen
Three doses of SP-HBPs i.e. 10, 20, 40 μg/mL semen were standardized to find out the optimum dose and 20 μg/mL was found to be an optimum dose. Semen was divided into two fractions: one fraction, supplemented with SP-HBP and other as such (control), were incubated at 37°C for 20 min. After incubation, both fractions were diluted with sodium citrate extender. The pre-freezing sample was kept at 4°C in cold handling cabinet for equilibration and aliquot for freezing was filled and sealed by manual filling and sealing method. The filled and sealed straws were cryo-preserved in liquid nitrogen by following standard procedure. Semen at pre-freeze and frozen-thaw phase was analyzed for sperm function (motility, viability and Hypo-osmotic swelling test [HOST]), acrosome damage, acrosome reaction and lipid peroxidation [LPO]) as detailed below.
Individual motility
About 10 μL of semen was placed on a pre-warmed (37°C) glass slide, covered with a cover slip and placed on the pre-warmed stage (Magnus Mumbai, Maharashtra, India) attached to a phase contract microscope (Olympus CX21FS1, Japan). Linear progressive motility was assessed by subjective method viewing 10 fields with the aid of closed-circuit television attached to the microscope (400×). A total of 200 spermatozoa were observed under phase contrast microscope (Olympus, Japan) at 400× in different fields and percentage of motile spermatozoa was calculated.
Sperm viability
Sperm viability was assessed by Syber green/propium iodide (Life technologies staining kit, USA). A total of 200 live (green) and dead (red) spermatozoa were counted in different fields and percentage of viability was calculated (Figure 1).
Figure 1.
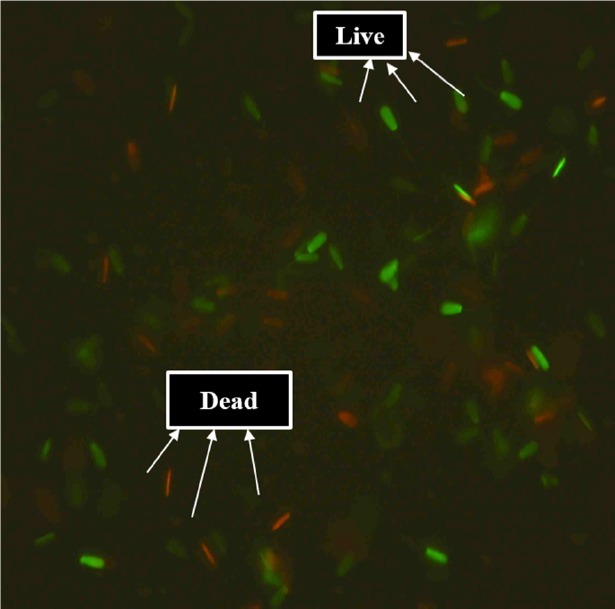
Cattle bull spermatozoa stained with syber green/propidium iodide (400×).
Hypo-osmotic swelling test
Briefly, 10 μL of semen was incubated in 100 μL of hypo-osmotic solution at 37°C for 30 min. A total of 150 spermatozoa with coiled and un-coiled tails were counted in different fields (Jeyendran et al., 1984). A control was also run in PBS (pH 7.4). The number of spermatozoa with coiled tailed/swollen heads in PBS was deducted from the number in hypo-osmotic solution and the resultant figure was taken as the Hypo-osmotic swelling (HOS)-reactive spermatozoa.
Acrosomal damage
Acrosome damage was assessed by chlortetracycline cysteine stain (CTC, Figure 2) (Ward and Storey, 1984). Briefly 25 μL of sperm suspension was mixed with 25 μL of 750 mM CTC in 20 mM Tris-HCL, 130 mM NaCl and 5 mM cystein, incubated at 37°C for 30 min. After adding 100 μL of 12.5% paraformaldehyde and one drop of 0.22 M 1, 4-Diazabicyclo[2.2.2]octane, a drop of sperm suspension was observed under fluorescence microscope (Olympus, Japan) at 400× using blue filter at 480 nm. At least 200 spermatozoa with normal (whole sperm head with bright fluorescence) and cryo capacitated sperm (acrosome-intact sperm with fluorescence ring at equatorial segment) and acrosome-reacted (sperm with fluorescence on postacrosomal region) were counted in different fields at 400× and percentage of cryo-capacitated and acrosome-reacted spermatozoa was calculated (Figure 2).
Figure 2.
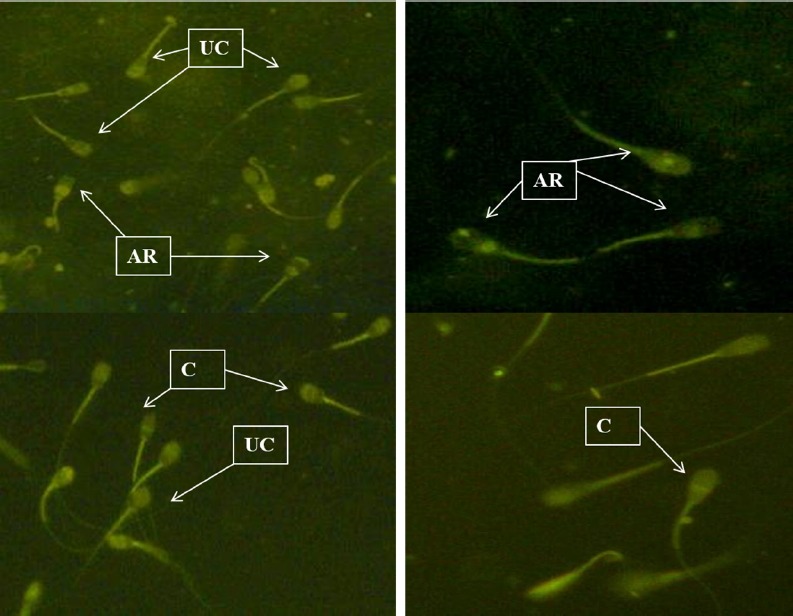
CTC staining of acrosome damage and in vitro capacitated/acrosome reacted cattle bull spermatozoa (400×). CTC, chlortetracycline cysteine; UC, uncapacitated; C, capacitated; AR, acrosome reacted.
In vitro capacitation/acrosome reaction
Frozen-thawed spermatozoa washed twice with tyrode albumin lactate pyruvate medium (TALP, 92.9 mM NaCl, 4 mM KCl, 25.9 mM NaHCO3, Na2HPO4, 10 mM CaCl2·2H2O, 0.5 mM MgCl2·6H2O, 1.3 mM sodium pyruvate, 7.6 mM sodium lactate and 20 mM 4-2-hydroxyethyl-1-piperazineethanesulfonic acid, pH 7.4 were subjected to swim up and only motile spermatozoa were incubated in TALP supplemented with 0.6% bovine serum albumin, 5 mM glucose and 165 units of heparin for 4 hrs at 37°C in an incubator (Fraser et al., 1995). Capacitation/acrosome reaction was assessed by counting 200 spermatozoa in CTC stained slides under fluorescence microscope at 400× (Olympus, Japan).
Lipid peroxidation
Lipid peroxidation was estimated by the end point generation of malondialdehyde (MDA) determined by the thiobarbituric acid (TBA) test (Buege and Steven, 1978). Briefly, extended spermatozoa ([40 to 70]×106 cells) in 0.5 mL of cold 10% (wt/vol) chilled trichloroacetic acid to proliferate proteins. The precipitate was pelleted by centrifugation (6,000 rpm for 10 min) and 1 mL of 0.375% (wt/vol) TBA was added to the supernatant and kept in a boiling water bath at 100°C for 10 minutes. After cooling, the absorbance was read at 532 nm (UV-VIS spectrophotometer, systronics, Ahmedabad, Gujrat, India).
MDA production was calculated by the formula:
*Molar extinction coefficient = (1.56×105 M-1C-1)
Statistical analysis
The data is presented as mean±standard error of the mean and analyses were performed with CPCS1 program package. One way analysis of variance was applied to evaluate the level of significance at 5% between treatments.
RESULTS AND DISCUSSION
Purification of HBPs from seminal plasma and verification of binding of SP-HBP to spermatozoa
Graphic image of HBP purified from pooled seminal plasma of cross-bred bulls is shown in Figure 3, which indicated two peaks of unbound (non-HBP) and bound proteins (HBP). Addition of 20 μg/mL of SP-HBP to neat semen and incubation at 37°C for 20 min before cryopreservation resulted in its binding mainly to head and mid piece. But binding was more on post-acrosomal cap in majority of the spermatozoa (Figure 4). A study done by Cheema et al. (2015) on natural localization of HBPs in cross-bred bull spermatozoa using anti-AZU-1 revealed that these proteins are mainly localized on acrosomal cap.
Figure 3.
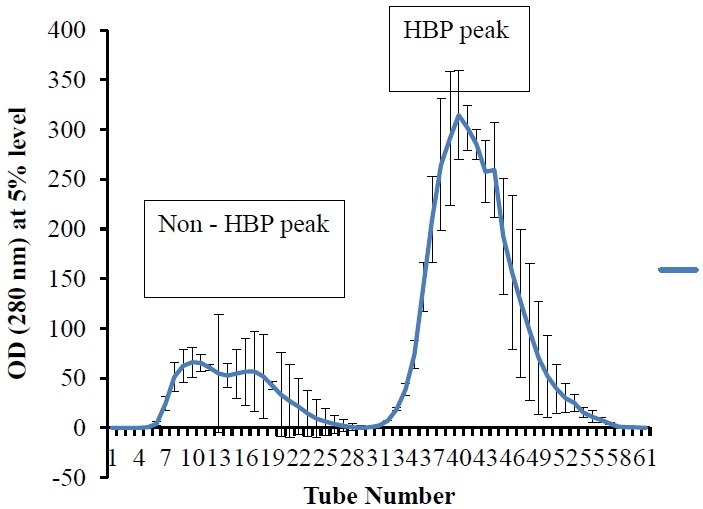
Graphic image of heparin-binding and non-heparin-binding proteins in seminal plasma of cross bred cattle bulls separated on heparin-affinity chromatography.
Figure 4.
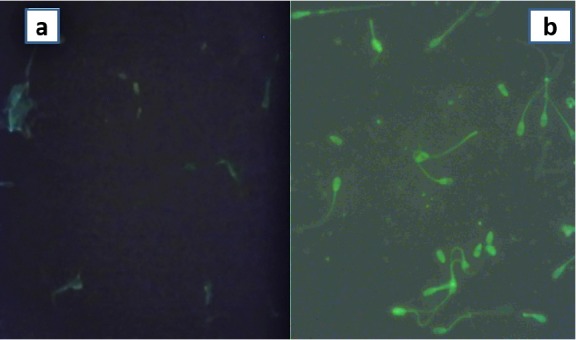
Visualization of binding of SP-HBP to spermatozoa (400×). (a) control (Semen incubated without SP-HBP), and (b) treated (Neat semen incubated with SP-HBP). SP-HBP, seminal plasma-heparin binding protein.
Effect of SP-HBP on sperm function tests
Standardization of dose of SP-HBP
Effects of different doses of SP-HBP on percent motility, viability, HOST, acrosome damage, in vitro capacitation and LPO in pre-freeze and frozen-thaw semen are given in Table 1. Individual motility (%), viability (%), HOST (%) were found to be higher in pre-freeze and post-thaw semen supplemented with 20 μg/mL of SP-HBP as compared to 0, 10, and 40 μg of SP-HBP. Percentage of in vitro capacitated/acrosome reacted spermatozoa was higher only in frozen-thawed semen supplemented with 20 μg/mL SP-HBP as compared to 0, 10, and 40 μg. Acrosome damage (%) and MDA (μM/109 spermatozoa) showed reduction in semen supplemented with 20 μg/mL of SP-HBP in comparison to 0, 10, and 40 μg SP-HBP at both phases of cryopreservation. Increase in motility, viability, in vitro acrosome reaction and decrease in LPO (MDA) in semen supplemented with SP-HBP was statistical significant (p<0.05). Manjunath et al. (2002) also suggested that at a greater concentration of bovine seminal plasma proteins, more cholesterol and phospholipids are removed from the sperm membrane, resulting in its destabilization that decreases sperm resistance to cold shock and freezing. Harshan et al. (2006) also postulated that higher loss of semen parameters with higher amount of HBP could be due to an increased cholesterol efflux in the spermatozoa which led to more pronounced cryo-damage. Therefore, effects of 20 μg/mL of SP-HBP on semen cryopreservation of six bulls were evaluated.
Table 1.
Effect of different doses of affinity purified SP-HBP on sperm function and fertility tests during pre-freeze and frozen-thaw phases of cryopreservation
| Sperm parameter | Pre-freeze | Frozen-thaw | ||||||
|---|---|---|---|---|---|---|---|---|
|
|
|
|||||||
| C1 | 10 | 20 | 40 | C | 10 | 20 | 40 | |
| Motility (%) | 73.3±1.7a | 76.7±1.7a | 83.3± 1.7b | 75.0±0.0a | 45.0±2.9c | 46.7±1.7c | 58.3±4.4d | 48.3±4.4 c |
| Viability (%) | 68.7±3.6ab | 61.2±1.2a | 76.2±3.5a b | 72.4±3.1 b | 48.7±8.7c | 47.8±6.2c | 56.0±7.6c | 52.4±8.2c |
| HOST (%) | 35.3±2.4a | 38.2±4.3a | 42.1±4.1a | 35.72±2.2a | 35.0±2.2a | 12.1±2.4b | 18.6±3.9b | 14.4±3.0b |
| Acrosome damage (%) | ||||||||
| Cryo-Cap | 22.9±2.6a | 22.1±1.6a | 23.3±1.5a | 24.5±1.8a | 29.2±2.9b | 28.1±3.4b | 28.4±1.7b | 30.1±2.4b |
| Cryo-AR | 12.1±4.5a | 10.1±3.8a | 10.5±4.4a | 11.5±4.9a | 16.3±4.3a | 13.8±2.8a | 14.6±3.7a | 17.0±4.2a |
| In vitro capacitation/acrosome reaction (%) | ||||||||
| Cap | - | - | - | - | 39.1±3.4a | 39.4±2.8a | 44.4±3.0a | 42.5±3.2a |
| AR | - | - | - | - | 27.9±5.8a | 30.1±6.1ab | 34.0±5.1b | 33.4±5.0 b |
| LPO (MDA, μM/109 spermatozoa) | 14.7±5.7a | 12.4±3.8a | 11.9±3.6a | 14.5±4.6a | 50.3±2.8b | 44.3±4.1bc | 40.3±2.6c | 45.3±4.9bc |
HOST, hypo-osmotic swelling test; Cap, capacitated; AR, acrosome reacted; LPO, lipid peroxidation; MDA, malondialdehyde.
C, control (0), 10, 20, 40 (Dose of HBP in μg/mL).
Superscripts (a, b, ab, bc) indicate the difference at 5% level of significance within the rows.
Effect of 20 μg SP-HBP on sperm function and fertility tests of six bulls
Motility, Viability and HOST
Mean percentage of motile, live and HOS responsive spermatozoa showed a variation among six cross breed bulls, which ranged from 61.7%±1.7% to 70±2.9%; 55.7%±2.8% to 73.0%±6.6%; 16.0%±0.7% to 36.1%±8.0%, and 41.7%±4.4% to 58.3%±6.7%; 38.2%±3.1% to 59.5%±2.2%; 8.8%±4.6% to 27.5%±7.5% at the time of pre-freezing and freezing-thawing, respectively (Table 2). Decrease observed in percent motility, viability and HOST from pre-freeze to post thaw phase of cryopreservation was significant (p<0.05). Lessard et al. (2000) were of the opinion that sperm viability decrease by 50%, whereas, fertilizing capacity is affected by a factor of sevenfold after cryopreservation. However, incorporation of SP-HBP to the sperm surface resulted in an increase of 5.0% to 10.0%; 3.6% to 7.8%; 4.8% to 22.1%, and 1.6% to 8.3%; 4.2% to 11.0%; 3.0% to 12.0% in motile, live and HOS positive spermatozoa at pre-freeze and frozen-thaw phase of cryo-preservation as compared to control, respectively. However, increase in only motility and HOST due to supplementation of SP-HBP was significant (p<0.05) at both phases of cryopreservation. Harshan et al. (2006) revealed that 40 μg dose of HBP improved the motility, HOST and cervical mucus penetration test at pre-freeze phase of buffalo semen cryopreservation, but they did not observe positive effect of this dose on frozen-thawed semen. Barrios et al. (2000) also demonstrated that the absorption of isolated proteins from seminal plasma may reduce the cold shock effect.
Table 2.
Effect of affinity purified SP-HBP on sperm motility, viability and HOST (mean±SE) during pre-freeze and frozen-thaw phases of cryopreservation
| Bull number | Motility (%) | Viability | HOST | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
|
|
||||||||||
| Pre-freeze | Frozen-thawed | Pre-freeze | Frozen-thawed | Pre-freeze | Frozen-thawed | |||||||
|
|
|
|
|
|
|
|||||||
| C1 | T1 | C | T | C | T | C | T | C | T | C | T | |
| 1 | 70.0±0.0 | 80.0±0.0 | 58.3±3.4 | 66.6±2.9 | 73.0±6.6 | 79.6±4.5 | 59.5±2.2 | 64.8±2.7 | 30.6±3.7 | 36.2±3.3 | 18.1±3.2 | 25.1±4.9 |
| 2 | 70.0±2.9 | 78.3±3.3 | 55.0±2.9 | 61.7±1.7 | 69.1±7.5 | 76.9±5.8 | 54.9±7.1 | 61.6±6.0 | 32.5±8.1 | 54.6±2.5 | 16.9±5.9 | 27.8±4.8 |
| 3 | 68.3±4.4 | 73.3±4.4 | 50.0±5.8 | 56.6±4.4 | 65.9±3.8 | 71.0±3.6 | 50.4±8.8 | 56.4±6.1 | 36.1±8.0 | 46.3±6.2 | 27.5±7.5 | 35.3±5.7 |
| 4 | 61.7±1.7 | 70.0±2.9 | 45.0±2.9 | 50.0±2.9 | 61.3±2.1 | 67.0±1.0 | 43.0±7.2 | 47.2±7.2 | 25.3±5.1 | 30.1±3.3 | 17.7±1.7 | 20.7±2.1 |
| 5 | 63.3±3.3 | 68.3±3.3 | 41.7±4.4 | 43.3±3.3 | 55.7±2.8 | 59.3±3.3 | 38.2±3.1 | 44.9±0.2 | 16.0±0.7 | 22.0±2.6 | 8.8±4.6 | 12.6±3.2 |
| 6 | 65.0±2.9 | 71.7±1.7 | 46.7±6.0 | 51.7±6.0 | 65.1±0.5 | 69.3±1.8 | 48.5±1.4 | 59.5±0.7 | 21.6±4.6 | 32.5±4.0 | 12.6±2.0 | 24.6±1.5 |
| Combination factor mean | 66.4a | 73.6 b | 49.4c | 55.0d | 65.0a | 70.5a | 49.1b | 55.7b | 27.0a | 36.9b | 16.9c | 24.3a |
SP-HBP, seminal plasma-heparin binding protein; HOST, hypo-osmotic swelling test; SE, standard error.
C, control without HBP; T, supplemented with SP-HBP.
Superscripts a, b, c, d indicate the difference at 5% level of significance within the columns.
Acrosome damage
Two stages of acrosome damage were revealed by CTC staining i.e. partial (cryo-capacitated) and complete (cryo-acrosome reacted). These two phases of sperm due to cryo-injury are demonstrated as premature capacitation and spontaneous acrosome reaction (Srivastava et al., 2013). Mean percentage of cryo-capacitated and acrosome reacted spermatozoa showed a variation among the bulls and ranged from 20.1%±1.1% to 30.6%±1.5% and 5.3%±0.3% to 14.3%±4.1% at pre-freeze phase, which was further significantly (p<0.05) increased to 25.6%±4.5% to 37.5%±0.7% and 10.9%±1.3% to 21.1%±0.5% in frozen-thaw phase of cryopreservation in six bulls, respectively (Table 3). Altered mitochondrial function, reduction in motility and failure of chromatin condensation, all of which influence the viability and fertility of sperm have also been reported due to induction of pre-mature acrosome reaction (Watson, 2000; Chaveiro et al., 2006). Acrosomal cap is of utmost importance for fertilization and is related to fertility. Spermatozoa with higher motility cannot be fertile due to acrosomal loss (Raval and Dhami, 2010).
Table 3.
Effect of affinity purified SP-HBP on acrosome damage (mean±SE) during pre-freeze and frozen-thaw phase of cryopreservation
| Bull number | Pre-freeze | Frozen-thawed | ||||||
|---|---|---|---|---|---|---|---|---|
|
|
|
|||||||
| Control1 | Treated1 | Control | Treated | |||||
|
|
|
|
|
|||||
| Capacitated | Acrosome reacted | Capacitated | Acrosome reacted | Capacitated | Acrosome reacted | Capacitated | Acrosome reacted | |
| 1 | 20.2±3.4 | 14.3±4.1 | 21.2±3.6 | 11.7±3.7 | 25.6±4.5 | 16.8±3.6 | 23.6±3.4 | 15.8±3.5 |
| 2 | 25.1±3.8 | 9.6±2.1 | 25.2±3.7 | 8.5±2.8 | 30.8±3.3 | 13.3±2.0 | 21.3±0.8 | 4.9±0.6 |
| 3 | 20.1±1.1 | 5.3±0.3 | 20.3±0.8 | 4.9±0.6 | 26.1±1.4 | 10.9±1.3 | 26.3±1.4 | 10.1±0.8 |
| 4 | 22.9±1.0 | 6.6±0.1 | 24.4±0.7 | 5.2±0.1 | 32.9±0.8 | 14.0±3.1 | 33.9±0.4 | 12.4±2.6 |
| 5 | 30.6±1.5 | 9.0±0.4 | 29.6±0.9 | 10.8±0.7 | 37.5±0.7 | 13.5±2.0 | 35.0±1.2 | 15.0±2.0 |
| 6 | 21.5±1.0 | 14.1±2.1 | 22.1±0.5 | 12.2±2.6 | 29.5±0.6 | 21.1±0.5 | 29.6±0.2 | 19.8±1.4 |
| Combination factor mean | 23.4a | 9.8a | 23.08 a | 8.8a | 30.4b | 14.9bc | 28.2bc | 13.0c |
SP-HBP, seminal plasma-heparin binding protein; SE, standard error.
Control, without HBP); Treated, supplemented with HBP.
Superscripts a, b, bc, c indicates the difference at 5% level of significance within column.
Acrosome is required to undergo biochemical changes in the female reproductive tract before fertilization and premature loss of acrosome and hence loss of enzymes will affect the fertilization process. Supplementation of SP-HBP to cattle bull semen resulted in a non-significant (p<0.05) reduction of 1.0% to 1.4% and 0.6% to 17.9% in acrosome damage at pre-freeze and frozen-thaw phase, respectively. Our study revealed a reduction in cold shock effect on treating the spermatozoa with SP-HBP. Result of a study by Mogielnicka-Brzozowska et al. (2011) also indicated that zona binding proteins of boar seminal plasma have a shielding effect on the plasma membrane and the acrosome of spermatozoa by protecting their structures against the damage caused by cold shock. Mogielnicka-Brzozowska and Kordan (2011) mentioned that addition of specific plasma protein to sperm probably may retain the features responsible for the efficient fertilization after storage.
In vitro capacitation and acrosome reaction
A variation in response of frozen-thawed spermatozoa to in vitro capacitation and acrosome reaction was also observed irrespective of treatment among the six bulls. Incubation of frozen thawed spermatozoa of six bulls in TALP energy medium resulted in 36.6%±2.1% to 45.6%±1.8% capacitated and 23.9%±2.2% to 31.7%±1.7% acrosome reacted spermatozoa (Table 4). Incorporation of SP-HBP enhanced the percentage of capacitated (41.9%±1.7% to 48.5%±0.2%) and acrosome reacted (26.6%±2.5% to 33.5%±5.4%) spermatozoa in frozen-thawed semen of six bulls. It indicated a significant (p<0.05) increase of 6.2% to 11.4% in capacitated/acrosome reacted spermatozoa upon binding of SP-HBP to the sperm membrane. Arangasamy (2010) performed in vitro capacitation of stallion spermatozoa in the presence of SP-HBP/heparin and found significant difference in treated and control groups. Heparin and heparin like molecules are known to effectively capacitate bovine sperm. High density lipoproteins (HDL) have also been shown to capacitate bull sperm (Therein et al., 1998) and HDL by binding to the bovine seminal plasma proteins (Manjunath et al., 1988). Addition of purified fraction of 15 to 17 kDa, 24 kDa, and 31 kDa HBPs (Miller et al., 1990) to epididymal sperm revealed the effect of heparin. They further conveyed that fraction containing 24 and 31 kDa proteins possessed greatest activity/unit of protein to induce acrosome exocytosis.
Table 4.
Effect of affinity purified SP-HBP on in vitro capacitation/acrosome reaction (mean±SE) during pre-freeze and frozen-thaw phase of cryopreservation
| Bull number | Control | Treated | ||
|---|---|---|---|---|
|
|
|
|||
| Capacitated | Acrosome reacted | Capacitated | Acrosome reacted | |
| 1 | 41.7±1.6 | 28.3±5.7 | 44.1±1.6 | 33.5±5.4 |
| 2 | 40.9±0.4 | 24.4±2.2 | 46.9±0.6 | 26.6±2.5 |
| 3 | 36.6±2.1 | 23.9±2.2 | 41.9±1.7 | 30.0±2.5 |
| 4 | 45.6±1.8 | 25.0±3.3 | 47.1±0.5 | 30.4±2.9 |
| 5 | 43.0±0.4 | 27.0±2.9 | 48.5±0.2 | 27.7±2.1 |
| 6 | 40.0±2.0 | 31.7±1.7 | 44.3±0.5 | 32.9±3.7 |
| Average±SE | 41.4a | 26.3b | 45.5c | 30.2d |
SP-HBP, seminal plasma-heparin binding protein; SE, standard error.
Control, without HBP; Treated, supplemented with SP-HBP.
Superscripts a, b, c, d indicate the difference at 5% level of significance within the columns.
Lipid peroxidation
An increase of 35.8 to 108.1 μmoles of MDA/109 spermatozoa was noticed from pre-freeze to freezing-thawing process of cryopreservation (Table 5). MDA has been used in biochemical assays to monitor the degree of peroxidative damage in spermatozoa and exhibited an excellent correlation with the degree to which sperm function is impaired in terms of motility and the capacity for sperm-oocyte fusion (Sidhu et al., 1998). Increase in MDA production due to freezing-thawing was non- significantly (p>0.05) less (23.1 to 86.9 μmoles/109 spermatozoa) in semen supplemented with SP-HBP in comparison to control.
Table 5.
Effect of affinity purified SP-HBP on MDA production (μM/109 spermatozoa, mean ± SE) during pre-freeze and frozen-thaw phase of cryopreservation
| Bull number | Pre-freeze | Frozen-thaw | ||
|---|---|---|---|---|
|
|
|
|||
| Control1 | Treated1 | Control | Treated | |
| 1 | 73.5±51.3 | 51.6±35.0 | 112.4±38.0 | 94.9±37.8 |
| 2 | 46.8±19.4 | 24.9±12.8 | 83.3±19.4 | 60.3±15.1 |
| 3 | 22.1±10.3 | 50.2±33.4 | 115.2±34.1 | 98.9±31.6 |
| 4 | 72.9±20.8 | 32.8±5.4 | 130.3±47.5 | 65.6±16.8 |
| 5 | 97.0±24.5 | 62.3±22.6 | 132.8±49.6 | 85.4±31.5 |
| 6 | 51.6±20.7 | 45.8±20.9 | 159.7±69.9 | 132.7±51.1 |
| Combination factor mean | 66.7ac | 44.6 a | 122.3bd | 89.6cd |
SP-HBP, seminal plasma-heparin binding protein; MDA, malondialdehyde; SE, standard error.
Control, without HBP; Treated, supplemented with SP-HBP.
Superscripts a, ac, bd, cd indicates the difference at 5% level of significance within the colums.
There was a non significant (p>0.05) decline of 5.8 to 40.1 μmoles/109 and 16.3 to 64.7 μmoles/109 spermatozoa in pre-frozen and frozen-thawed semen supplemented with SP-HBP, respectively. Karunakaran et al. (2012) added 25 μg of 28 to 30 kDa HBP to the frozen-thawed bull semen in control and H2O2 treated group and concluded that excessive H2O2 causes significant increase in LPO levels and 28 to 30 kDa HBP of sperm membrane helped in controlling the oxidative stress of sperm cells. Kumar et al. (2008) were also of the opinion that HBPs protected the buffalo bull spermatozoa from lipid peroxidation stress during cryopreservation.
Seminal oxidative stress develops as a result of an imbalance between reactive oxygen species and scavenging activities. Spermatozoa are particularly susceptible to the oxidative stress induced damage as their plasma membrane contains large quantity of polyunsaturated fatty acids. Cytoplasm of spermatozoa has low concentration of scavenging enzymes (Sharma and Agarwal, 1996). Moreover mature sperm is devoid of cytoplasm and intracellular antioxidant enzymes cannot protect the plasma membrane that surrounds acrosome and tail. Therefore, for their protection spermatozoa depend upon seminal plasma. Small amounts of reactive oxygen species (ROS) have been presented to be required for several functions of spermatozoa, but their excessive levels can negatively impact the quality of spermatozoa and impair the fertilizing capacity (Tvrda et al., 2011). Reactive oxygen species are usually formed during normal enzymatic reaction of inter and intracellular signalling (Agarwal and Allamaneni, 2006). Therefore, declined level of MDA production in the spermatozoa treated with SP-HBP may be due to its interference with enzymatic reaction of inter and intracellular signalling responsible for ROS production.
Oxidative stress affects the fluidity of sperm plasma membrane as well as integrity of DNA in the nucleus (Aitken, 1999). Increase in membrane fluidity leads to sperm capacitation. Therefore, higher level of acrosome damage (cryocapacitation like changes ) observed in frozen-thawed spermatozoa as compared to pre-frozen spermatozoa may be due to increase in ROS level during cryopreservation of semen, which was also evidenced from increased level of MDA production. Spermatozoa itself produce small amounts of ROS that are essential to many of physiological process i.e. capacitation, hyperactivation and sperm oocyte fusion (Aitken et al., 2003). Low level of ROS has also been shown to be essential for fertilization, acrosome reaction and motility. Production of higher level of ROS as observed by a rising level of MDA production in frozen-thawed spermatozoa was repressed by addition of SP-HBP. Therefore reduced level of oxidative stress in frozen thawed spermatozoa treated with SP-HBP may be responsible for higher percentage of motile, viable, HOS positive, in vitro capacitated/acrosome reacted spermatozoa and spermatozoa with intact acrosomes in pre-frozen and frozen-thawed semen of six cross bred bulls.
It was observed during the present study that cryopreservation of semen of six bulls resulted in reduced motility, viability, HOST and enhanced acrosome damage/oxidative stress with a variation among the bulls. Such a variation may be attributed to varying concentration of membrane cholesterol. Fresh spermatozoa with higher cholesterol content are expected to resist destabilization of membrane following cryopreservation (Srivastava et al., 2013). Since membrane integrity is altered with cryopreservation, spermatozoa with more cholesterol content prior to cryopreservation will also resist membrane damage and effect of membrane damage and cryocapacitation will be less evident in these bulls (Srivastava et al., 2013). Treatment of semen with SP-HBP improved motility, viability, HOST, acrosome integrity, in vitro capacitation/acrosome reaction by diminishing oxidative stress, but with a variation among the bulls. HBPs are acquired by spermatozoa at the time of ejaculation and have been associated with removal of cholesterol from sperm membrane during capacitation (Therien et al., 2001). Therefore, variation in response of spermatozoa to SP-HBP treatments may also be associated to varying concentration of sperm membrane cholesterol.
Our observations revealed that absorption of SP-HBP to the sperm surface reduced the damage that cold shock can have on the sperm membrane during the process of freezing, consequently as pointed out by Lessard et al. (2000), increase in fertilizing capacity of SP-HBP treated cryopreserved semen should be expected.
Footnotes
CONFLICT OF INTEREST
We certify that there is no conflict of interest with any financial organization regarding the material discussed in the manuscript.
REFERENCES
- Agarwal A, Allamaneni S. Oxidative stress and human reproduction. In: Singh KK, editor. Oxidative Stress, Disease and Cancer. Imperial College Press; New York, NY, USA.: 2006. pp. 687–703. [Google Scholar]
- Aitken RJ. The Amoroso lecture the human spermatozoon—a cell in crisis? J Reprod Fertil. 1999;115:1–7. doi: 10.1530/jrf.0.1150001. [DOI] [PubMed] [Google Scholar]
- Aitken RJ, Baker MA, Sawyer D. Oxidative stress in the male germ line and its role in the aetiology of male infertility and genetic disease. Reprod Biomed Online. 2003;7:65–70. doi: 10.1016/s1472-6483(10)61730-0. [DOI] [PubMed] [Google Scholar]
- Alvarez-Gallardo H, Kjelland ME, Moreno JF, Welsh TH, Jr, Randel RD. Gamete therapeutics: Recombinant protein adsorption by sperm for increasing fertility via artificial insemination. Plos One. 2013;8:e65083. doi: 10.1371/journal.pone.0065083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arangasamy A. Effect of stallion seminal plasma proteins on in-vitro capacitation of equine spermatozoa. Ind J Anim Sci. 2010;80:394–397. [Google Scholar]
- Ardon F, Sauarez SS. Cryopreservation increases coating of bull sperm by seminal plasma binder of sperm proteins BSP1, BSP3, and BSP5. Reproduction. 2013;146:111–117. doi: 10.1530/REP-12-0468. [DOI] [PubMed] [Google Scholar]
- Ax RL, Hawkins HE, DeNise SK, Holm TR, Zhang HM. Boca Raton, FL, USA . New developments in managing the bull. In: Fields MJ, Sand RS, Yelich JV, editors; Factors Affecting Calf Crop: Biotechnology of Reproduction. CRC Press LLC; 2002. pp. 287–295. [Google Scholar]
- Barrios B, Perez-Pe R, Gallego M, Tato A, Osada J, Muino-Blanco T, Cebrian-Perez JA. Seminal plasma proteins revert the cold-shock damage on ram sperm membrane. Biol Reprod. 2000;63:1531–1537. doi: 10.1095/biolreprod63.5.1531. [DOI] [PubMed] [Google Scholar]
- Buege JA, Aust SD. Microsomal lipid peroxidation. Methods Enzymol. 1978;52:302–310. doi: 10.1016/s0076-6879(78)52032-6. [DOI] [PubMed] [Google Scholar]
- Chaveiro A, Machado L, Frijters A, Engel B, Woelders H. Improvement of parameters of freezing medium and freezing protocol for bull sperm using two osmotic supports. Theriogenology. 2006;65:1875–1890. doi: 10.1016/j.theriogenology.2005.10.017. [DOI] [PubMed] [Google Scholar]
- Cheema RS, Zodinsanga V, Bansal AK. Characterization and immunolocalization of HBP, FA-1 and TIMP-2 like proteins in cattle bull semen: HBP modification in in vitro capacitated spermatozoa. Theriogenol Insight. 2015;5:53–67. [Google Scholar]
- Farooqui AA. Purification of enzymes by heparin-sepharose affinity chromatography. J Chromatogr A. 1980;184:335–345. doi: 10.1016/s0021-9673(00)89004-4. [DOI] [PubMed] [Google Scholar]
- Fraser LR, Abeydeera LR, Niwa K. Ca2+-Regulating mechanisms that modulate bull sperm capacitation and acrosomal exocytosis as determined by chlortetracycline analysis. Mol Reprod Dev. 1995;40:233–241. doi: 10.1002/mrd.1080400213. [DOI] [PubMed] [Google Scholar]
- Harshan HM, Singh LP, Arangasamy A, Ansari MR, Umar S. Effect of buffalo seminal plasma heparin binding protein (HBP) on freezability and in vitro fertility of buffalo cauda spermatozoa. Anim Reprod Sci. 2006;93:124–133. doi: 10.1016/j.anireprosci.2005.07.010. [DOI] [PubMed] [Google Scholar]
- Jeyendran RS, Vander-Ven HH, Perez-Pelaez M, Crabo BG, Zanevld LJD. Development of an assay to assess the functional integrity of the human sperm membrane and its relationship to other semen characteristics. J Repord Fertil. 1984;70:219–228. doi: 10.1530/jrf.0.0700219. [DOI] [PubMed] [Google Scholar]
- Karunakaran M, Devanathan TG, Kulasekar K, Sridevi P, Jawahar T, Loganatahsamy K, Dhali A, Selvaraju S. Effect of fertility associated protein on oxidative Stress of bovine sperm cells. Ind J Anim Reprod. 2012;33:43–47. [Google Scholar]
- Kumar A, Singh LP, Harshan HM, Majumdar AC. Seminal plasma non-heparin binding proteins (NHBP) reduce the cryoinjury to buffalo cauda epididymal spermatozoa induced by heparin binding proteins (HBP) Anim Reprod Sci. 2008;104:220–226. doi: 10.1016/j.anireprosci.2007.02.014. [DOI] [PubMed] [Google Scholar]
- Lessard C, Parent S, Leclerc P, Bailey JL, Sullivan R. Cryopreservation alters the levels of the bull sperm surface protein P25b. J Androl. 2000;21:700–707. [PubMed] [Google Scholar]
- Lowry OH, Rosenberg NJ, Farr AL, Randall RJ. Protein measurement with the folin phenol reagent. J Biol Chem. 1951;193:265–275. [PubMed] [Google Scholar]
- Manaskova P, Liberda J, Ticha M, Jonakova V. Isolation of non-heparin-binding and heparin-binding proteins of boar prostate. J Chromatograph B. 2002;770:137–143. doi: 10.1016/s0378-4347(01)00480-7. [DOI] [PubMed] [Google Scholar]
- Manjunath P, Nauc V, Bergeron A, Ménard M. Major proteins of bovine seminal plasma bind to the low-density lipoprotein fraction of hen’s egg yolk. Biol Reprod. 2002;67:1250–1258. doi: 10.1095/biolreprod67.4.1250. [DOI] [PubMed] [Google Scholar]
- Miller DJ, Winer MA, Ax RL. Heparin-binding proteins from seminal plasma bind to bovine spermatozoa and modulate capacitation by heparin. Biol Reprod. 1990;42:899–915. doi: 10.1095/biolreprod42.6.899. [DOI] [PubMed] [Google Scholar]
- Mogielnicka-Brzozowska M, Kordan W. Characteristics of selected seminal plasma proteins and their application in the improvement of the reproductive processes in mammals. Pol J Vet Sci. 2011;14:489–499. doi: 10.2478/v10181-011-0074-z. [DOI] [PubMed] [Google Scholar]
- Mogielnicka-Brzozowska M, Wysocki P, Strzezek J, Kordan W. Zinc-binding proteins from boar seminal plasma – isolation, biochemical characteristics and influence on spermatozoa stored at 4°C. Acta Biochim Pol. 2011;58:171–177. [PubMed] [Google Scholar]
- Mota Filho AC, Silva HV, Nunes TG, de Souza MB, de Freitas LA, de Araujo AA, da Silva LDM. Cryopreservation of canine epididymal sperm using ACP-106c and TRIS. Cryobiology. 2014;69:17–21. doi: 10.1016/j.cryobiol.2014.04.013. [DOI] [PubMed] [Google Scholar]
- Nass SJ, Miller DJ, Winer MA, Ax RL. Male accessory sex glands produce heparin-binding proteins that bind to cauda epididymal spermatozoa and are testosterone dependent. Mol Reprod Dev. 1990;25:237–246. doi: 10.1002/mrd.1080250305. [DOI] [PubMed] [Google Scholar]
- Olaciregui, Gil ML, Monton A, Luno V, Jerez RA, Marti JI. Cryopreservation of epididymal stallion sperm. Cryobiology. 2014;68:91–95. doi: 10.1016/j.cryobiol.2013.12.009. [DOI] [PubMed] [Google Scholar]
- Raval RJ, Dhami AJ. Effect of additives on various spermatozoal attributes of fresh, frozen-thawed and refrigerated semen. Ind J Anim Reprod. 2010;31:33–36. [Google Scholar]
- Sharma RK, Agarwal A. Role of reactive oxygen species in male infertility. Urology. 1996;48:835–850. doi: 10.1016/s0090-4295(96)00313-5. [DOI] [PubMed] [Google Scholar]
- Shi Y, Mosser DD, Morimoto RI. Molecularchaperones as HSF1 -specific transcriptional repressors. Genes Dev. 1998;12:654–666. doi: 10.1101/gad.12.5.654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sidhu RS, Sharma RK, Jr, Thomas AJ, Agarwal A. Relationship between creatine kinase activity and semen characteristics in subfertile men. Int J Fertil Women’s Med. 1998;43:192–197. [PubMed] [Google Scholar]
- Srivastava N, Srivastava SK, Ghosh SK, Kumar A, Perumal P, Jerome A. Acrosome membrane integrity and cryocapacitation are related to cholesterol content of bull spermatozoa. Asian Pac J Reprod. 2013;2:126–131. [Google Scholar]
- Therien I, Bousquet D, Manjunath P. Effect of seminal phospholipid-binding proteins and follicular fluid on bovine sperm capacitation. Biol Reprod. 2001;65:41–51. doi: 10.1095/biolreprod65.1.41. [DOI] [PubMed] [Google Scholar]
- Therien I, Moreau R, Manjunath P. Major proteins of bovine seminal plasma and high-density lipoprotein induce cholesterol efflux from epididymal sperm. Biol Reprod. 1998;59:768–776. doi: 10.1095/biolreprod59.4.768. [DOI] [PubMed] [Google Scholar]
- Tvrda E, Knazicka Z, Bardos L, Massanyi P, Lukac N. Impact of oxidative stress on male fertility—A review. Acta Vet Hung. 2011;59:465–484. doi: 10.1556/AVet.2011.034. [DOI] [PubMed] [Google Scholar]
- Ward CR, Storey BT. Determination of the time course of capacitation in mouse spermatozoa using a chlortetracycline fluorescence assay. Dev Biol. 1984;104:287–296. doi: 10.1016/0012-1606(84)90084-8. [DOI] [PubMed] [Google Scholar]
- Watson PF. The causes of reduced fertility with cryopreserved semen. Anim Reprod Sci. 2000;61:481–492. doi: 10.1016/s0378-4320(00)00099-3. [DOI] [PubMed] [Google Scholar]
- Yoon SJ, Kwon WS, Rahman MS, Lee JS, Pang MG. A novel approach to identifying physical markers of cryo-damage in bull spermatozoa. PLoS One. 2015;10:e0126232. doi: 10.1371/journal.pone.0126232. [DOI] [PMC free article] [PubMed] [Google Scholar]