

Abstract
Thymic stromal lymphopoietin (TSLP) stimulates in vitro proliferation of human fetal B-cell precursors. However, its in vivo role during normal human B lymphopoiesis is unknown. Genetic alterations that cause overexpression of its receptor component, cytokine receptor-like factor 2 (CRLF2), lead to high-risk B-cell acute lymphoblastic leukemia implicating this signaling pathway in leukemogenesis. We show that mouse thymic stromal lymphopoietin does not stimulate the downstream pathways (JAK/STAT5 and PI3K/AKT/mTOR) activated by the human cytokine in primary high-risk leukemia with overexpression of the receptor component. Thus, the utility of classic patient-derived xenografts for in vivo studies of this pathway is limited. We engineered xenograft mice to produce human thymic stromal lymphopoietin (+T mice) by injection with stromal cells transduced to express the cytokine. Control (−T) mice were produced using stroma transduced with control vector. Normal levels of human thymic stromal lymphopoietin were achieved in sera of +T mice, but were undetectable in −T mice. Patient-derived xenografts generated from +T as compared to −T mice showed a 3–6-fold increase in normal human B-cell precursors that was maintained through later stages of B-cell development. Gene expression profiles in high-risk B-cell acute lymphoblastic leukemia expanded in +T mice indicate increased mTOR pathway activation and are more similar to the original patient sample than those from −T mice. +T/−T xenografts provide a novel pre-clinical model for understanding this pathway in B lymphopoiesis and identifying treatments for high-risk B-cell acute lymphoblastic leukemia with overexpression of cytokine-like factor receptor 2.
Introduction
Thymic stromal lymphopoietin (TSLP) is an IL-7-like cytokine that plays key roles at several points in normal hematopoietic cell development and function.1–4 The role of TSLP in B lymphopoiesis has been evaluated almost exclusively in mice.5–8 Knowledge of TSLP in normal human B-cell development is limited to a single in vitro study showing that TSLP increases production of fetal B-cell precursors.9 Genetic alterations that cause overexpression of the TSLP receptor component, CRLF2, have been linked to B-cell acute lymphoblastic leukemia (ALL), thus implicating the TSLP pathway in leukemogenesis.10–13 The low homologies of murine versus human TSLP and CRLF2 (approx. 40%)14,15 suggest the need for novel models to study the in vivo role of TSLP in normal and malignant human B lymphopoiesis.
Acute lymphoblastic leukemia is the most common childhood malignancy and primarily involves the B lineage (B-ALL). Although cure rates approach 90%, approximately 10%–20% of patients still relapse.16,17 Genomic profiling has identified several high-risk B-ALL subtypes that are chemoresistant.18–21 These include CRLF2 B-ALL, a leukemia with genetic alterations causing overexpression of the CRLF2 protein on the surface of B-ALL cells.10–13 CRLF2 and the IL-7 receptor alpha chain (IL-7Rα) together form the TSLP receptor signaling complex (Figure 1A).8,22 Binding of TSLP induces CRLF2 and IL-7Rα dimerization leading to activation of the JAK-STAT523,24 and the PIK3/AKT/mTOR pathways,25,26 as demonstrated in CRLF2 B-ALL.27 The finding that JAK kinases are mutated in CRLF2 B-ALL28 suggested that CRLF2 and mutated JAK co-operate to induce constitutive STAT5 activation in CRLF2 B-ALL.29,30 However, approximately half of CRLF2 B-ALL lack JAK mutations. Thus, the role of TSLP in the leukemogenesis of CRLF2 B-ALL remains unclear and the mechanisms for its contribution to chemoresistance are unknown.
Figure 1.
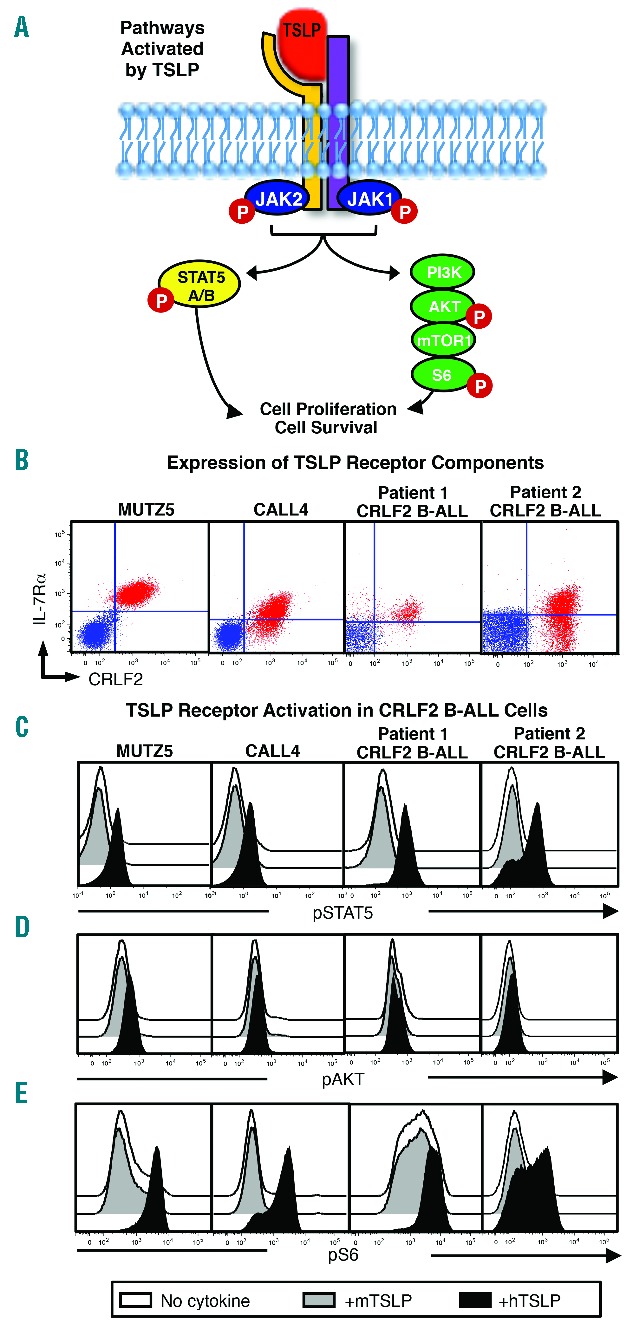
Mouse TSLP does not activate the human TSLP receptor complex. (A) Pathways activated downstream of TSLP receptor in human cells. (B) CRLF2 BALL cell lines (MUTZ5, MHH-CALL4) and primary CRLF2 B-ALL cells used to produce patient-derived xenografts (PDX) used in the studies described here were stained for flow cytometry to detect the TSLP receptor components (IL-7Rα and CRLF2). Plotted in red are CRLF2 B-ALL cells within living cell light scatter. Quadrants shown are set based on unstained controls (blue overlay) (C–E) CRLF2 B-ALL cell lines and primary CRLF2 B-ALL cells were stimulated with human TSLP (hTSLP), mouse TSLP (mTSLP), or no cytokine and evaluated for phosphorylated STAT5 (pSTAT5) AKT (pAKT), and S6 (pS6) by phospho-flow cytometry.
The genetic landscape produced by inherited germline variations contributes to leukemogenesis and disease outcome,31 and is a biological component that contributes to racial, ethnic and other health disparities in ALL.32 This is particularly relevant in CRLF2 B-ALL which occurs five times more often in Hispanic children than others28 and comprises more than half of the ALL cases in children with Down Syndrome.11,18,33 Patient-derived xenograft (PDX) models produced by injecting human cells into immune deficient mice provide in vivo pre-clinical models for understanding disease mechanisms and identifying effective therapies in the context of the range of genetic landscapes present in the human population. However, engineered cellular models8 suggest that mouse TSLP (mTSLP) is species-specific and unlikely to stimulate CRLF2-mediated signaling in human cells.8,22 Given the role of TSLP in activating the CRLF2 pathway,27 and the identification of CRLF2 as a biological component of health disparities in CRLF2 B-ALL,11,18,28,33 it is important that studies to identify disease mechanisms and potential therapies for this leukemia be performed in pre-clinical models that provide human TSLP (hTSLP).
Our goal was to develop and validate a xenograft model that can be used to study the role of hTSLP in normal and malignant B lymphopoiesis. Here we describe the development of a novel xenograft model system comprised of mice that provide hTSLP (+T mice) and mice that do not (−T mice). PDX generated from +T mice show functional in vivo hTSLP effects, expanding the production of normal B-cell precursors from hematopoietic stem cells (HSCs) and inducing changes in mTOR-regulated gene expression in primary CRLF2 B-ALL cells. These data validate the +T/−T xenograft model system as an important new preclinical model for understanding the role of hTSLP in normal and malignant B lymphopoiesis and for identifying therapies to treat CRLF2 B-ALL.
Methods
Human samples and cell lines
Human CRLF2 B-ALL cell lines MUTZ5 and MHH-CALL4 were from DSMZ (Braunschweig, Germany) and the human stroma line HS-27a (HS27) from ATCC (Manassas, VA, USA). Primary human CRLF2 B-ALL cells (Online Supplementary Table S1) and umbilical cord blood (CB) were obtained in accordance with protocols approved by the Loma Linda University Institutional Review Board (IRB) and with the 1975 Declaration of Helsinki, as revised in 2008. Details of HS27 stroma characteristics, CB CD34+ cell isolation, and culture media are described in the Online Supplementary Methods.
Mice
Studies were performed using NOD.Cg-Prkdcscid Il2rgtm1Wjl/SzJ (NSG) mice (Jackson Laboratory) housed under pathogen-free conditions at Loma Linda University Animal Care Facility. Studies were conducted in accordance with the Loma Linda University Institutional Animal Care and Use Committee (IACUC) approved protocols.
Flow cytometry
Cells were prepared for flow cytometry using specific antibody clones as described in the Online Supplementary Methods. Flow cytometry analysis was performed using a MACSQuant analyzer (Miltenyi Biotec Inc., Auburn, CA, USA) and FlowJo analysis software (FlowJo, Ashland, OR, USA).
Stromal cell transduction
Human HS27 stromal cells were transduced to express hTSLP (+T stroma) or with control vector (−T stroma) at a multiplicity of infection of 0.5–5/cell overnight. After transduction, cells were washed, expanded in culture, and frozen in aliquots. For the generation of +T and −T mice, stromal cells were thawed, cultured, and injected into mice within 15 passages after thawing. Details of lentiviral vector production and ELISAs to determine hTSLP levels are available in the Online Supplementary Methods.
Xenograft transplantation
HS27 stroma transduced to be +T or −T were injected intraperitoneally into NSG mice at doses of 5 or 10×106 cells at time points described in the Results section. Mice were sub-lethally irradiated with 225 cGy, then transplanted by tail vein injection with freshly thawed human umbilical CB CD34+ cells (1×105) or primary CRLF2 B-ALL cells (5×105 Patient 1 and 1.5×106 Patient 2).
Gene expression analysis in CRLF2 B-ALL cells
Microarray analyses were performed by Miltenyi Biotec’s Genomic Services (Miltenyi Biotec GmbH, Bergisch Gladbach) using Agilent Whole Human Genome Oligo Microarrays (Design ID 039494, 8×60K v.2). Libraries for RNA sequencing were prepared and sequenced by the UCLA Clinical Microarray Core. Data discussed in this publication are deposited in the NCBI Gene Expression Omnibus (GEO).34
Microarray and RNA sequencing data are available through accession numbers GSE65274 and GSE74339, respectively, at http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSExxx. Gene set enrichment analysis (GSEA) was performed by Novogenix Laboratories, LLC (Los Angeles, CA, USA). Details of microarray, RNA sequencing, data processing, differential gene expression analysis, GSEA, and quantitative RT-PCR analysis are available in the Online Supplementary Appendix.
Further details are available in the Online Supplementary Methods.
Results
Mouse TSLP does not activate the human TSLP receptor complex
Activation of the human TSLP receptor complex can be induced by hTSLP, but not by mTSLP, in murine Ba/F3 cells transduced to express human CRLF2 and IL-7Rα.8 However, the ability of mTSLP to activate CRLF2-signaling in human cells, particularly those expressing high levels of CRLF2 observed in CRLF2 B-ALL, has not been reported. To determine if mTSLP can induce CRLF2-mediated signals, we tested its ability to activate downstream pathways in human CRLF2 B-ALL cells. First, we verified that the CRLF2 B-ALL cell lines and primary CRLF2 B-ALL cells used in our studies express the TSLP receptor components. Expression of IL-7Rα, as well as high levels of CRLF2, was observed in both CRLF2 B-ALL cell lines and in the 2 primary CRLF2 B-ALL samples used in studies described here (Figure 1B).
We then used phospho-flow cytometry to determine whether mTSLP can activate the JAK-STAT5 and PI3K/AKT/mTOR pathways activated by hTSLP in CRLF2 B-ALL cells.27 hTSLP induced robust increases in phosphorylated STAT5 (pSTAT5) and S6 (pS6, activated downstream of mTOR) and more modest increases in phosphorylated AKT (pAKT) in CRLF2 B-ALL cell lines and primary CRLF2 B-ALL cells (Figure 1C–E). In contrast, stimulation of CRLF2 B-ALL cells with mTSLP showed no effect (Figure 1C–E).35 These data show that mTSLP fails to induce the STAT5, AKT, or S6 phosphorylation that are characteristic of CRLF2-mediated signals induced by hTSLP in human CRLF2 B-ALL cells. Thus, TSLP produced in the existing mouse xenograft models does not induce human CRLF2-mediated signals.
Engineering xenograft mice to express normal serum levels of human TSLP
We developed a strategy for engineering xenograft mice that provide hTSLP to activate CRLF2-mediated signals (Figure 2A) by building on a previous cytokine delivery approach.36 Our goal was to produce stable, sustained serum levels of hTSLP, similar to normal levels reported in children (range 13–32 pg/mL)37 by intraperitoneal injection of stromal cells transduced to express high levels of hTSLP (+T mice) (Figure 2A). Control mice (−T mice) were produced by injecting stroma transduced with empty vector. Because the only difference between +T and −T mice is the presence of hTSLP in +T mice, this model would allow us to identify the in vivo effects of hTSLP by comparing hematopoiesis in +T and −T PDX generated from these mice.
Figure 2.
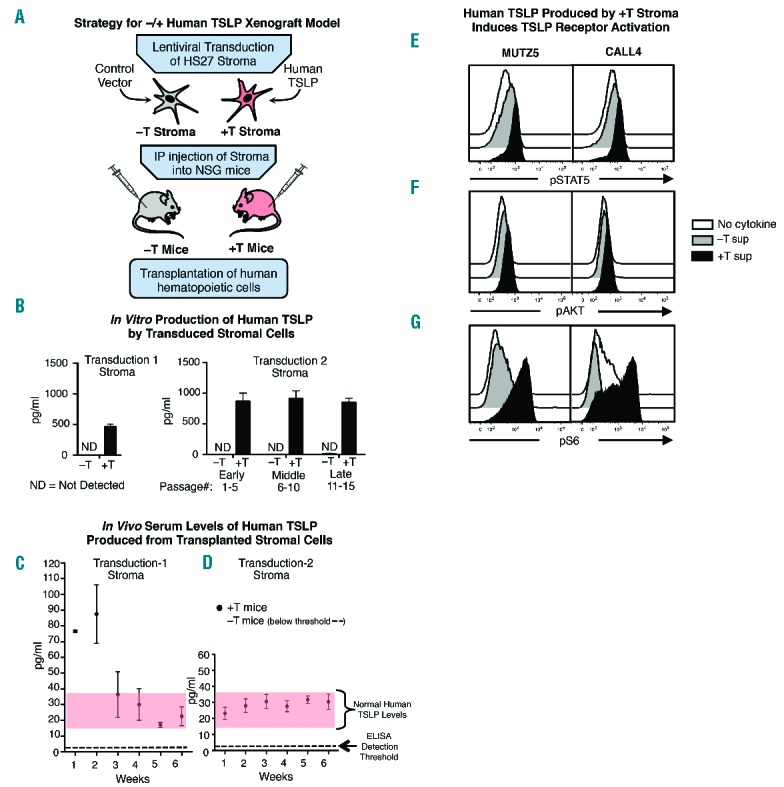
Engineering xenograft mice to express normal serum levels of human TSLP. (A) Strategy for developing the −/+hTSLP xenograft model. (B) ELISA assays of hTSLP protein in culture supernatant from HS27 +T and HS27 −T stromal cells collected after multiple passages following the initial transduction (Transduction 1) or at early (1–5), mid (6–10) and late (11–15) post-thaw passages following Transduction 2. (C–D) Elisa assays of serum hTSLP in blood collected at indicated time points from +T or −T mice injected with stroma as follows: (C) loading dose (3 doses of 1×107 in first week) followed by weekly doses of 1×107 stromal cells (Transduction 1), graphed are the means ± SEM of n = 2–7 mice each time point. (D) injection of 5×106 stromal cells (Transduction 2) at weekly intervals; graphed are the means ± SEM n ≥ 24 mice at each time point. (E–G) CRLF2 B-ALL cell lines were stimulated with stromal cell supernatant (Transduction 2) and levels of pSTAT5, pAKT and pS6 were measured by phospho-flow cytometry.
Human HS27 stromal cells38 were transduced with lentiviral vector containing hTSLP (+T stroma) or with control vector (−T stroma). Culture supernatant collected from stroma after the initial transduction showed hTSLP levels of approximately 500 pg/mL (Figure 2B, Transduction 1). The ability of +T stroma to generate measureable serum levels of hTSLP in vivo was evaluated. NSG mice receiving weekly intraperitoneal injections of 5×106+T stroma showed serum hTSLP levels that were low but detectable (4–10 pg/mL) (data not shown). To achieve higher levels of sustained hTSLP expression, NSG mice were injected with 3 doses of 1×107 +T or −T stroma over a 1-week period (loading dose) followed by weekly injections of 1×107 stromal cells. Serum collected at weekly time points showed serum hTSLP levels of approximately 80 pg/mL following the initial 3 doses of +T stroma but quickly dropped to 15–20 pg/mL with +T stroma injections (Figure 2C, left panel). hTSLP was not detectable in the sera of mice transplanted with −T stroma (Figure 2C). These data show that +T stroma can generate normal serum levels of hTSLP and that serum levels of hTSLP correspond to the number of stromal cells injected during the previous 1–2 week period. These data suggested that stable, sustained hTSLP production might be more likely with weekly injections of stroma that could deliver slightly higher doses of hTSLP.
A second stromal cell transduction produced hTSLP levels averaging approximately 900 pg/mL in culture supernatants harvested at early, mid and late passages from +T stroma across multiple thaws of frozen aliquots (Figure 2, Transduction 2). (hTSLP was undetectable in supernatant from similarly passaged −T stroma (Figure 2, Transduction 2). NSG mice transplanted at weekly time points with 5×106 stroma from Transduction 2 showed serum hTSLP levels of approximately 20 pg/mL that were stable for up to 11 weeks (Figure 2D and data not shown). Evaluation of BM and spleen of +T and −T mice by flow cytometry showed that HS27 stroma (CD29+CD90+) were not detectable in either the spleen or BM of PDX mice (Online Supplementary Figure S1). These data provide evidence that the HS27 +T stroma in the peritoneal cavity give rise to normal serum levels of hTSLP in PDX mice.
To verify that the hTSLP produced in our model induces CRLF2-mediated signals, CRLF2 B-ALL cell lines were assessed for activation of the JAK/STAT5 and PI3K/AKT/mTOR pathways. Supernatant from +T stroma induced robust pSTAT5 and pS6 and minimal pAKT (Figure 2E–G). In contrast, CRLF2 B-ALL cells stimulated with supernatant from −T stroma showed background levels of pSTAT5, pAKT, and pS6, similar to cells incubated with medium alone (Figure 2E–G). These data show that +T stroma, but not −T stroma, used in our model produce hTSLP that can activate signaling in human CRLF2 BALL cells.
hTSLP produced in xenograft mice exerts in vivo functional effects on normal human B-lineage cells
We evaluated the ability of hTSLP in +T PDX to induce in vivo functional effects on human B-lineage cells. hTSLP has been shown to increase the in vitro production of normal human B-cell precursors9 despite their low levels of CRLF2 expression (Online Supplementary Figure S2). Thus, expansion of normal human B-cell precursors provides a sensitive in vivo functional test of the hTSLP produced in our xenograft model. We compared normal B-cell production in +T and −T NSG mice transplanted with umbilical CB CD34+ cells from 3 different donors (Figure 3A and Online Supplementary Figures S3A and S4A). Five weeks post-transplant mice were euthanized and bone marrow (BM) harvested for flow cytometry to evaluate the production of B-lineage and other hematopoietic cells. Mouse and human cells were identified based on expression of mouse CD45 (mCD45) or human CD45 (hCD45), respectively (Figure 3B and Online Supplementary Figures S3B and S4B). Flow cytometry analysis of hCD19 expression on gated human cells showed an increase in the percentage of B-lineage cells in +T as compared to −T PDX mice generated from all 3 CB donors (Figure 3C and Online Supplementary Figures S3C and S4C). Pooled data comparing BM cellularity in all 3 experiments showed that the total number of human cells (hCD45+) was increased by 3–4-fold in +T as compared to −T PDX and this was due to significant increases in CD19+ (B-lineage) cells but not CD19− cells (Figure 3D). These data suggest that hTSLP in +T PDX selectively expands human B-lineage cells rather than exerting global effects on hematopoiesis.
Figure 3.
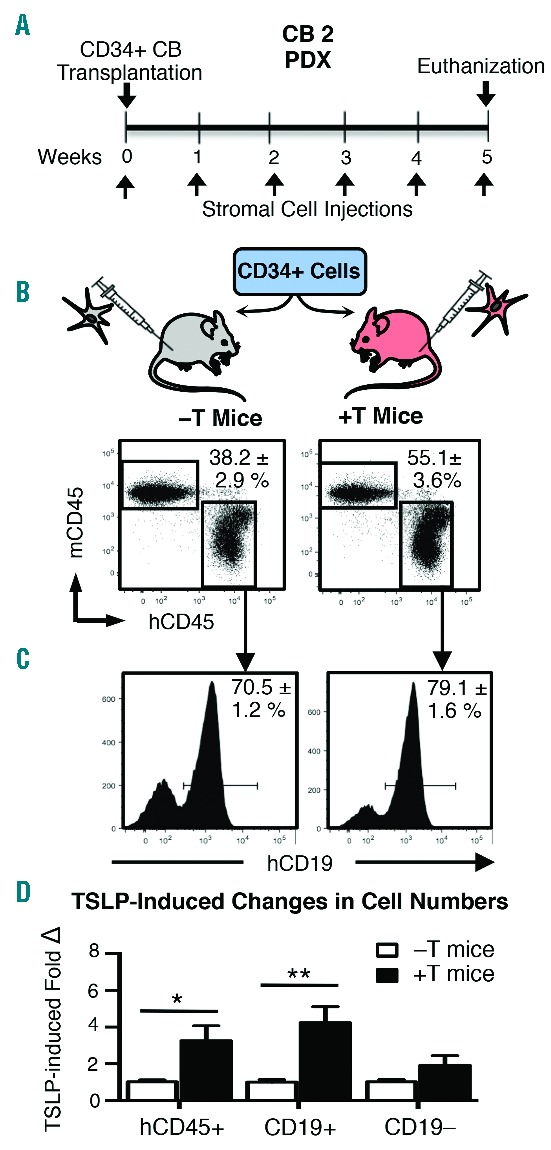
hTSLP produced in xenograft mice exerts in vivo functional effects on normal human B-lineage cells. (A) −T and +T NSG mice were transplanted with 1×105 CD34+ human cord blood (CB) cells and injected with weekly doses of 5×105 stromal cells (Transduction 2 stroma). (B) Five weeks post transplant, mice were euthanized, BM harvested and cells were stained for flow cytometry to detect mouse CD45 (mCD45), human CD45 (hCD45), and hCD19. Total living cells were gated and mCD45 versus hCD45 is plotted with gates to identify mouse and human leukocytes, respectively, in the BM. (C) hCD45+ cells were gated and hCD19 expression is shown by histogram. Percentages are mean ± SEM. Data shown in A–C were obtained from CB2, n=2 −T PDX and n=4 +T PDX; data from two additional CB samples (CB1 and CB3) are shown in Online Supplementary Figures S3–S4. (D) Graphed are the fold change in total human cells (hCD45+), human B-lineage cells (CD19+), and human non-B-lineage cells (CD19−). Fold change was obtained by normalizing cell numbers in +T animals to the average cell numbers in −T animals for each experiment. (D) Data pooled from 3 different experiments performed with CB samples 1–3. Total n=9 −T PDX mice and n=14 +T PDX mice. Statistical significance was calculated using a two-tailed, unpaired t-test (*P≤0.05; ** P≤0.05).
B-cell precursor production is increased in +T PDX mice
To confirm that normal human B-cell precursors are expanded in +T PDX, we compared the production of human B-cell subsets (Figure 4A) and non-B-cell lineages in −T and +T PDX. Staining of PDX BM for markers of human non-B cells showed that human CD19− cells are comprised primarily of monocytes (CD14+), but also include granulocytes (CD66B+), NK cells (CD56+), and CD34+ cells. The number of cells in each non-B lineage was similar (data not shown) in the BM from −T and +T PDX, indicating that the production of these cells is not skewed. However, the production of B-cell precursors was increased 3–6-fold at each stage of B-cell development in +T PDX as compared to −T PDX, beginning with the earliest CD34+ pro-B cells (Figure 4B). This increase was maintained through the mature B-cell stage in BM and spleen (Figure 4B) with no skewing of B-cell subsets in the BM (Figure 4C) or spleen (data not shown). These data provide evidence that hTSLP produced in vivo in +T PDX has a functional effect on normal human B-cell precursors that express low levels of CRLF2.
Figure 4.
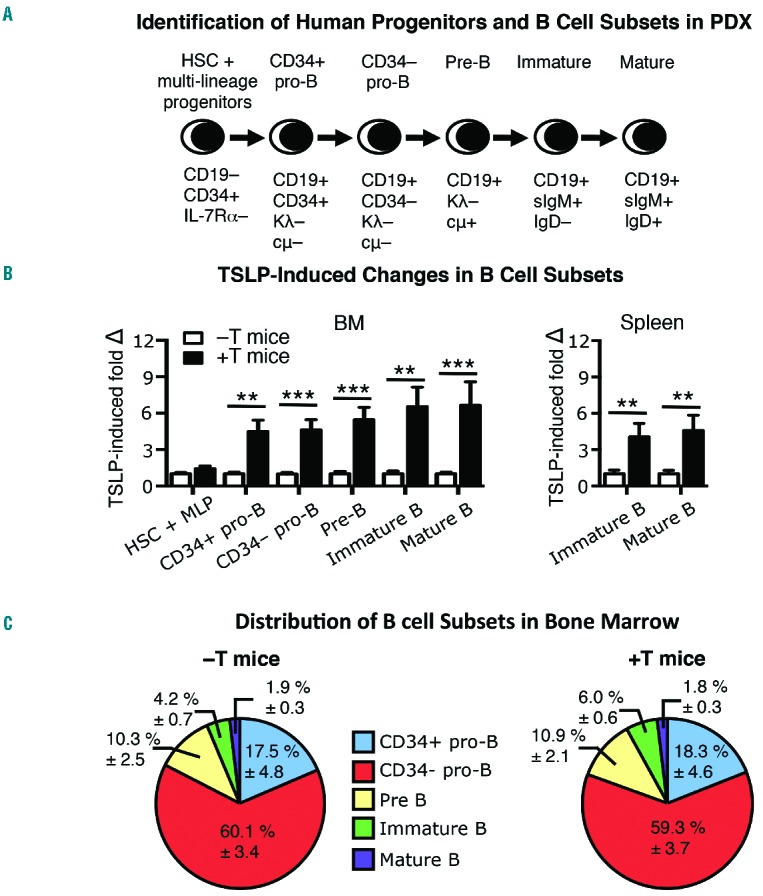
B-cell precursor production is increased in +T PDX. Bone marrow (BM) and spleen were harvested from −T and +T PDX mice generated from 3 different CB samples as shown in Figure 3, and in Online Supplementary Figures S3 and S4. (A) Harvested cells were counted and stained for flow cytometry to identify progenitors and B-cell subsets in the BM and spleen based on indicated immunophenotypes. (B) The number of cells and fold change in each subset were obtained by normalizing cell numbers in +T animals to the average cell numbers in −T animals for each experiment. (C) Graphed is the frequency of each subset within the total CD19+ B-lineage pool. Data shown are pooled from experiments performed using PDX generated from CB 1–3. n=9 −T PDX mice and n=15 +T PDX mice. Statistical significance was calculated using a two-tailed, unpaired t-test (**P≤0.01; ***P ≤0.001).
hTSLP in xenograft mice induces mTOR-regulated genes in primary CRLF2 B-ALL
To determine the ability of hTSLP in +T xenografts to activate CRLF2 downstream pathways in CRLF2-B-ALL cells, we used primary samples obtained from 2 different Hispanic pediatric patients. Patients’ characteristics are shown in Online Supplementary Table S1. First, we evaluated the ability of NSG mice transplanted with HS27 stroma to support engraftment of primary CRLF2 B-ALL cells. PDX from Patient 1 were generated using NSG mice injected with +T stroma from transduction 1 (stroma injection schedule) (Figure 5A). Serum levels of hTSLP produced in these mice were detectable but below normal levels (4–10 pg/mL) and therefore provided a sensitive test of in vivo hTSLP activity in +T PDX. +T and −T NSG mice were transplanted with primary CRLF2 B-ALL cells (Figure 1B, right panel) from Patient 1. Mice were euthanized at nine weeks when peripheral blood chimerism reached 80%. BM was harvested and stained for mCD45 and human-specific markers (CRLF2 and CD19) to identify CRLF2 B-ALL cells (Figure 5B). Flow cytometry analysis showed BM chimerism of human CRLF2 B-ALL cells was more than 90% in +T and −T PDX (Figure 5B). These data show that NSG mice support engraftment of primary BALL cells when injected with −T or with +T stroma.
Figure 5.
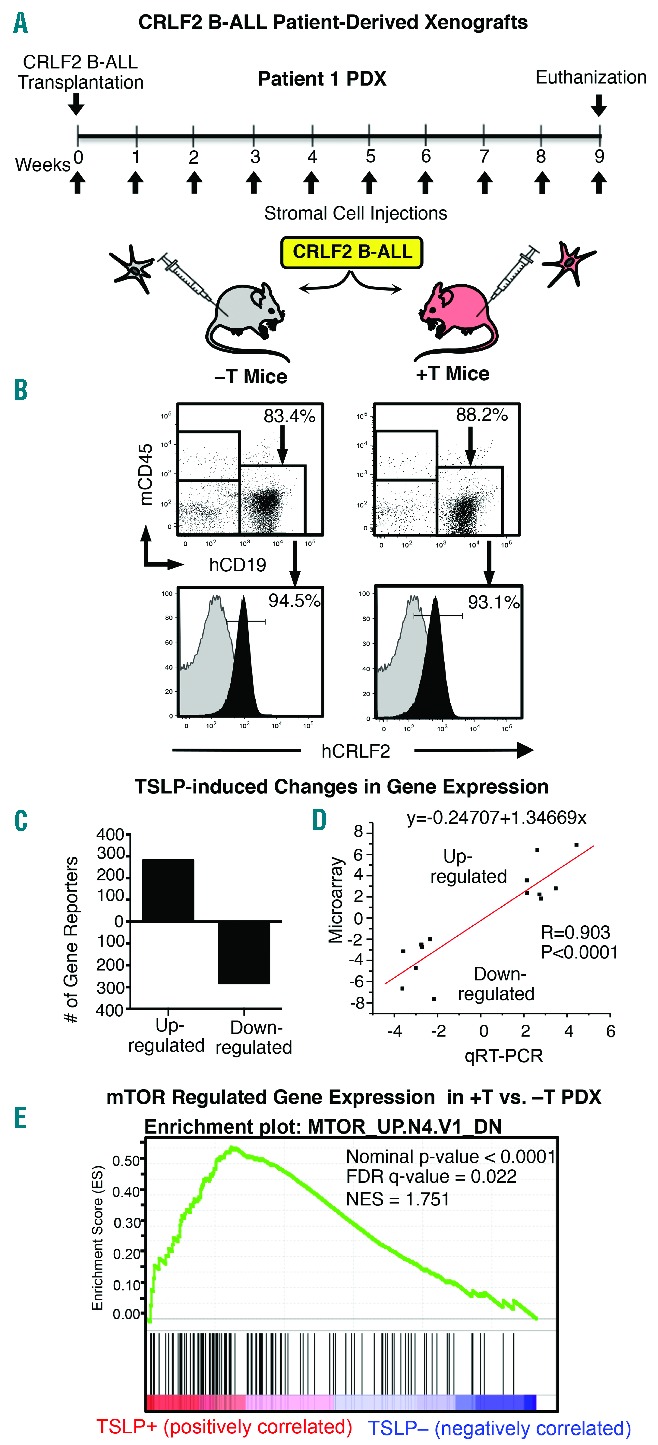
hTSLP in +T xenograft mice induces mTOR-regulated genes in primary CRLF2 B-ALL. (A) −T and +T NSG mice were transplanted with primary CRLF2 B-ALL cells from Patient 1 and injected weekly with 5×106 stromal cells (transduction 1). (B) Nine weeks post transplant, mice were euthanized, BM was harvested and cells were stained with mCD45 and hCD19 to identify human BALL cells. CRLF2 staining (black overlay) on gated hCD19+ cells is shown in bottom panels. Isotype control is shown in gray. (C) Human hCD19+ B-ALL cells were isolated by magnetic separation and analyzed using whole genome microarray to identify genes that are differentially regulated in +T as compared to −T PDX. Graphed are the numbers of Agilent gene reporters up or down-regulated (≥2 fold, P≤0.05) in CRLF2 B-ALL cells from +T as compared to −T PDX. (D) qRT-PCR validation of whole genome microarray. Regression analysis of fold changes measured by microarray versus fold changes measured by qRT-PCR. (E) Gene Set Enrichment Analysis (GSEA) of whole genome microarray data shows that CRLF2 B-ALL cells harvested from +T PDX exhibit an increase in expression of genes regulated downstream of mTOR signaling as compared to cells from −T PDX. The top half of the GSEA enrichment plot shows the enrichment score for each gene and the bottom half shows the values of the ranking metric moving down the list of the ranked genes.
The leukemia cell burden in the BM of −T and +T PDX generated from Patient 1 and from Patient 2 showed no consistent differences (Online Supplementary Figure S6). Thus, our next question was whether hTSLP in +T animals induces gene expression profiles associated with pathway activation downstream of CRLF2-mediated signaling in primary CRLF2 B-ALL cells. Human CRLF2 B-ALL cells were isolated from the BM of +T and −T PDX generated from Patient 1 by magnetic separation. Gene expression was compared by whole genome microarray performed in triplicate. Differential gene expression analysis identified 280 gene reporters that were up-regulated and 281 gene reporters that were down-regulated at least 2-fold (Figure 5C). Microarray was validated by quantitative RT-PCR (qRT-PCR) analysis of selected differentially expressed genes (Online Supplementary Table S2). Changes in gene expression by microarray and qRT-PCR were strongly correlated as indicated by linear regression (Figure 5D). GSEA39,40 of whole genome microarray data showed a strong enrichment for genes regulated downstream of mTOR activation in CRLF2 B-ALL cells from +T PDX as compared to −T PDX (FDR q-value=0.022) (Figure 5E). These data provide evidence that the hTSLP in +T PDX, even when present at low levels, acts on CRLF2 B-ALL cells to increase mTOR pathway activation downstream of CRLF2.
+T PDX show gene expression profiles that are more similar to the original patient sample
Next we performed experiments to determine whether +T PDX provide an in vivo environment more like the patient than −T PDX and if this could be achieved with short-term in vivo hTSLP exposure. PDX were established using CRLF2 B-ALL cells from Patient 2. Injection of stromal cells from transduction 2 were not initiated until two weeks prior to euthanasia (Figure 6A). At euthanasia, BM was harvested and stained for flow cytometry to verify CRLF2 B-ALL engraftment (Figure 6B). Human leukemia cells were isolated from the BM of +T and −T PDX by magnetic separation for RNA sequencing. Differential gene expression analysis identified 503 genes that were up-regulated and 117 that were down-regulated at least 2-fold in CRLF2 B-ALL cells from +T as compared to −T PDX (Figure 6C). Similar to what we observed with Patient 1, GSEA analysis of RNA sequencing data showed an enrichment for genes regulated downstream of mTOR in CRLF2 B-ALL cells from +T as compared to −T PDX (FDR q-value = 0.00048) (Figure 6D). A comparison of +T PDX to −T PDX showed similar immunophenotypes to each other and to that reported for the respective original Patient 1 and Patient 2 samples (Online Supplementary Table S1 and Figure S7), indicating that the differences in mTOR-regulated gene expression are unlikely to be due to selective survival of different clones. These data demonstrate that in vivo hTSLP-induced changes in mTOR regulated gene expression can be observed after short-term stimulation and validate our model in a second primary CRLF2 B-ALL patient sample.
Figure 6.
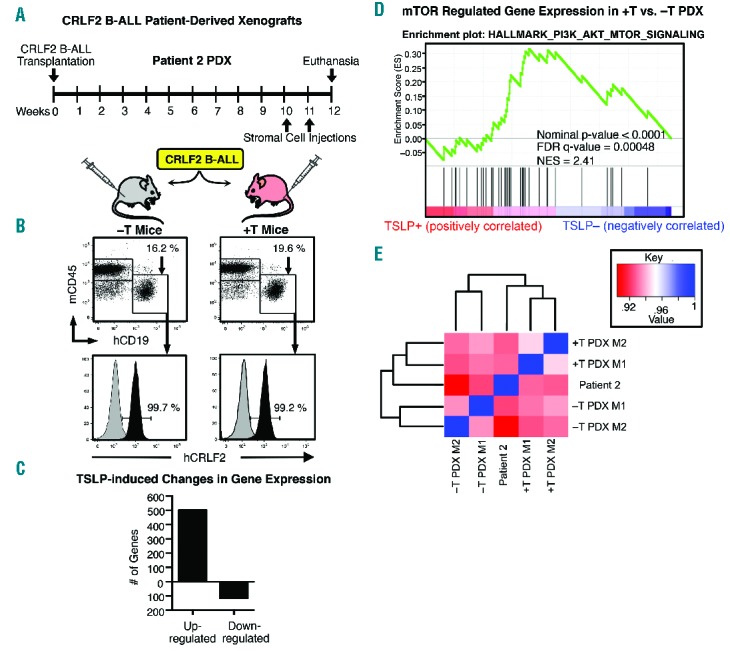
+T PDX show gene expression profiles that are more similar to the original patient sample. (A) −T and +T NSG mice were transplanted with primary CRLF2 B-ALL cells from Patient 2. Beginning at ten weeks after leukemia transplant mice received two injections with 5×106 stromal cells (Transduction 2 stroma) one week apart. (B) Twelve weeks post transplant, mice were euthanized, BM was harvested stained with mCD45 and hCD19 to identify human B-ALL cells. CRLF2 staining (black overlay) on gated hCD19+ cells is shown in bottom panels. Isotype control is shown in gray. (C–E) RNA sequencing was performed on CRLF2 B-ALL cells isolated by magnetic separation from 2 +T PDX mice (+T PDX M1 and M2) and 2 −T PDX mice (−T PDX M1 and M2) and from the original Patient 2 primary sample used to generate them. (C) Differential gene expression analysis was performed and graphed are the numbers of significantly up- or down-regulated genes (≥2-fold, P≤0.05) in CRLF2 B-ALL cells from +T as compared to −T PDX. (D) GSEA of RNA sequencing data shows that CRLF2 B-ALL cells from +T PDX exhibit an increase in expression of genes regulated downstream of mTOR signaling as compared to cells from −T PDX. (E) Graphed is a dendrogram of unsupervised hierarchical clustering and heatmap of Spearman rank correlation values showing that gene expression in +T PDX is more closely correlated (P<0.0001) to the original sample than is −T PDX. Spearman rank coefficients are shown in Online Supplementary Figure S5.
To determine whether +T PDX provide an in vivo environment that is more similar to the patient than −T PDX, we compared gene expression profiles of +T and −T PDX to the original ex vivo CRLF2 B-ALL patient sample. Human leukemia cells were isolated from the BM of +T and −T PDX generated from Patient 2, and from the original primary Patient 2 sample by magnetic separation. RNA sequencing was performed to determine whole genome expression. Analysis of gene expression by unsupervised clustering and Spearman rank correlation show that gene expression in +T PDX, as compared to −T PDX, is significantly more closely correlated to the original patient sample (Figure 6E). These data validate the use of +T/−T xenograft mice as an in vivo pre-clinical model system for studies to understand the role of hTSLP in normal human B-cell production and in the development and progression of CRLF2 B-ALL.
Discussion
Patient-derived xenografts provide a critical complement to other models in understanding normal and malignant human B lymphopoiesis in the context of the broad background of human genetic variation. A limitation of PDX is the reduced cross-species activity of some cytokines. Classic immune deficient mice, including NSG mice, selectively support B-lineage differentiation. This is because IL-7, a cytokine important for human B-cell production,41 shows high species cross reactivity42 while myeloid supportive cytokines do not. Here we show that mTSLP does not induce the increases in STAT5 and PI3K/AKT/mTOR phosphorylation induced by hTSLP in CRLF2 B-ALL cells (Figure 1). CRLF2 B-ALL cells express abundant IL-7Rα and CRLF2 as compared to normal B-cell precursors (Figure 1 and Online Supplementary Figure S1) and thus provide a sensitive test for mTSLP activity. These data establish that the classic mouse xenograft models do not provide cross-species activation of the human TSLP receptor.
The goals of the studies described here were to develop a xenograft model that expresses hTSLP and to validate the effects of hTSLP produced in the model on normal and malignant B-cell precursors. We engineered NSG mice that express normal serum levels of hTSLP (+T mice) or that lack hTSLP (−T mice) (Figure 2) by intraperitoneal injection with stroma transduced to express hTSLP (+T stroma) or with a control vector (−T stroma) (Figure 2). hTSLP produced by +T stroma induced in vitro STAT5 and PI3K/AKT/mTOR pathway activation (Figure 2) and showed in vivo functional effects as indicated by selective expansion of normal B-cell precursors (Figures 3 and 4 and Online Supplementary Figures S3 and S4) and upregulation of the mTOR pathway in primary CRLF2 B-ALL cells (Figures 5 and 6). The studies described here establish and validate a novel hTSLP+/− xenograft model system while providing the first data on the in vivo impact of hTSLP on normal B-cell development and primary CRLF2+ B-ALL cells.
Normal serum levels of hTSLP were achieved in our model; however, a novel aspect of our model is the ability to modulate serum levels of hTSLP based on the timing and number of stromal cells injected intraperitoneally. The ability to vary hTSLP levels or initiate hTSLP at time points after transplant of hematopoietic cells offers advantages for experimental design. The ability to modulate serum levels of hTSLP is also biologically relevant because TSLP production is increased by environmental factors such as pollutants and allergens, and is up-regulated in some autoimmune diseases and solid tumors.43 TSLP is primarily produced by cells residing outside the BM (immune cells, smooth muscle cells, gut and skin epithelial cells),44 although human BM stroma do produce hTSLP.45 Thus, the major source of hTSLP that acts on normal and malignant B cells in vivo in patients is likely to be produced outside the marrow as it is in our model.
Little is known about cellular mechanisms that contribute to the initiation, progression and maintenance of CRLF2 B-ALL or the biological factors that contribute to the health disparities associated with this disease. Studies of the relationship between inherited genetic variation and susceptibility to ALL provide some clues to the relationship of ALL to ethnicity.46 Patient-derived xenograft (PDX) models are particularly critical for health disparities in diseases where inherited genetic variation or alterations such as Down Syndrome are likely to play a role in disease progression.47,48 The ability to generate PDX mice that provide hTSLP makes it possible to study the initiation, progression and maintenance of CRLF2 B-ALL from normal human progenitors transduced to over-express CRLF2 on a range of genetic backgrounds.
The +T and −T PDX generated in the studies described here did not show consistent differences in the leukemia cell burden (Online Supplementary Figure S6). However, changes in gene expression that contribute to chemoresistance would not necessarily be expected to impact leukemia burden in untreated PDX, but would instead be revealed following chemotherapy. CRLF2 B-ALL is a leukemia at high risk for relapse and the mTOR pathway is known to play a role in chemoresistance in multiple malignancies.49 The ability of +T PDX to model TSLP-induced mTOR activation makes this an important model for understanding and identifying therapeutic strategies for overcoming chemoresistance in CRLF2 B-ALL. Our finding that gene expression in +T PDX is significantly more like the original patient sample than −T PDX (Figure 6E and Online Supplementary Figure S5) supports the use of +T PDX mice as a pre-clinical model of CRLF2 B-ALL. PDX generated from +T and −T mice and CRLF2 B-ALL samples obtained from Hispanic or Down Syndrome pediatric patients will allow us to study disease mechanisms: 1) in the context of a genetic background associated with CRLF2 B-ALL health disparities; and 2) in the context of TSLP-CRLF2-mediated signals present in vivo in the patient.
The model developed here provides an important new tool for understanding the mechanisms of TSLP activity in normal and malignant B lymphopoiesis. It also provides a pre-clinical model for identifying therapies to effectively treat CRLF2 B-ALL in an environment that can induce the TSLP-mediated signals present in vivo in patients and in the context of the genetic background that leads to health disparities in this disease.
Acknowledgments
The authors would like to thank Batul T. Suterwala, Jonathon L. Payne, and Michelle Choe for manuscript critique.
Footnotes
Check the online version for the most updated information on this article, online supplements, and information on authorship & disclosures: www.haematologica.org/content/101/04/417
Funding
This work was supported by NIH R21CA162259, NIH P20 MD006988, NIH 2R25 GM060507, a St. Baldrick’s Research Grant and a Hyundai Hope on Wheels Scholar Hope Grant, the Department of Pathology and Human Anatomy, the Department of Basic Sciences, and the Center for Health Disparities and Molecular Medicine at Loma Linda University School of Medicine (to KJP) and by a Grant to Promote Collaborative and Translational Research from Loma Linda University (to KJP and CLM). This work was also supported by NIH R01 HL095120, a St Baldrick’s Foundation Career Development Award, a Hyundai Hope on Wheels Scholar Grant Award, the Four Diamonds Fund of the Pennsylvania State University, and the John Wawrynovic Leukemia Research Scholar Endowment (to SD) The content is solely the responsibility of the authors and does not necessarily represent the official views of the NIH.
References
- 1.Ito T, Wang YH, Duramad O, et al. TSLP-activated dendritic cells induce an inflammatory T helper type 2 cell response through OX40 ligand. J Exp Med. 2005;202(9):1213–1223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Ying S, O’Connor B, Ratoff J, et al. Thymic stromal lymphopoietin expression is increased in asthmatic airways and correlates with expression of Th2-attracting chemokines and disease severity. J Immunol. 2005;174(12):8183–8190. [DOI] [PubMed] [Google Scholar]
- 3.Zhou BH, Comeau MR, De Smedt T, et al. Thymic stromal lymphopoietin as a key initiator of allergic airway inflammation in mice. Nat Immunol. 2005;6(10):1047–1053. [DOI] [PubMed] [Google Scholar]
- 4.Siracusa MC, Saenz SA, Hill DA, et al. TSLP promotes interleukin-3-independent basophil haematopoiesis and type 2 inflammation. Nature. 2011;477(7363):229–233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Ray RJ, Furlonger C, Williams DE, Paige CJ. Characterization of thymic stromal-derived lymphopoietin (TSLP) in murine B cell development in vitro. Eur J Immunol. 1996;26(1):10–16. [DOI] [PubMed] [Google Scholar]
- 6.Sims JE, Williams DE, Morrissey PJ, et al. Molecular cloning and biological characterization of a novel murine lymphoid growth factor. J Exp Med. 2000;192(5):671–680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Osborn MJ, Ryan PL, Kirchhof N, Panoskaltsis-Mortari A, Mortari F, Tudor KS. Overexpression of murine TSLP impairs lymphopoiesis and myelopoiesis. Blood. 2004;103(3):843–851. [DOI] [PubMed] [Google Scholar]
- 8.Reche PA, Soumelis V, Gorman DM, et al. Human thymic stromal lymphopoietin preferentially stimulates myeloid cells. J Immunol. 2001;167(1):336–343. [DOI] [PubMed] [Google Scholar]
- 9.Scheeren FA, van Lent AU, Nagasawa M, et al. Thymic stromal lymphopoietin induces early human B-cell proliferation and differentiation. Eur J Immunol. 2010;40(4):955–965. [DOI] [PubMed] [Google Scholar]
- 10.Yoda A, Yoda Y, Chiaretti S, et al. Functional screening identifies CRLF2 in precursor B-cell acute lymphoblastic leukemia. Proc Natl Acad Sci USA. 2010;107(1):252–257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Russell LJ, Capasso M, Vater I, et al. Deregulated expression of cytokine receptor gene, CRLF2, is involved in lymphoid transformation in B-cell precursor acute lymphoblastic leukemia. Blood. 2009;114(13):2688–2698. [DOI] [PubMed] [Google Scholar]
- 12.Chapiro E, Russell L, Lainey E, et al. Activating mutation in the TSLPR gene in B-cell precursor lymphoblastic leukemia. Leukemia. 2010;24(3):642–645. [DOI] [PubMed] [Google Scholar]
- 13.Cario G, Zimmermann M, Romey R, et al. Presence of the P2RY8-CRLF2 rearrangement is associated with a poor prognosis in non-high-risk precursor B-cell acute lymphoblastic leukemia in children treated according to the ALL-BFM 2000 protocol. Blood. 2010;115(26):5393–5397. [DOI] [PubMed] [Google Scholar]
- 14.Quentmeier H, Drexler HG, Fleckenstein D, et al. Cloning of human thymic stromal lymphopoietin (TSLP) and signaling mechanisms leading to proliferation. Leukemia. 2001;15(8):1286–1292. [DOI] [PubMed] [Google Scholar]
- 15.Tonozuka Y, Fujio K, Sugiyama T, Nosaka T, Hirai M, Kitamura T. Molecular cloning of a human novel type I cytokine receptor related to delta1/TSLPR. Cytogenet Cell Genet. 2001;93(1–2):23–25. [DOI] [PubMed] [Google Scholar]
- 16.Raetz EA, Bhatla T. Where do we stand in the treatment of relapsed acute lymphoblastic leukemia¿ Hematology Am Soc Hematol Educ Program. 2012;2012:129–136. [DOI] [PubMed] [Google Scholar]
- 17.Hunger SP, Lu X, Devidas M, et al. Improved survival for children and adolescents with acute lymphoblastic leukemia between 1990 and 2005: a report from the children’s oncology group. J Clin Oncol. 2012;30(14):1663–1669. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Mullighan CG, Collins-Underwood JR, Phillips LA, et al. Rearrangement of CRLF2 in B-progenitor- and Down syndrome-associated acute lymphoblastic leukemia. Nat Genet. 2009;41(11):1243–1246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Mullighan CG, Goorha S, Radtke I, et al. Genome-wide analysis of genetic alterations in acute lymphoblastic leukaemia. Nature. 2007;446(7137):758–764. [DOI] [PubMed] [Google Scholar]
- 20.Mullighan CG. Genomic profiling of B-progenitor acute lymphoblastic leukemia. Best Pract Res Clin Haematol. 2011;24(4):489–503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Chen IM, Harvey RC, Mullighan CG, et al. Outcome modeling with CRLF2, IKZF1, JAK, and minimal residual disease in pediatric acute lymphoblastic leukemia: a Children’s Oncology Group study. Blood. 2012;119(15):3512–3522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Park LS, Martin U, Garka K, et al. Cloning of the murine thymic stromal lymphopoietin (TSLP) receptor: Formation of a functional heteromeric complex requires interleukin 7 receptor. J Exp Med. 2000;192(5):659–670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Wohlmann A, Sebastian K, Borowski A, Krause S, Friedrich K. Signal transduction by the atopy-associated human thymic stromal lymphopoietin (TSLP) receptor depends on Janus kinase function. Biol Chem. 2010;391(2–3):181–186. [DOI] [PubMed] [Google Scholar]
- 24.Rochman Y, Kashyap M, Robinson GW, et al. Thymic stromal lymphopoietin-mediated STAT5 phosphorylation via kinases JAK1 and JAK2 reveals a key difference from IL-7-induced signaling. Proc Natl Acad Sci USA. 2010;107(45):19455–19460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Zhong J, Kim MS, Chaerkady R, et al. TSLP signaling network revealed by SILAC-based phosphoproteomics. Mol Cell Proteomics. 2012;11(6):M112.017764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Zhong J, Sharma J, Raju R, et al. TSLP signaling pathway map: a platform for analysis of TSLP-mediated signaling. Database (Oxford). 2014;2014:bau007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Tasian SK, Doral MY, Borowitz MJ, et al. Aberrant STAT5 and PI3K/mTOR pathway signaling occurs in human CRLF2-rearranged B-precursor acute lymphoblastic leukemia. Blood. 2012;120(4):833–842. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Harvey RC, Mullighan CG, Chen IM, et al. Rearrangement of CRLF2 is associated with mutation of JAK kinases, alteration of IKZF1, Hispanic/Latino ethnicity, and a poor outcome in pediatric B-progenitor acute lymphoblastic leukemia. Blood. 2010;115(26):5312–5321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Roll JD, Reuther GW. CRLF2 and JAK2 in B-progenitor acute lymphoblastic leukemia: a novel association in oncogenesis. Cancer Res. 2010;70(19):7347–7352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Malin S, McManus S, Cobaleda C, et al. Role of STAT5 in controlling cell survival and immunoglobulin gene recombination during pro-B cell development. Nat Immunol. 2010;11(2):171–179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Moriyama T, Relling MV, Yang JJ. Inherited genetic variation in childhood acute lymphoblastic leukemia. Blood. 2015;125(26): 3988–3995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Lim JY, Bhatia S, Robison LL, Yang JJ. Genomics of racial and ethnic disparities in childhood acute lymphoblastic leukemia. Cancer. 2014;120(7):955–962. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Hertzberg L, Vendramini E, Ganmore I, et al. Down syndrome acute lymphoblastic leukemia, a highly heterogeneous disease in which aberrant expression of CRLF2 is associated with mutated JAK2: a report from the International BFM Study Group. Blood. 2010;115(5):1006–1017. [DOI] [PubMed] [Google Scholar]
- 34.Edgar R, Domrachev M, Lash AE. Gene Expression Omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res. 2002;30(1):207–210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Dann SG, Selvaraj A, Thomas G. mTOR Complex1-S6K1 signaling: at the crossroads of obesity, diabetes and cancer. Trends Mol Med. 2007;13(6):252–259. [DOI] [PubMed] [Google Scholar]
- 36.Dao MA, Pepper KA, Nolta JA. Long-term cytokine production from engineered primary human stromal cells influences human hematopoiesis in an in vivo xenograft model. Stem Cells. 1997;15(6):443–454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Lee EB, Kim KW, Hong JY, Jee HM, Sohn MH, Kim KE. Increased serum thymic stromal lymphopoietin in children with atopic dermatitis. Pediatr Allergy Immunol. 2010;21(2 Pt 2):e457–460. [DOI] [PubMed] [Google Scholar]
- 38.Graf L, Iwata M, Torok-Storb B. Gene expression profiling of the functionally distinct human bone marrow stromal cell lines HS-5 and HS-27a. Blood. 2002;100(4):1509–1511. [DOI] [PubMed] [Google Scholar]
- 39.Mootha VK, Lindgren CM, Eriksson KF, et al. PGC-1alpha-responsive genes involved in oxidative phosphorylation are coordinately downregulated in human diabetes. Nat Genet. 2003;34(3):267–273. [DOI] [PubMed] [Google Scholar]
- 40.Subramanian A, Tamayo P, Mootha VK, et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc Natl Acad Sci USA. 2005;102(43):15545–15550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Parrish YK, Baez I, Milford TA, et al. IL-7 Dependence in human B lymphopoiesis increases during progression of ontogeny from cord blood to bone marrow. J Immunol. 2009;182(7):4255–4266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Johnson SE, Shah N, Panoskaltsis-Mortari A, LeBien TW. Murine and human IL-7 activate STAT5 and induce proliferation of normal human pro-B cells. J Immunol. 2005;175(11):7325–7331. [DOI] [PubMed] [Google Scholar]
- 43.Ziegler SF, Roan F, Bell BD, Stoklasek TA, Kitajima M, Han H. The biology of thymic stromal lymphopoietin (TSLP). Adv Pharmacol. 2013;66:129–155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Takai T. TSLP expression: cellular sources, triggers, and regulatory mechanisms. Allergol Int. 2012;61(1):3–17. [DOI] [PubMed] [Google Scholar]
- 45.Allakhverdi Z, Comeau MR, Armant M, et al. Mast Cell-Activated Bone Marrow Mesenchymal Stromal Cells Regulate Proliferation and Lineage Commitment of CD34(+) Progenitor Cells. Front Immunol. 2013;4(461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Xu H, Yang W, Perez-Andreu V, et al. Novel susceptibility variants at 10p12.31-12.2 for childhood acute lymphoblastic leukemia in ethnically diverse populations. J Natl Cancer Inst. 2013;105(10):733–742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Liem NL, Papa RA, Milross CG, et al. Characterization of childhood acute lymphoblastic leukemia xenograft models for the preclinical evaluation of new therapies. Blood. 2004;103(10):3905–3914. [DOI] [PubMed] [Google Scholar]
- 48.Lock RB, Liem NL, Papa RA. Preclinical testing of antileukemic drugs using an in vivo model of systemic disease. Methods Mol Med. 2005;111:323–334. [DOI] [PubMed] [Google Scholar]
- 49.Jiang BH, Liu LZ. Role of mTOR in anticancer drug resistance: perspectives for improved drug treatment. Drug Resist Updat. 2008;11(3):63–76. [DOI] [PMC free article] [PubMed] [Google Scholar]