

ABSTRACT
Translation control plays an important role in the regulation of gene expression in the malaria parasite Plasmodium falciparum, especially in transition stages between the vertebrate host and mosquito vector. Here, we determined the function of the Puf-family member Puf1 (denoted as PfPuf1 for the P. falciparum protein) during P. falciparum sexual development. We show that PfPuf1 was expressed in all gametocyte stages and at higher levels in female gametocytes. PfPuf1 disruption did not interfere with the asexual erythrocyte cycle of the parasite but resulted in an approximately tenfold decrease of mature gametocytes. In the PfPuf1-disrupted lines, gametocytes appeared normal before stage III but subsequently exhibited a sharp decline in gametocytemia. This was accompanied by a concomitant accumulation of dead and dying late-stage gametocytes, which retained normal gross morphology. In addition, significantly more female gametocytes were lost in the PfPuf1-disrupted lines during development, resulting in a reversed male-to-female sex ratio. These results indicate that PfPuf1 is important for the differentiation and maintenance of gametocytes, especially female gametocytes.
KEY WORDS: Plasmodium falciparum, Puf family, Translation regulation, Gametocyte, Sex ratio
Highlighted Article: Characterization of the RNA-binding protein Puf1 as an important regulator required for the maintenance of late-stage gametocytes, the sexual form required for transmission to mosquitoes, in the malaria parasite.
INTRODUCTION
Malaria is a devastating tropical disease killing an estimated 438,000 people in 2015 (WHO, 2015). The malaria parasite Plasmodium falciparum is responsible for more than 90% of the fatal cases. The malaria parasites undergo a complex life cycle, involving multiple stages in both invertebrate and vertebrate hosts. Transition between the two hosts requires specialized parasite stages – the gametocytes and sporozoites. The parasite developmental program is tightly regulated at both the transcriptional and translational levels (Coulson et al., 2004; Le Roch et al., 2003; Lindner et al., 2013; Mair et al., 2006; Painter et al., 2011). In particular, translation regulation plays a crucial role during host transition, depending on translational activation of stored and silent mRNAs (Cui et al., 2015). Translational regulation in Plasmodium has long been recognized. It has been found that the mRNAs for the major ookinete surface antigens P25 and P28 are already abundantly expressed in gametocytes but active protein translation only occurs in the mosquito stages (del Carmen Rodriguez et al., 2000; Kuersten and Goodwin, 2003; Paton et al., 1993). The recent comprehension of translational regulation as a widespread phenomenon in malaria parasites came from comparisons of the transcriptomes and proteomes in P. falciparum (Foth et al., 2011; Le Roch et al., 2004; Otto et al., 2010) and the rodent parasite Plasmodium berghei (Hall et al., 2005).
RNA-binding proteins (RBPs) are crucial players in translational regulation. In P. berghei, translationally silent mRNAs that have accumulated in the cytoplasm of female gametocytes appear to be stabilized by an RBP, the DEAD-box RNA helicase DOZI (Mair et al., 2006), whereas in P. falciparum, some of the mRNAs are regulated by an RBP of the Puf protein family (Miao et al., 2013). Puf proteins, named after two founding members Pumilio in Drosophila and fem-3-binding factor (FBF) in Caenorhabditis elegans, are evolutionarily conserved and widespread in eukaryotes (Spassov and Jurecic, 2003). Puf proteins are characterized by a conserved RNA-binding domain (RBD), which contains eight tandem α-helical repeats of ∼36 amino acids (Edwards et al., 2001). Both Pumilio and FBF proteins recognize a UGUR motif in the 3′ UTRs of target mRNAs and have a conserved mechanism of translational regulation. In most cases, Puf mediates translation repression and/or acceleration of decay of target mRNAs (Goldstrohm et al., 2007; Olivas and Parker, 2000; Wharton et al., 1998), whereas in some cases, Puf also contributes to activation of mRNA expression (Archer et al., 2009; Kaye et al., 2009; Pique et al., 2008; Suh et al., 2009). Puf proteins play diverse biological roles in stem cell maintenance, neurogenesis and embryonic development in different organisms (Mee et al., 2004; Menon et al., 2004; Muraro et al., 2008; Wickens et al., 2002; Ye et al., 2004). Two conserved Puf proteins in P. falciparum, Puf1 and Puf2 (denoted as PfPuf1 and PfPuf2 for the P. falciparum proteins), are preferentially expressed in gametocytes and sporozoites (Cui et al., 2002; Fan et al., 2004; Le Roch et al., 2003; Young et al., 2005). Disruption of PfPuf2 results in increased gametocyte differentiation and elevation of the male-to-female sex ratio (Miao et al., 2013). Studies on the Puf2 protein in P. berghei and P. yoelii have shown that knockout of Puf2 results in failure of maintenance of sporozoites in mosquito salivary glands and premature transformation of sporozoites into liver-stage-like parasites (Gomes-Santos et al., 2011; Lindner et al., 2013; Muller et al., 2011). However, Puf1 knockout in P. berghei did not reveal any noticeable phenotypic changes during parasite development (Gomes-Santos et al., 2011; Muller et al., 2011).
Recent studies on the underlying molecular mechanisms of gametocytogenesis have identified a transcriptional factor, AP2-G, as the switch for the commitment of gametocyte development in both P. falciparum and P. berghei (Kafsack et al., 2014; Sinha et al., 2014). Nevertheless, P. falciparum still differs in the process of sexual development in several aspects from that in other malaria parasites. Most notably, gametocyte development takes 10–12 days in P. falciparum as compared to about 30 h in most Plasmodium species. In addition, gametocytes in other species are round in shape, whereas P. falciparum gametocytes develop with five morphologically distinct stages, and the final mature gametocytes take a crescent form, where this parasite derived its name (Carter and Graves, 1988; Sinden, 1998). For P. falciparum, only the youngest stage-I and mature gametocytes are observed in blood circulation, whereas other immature stages of gametocytes are sequestered in the bone marrow and possibly the spleen (Farfour et al., 2012; Joice et al., 2014). These distinctions in gametocyte development between P. falciparum and other malaria parasites prompted us to analyze the functions of PfPuf1 during gametocytogenesis in P. falciparum. Through genetic disruption of PfPuf1, we demonstrate that it plays an important role in the maintenance of gametocytes, especially at later developmental stages.
RESULTS
PfPuf1 is expressed at all gametocyte stages
To study the expression pattern and subcellular localization of PfPuf1, NF54 parasites were transfected with a pHD22y-Puf1-GFP construct to tag the endogenous PfPuf1 at the C-terminus with the green fluorescent protein (GFP) (Fig. 1A). We used integration-specific genomic DNA PCR to confirm the correct integration of the plasmid into the PfPuf1 locus in a selected parasite clone (Fig. 1B). Expression of the PfPuf1–GFP fusion protein was analyzed by western blotting and quantified by indirect enzyme-linked immunosorbent assay (iELISA). Western blot analysis of protein lysates from mature gametocytes with antibodies against GFP detected a protein band of ∼250 kDa only in the GFP-tagged parasite line, which is close to the estimated size of 252 kDa for PfPuf1–GFP (Fig. 1C). To determine the expression dynamics of PfPuf1 during gametocytogenesis, RNA and proteins were obtained from day-4, -8 and -12 gametocytes after induction. Consistent with a previous finding (Cui et al., 2002), real-time reverse-transcriptase PCR (real-time RT-PCR) analysis detected a gradual increase in the transcript level of PfPuf1 during gametocyte development (Fig. 1D, upper panel). Concomitantly, PfPuf1–GFP protein expression was also increased during development (Fig. 1D, lower panel). GFP signal was monitored with fluorescence microscopy throughout the entire duration of gametocyte development (Fig. 1E), but was not noticeable during asexual stages, which is in concordance with the lack of PfPuf1 transcript in asexual stages as revealed by microarray (Le Roch et al., 2003) and RNAseq (Lopez-Barragan et al., 2011) analyses. PfPuf1–GFP was localized in the cytoplasm of developing gametocytes, sometimes with a punctate appearance, especially at later stages. An indirect immunofluorescence assay with antibodies against the gametocyte surface marker Pfs230 indicated that PfPuf1 expression was indeed intracellular (Fig. 1F).
Fig. 1.

PfPuf1 expression and localization in P. falciparum gametocytes. (A) GFP tagging of PfPuf1 in NF54 strain. Schemes of the endogenous PfPuf1 locus on chromosome 5, the pHD22Y-Puf1-GFP plasmid and the resultant PfPuf1 locus with integration of GFP from a single crossover event. Filled boxes represent the gene region used for homologous recombination. (B) Confirmation of GFP integration by performing integration-specific PCR using primers F1 and R2. (C) Western blot analysis showing expression of the PfPuf1–GFP fusion protein from day-12 gametocytes using anti-GFP antibody. M indicates molecular marker in kDa. (D) Dynamics of PfPuf1 expression during gametocytogenesis. Upper panel: The relative abundance of PfPuf1 mRNA in day-4, -8 and -12 gametocytes as determined with real-time RT-PCR using primers GFP-F and GFP-R shown in panel A. Lower panel: PfPuf1–GFP protein expression quantified with iELISA using anti-GFP antibodies. (E) Representative microscopy images of stage-II to stage-V gametocytes showing localization of Puf1–GFP. Nuclei were counterstained with Hoechst 33342. DIC, differential interference contrast microscopy. (F) Localization of PfPuf1 was defined by immunofluorescence staining of the surface marker Pfs230. Scale bars: 5 μm.
PfPuf1 is expressed differentially in male and female gametocytes
Under a fluorescence microscope, we observed variations in the GFP signal intensity in PfPuf1–GFP gametocytes (Fig. 2A). To further confirm this observation and quantify these changes, we performed flow cytometry analysis of the fluorescent gametocytes on days 4 and 8 after induction. Based on gating for GFP fluorescence intensity, purified gametocytes from the PfPuf1–GFP line formed two populations, which could be roughly divided into low-GFP and high-GFP populations, with the high-GFP population being approximately twice as abundant as that of the low-GFP population (Fig. 2B). To test whether these gametocyte populations were enriched for different sexes, we sorted these populations and performed real-time RT-PCR analysis using male (α-tubulin II) and female (Pfs47) gametocyte markers. Interestingly, the α-tubulin-II mRNA level in the low-GFP population was approximately three times higher than in the high-GFP population, whereas Pfs47 transcript levels showed the opposite (Fig. 2C, left panel). This resulted in a more than fivefold higher α-tubulin II:Pfs47 ratio in the low-GFP population than in the high-GFP population, indicating that low- and high-GFP populations were enriched for male and female gametocytes, respectively (P<0.05, t-test) (Fig. 2C, right panel). These two populations at the mature stage were also observed by using microscopy after Giemsa staining, which showed that more gametocytes in the low- and high-GFP populations morphologically resembled males and females, respectively (data not shown). Moreover, an indirect immunofluorescence assay with an antibody against α-tubulin II showed a higher level of α-tubulin II expression in the low-GFP than in the high-GFP gametocytes (Fig. 2D), similar to the pattern of α-tubulin II staining in male versus female gametocytes observed in a previous study (Schwank et al., 2010). These results indicate that PfPuf1–GFP was expressed at a higher level in female gametocytes.
Fig. 2.

Differential expression of PfPuf1–GFP in P. falciparum male and female gametocytes. (A) Microscopy images of stage-V gametocytes of the NF54 line with PfPuf1–GFP showing differences in GFP fluorescence intensities in individual gametocytes. (B) Flow cytometry analysis of early- (stage II, day 4) and late-stage (stage IV, day 8) gametocytes based on GFP fluorescence intensities. The gametocytes were roughly divided into two populations (circled), which were individually sorted using a cell sorter and used for analysis in C. The GFP signal was defined at 488-nm laser excitation with one bandpass filter, GFP (513/22 nm). FSC, forward scatter channel. (C) Real-time RT-PCR analysis of transcript abundance of male (α-Tubulin II) and female (Pfs47) gametocyte markers in the two GFP populations. Left: relative transcript abundance of male and female gametocyte markers in GFP-positive populations. Total RNA was extracted from separated gametocytes with different GFP fluorescence intensities and used for cDNA synthesis and real-time RT-PCR. Data are shown in mean±s.d. from three replicates. Right: the male-to-female ratio represented by the α-Tubulin II:Pfs47 ratio (α-Tubulin II/Pfs47) was significantly different in low- and high-GFP gametocyte populations (P<0.05, t-test). (D) GFP fluorescence was analyzed with immunofluorescence with an antibody against α-tubulin II. DIC, differential interference contrast microscopy. Scale bars: 5 μm.
Disruption of PfPuf1 leads to decreased gametocytemia
To determine the role of PfPuf1 during P. falciparum development, we constructed a plasmid to disrupt the PfPuf1 RBD. Integration of the plasmid through homologous recombination of a ∼1-kb fragment upstream of the RBD results in the expression of an N-terminally truncated PfPuf1–GFP fusion protein (Fig. S1A). NF54 parasites were transfected and selected after three alternating drug-on and drug-off cycles with WR99210 to enrich for parasites with chromosomal integration of the plasmid, which was followed by parasite cloning. Given the prohibitory large size of PfPuf1 for complementation studies, multiple transfection experiments were performed in order to obtain a more conclusive analysis of PfPuf1 function. PfPuf1-disrupted clones (ΔPfPuf1–GFP) were selected based on the GFP signal in gametocytes under a microscope and confirmed by Southern blot and genomic DNA PCR analysis. Three clones with GFP signals from different transfection experiments were selected for confirmation of PfPuf1 disruption. Genomic Southern blot showed a DNA banding pattern in all three clones that was consistent with the correct disruption of the PfPuf1 locus (Fig. S1B). In agreement, genomic DNA PCR using primers flanking the homologous recombination fragment in PfPuf1 only amplified the ∼2-kb fragment from the wild-type NF54 genomic DNA, but not from the GFP-positive clones because the expected size of the genomic DNA PCR fragment (10.3 kb) in the ΔPfPuf1–GFP clones would be too large to be amplified under the genomic DNA PCR conditions used (Fig. S1C). Similarly, RT-PCR analysis with gametocyte RNA could only amplify the PfPuf1 3′ region in NF54 but not in the ΔPfPuf1–GFP clones (Fig. S1C).
To investigate phenotypic changes associated with PfPuf1 disruption, we quantified asexual multiplication and gametocyte development using Giemsa-stained smears from the cultures. Consistent with preferential expression of PfPuf1 during gametocytogenesis, there was no noticeable difference in the in vitro growth rate of asexual-stage parasites between the ΔPfPuf1-GFP clones and wild-type NF54 (Fig. S1D). Although the gametocytemia was not significantly different between the ΔPfPuf1-GFP and wild-type NF54 clones during the first 7 days after gametocyte induction, it was significantly lower in the ΔPfPuf1-GFP clones from day 8 onwards (P<0.001, t-test) (Fig. 3), suggesting that PfPuf1 plays a more prominent role in the development of late-stage gametocytes.
Fig. 3.
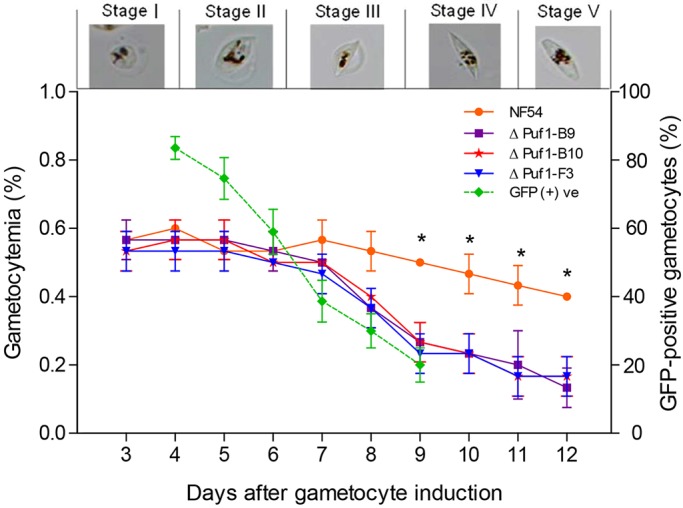
Gametocytogenesis after PfPuf1 disruption. Daily gametocytemia of three PfPuf1-disrupted clones (ΔPfPuf1-B9, ΔPfPuf1-B10 and ΔPfPuf1-F3) was determined by counting Giemsa-stained gametocytes under a microscope from day 4 through day 12 after induction. The proportion of GFP-positive gametocytes was determined on day 4–9 under an epifluorescence microscope. Images above the growth curve represent different stages of gametocyte development of P. falciparum. Data are shown in mean±s.d. from three replicates. *Significant differences in gametocytemia between the NF54 and ΔPfPuf1 parasites (P<0.05, Fisher's exact test).
PfPuf1 is required for maintenance of late-stage gametocytes
Next we wanted to determine whether PfPuf1 disruption differentially affects male and female gametocytes. Based on the similar course of gametocyte development among the three ΔPfPuf1-GFP clones, we randomly chose the B10 clone of ΔPfPuf1–GFP for further characterization. When gametocytes were examined under a fluorescence microscope, we observed a low- and a high-GFP population in the ΔPfPuf1-B10 clone on day 4, when parasitemia was not different between the three ΔPfPuf1-GFP clones and NF54 (Fig. 3). However, in subsequent days, gametocytes that were morphologically healthy but that lacked GFP fluorescence appeared and became more abundant as days went on (Fig. 4A). To further illustrate the consequence of PfPuf1 disruption on gametocyte development, we compared Giemsa-stained cultures of wild-type NF54 and the PfPuf1-disrupted line. The results showed increases in and accumulation of unhealthy or dying parasites in the ΔPfPuf1-GFP culture starting from stage III. These dying parasites gradually lost their membranous structures, leaving only cell debris at later stages of gametocytogenesis, and such cell debris was not observed after Percoll purification of gametocytes (Fig. S2). Concomitantly, the proportions of gametocytes with GFP fluorescence continued to decline from >80% on day 4 to ∼20% on day 9 (Fig. 3). This change in gametocyte fluorescence in the ΔPfPuf1-GFP clone was also reflected in quantitative analysis of Percoll-purified gametocytes with flow cytometry (Fig. 4B). There were mainly two populations of gametocytes – with low- or high-GFP fluorescence – on day 4 after gametocyte induction, whereas three gametocyte populations with no-, low- and high-GFP fluorescence were found in subsequent days (Fig. 4B). Consistent with the quantification under a fluorescence microscope, the non-fluorescent gametocyte population increased dramatically from day 6 and reached >80% in the Percoll-purified gametocyte population on day 12 (Fig. 4C). Conversely, the GFP-positive populations experienced a sharp decline as gametocytes matured (Fig. 4B,C). Transmission electron microscopy (TEM) examination of day 8 (stage IV) gametocytes showed that organelles and membranous structures – such as the erythrocyte membrane, parasitophorous vacuole membrane and parasite plasma membrane – were clearly visible in the healthy NF54 cells, whereas gametocytes from the PfPuf1-disrupted line, despite appearing morphologically normal with light microscopy, contained homogenous cytoplasmic content and ill-defined single-membrane vacuoles of different sizes. These vacuoles occupied the majority of the gametocyte cytoplasm. Some vacuoles contained hemozoin pigments and smaller vesicles. Additional abnormalities included discontinuity of the membrane of the infected erythrocytes. Some parasites lost the entire integrity of the membrane complex (Fig. S3). These abnormal parasites constituted 73.6% (81 out of 110) of the total gametocyte population in the PfPuf1-disrupted line on day 8. To further define these three gametocyte populations in the ΔPfPuf1-B10 clone, gametocytes from day-4 and day-8 cultures were sorted, and RNA was extracted for real-time RT-PCR analysis of α-tubulin II and Pfs47 expression. At both time points, the α-tubulin II:Pfs47 expression ratio was more than threefold higher in the low-GFP population than in the high-GFP population, consistent with the earlier indication of enriched male and female gametocytes in the low- and high-GFP populations, respectively (Fig. 4D). Analysis of three gametocyte populations by using an immunofluorescence assay to detect α-tubulin II further confirmed higher levels of α-tubulin II expression in the low-GFP parasites than the high-GFP parasites (Fig. 4E). To define the effect of PfPuf1 disruption on different sexes, day-12 (stage V) mature gametocytes of the ΔPfPuf1-GFP line were sorted and their sexes determined by performing microscopy. The results showed that >80% of high-GFP gametocytes were female, whereas >75% of low-GFP gametocytes were male, consistent with the α-tubulin II:Pfs47 expression ratio determined for these populations (Fig. S4A). The flow cytometry results also clearly showed a reverse of the sex ratio in the ΔPfPuf1-GFP line, which became male-biased (Fig. 4B). To determine whether PfPuf1 deletion affects the function of male gametocytes, we compared the exflagellation ability of the male gametocytes in both wild-type NF54 and PfPuf1-disrupted strains. The number of exflagellation centers formed in both lines on day 13, normalized to the number by male gametocytes, did not show a significant difference (P=0.795, t-test) (Fig. S4B).
Fig. 4.

Characterization of gametocyte populations after PfPuf1 disruption. (A) Microscopy images of P. falciparum gametocytes after PfPuf1 gene disruption. Gametocytes during their late stages (stage IV, day 8; and stage V, day 12) were observed under an epifluorescence microscope. Gametocytes were marked as no-, low- or high-GFP based on their GFP level. Blue arrows, three morphologically similar gametocytes (light-microscopy image); white arrows, the same three gametocytes showing high, low and no GFP fluorescence (fluorescence microscopy image). Scale bar: 5 μm. (B) Flow cytometry analysis from day-4, -6, -8 and -12 gametocytes to represent early- through late-stage gametocytes. The gametocytes were roughly divided into 2 or 3 populations (circled), which were individually sorted and used for analysis in D. FSC, forward scatter channel. (C) Analysis of three GFP populations of gametocytes with flow cytometry. (D) Relative transcript abundance of male and female gametocyte markers in GFP-positive populations. Male-to-female ratio, represented by the α-Tubulin II:Pfs47 ratio (α-Tubulin II/Pfs47), was analyzed by performing real-time RT-PCR analysis of early- (stage II, day 4) and late-stage (stage IV, day 8) gametocytes. The ratio was significantly different between low- and high-GFP gametocyte populations (P<0.05, t-test). (E) GFP fluorescence was analyzed by performing immunofluorescence analysis with an antibody against α-tubulin II. DIC, differential interference contrast microscopy. Scale bar: 5 μm. (F) Fluorescence intensities of gametocytes after staining with Mitotracker RedFM. Gametocytes at day 4, 8 and 12 were treated with Mitotracker RedFM and quantified under an epifluorescence microscope. The numbers of Mitotracker-RedFM-negative gametocytes in the PfPuf1-disrupted line were significantly different from those in control NF54 parasites (P<0.0001, Fisher's exact-test). Data are mean±s.d. from three replicates.
In contrast, the α-tubulin II, Pfs47 and reference gene could not be amplified with the RNA isolated from the non-fluorescent gametocytes, suggesting that RNA from this gametocyte population had been degraded. Consistently, indirect immunofluorescence assay did not detect α-tubulin II expression in parasites that lacked GFP signals (Fig. 4E). The lack of mRNAs or proteins for several gametocyte markers in these non-fluorescent parasites suggested that they might represent dead and/or dying gametocytes. To verify this observation, Mitotracker RedFM, which labels parasites with active mitochondria, was used to differentiate live and dead parasites. Whereas both low- and high-GFP gametocytes were labeled with Mitotracker RedFM, the non-fluorescent gametocytes and gametocytes that had been killed upon treatment with carbonyl cyanide m-chlorophenylhydrazone (CCCP), which induces mitochondrial permeability transition, were not labeled with Mitotracker RedFM (Fig. S4C). Further, flow cytometry analysis of gametocytes on day 4, 8 and 12 after gametocyte induction showed that the numbers of Mitotracker-Red-negative cells (presumably dead or dying) were significantly higher in B10 clones of ΔPfPuf1-GFP than in control NF54 cells, and this cell population reached almost 60% in day 12 gametocytes (P<0.0001, Fisher's exact-test) (Fig. 4F). These findings, collectively, were consistent with significantly elevated death rates in later-stage gametocytes in the ΔPfPuf1-GFP clones, which corresponded to the lower gametocytemia observed by microscopy (Fig. 3). If these dead cells were excluded, PfPuf1 disruption led to a more than tenfold reduction of mature gametocytes (Fig. 3). Altogether, these results indicate that PfPuf1 is important for maintaining gametocyte viability in later stages of development, and that this effect is more crucial for female gametocytes.
DISCUSSION
In this study, we functionally characterized a member of the Puf protein family, PfPuf1, in sexual development of P. falciparum. The GFP-tagged PfPuf1 protein was expressed in all gametocyte stages, and both transcript as well as protein expression levels were increased during gametocyte development. Disruption of PfPuf1 resulted in a significant loss of gametocytes and was reflected in much reduced gametocytemia during the late stages of gametocytogenesis. Consistent with expression of PfPuf1 at a higher level in female gametocytes, PfPuf1 disruption led to a more severe developmental defect in female gametocytes.
In malaria parasites, translation regulation is especially crucial during the development of specialized transition stages, gametocytes and sporozoites (Cui et al., 2015). Transcripts to be used in subsequent stages are deposited in these transition stages in translationally repressed forms. Translation repression of these target mRNAs is often mediated by RBPs, such as DOZI and the Puf-family proteins. In P. berghei, the translation repressor complex of the Dhh1 RNA helicase DOZI with CAR-I/trailer hitch homolog (CITH) is essential for protecting and silencing the stored mRNAs in female gametocytes (Mair et al., 2006, 2010). A recent RNA immunoprecipitation and microarray analysis revealed a large repertoire of 731 mRNAs to be associated with the DOZI–CITH complex, and the stored mRNA pool enables the zygote–ookinete developmental transition without requiring de novo transcription (Guerreiro et al., 2014). Although the functions of the DOZI–CITH complex in P. falciparum remain to be characterized, at least four such translationally repressed mRNAs have been found to be controlled by a Puf-family member (Miao et al., 2013).
Many eukaryotes contain multiple Puf members with different functions (Spassov and Jurecic, 2003). In Plasmodium, two evolutionarily conserved Puf proteins, Puf1 and Puf2, are differentially expressed in gametocytes (Cui et al., 2002; Fan et al., 2004). In P. falciparum, these two Puf proteins appeared to play distinct functions in gametocytogenesis. Their differences in the residues that are predicted to mediate RNA binding in the RBDs suggest that they recognize different subsets of target genes. Functional studies of PfPuf2 have shown that its disruption promotes gametocytogenesis and increases the male-to-female sex ratio (Miao et al., 2010b). In comparison, an opposite effect was observed here, where PfPuf1 disruption led to increased death of gametocytes and reduced gametocytemia, especially at the late stages of gametocytogenesis. With stronger effects on female gametocytes, PfPuf1 disruption resulted in more considerable losses of the female gametocytes and a reversal in the sex ratio. However, the two Puf proteins in the rodent parasites play different roles, probably reflecting their substantial evolutionary divergence from the human malaria parasites. Puf1 deletion does not result in noticeable changes in gametocytogenesis in P. berghei (Muller et al., 2011). Intriguingly, in one study, Puf2 knockout in P. berghei altered the gametocyte sex ratio, as observed in P. falciparum (Muller et al., 2011), whereas sex ratio was unchanged with Puf2 knockout in either P. berghei (Gomes-Santos et al., 2011) or P. yoelii (Lindner et al., 2013). The functions of Puf2 during sporozoite development appear to be conserved in these two rodent parasite species (Gomes-Santos et al., 2011; Lindner et al., 2013). Puf2 deletion in both rodent parasite species resulted in the loss of infectivity and in premature transformation of sporozoites to exoerythrocytic forms, though such changes occurred much earlier during sporogony in P. berghei than in P. yoelii. In the present study, it was not possible to examine the functions of PfPuf1 in sporozoites because mature gametocytes in the PfPuf1-disrupted lines were too low to infect mosquitoes.
The lengthy gametocytogenesis process in P. falciparum allowed us to observe the changes that were due to PfPuf1 disruption in greater detail. Consistent with the gradual increase in the expression level of PfPuf1 during gametocytogenesis, gametocyte development showed sharper declines in PfPuf1-disrupted lines in later stages (after stage III). This was accompanied by a rapid increase in cell population that exhibited normal morphology but lacked GFP fluorescence. The lack of Mitotracker RedFM staining in the non-fluorescent cells indicated the loss of mitochondrial integrity in these cells and is consistent with them being dead or dying gametocytes. These dead cells having the morphology of mature gametocytes were especially abundant during late gametocyte development in the PfPuf1-disrupted lines. This suggests that PfPuf1 disruption does not prevent maturation of gametocytes but rather that it interferes with the maintenance of later-stage gametocytes. Given that most Puf proteins function as translation repressors (Wickens et al., 2002), the loss of PfPuf1 probably leads to de-repression of target mRNAs and premature expression of proteins, resulting in the death of the gametocytes. Identification of the target mRNAs for the two Puf proteins in malaria parasites will help to decipher the exact molecular mechanisms of Puf regulation during gametocytogenesis.
MATERIALS AND METHODS
Parasite culture, gametocytogenesis and exflagellation
Parasite cultures were maintained as described previously (Trager and Jensen, 1976) and synchronized with sorbitol treatment (Lambros and Vanderberg, 1979). A modified method of gametocyte induction was followed to obtain highly synchronous gametocyte cultures (Fivelman et al., 2007). Gametocytemia was determined by counting gametocytes in at least 5000 red blood cells (RBCs) on Giemsa-stained thin smears for each treatment. Asexual erythrocytic development was followed as previously described (Miao et al., 2010b). Exflagellation assays were performed as described previously (Miao et al., 2010b).
Plasmid construction and parasite transfection
To tag the endogenous PfPuf1 protein with GFP, a 3′ fragment (nucleotides 4634–5682) of the PfPuf1 gene (Pf3D7_0518700) was PCR amplified from P. falciparum genomic DNA by using primers Puf1 F1 and Puf1 R1 (Table S1) cloned into a modified pBluescript plasmid to fuse the fragment with GFP and with pDT3′UTR (Fan et al., 2009). This 3′ PfPuf1 cassette was subcloned into pHD22Y between BamH1 and Not1 sites to produce the plasmid pHD22Y-PfPuf1-GFP. For disruption of PfPuf1, a 5′ fragment of the PfPuf1 coding region (nucleotides 79–1353) was amplified using primers ΔPfPuf1-GFPF and ΔPfPuf1-GFPR (Table S1) and cloned into the modified pBluescript plasmid to fuse the fragment with GFP and pDT3′UTR. The 5′ PfPuf1 cassette was subcloned into pHD22Y to produce the plasmid pHD22Y-ΔPfPuf1-GFP. Parasite transfection was performed by electroporation of 100 µg of plasmid DNA using the RBC-loading method (Deitsch et al., 2001). Transfected parasites were selected and cloned as described before (Miao et al., 2010a,b).
Southern blot analysis
To analyze the integration events in the transfected parasites, Southern blot was performed as described previously (Miao et al., 2010b). Briefly, genomic DNA was digested with HindIII and XbaI, and separated on a 0.8% agarose gel. Then, the DNA was transferred to a nylon membrane, denatured and UV-crosslinked using a Stratalinker 2400 instrument (Stratagene). A 506-bp PfPuf1 genomic DNA PCR fragment (nucleotides 168–674), obtained by using primers PfPuf1-probe F and PfPuf1-probe R (Table S1), was labeled with DIG using Probe synthesis kit (Roche) according to the manufacturer's protocol. The membrane was pre-hybridized in a pre-hybridization buffer, purchased from Roche Diagnostics, containing denatured salmon sperm DNA for 1 h at 42°C and hybridized with a denatured probe for 12 h at 42°C. After washing with 1× washing buffer (Roche Diagnostics), the membrane was exposed to a Kodak X-ray film at room temperature for 10 min for autoradiography.
Genomic DNA PCR and RT-PCR
Genomic DNA PCR was performed to confirm the correct integration for GFP tagging and disruption of PfPuf1 (Table S1). Total RNA was extracted from the parasites using the Trizol reagent (Invitrogen) and treated with RNase-free DNAse I (Promega) to remove contaminating genomic DNA. One microgram of total RNA was used to synthesize cDNA using oligo-dT primer in a 20-µl total reaction volume. RT-PCR was performed with primers Puf1 PD F2 and Puf1 PD R2 to detect the expression of PfPuf1 3′ region in PfPuf1-disrupted clones and NF54 control (Table S1). Real-time RT-PCR analysis was performed to determine the expression level of sex-specific markers α-tubulin II and Pfs47 for male and female gametocytes, by using primers α-tubulin II F or α-tubulin II R and Pfs47 F or Pfs47 R, respectively, and to detect the expression of PfPuf1 in a GFP-tagged line (Table S1). The constitutively expressed Pf07_0073 gene was used as a control by using primers Pf07_0073 F and Pf07_0073 R (Table S1). The relative expression level of each gene was calculated as described previously using the ΔCt method (Miao et al., 2006).
Quantification of PfPuf1 during gametocyte development
To analyze PfPuf1 protein during gametocyte development in P. falciparum, day-4, -8 and -12 gametocytes of the PfPuf1–GFP line were purified on a 35 and 65% Percoll step gradient. The lower interface containing infected RBCs with gametocytes was collected into a fresh tube, washed with cold PBS (pH 7.4) and lysed with 10% saponin. The freed gametocytes were washed with cold PBS and resuspended in cold PBS containing protease inhibitors. The cells were then subjected to protein extraction by sonication for 20 s on and 30 s off for five cycles on ice using a biorupter (Microson Ultrasonic cell disruptor). Protein concentration was determined by using the Micro BCA™ protein assay kit (Thermo Scientific) using bovine serum albumin (BSA) as the standard. PfPuf1-GFP expression was quantified by using iELISA. Gametocyte lysates were first diluted to 10 µg ml−1 protein concentrations in a coupling buffer containing 0.1 M sodium carbonate-bicarbonate (pH 9.6). Flat-bottomed 96-well plates (Becton Dickinson Labware) were coated with the diluted proteins at 4°C overnight. The coated wells were washed three times with PBS containing 0.05% Tween-20 (PBST) at room temperature and blocked with a blocking buffer (3% non-fat dry milk in PBST) for 1 h at 37°C. The primary antibody against GFP (cat. no. A-11122, Thermo-Fisher Scientific) at 1:1000 was added in each well and incubated for 1.5 h at 37°C, this was followed by three washes with PBST before adding the anti-rabbit horseradish peroxidase (HRP)-conjugated antibody (cat. no. A6154, Sigma-Aldrich) at 1:5000 for 1 h at 37°C. Subsequently, the wells were washed 5–7 times with PBST. The plates were developed with freshly prepared O-phenylenediamine dihydrochloride solution as the substrate (Sigma-Aldrich) for 5 min and 2 M H2SO4 was added to terminate the reaction. The absorbance was determined using a microplate reader (SpectraMax 250, Molecular device). Each iELISA was performed on three independent replicates, and the data were expressed as the mean optical density value.
Western blot
Immunoblotting was performed to detect PfPuf1–GFP protein expression. For this, protein from an equal number of mature gametocytes (106) were separated in an 8% SDS-PAGE gel and transferred to a nitrocellulose membrane, which was then probed with monoclonal antibody against GFP (Roche) at 1:1000, followed by detection with anti-rabbit HRP-conjugated antibody (Sigma-Aldrich) at 1:5000. The detected proteins were visualized using an enhanced chemiluminescence ECL kit (Invitrogen).
Indirect immunofluorescence assay
The expression of PfPuf1–GFP and its subcellular localization were directly visualized using the GFP signal under a Nikon ECLIPSE E600 epifluorescence microscope. The immunofluorescence assay using an antibody against male-specific α-tubulin II antibody (1:500) and gametocyte-specific Pfs230 (1:500) was performed based on a previously described method (Tonkin et al., 2004) with nuclei counterstained with Hoechst 33342. Both anti-α-tubulin-II (MRA-37) and anti-Pfs230 (MRA-27) antibodies were obtained from MR4 (Malaria Research and Reference Reagent Center). To check the viability of parasites, the carbocyanine-based Mitotracker RedFM dye (Molecular Probes) was used to stain the mitochondria of live gametocytes according to the manufacturer's instructions. Approximately 200 µl of the culture from ΔPfPuf1-GFP clones was centrifuged at 1000 g for 3 min at room temperature. The pellet was washed once with PBS and then resuspended gently in PBS containing 69 nM of the staining solution. The sample was incubated at 37°C for 15 min in the dark, washed three times with PBS and observed under an epifluorescence microscope. As a negative control, the gametocyte suspension was treated with 2 µM of CCCP for 1 h at 37°C in the dark, followed by standard Mitotracker RedFM staining.
Flow cytometry analysis
Flow cytometry was performed on a Beckman Coulter MoFlo Astrios (Brea, CA) system with 488-nm laser excitation. The GFP signal from cells was detected using a 513/26 nm bandpass detector. To discriminate between auto-fluorescence and true fluorescence, we used a phycoerythrin (PE)-Cy5 channel at 488 nm excitation with a 644/22 nm bandpass detector. To define green emission signals only from the cells, gametocytes from the PfPuf1–GFP line was used to define the gate, whereas gametocytes from NF54 with no signal at the initial fluorescence gate were used as the negative control. Data were collected with 150,000 events per sample.
Transmission electron microscopy
Mature gametocytes at stage IV on day 8 from NF54 wild-type and PfPuf1-disrupted lines were purified on a 35 and 65% Percoll step gradient followed by concentration using a magnetic column. The cells were then fixed with 4% paraformaldehyde in PBS for 30 min at room temperature and washed once with PBS. Then the cells were mixed with a fixative of 1.5% glutaraldehyde and 0.5% tannic acid in 0.05 M cacodylate buffer (pH 7.2). The cells were then centrifuged at 3000 rpm for 10 min and overlaid with 5 µl of non-infected human red blood cells, and incubated in the same fixative for another 2 h. The cells were then post-fixed in 1% osmium tetroxide for 2 h and en bloc stained in 2% aqueous uranyl acetate for 1 h. The cells were embedded in Eponite 12 (Ted Pella, CA). The sections were contrasted with uranyl acetate and lead citrate, and examined with a JEOL JEM1200 EXII transmission electron microscope (Peabody, MA).
Statistical analysis
Flow cytometry data was analyzed by using FlowJo software version 10. All assays were performed in three independent replicates and t-tests were performed in Microsoft Excel and GraphPad Prism 5.0. t-tests were used to compare specific gene expression levels between two GFP populations, and gametocytemia and exflagellation between the wild-type and PfPuf1-disrupted parasites. Fisher's exact test was used to compare total gametocytemia between the wild-type and PfPuf1-disrupted parasite lines. A P value of <0.05 was considered significant.
Acknowledgements
We would like to thank Dr Zenglei Wang for helping with the statistical analysis.
Footnotes
Competing interests
The authors declare no competing or financial interests.
Author contributions
S.S. performed most experiments except parasite transgenics, and drafted the manuscript. X.L. helped with parasite culture. G.N. assisted with the EM study. J.M. performed the parasite transfection and revised the manuscript. L.C. conceived the study and reviewed drafts of the manuscript. All authors read and approved the final manuscript.
Funding
This work was funded by National Institute of Allergy and Infectious Diseases, National Institutes of Health [grant numbers R01AI104946 and U19AI089672]. Deposited in PMC for release after 12 months.
Supplementary information
Supplementary information available online at http://jcs.biologists.org/lookup/doi/10.1242/jcs.186908.supplemental
References
- Archer S. K., Luu V.-D., de Queiroz R. A., Brems S. and Clayton C. (2009). Trypanosoma brucei PUF9 regulates mRNAs for proteins involved in replicative processes over the cell cycle. PLoS Pathog. 5, e1000565 10.1371/journal.ppat.1000565 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carter R. and Graves P. M. (1988). Gametocytes. In Malaria: Principles and Practice of Malariology, Vol. 1 (ed. Wernsdorfer W. H. and Sir McGregor I.), pp. 253-305. London: Churchill Livingstone. [Google Scholar]
- Coulson R. M. R., Hall N. and Ouzounis C. A. (2004). Comparative genomics of transcriptional control in the human malaria parasite Plasmodium falciparum. Genome Res. 14, 1548-1554. 10.1101/gr.2218604 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cui L., Fan Q. and Li J. (2002). The malaria parasite Plasmodium falciparum encodes members of the Puf RNA-binding protein family with conserved RNA binding activity. Nucleic Acids Res. 30, 4607-4617. 10.1093/nar/gkf600 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cui L., Lindner S. and Miao J. (2015). Translational regulation during stage transitions in malaria parasites. Ann. N. Y. Acad. Sci. 1342, 1-9. 10.1111/nyas.12573 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deitsch K. W., Driskill C. L. and Wellems T. E. (2001). Transformation of malaria parasites by the spontaneous uptake and expression of DNA from human erythrocytes. Nucleic Acids Res. 29, 850-853. 10.1093/nar/29.3.850 [DOI] [PMC free article] [PubMed] [Google Scholar]
- del Carmen Rodriguez M., Gerold P., Dessens J., Kurtenbach K., Schwartz R. T., Sinden R. E. and Margos G. (2000). Characterisation and expression of Pbs25, a sexual and sporogonic stage specific protein of Plasmodium berghei. Mol. Biochem. Parasitol. 110, 147-159. 10.1016/S0166-6851(00)00265-6 [DOI] [PubMed] [Google Scholar]
- Edwards T. A., Pyle S. E., Wharton R. P. and Aggarwal A. K. (2001). Structure of Pumilio reveals similarity between RNA and peptide binding motifs. Cell 105, 281-289. 10.1016/S0092-8674(01)00318-X [DOI] [PubMed] [Google Scholar]
- Fan Q., Li J., Kariuki M. and Cui L. (2004). Characterization of PfPuf2, member of the Puf family RNA-binding proteins from the malaria parasite Plasmodium falciparum. DNA Cell Biol. 23, 753-760. 10.1089/dna.2004.23.753 [DOI] [PubMed] [Google Scholar]
- Fan Q., Miao J., Cui L. and Cui L. (2009). Characterization of PRMT1 from Plasmodium falciparum. Biochem. J. 421, 107-118. 10.1042/BJ20090185 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Farfour E., Charlotte F., Settegrana C., Miyara M. and Buffet P. (2012). The extravascular compartment of the bone marrow: a niche for Plasmodium falciparum gametocyte maturation? Malaria J. 11, 285 10.1186/1475-2875-11-285 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fivelman Q. L., McRobert L., Sharp S., Taylor C. J., Saeed M., Swales C. A., Sutherland C. J. and Baker D. A. (2007). Improved synchronous production of Plasmodium falciparum gametocytes in vitro. Mol. Biochem. Parasitol. 154, 119-123. 10.1016/j.molbiopara.2007.04.008 [DOI] [PubMed] [Google Scholar]
- Foth B. J., Zhang N., Chaal B. K., Sze S. K., Preiser P. R. and Bozdech Z. (2011). Quantitative time-course profiling of parasite and host cell proteins in the human malaria parasite Plasmodium falciparum. Mol. Cell. Proteomics 10, M110.006411 10.1074/mcp.m110.006411 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldstrohm A. C., Seay D. J., Hook B. A. and Wickens M. (2007). PUF protein-mediated deadenylation is catalyzed by Ccr4p. J. Biol. Chem. 282, 109-114. 10.1074/jbc.M609413200 [DOI] [PubMed] [Google Scholar]
- Gomes-Santos C. S. S., Braks J., Prudencio M., Carret C., Gomes A. R., Pain A., Feltwell T., Khan S., Waters A., Janse C. et al. (2011). Transition of Plasmodium sporozoites into liver stage-like forms is regulated by the RNA binding protein Pumilio. PLoS Pathog. 7, e1002046 10.1371/journal.ppat.1002046 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guerreiro A., Deligianni E., Santos J. M., Silva P. A. G. C., Louis C., Pain A., Janse C. J., Franke-Fayard B., Carret C. K., Siden-Kiamos I. et al. (2014). Genome-wide RIP-Chip analysis of translational repressor-bound mRNAs in the Plasmodium gametocyte. Genome Biol. 15, 493 10.1186/s13059-014-0493-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hall N., Karras M., Raine J. D., Carlton J. M., Kooij T. W. A., Berriman M., Florens L., Janssen C. S., Pain A., Christophides G. K. et al. (2005). A comprehensive survey of the Plasmodium life cycle by genomic, transcriptomic, and proteomic analyses. Science 307, 82-86. 10.1126/science.1103717 [DOI] [PubMed] [Google Scholar]
- Joice R., Nilsson S. K., Montgomery J., Dankwa S., Egan E., Morahan B., Seydel K. B., Bertuccini L., Alano P., Williamson K. C. et al. (2014). Plasmodium falciparum transmission stages accumulate in the human bone marrow. Sci. Transl. Med. 6, 244re5 10.1126/scitranslmed.3008882 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kafsack B. F. C., Rovira-Graells N., Clark T. G., Bancells C., Crowley V. M., Campino S. G., Williams A. E., Drought L. G., Kwiatkowski D. P., Baker D. A. et al. (2014). A transcriptional switch underlies commitment to sexual development in malaria parasites. Nature 507, 248-252. 10.1038/nature12920 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaye J. A., Rose N. C., Goldsworthy B., Goga A. and L'Etoile N. D. (2009). A 3′UTR pumilio-binding element directs translational activation in olfactory sensory neurons. Neuron 61, 57-70. 10.1016/j.neuron.2008.11.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuersten S. and Goodwin E. B. (2003). The power of the 3′ UTR: translational control and development. Nat. Rev. Genet. 4, 626-637. 10.1038/nrg1125 [DOI] [PubMed] [Google Scholar]
- Lambros C. and Vanderberg J. P. (1979). Synchronization of Plasmodium falciparum erythrocytic stages in culture. J. Parasitol. 65, 418-420. 10.2307/3280287 [DOI] [PubMed] [Google Scholar]
- Le Roch K. G., Zhou Y., Blair P. L., Grainger M., Moch J. K., Haynes J. D., De La Vega P., Holder A. A., Batalov S., Carucci D. J. et al. (2003). Discovery of gene function by expression profiling of the malaria parasite life cycle. Science 301, 1503-1508. 10.1126/science.1087025 [DOI] [PubMed] [Google Scholar]
- Le Roch K. G., Johnson J. R., Florens L., Zhou Y., Santrosyan A., Grainger M., Yan S. F., Williamson K. C., Holder A. A., Carucci D. J. et al. (2004). Global analysis of transcript and protein levels across the Plasmodium falciparum life cycle. Genome Res. 14, 2308-2318. 10.1101/gr.2523904 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lindner S. E., Mikolajczak S. A., Vaughan A. M., Moon W., Joyce B. R., Sullivan W. J. Jr and Kappe S. H. I. (2013). Perturbations of Plasmodium Puf2 expression and RNA-seq of Puf2-deficient sporozoites reveal a critical role in maintaining RNA homeostasis and parasite transmissibility. Cell. Microbiol. 15, 1266-1283. 10.1111/cmi.12116 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lopez-Barragan M. J., Lemieux J., Quinones M., Williamson K. C., Molina-Cruz A., Cui K., Barillas-Mury C., Zhao K. and Su X.-Z. (2011). Directional gene expression and antisense transcripts in sexual and asexual stages of Plasmodium falciparum. BMC Genomics 12, 587 10.1186/1471-2164-12-587 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mair G. R., Braks J. A. M., Garver L. S., Wiegant J. C. A. G., Hall N., Dirks R. W., Khan S. M., Dimopoulos G., Janse C. J. and Waters A. P. (2006). Regulation of sexual development of Plasmodium by translational repression. Science 313, 667-669. 10.1126/science.1125129 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mair G. R., Lasonder E., Garver L. S., Franke-Fayard B. M. D., Carret C. K., Wiegant J. C. A. G., Dirks R. W., Dimopoulos G., Janse C. J. and Waters A. P. (2010). Universal features of post-transcriptional gene regulation are critical for Plasmodium zygote development. PLoS Pathog. 6, e1000767 10.1371/journal.ppat.1000767 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mee C. J., Pym E. C. G., Moffat K. G. and Baines R. A. (2004). Regulation of neuronal excitability through pumilio-dependent control of a sodium channel gene. J. Neurosci. 24, 8695-8703. 10.1523/JNEUROSCI.2282-04.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Menon K. P., Sanyal S., Habara Y., Sanchez R., Wharton R. P., Ramaswami M. and Zinn K. (2004). The translational repressor Pumilio regulates presynaptic morphology and controls postsynaptic accumulation of translation factor eIF-4E. Neuron 44, 663-676. 10.1016/j.neuron.2004.10.028 [DOI] [PubMed] [Google Scholar]
- Miao J., Fan Q., Cui L., Li J., Li J. and Cui L. (2006). The malaria parasite Plasmodium falciparum histones: organization, expression, and acetylation. Gene 369, 53-65. 10.1016/j.gene.2005.10.022 [DOI] [PubMed] [Google Scholar]
- Miao J., Fan Q., Cui L., Li X., Wang H., Ning G., Reese J. C. and Cui L. (2010a). The MYST family histone acetyltransferase regulates gene expression and cell cycle in malaria parasite Plasmodium falciparum. Mol. Microbiol. 78, 883-902. 10.1111/j.1365-2958.2010.07371.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miao J., Li J., Fan Q., Li X., Li X. and Cui L. (2010b). The Puf-family RNA-binding protein PfPuf2 regulates sexual development and sex differentiation in the malaria parasite Plasmodium falciparum. J. Cell Sci. 123, 1039-1049. 10.1242/jcs.059824 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miao J., Fan Q., Parker D., Li X., Li J. and Cui L. (2013). Puf mediates translation repression of transmission-blocking vaccine candidates in malaria parasites. PLoS Pathog. 9, e1003268 10.1371/journal.ppat.1003268 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Muller K., Matuschewski K. and Silvie O. (2011). The Puf-family RNA-binding protein Puf2 controls sporozoite conversion to liver stages in the malaria parasite. PLoS ONE 6, e19860 10.1371/journal.pone.0019860 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Muraro N. I., Weston A. J., Gerber A. P., Luschnig S., Moffat K. G. and Baines R. A. (2008). Pumilio binds para mRNA and requires Nanos and Brat to regulate sodium current in Drosophila motoneurons. J. Neurosci. 28, 2099-2109. 10.1523/JNEUROSCI.5092-07.2008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Olivas W. and Parker R. (2000). The Puf3 protein is a transcript-specific regulator of mRNA degradation in yeast. EMBO J. 19, 6602-6611. 10.1093/emboj/19.23.6602 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Otto T. D., Wilinski D., Assefa S., Keane T. M., Sarry L. R., Böhme U., Lemieux J., Barrell B., Pain A., Berriman M. et al. (2010). New insights into the blood-stage transcriptome of Plasmodium falciparum using RNA-Seq. Mol. Microbiol. 76, 12-24. 10.1111/j.1365-2958.2009.07026.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Painter H. J., Campbell T. L. and Llinas M. (2011). The Apicomplexan AP2 family: integral factors regulating Plasmodium development. Mol. Biochem. Parasitol. 176, 1-7. 10.1016/j.molbiopara.2010.11.014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paton M. G., Barker G. C., Matsuoka H., Ramesar J., Janse C. J., Waters A. P. and Sinden R. E. (1993). Structure and expression of a post-transcriptionally regulated malaria gene encoding a surface protein from the sexual stages of Plasmodium berghei. Mol. Biochem. Parasitol. 59, 263-275. 10.1016/0166-6851(93)90224-L [DOI] [PubMed] [Google Scholar]
- Pique M., Lopez J. M., Foissac S., Guigo R. and Mendez R. (2008). A combinatorial code for CPE-mediated translational control. Cell 132, 434-448. 10.1016/j.cell.2007.12.038 [DOI] [PubMed] [Google Scholar]
- Schwank S., Sutherland C. J. and Drakeley C. J. (2010). Promiscuous expression of alpha-tubulin II in maturing male and female Plasmodium falciparum gametocytes. PLoS ONE 5, e14470 10.1371/journal.pone.0014470 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sinden R. E. (1998). Gametocytes and sexual development. In Malaria: Parasite Biology, Pathogenesis and Protection (ed. Sherman I. W.), pp. 25-48: ASM Press; Washington DC, USA. [Google Scholar]
- Sinha A., Hughes K. R., Modrzynska K. K., Otto T. D., Pfander C., Dickens N. J., Religa A. A., Bushell E., Graham A. L., Cameron R. et al. (2014). A cascade of DNA-binding proteins for sexual commitment and development in Plasmodium. Nature 507, 253-257. 10.1038/nature12970 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spassov D. S. and Jurecic R. (2003). The PUF family of RNA-binding proteins: does evolutionarily conserved structure equal conserved function? IUBMB Life 55, 359-366. 10.1080/15216540310001603093 [DOI] [PubMed] [Google Scholar]
- Suh N., Crittenden S. L., Goldstrohm A., Hook B., Thompson B., Wickens M. and Kimble J. (2009). FBF and its dual control of gld-1 expression in the Caenorhabditis elegans germline. Genetics 181, 1249-1260. 10.1534/genetics.108.099440 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tonkin C. J., van Dooren G. G., Spurck T. P., Struck N. S., Good R. T., Handman E., Cowman A. F. and McFadden G. I. (2004). Localization of organellar proteins in Plasmodium falciparum using a novel set of transfection vectors and a new immunofluorescence fixation method. Mol. Biochem. Parasitol. 137, 13-21. 10.1016/j.molbiopara.2004.05.009 [DOI] [PubMed] [Google Scholar]
- Trager W. and Jensen J. B. (1976). Human malaria parasites in continuous culture. Science 193, 673-675. 10.1126/science.781840 [DOI] [PubMed] [Google Scholar]
- Wharton R. P., Sonoda J., Lee T., Patterson M. and Murata Y. (1998). The Pumilio RNA-binding domain is also a translational regulator. Mol. Cell 1, 863-872. 10.1016/S1097-2765(00)80085-4 [DOI] [PubMed] [Google Scholar]
- WHO. (2015). World Malaria Report. Geneva: World Health Organization. [Google Scholar]
- Wickens M., Bernstein D. S., Kimble J. and Parker R. (2002). A PUF family portrait: 3′UTR regulation as a way of life. Trends Genet. 18, 150-157. 10.1016/S0168-9525(01)02616-6 [DOI] [PubMed] [Google Scholar]
- Ye B., Petritsch C., Clark I. E., Gavis E. R., Jan L. Y. and Jan Y. N. (2004). Nanos and Pumilio are essential for dendrite morphogenesis in Drosophila peripheral neurons. Curr. Biol. 14, 314-321. 10.1016/j.cub.2004.01.052 [DOI] [PubMed] [Google Scholar]
- Young J. A., Fivelman Q. L., Blair P. L., de la Vega P., Le Roch K. G., Zhou Y., Carucci D. J., Baker D. A. and Winzeler E. A. (2005). The Plasmodium falciparum sexual development transcriptome: a microarray analysis using ontology-based pattern identification. Mol. Biochem. Parasitol. 143, 67-79. 10.1016/j.molbiopara.2005.05.007 [DOI] [PubMed] [Google Scholar]