

Abstract
Arctic marine mammals (AMMs) are icons of climate change, largely because of their close association with sea ice. However, neither a circumpolar assessment of AMM status nor a standardized metric of sea ice habitat change is available. We summarized available data on abundance and trend for each AMM species and recognized subpopulation. We also examined species diversity, the extent of human use, and temporal trends in sea ice habitat for 12 regions of the Arctic by calculating the dates of spring sea ice retreat and fall sea ice advance from satellite data (1979–2013). Estimates of AMM abundance varied greatly in quality, and few studies were long enough for trend analysis. Of the AMM subpopulations, 78% (61 of 78) are legally harvested for subsistence purposes. Changes in sea ice phenology have been profound. In all regions except the Bering Sea, the duration of the summer (i.e., reduced ice) period increased by 5–10 weeks and by >20 weeks in the Barents Sea between 1979 and 2013. In light of generally poor data, the importance of human use, and forecasted environmental changes in the 21st century, we recommend the following for effective AMM conservation: maintain and improve comanagement by local, federal, and international partners; recognize spatial and temporal variability in AMM subpopulation response to climate change; implement monitoring programs with clear goals; mitigate cumulative impacts of increased human activity; and recognize the limits of current protected species legislation.
Keywords: circumpolar assessment, climate change, management, subsistence harvest, cambio climático, caza para la subsistencia, evaluación circumpolar, manejo
Abstract
Estado de las Poblaciones de Mamíferos Marinos del Ártico, la Pérdida del Hábitats de Hielo Marino y Recomendaciones de Conservación para el Siglo XXI
Resumen
Los mamíferos marinos del Ártico (MMA) son emblemas del cambio climático, principalmente por su asociación cercana con el hielo marino. Sin embargo, no se encuentran disponibles ni una evaluación circumpolar del estado de los MMA ni una medida estandarizada del cambio en el hábitat de hielo marino. Resumimos los datos disponibles sobre la abundancia y la tendencia de cada especie de MMA y reconocimos subpoblaciones. También examinamos la diversidad de especies, la extensión del uso por parte de los humanos y las tendencias temporales en el hábitat de hielo marino para doce regiones del Ártico al calcular las fechas del retroceso de los hielos en la primavera y de su avance en el otoño a partir de datos satelitales (1979–2013). Los estimados de abundancia de MMA variaron enormemente en calidad y pocos estudios fueron lo suficientemente largos como para realizar un análisis de tendencia. De las subpoblaciones de MMA, el 78% (61 de 78) son cazadas legalmente por razones de subsistencia. Los cambios en la fenología del hielo marino han sido profundos. En todas las regiones, salvo el Mar de Bering, la duración del periodo de verano (es decir, la reducción del hielo) incrementó por 5–10 semanas y por >20 semanas en el Mar de Barents entre 1979 y 2013. A razón de los datos generalmente pobres, la importancia del uso por parte de los humanos y los cambios ambientales pronosticados para el Siglo XX1, recomendamos lo siguiente para la conservación efectiva de los MMA: mantener y mejorar el co‐manejo por parte de los socios locales, federales e internacionales; reconocer la variabilidad temporal y espacial en la respuesta de las sub‐poblaciones de MMA al cambio climático; implementar el monitoreo de programas con objetivos claros; mitigar los impactos acumulativos del incremento de la actividad humana; y reconocer los límites de la legislación actual para las especies protegidas.
Introduction
The world's marine mammals are disproportionately threatened and data poor compared with their terrestrial counterparts (Schipper et al. 2008), and the 11 species of arctic marine mammals (AMMs) are particularly vulnerable due to their dependence on sea ice (Laidre et al. 2008 a; Kovacs et al. 2012; Reid & Laidre 2013). Some AMMs are sea ice obligates, meaning their life history events (e.g., reproduction, molting, resting) and feeding depend on sea ice, whereas others use ice but do not depend on it completely (Laidre et al. 2008 a). By AMMs we mean species that occur north of the Arctic Circle (66° 33′ N) for most of the year and depend on the Arctic marine ecosystem for all aspects of life and selected species that seasonally inhabit Arctic waters but may live outside the Arctic for part of the year. AMMs include 3 cetaceans (narwhal [Monodon monoceros], beluga [Delphinapterus leucas], and bowhead [Balaena mysticetus] whales); 7 pinnipeds (ringed [Pusa hispida], bearded [Erignathus barbatus], spotted [Phoca largha], ribbon [Histriophoca fasciata], harp [Pagophilus groenlandicus], and hooded [Cystophora cristata] seals and walrus [Odobenus rosmarus]); and the polar bear (Ursus maritimus). Throughout much of their range, these animals are important cultural and nutritional resources for indigenous and nonindigenous peoples. Recent reviews outline AMM vulnerabilities to climate change (e.g., Laidre et al. 2008 a; Kovacs et al. 2011).
Warming in the Arctic over the past few decades has been about 2 times greater than the global mean (IPCC 2013). The rate of loss of Arctic sea ice is faster than predicted by climate models (Stroeve et al. 2012), and projections suggest an ice‐free Arctic in summer by 2040 (Overland & Wang 2013). Even if greenhouse gases, the primary driver of climate change, are limited immediately, sea ice loss is likely to continue for several decades (Overland & Wang 2013). Therefore, it appears that continued unprecedented changes in AMM habitats are inevitable.
We reviewed what is known about population abundance, trends in abundance, and sea ice habitat loss for each AMM subpopulation or recognized stock. We evaluated species richness across 12 regions and quantified the extent of human subsistence use. We also assessed trends in the dates of spring sea ice retreat and fall sea ice advance to provide the first comparative circumpolar measure of AMM habitat change. Based on our findings, we make recommendations for AMM conservation relative to data gaps, sea ice forecasts, anthropogenic activities, and the complex social, economic, and political context of a rapidly warming Arctic.
Methods
We compiled current estimates of AMM abundance and population trend data from published and unpublished sources. Subpopulations included are those recognized by management bodies and advisory groups such as the International Whaling Commission (IWC), the North Atlantic Marine Mammal Commission (NAMMCO), and International Union for the Conservation of Nature (IUCN) specialist groups. Trends and their associated time frames are those reported by advisory groups or authors as noted. We delineated 12 regions, modified slightly from the Conservation of Arctic Flora and Fauna (CAFF) Circumpolar Biodiversity Monitoring Plan (CAFF 2011) (Fig. 1). The central Arctic Basin was excluded due to a paucity of data. We calculated the number of species per region and quantified the extent of human use by determining whether each subpopulation had a legal subsistence or commercial harvest.
Figure 1.

Geographic regions important to assemblages of Arctic marine mammals and used for sea ice analyses. Subpopulations and species may cross multiple regional boundaries.
Habitat change was quantified by calculating the dates of spring sea ice retreat and fall sea ice advance in each region for 1979–2013 from daily sea ice concentration data from satellites (Supporting Information). The date of spring retreat in a given region was when the area of sea ice fell below a specific threshold, and the date of fall advance was when the area rose above that same threshold. We used a region‐specific threshold halfway between the mean March and mean September sea ice areas over a baseline decade (1982–1991) (Supporting Information). The spring and fall dates characterize the biologically important transitions between winter and summer sea ice conditions.
Results
Circumpolar Diversity, Abundance, and Trends of AMMs
The highest species richness of AMMs was in the Atlantic regions of Baffin Bay, Davis Strait, and the Barents Sea; the lowest species richness was in the Sea of Okhotsk and the Beaufort Sea. The availability and quality of AMM subpopulation abundance estimates reviewed here through 2015 varied widely (Table 1 & Fig. 2). In many cases, knowledge of abundance consisted of a single point estimate with large uncertainty or an estimate based on expert opinion without formal assessment of uncertainty or bias. For cetaceans, trend data were available for 5 of 19 beluga subpopulations, 0 of 11 narwhal subpopulations, and 2 of 4 bowhead subpopulations. Abundance estimates for ringed and bearded seal were poor and outdated, and trends were not available except for some small areas surveyed repeatedly for ringed seals. Abundance estimates for walrus were available in several areas; however, some estimates represented only a portion of a subpopulation and the discreteness of some subpopulations was uncertain. Trend data suggested harp seal abundance increased, whereas hooded seal abundance was stable or declined. Of the 19 subpopulations of polar bears, abundance estimates were available for 14, although many of the estimates were out of date or had large uncertainty. Current trends were available for 10 subpopulations, although several of these were derived from projection models with untested assumptions (Table 1).
Table 1.
Estimates of abundance and trend for subspecies and subpopulations of Arctic marine mammals (with associated estimate of uncertainty if available) from dedicated surveys, population viability analyses, or expert opinion
| Species | Subpopulation or stocka,b | Abundance (95% CI or coefficient of variation [CV] if available) | Year | Trend | Survey/trend referencea | Legal subsistence (S) or commercial (C) harvest |
|---|---|---|---|---|---|---|
| Beluga | E Siberian and W Chukchi Seas | unknown | unknown | S | ||
| Eastern Chukchi Sea | 3700 | 1992 | unknown | 1 | S | |
| Eastern Beaufort Sea | 39258 (CV 0.23) | 1999 | unknown | 2 | S | |
| Eastern Bering Sea | 18000 | 1989–1991 | unknown | 2 | S | |
| Bristol Bay | 2877 (CV 0.23) | 2005 | increasing | 3 | S | |
| Cook Inlet | 315 (CV 0.13) | 2011 | declining | 4 | none | |
| Western Hudson Bay | 57300 (CI 37700–87100) | 2004 | unknown | 5 | S | |
| James Bay | 14967 (CI 8316–26939) | 2011 | unknown | 6 | S | |
| Eastern Hudson Bay | 3351 (CI 1552–7855) | 2011 | stable | 6 | S | |
| St Lawrence Estuary | 979 (CV 0.14) | 2009 | declining | 7, 8 | none | |
| Ungava Bay | 32 (CI 0–94) | 2012 | unknown | 9 | S | |
| Cumberland Sound | 1547 (CI 1187–1970) | 2001 | unknown | 10 | S | |
| E high Arctic‐Baffin Bay | 21200 (CV 0.25) | 1996 | unknown | 11 | S | |
| West Greenland winter | 10595 (CI 4904–24650) | 2006 | unknown | 12 | S | |
| White Sea | 6498 (CI 4664–8818) | 2008 | declining | 13 | none | |
| Svalbard | unknown | unknown | 14 | none | ||
| Kara & Laptev Seas | unknown | unknown | none | |||
| Gulf of Anadyr | 15127 (CI 7447–30741) | 2006 | unknown | 15 | S | |
| Okhotsk Sea | 12226 (CV 6.8) | 2010 | unknown | 16 | none | |
| Narwhal | Eclipse Sound | 20225 (CI 9471–37096) | 2004 | unknown | 17 | S |
| Admiralty Inlet | 18049 (CI 11613–28053) | 2010 | unknown | 17, 18 | S | |
| Somerset Island | 27656 (CI 9080–66061) | 2002 | unknown | 17 | S | |
| Jones Sound/Smith Sound | unknown | unknown | S | |||
| E Baffin Island fjords | 10073 (CI 5333–17474) | 2003 | unknown | 17 | S | |
| Northern Hudson Bay | 12485 (CV 0.26) | 2011 | unknown | 19, 20 | S | |
| Inglefield Bredning, W Greenland | 8368 (CI 5209–13442) | 2007 | unknown | 21 | S | |
| Melville Bay, W Greenland | 6024 (CI 1403–25860) | 2007 | unknown | 21 | S | |
| W Greenland winter aggregation | 7819 (CI 4358–14029) | 2006 | unknown | 21 | S | |
| E Greenland | 6444 (CI 2505–16575) | 2008 | unknown | 21 | S | |
| Svalbard | unknown | unknown | none | |||
| Bowhead | Bering‐Chukchi‐Beaufort Seas (BCB) | 16892 (CI 15704–18928) | 2011 | increasing | 22 | S |
| E Canada‐W Greenland (BBDS and FBHB) | >6500 | 2002‐2009 | increasing | 23 | S | |
| Svalbard‐Barents Sea | <100 | – | unknown | 24 | none | |
| Okhotsk Sea | <400 | 1979 | unknown | 25 | none | |
| Ringed seal | Arctic subspecies | unknown total | unknown | S | ||
| ‐Beaufort & Chukchi seas | 1000000 | unspecified | unknown | 26 | ||
| ‐Bering Sea | 340000 | 1976‐2012 | unknown | 27, 28 | ||
| ‐Hudson and James Bay | 516000 | 1974 | unknown | 29 | ||
| ‐Baffin Bay | 787000 | 1979 | unknown | 30 | ||
| ‐Portion of Greenland Sea/Spitsbergen | 7585 (CI 6332–9085) | 2002 | unknown | 31 | ||
| ‐Svalbard | 6332–9085 | 2001 | unknown | 31 | ||
| ‐White, Barents, Kara, and East Siberian seas | 220000 | unspecified | unknown | 26 | ||
| Okhotsk Sea subspecies | 676000–855000 | 1971 | unknown | 27 | S | |
| Bearded seal | E. nauticus subspecies | unknown total | S | |||
| ‐Sea of Okhotsk | 200000–250000 | 1968–1969 | unknown | 27 | ||
| ‐Bering Sea | >299000 | 2012 | unknown | 28 | ||
| ‐Chukchi Sea | 27000 | 2000 | unknown | 32 | ||
| ‐Beaufort Sea | unknown | unknown | ||||
| ‐E. Siberian Sea | unknown | unknown | ||||
| E. barbatus subspecies | unknown total | S | ||||
| ‐Canadian waters | 190000 | 1958–1979 | unknown | 33 | ||
| ‐Greenland | unknown | – | unknown | |||
| ‐Svalbard | unknown | unknown | ||||
| ‐Barents, White, Kara, and Laptev seas | unknown | unknown | ||||
| Spotted seal | Bering Sea | >460000 | 2012 | unknown | 31 | S |
| Sea of Okhotsk | 180000–240000 | 1968–1990 | unknown | 27 | S | |
| Yellow Sea and Sea of Japan | 3300 | 2007 | unknown | 34, 35 | none | |
| Ribbon seal | Bering Sea | 143000 | 1960s, 2007 | unknown | 36, 37 | S |
| Sea of Okhotsk | 124000 | 1960s, 2007 | unknown | 36, 37 | S | |
| Harp seal | Northwest Atlantic | 7400000 (CI 5000000–8000000) | 2008 | increasing | 38 | S (Greenland) and C (Canada) |
| Greenland Sea | 627410 (CI 470540–784280) | 2012 | increasing | 39 | S (Greenland) and C (Norway) | |
| White Sea | 1419800 (CI 1266910–1572690) | 2013 | increasing (or stable) | 39 | C (Norway and Russia) | |
| Hooded seal | Northwest Atlantic | 593500 (CI 465600–728300) | 2005 | stable | 40 | S (Greenland) and C (Canada) |
| Greenland Sea | 84020 (CI 68060–99980) | 2012 | decreasing | 39 | S | |
| Walrus | O. r. divergens subspecies (Pacific) | |||||
| Bering‐Chukchi Seas | ∼129000 (CI 55000–507000) | 2006 | unknown | 41 | S | |
| Laptev Sea | 3000–5000 | 1992 | unknown | 42 | none | |
| O. r. rosmarus subspecies (Atlantic) | ||||||
| North and Central Foxe Basin | 13452 (CV 0.43) | 2011 | unknown | 43 | S | |
| South and East Hudson Bay | low hundreds | 2006 | unknown | 44 | S | |
| N Hudson Bay‐Hudson Strait‐SE Baffin Island‐N Labrador (estimates for portions of the range below) | unknown | unknown | S | |||
| SE Baffin Island summer aggregation | 2502 (CI 1660–3345) | 2007 | 45 | |||
| N Hudson Bay summer aggregation | 1376 | 1990 | 44 | |||
| Hudson Strait winter aggregation | 6020 (CI 2485–14,85) | 2012 | 46 | |||
| West Greenland winter aggregation | 1408 (CI 922–2150) | 2012 | 47, 48 | |||
| West Jones Sound | 503 (CI 473–534) | 2008 | stable | 49 | S | |
| Penny Strait/Lancaster Sound | 727 (CI 623–831) | 2009 | stable | 49 | S | |
| Baffin Bay winter summer | 1499 (CI 1077–2087) | 2009‐2010 | increasing | 47, 50, 51 | S | |
| 1251 (CL 571–2477)b | 2009 | |||||
| East Greenland | 1429 (CI 705–2896) | 2009 | increasing | 47 | S | |
| Svalbard/Franz Josef Land | 2629 (CI 2318–2998) | 2006 | increasing | 52 | none | |
| Novaya Semlya‐Eastern Barents‐Pechora‐White Seas | 3943 (CI 3605‐4325) | 2010 | unknown | 53 | none | |
| Polar bear | Chukchi Sea | unknown | – | unknown | 54, 55 | S |
| Southern Beaufort Sea | 900 (CI 606–1212)c | 2010 | declining | 56 | S | |
| Northern Beaufort Sea | 980 (CI 825–1135) | 2006 | stable | 54, 57 | S | |
| Viscount Melville | 161 (CI 121–201) | 1992 | unknown | 54, 58 | S | |
| M'Clintock Channel | 284 (CI 166–402) | 2000 | increasing | 54, 59 | S | |
| Lancaster Sound | 2541 (CI 1759–3323) | 1997 | unknown | 54, 60 | S | |
| Norwegian Bay | 203 (CI 115–291) | 1997 | unknown | 54, 60 | S | |
| Gulf of Boothia | 1592 (CI 870–2314) | 2000 | stable | 54, 61 | S | |
| Foxe Basin | 2580 (CI 2093–3180) | 2010 | stable | 54, 62 | S | |
| Western Hudson Bay | 1030 (CI 754–1406) | 2011 | seclining | 63, 64, 65 | S | |
| Southern Hudson Bay | 970 (CI 680–1383) | 2005 | stable | 54, 66 | S | |
| Kane Basin | 164 (CI 94–234) | 1998 | declining | 54, 67 | S | |
| Baffin Bay | 1546 (CI 690–2402) | 2004 | declining | 54, 68 | S | |
| Davis Strait | 2158 (CI 1833–2542) | 2007 | stable | 54, 69 | S | |
| East Greenland | unknown | – | unknown | 54 | S | |
| Barents Sea | 2644 (CI 1899‐3592) | 2004 | unknown | 54, 70 | none | |
| Kara Sea | unknown | – | unknown | 54 | none | |
| Laptev Sea | unknown | 1993 | unknown | 54 | none | |
| Arctic Basin | unknown | – | unknown | 54 | none |
In some cases, estimates represent only a portion of the subpopulation (e.g., aggregation within a subpopulation or haul‐out site).
Subpopulations are roughly listed from west to east beginning with the International Date Line.
The code for survey references is in Supporting Information.
Confidence limit (CL) was reported as the minimum population size.
Confidence interval was reported as 90%.
Figure 2.
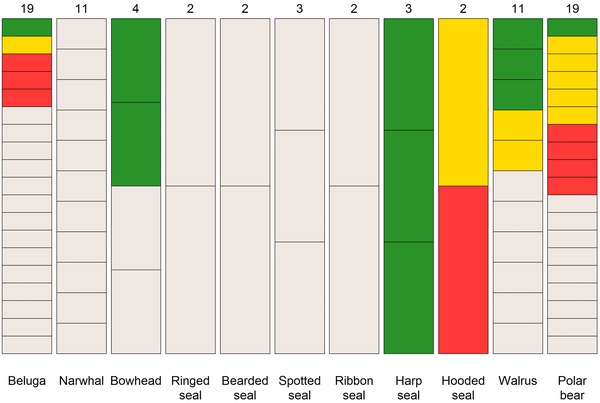
Trends in Arctic marine mammal abundance based on the most recent assessment for each recognized subpopulation of a species (red, declining trend in abundance; yellow, stable trend; green, increasing trend; gray, unknown trend). Number of subpopulations is given at the top of each bar. Each column is divided into equal segments, the sizes of which are not proportional to the size of the subpopulation. Ringed seal and bearded seal segments represent subspecies. Walrus segments represent subpopulations within subspecies. See Table 1 for details on abundance.
Evaluation of the abundance and trend of AMMs is complicated by unknown subpopulation structure or partial surveys of seasonal aggregations. High variability in survey methods, duration of surveys, and levels of survey precision made a summary of trend data difficult. Nonetheless, some assessment of trend was available for 35% of the 78 identified AMM subpopulations (Table 1 & Fig. 2).
Human Use of AMMs
We found that AMMs are taken for subsistence in all Arctic nations except Norway (including Svalbard). Based on our review of 78 subpopulations and subspecies, 78% (n = 61) were regularly and legally harvested for subsistence (Table 1). Of the cetaceans, 76% of subpopulations were harvested (74% of belugas, 91% of narwhals, 50% of bowhead whales), not including beluga stocks from which live whales were captured for aquaria (White Sea and Okhotsk Sea). Approximately 80% of pinniped subpopulations including walruses were harvested for subsistence, and 4 subpopulations were taken for commercial purposes by Norway, Canada, and Russia (harp and hooded seals [Table 1]). Of the 19 polar bear subpopulations, 80% were harvested for subsistence, not including the Kara Sea and Laptev Sea subpopulations where a small illegal harvest likely occurs.
Sea Ice Habitat Loss
Large changes in sea ice occurred in nearly all AMM habitats. Eleven of the 12 regions showed statistically significant trends for 1979–2013 toward earlier spring sea ice retreat, later fall sea ice advance, and, consequently, longer summers (Table 2 & Fig. 3, Supporting Information). Only the Bering Sea showed no trend. In 10 regions, the cumulative effect of the 34‐year trend was that the summer season was 5–10 weeks longer in 2013 than in 1979. Trends were largest in the Barents Sea, where summer was 20 weeks longer over this period.
Table 2.
Trends in the spring and fall sea ice transition dates and trends in the time interval between them (1 SD) for 1979–2013
| Region number | Region name | Spring (days/decade) | Fall (days/decade) | Interval (days/decade) | Correlation of spring and fall residualsa |
|---|---|---|---|---|---|
| 1 | East Siberian Sea | −6.8 (2.7)b | +8.6 (1.5)b | +15.4 (3.9)b | −0.64b |
| 2 | Sea of Okhotsk | −4.6 (1.6)b | +5.1 (2.2)b | +9.7 (3.1)b | −0.24 |
| 3 | Bering Sea | +1.8 (1.7) | +0.9 (2.2) | −0.9 (3.0) | −0.16 |
| 4 | Chukchi Sea | −5.9 (1.5)b | +7.0 (2.2)b | +12.9 (3.3)b | −0.59b |
| 5 | Beaufort Sea | −7.3 (3.3)b | +7.8 (1.9)b | +15.2 (4.5)b | −0.48b |
| 6 | Canadian Archipelago | −7.3 (2.2)b | +6.3 (1.2)b | +13.7 (3.1)b | −0.61b |
| 7 | Hudson Bay | −5.0 (1.0)b | +4.8 (1.2)b | +9.8 (1.9)b | −0.45b |
| 8 | Baffin Bay | −7.0 (1.2)b | +5.2 (1.4)b | +12.2 (2.3)b | −0.65b |
| 9 | Labrador Sea | −9.7 (3.1)b | +10.7 (2.5)b | +20.4 (4.5)b | −0.27 |
| 10 | Greenland Sea | −6.1 (1.7)b | +6.2 (2.7)b | +12.3 (3.7)b | −0.37 |
| 11 | Barents Sea | −17.2 (2.8)b | +25.1 (5.4)b | +41.8 (7.1)b | −0.39b |
| 12 | Laptev and Kara Seas | −9.4 (1.6)b | +7.0 (1.5)b | +16.4 (2.8)b | −0.66b |
Correlation of spring and fall transition dates after the trends are removed (i.e., correlation of residuals from trend line).
Statistical significance: P < 0.05 in a 2‐sided F test.
Figure 3.
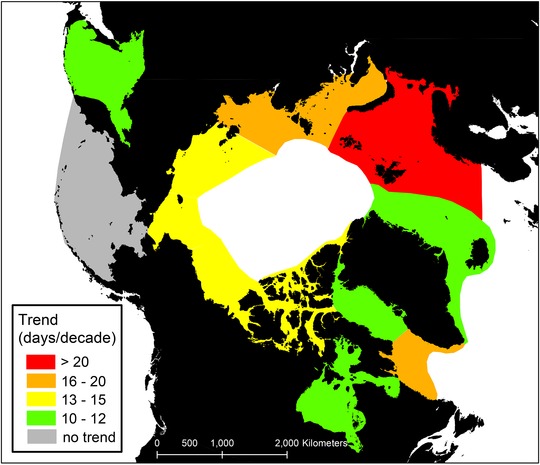
Trends in the length of the summer season (time from spring sea ice retreat to fall sea ice advance) in days per decade for 1979–2013. All trends in color are significant at P < 0.01. See Table 2 for specific values.
The dates of sea ice retreat and advance were not sensitive to the choice of the threshold by which they were defined (Supporting Information) because sea ice area typically changes rapidly in spring and fall. In all regions, the date of fall sea ice advance was negatively correlated with the date of spring sea ice retreat (Table 2).
Discussion
Climate change has had widespread ecological impacts on the Arctic (Rosenzweig et al. 2008; Gilg et al. 2012; Post et al. 2013), yet its effects are relatively underreported despite abiotic changes that exceed those in temperate, tropical, and montane biomes (ACIA 2005). Recent studies provide quantitative evidence of negative impacts of sea ice loss on some ice‐obligate AMMs (Stirling et al. 1999; Regehr et al. 2007; Øigard et al. 2010, 2013; Regehr et al. 2010). However, species‐ and subpopulation‐specific responses to climate change are likely to vary in time and space, as evidenced by delayed or even positive responses to sea ice loss for some AMMs (Moore & Laidre 2006; Quakenbush et al. 2011; Stirling et al. 2011; Rode et al. 2014; George et al. 2015). Variability in AMM responses can arise from differences in exploitation histories, life history strategies, biological productivity, and trophic interactions. Such heterogeneity with respect to climate change is widely recognized in terrestrial systems, which tend to be more data rich (Moritz & Agudo 2013).
Status and Trends of AMMs
Assessing status and trends of marine mammal populations in the Arctic is difficult because wide distributions and cryptic behavior are compounded by the logistical challenges of surveying in remote marine areas. Trend data are particularly important for understanding population status and identifying conservation priorities, but estimates of abundance and trend for most AMMs are absent or poor (Table 1 & Fig. 3), and lack of baseline data will limit the utility of future assessments. Although habitat loss is expected to lower the environmental carrying capacity for many ice‐dependent species, currently several AMMs are recovering from commercial exploitation (e.g., bowhead whales [George et al. 2015], polar bear sport hunting [Schliebe et al. 2006]) or previously insufficiently managed subsistence hunting (e.g., prior to 2000 West Greenland walrus [Witting & Born 2013]). In the short term, recovery from previous overexploitation could mask reductions in carrying capacity associated with habitat loss. In addition, increased biological productivity could, for an unknown period, offset potential negative impacts of sea ice loss and result in transient or longer term population increases (Quakenbush et al. 2011; Rode et al. 2014; George et al. 2015). Some populations (e.g., St Lawrence Estuary beluga and Cook Inlet beluga) show no evidence of recovery despite cessation of human harvesting (Wade et al. 2012).
Conservation and Management of AMM Human Use
In the modern world, it is rare for large wild mammals, in particular top predators, to support the nutritional and cultural well‐being of human communities, as AMMs do. Of the subpopulations and subspecies of AMMs we reviewed 78% are legally harvested for subsistence (Table 1). In addition, 4 subpopulations of pinnipeds are hunted commercially. Thus, management and conservation of AMMs is intertwined with their use as a renewable resource.
The primary responsibility for management of AMMs resides with federal and state government agencies and partners representing indigenous communities. Many AMMs are managed under regional, national, or international comanagement agreements that share decision‐making power between groups. Such agreements provide the framework for subsistence harvests and are supported by national laws (e.g., the U.S. Marine Mammal Protection Act, the Nunavut Land Claims Agreement). Although not a comprehensive list, examples of effective comanagement bodies including Alaska Native groups working with federal and state partners in the United States include the Alaska Beluga Whaling Committee, Alaska Eskimo Whaling Commission, Alaska Eskimo Walrus Commission, Alaska Nanuuq Commission, and the Ice Seal Committee. In Canada, such groups include Inuvialuit Fisheries Joint Management Committee; Wildlife Management Boards in Nunavut, Nunavik, and Nunatsiavut; and the Polar Bear Administrative Committee. In Greenland and Chukotka (Russia), the Greenland Associations of Hunters and Fisheries (KNAPK) and the Association of Traditional Marine Mammal Hunters of Chukotka, respectively, cooperate with federal and state agencies. Furthermore, in Greenland, all management of AMMs is carried out by a local government composed mainly of ethnic Inuit.
AMMs are highly mobile and undertake large seasonal movements, resulting in nearly half of subpopulations or stocks (Table 1) ranging across regional or international political boundaries, although this is difficult to quantify because movement data are poor. Management of transboundary subpopulations requires international collaboration. Currently, several subpopulations of polar bears are managed under joint international commissions (e.g., between Canada and Greenland; the United States and Russia) and information on polar bears is shared between the United States and Canada through the Inupiat‐Inuvialuit Agreement and the Canadian Polar Bear Technical Committee (also including Greenland). Scientific information on narwhals is shared though the Canada–Greenland Joint Commission for Narwhal‐Beluga and NAMMCO. Norway and Greenland receive management advice through NAMMCO, whereas catch limits for bowhead whales in Russia, the United States, and Greenland are set by the IWC. Advice on harp and hooded seals is coordinated by the International Council for the Exploration of the Seas and the North Atlantic Fisheries Organization.
Circumpolar Trends and Variability in Sea Ice Habitat Loss
It is well established that the area of Arctic sea ice has been declining in every month of the year (Stroeve et al. 2012). However, monthly areal trends do not capture the timing of annual sea ice retreat and advance, which influence reproduction, feeding, and life history events for AMMs. We therefore quantified sea ice habitat loss based on the timing of the seasonal change between winter and summer sea ice conditions. This is likely a biologically meaningful approach for most AMMs although other metrics may be appropriate for specific species or subpopulations. We found significant trends in the dates of spring sea ice retreat and fall sea ice advance for 1979‐2013, ranging from 5 to 17 days/decade earlier spring retreat and from 5 to 25 days/decade later fall advance, in 11 of the 12 regions (Table 2). This is consistent with the findings of Stammerjohn et al. (2012), who used methods similar to ours, and Stroeve et al. (2014), who used the detection of liquid water on the surface of the ice or snow to obtain statistically significant trends toward earlier melt onset and later freezing in most regions. In addition to declining sea ice extent, the thickness of sea ice has decreased substantially (Schweiger et al. 2011). Continuation of this trend is expected to induce greater variability in summer sea ice extent (Notz 2009) and possibly in the dates of spring retreat and fall advance because weather anomalies (e.g., a warm spring or a strong storm) have a greater impact on thin ice. The correlation we found between spring and fall transition dates (Table 2) is a manifestation of the ice‐albedo feedback, in which extra heat absorbed by the ocean during an early spring must be released into the atmosphere in the fall before sea ice can begin to form.
The direct and indirect impacts of sea ice loss on AMMs have been comprehensively reviewed by Laidre et al. (2008 a) and Kovacs et al. (2011). Loss of sea ice has affected survival in some subpopulations of polar bears (Regehr et al. 2007; Regehr et al. 2010). Pinniped pup survival is related to the timing of sea ice breakup because young need sufficient time for suckling prior to weaning (Øigard et al. 2010; 2013). Absence of sea ice is expected to lower Pacific walrus calf survival due to crushing at crowded haul‐out sites (Jay et al. 2012). Physical properties, such as ice roughness and snow depth (which has been decreasing in the Arctic [Webster et al. 2014]), have direct impacts on the suitability of sea ice for ringed seal lairs (Furgal et al. 1996; Iacozza & Ferguson 2014). Timing of sea ice breakup is linked to accessibility of seasonal foraging habitats and the spring primary production bloom that ultimately influences feeding for all AMMs (Carmack & Wassmann 2006). Indirect consequences of sea ice loss for AMM may include increased seasonal overlap in habitat with new species, including predators and competitors. Finally, loss of sea ice habitat will impact both AMM subpopulations and the ability of humans to access them for subsistence because a large fraction of subsistence hunting takes place on the sea ice or near the ice edge (e.g., Born et al. 2011).
Conservation Recommendations for the 21st Century
The environmental changes affecting the Arctic are not expected to abate in the immediate future. At present, there is no single international agreement to reduce global emissions of greenhouse gasses, the driver of climate warming and associated sea ice loss (IPCC 2013). Furthermore, forecasted sea ice loss for unabated emissions versus aggressive mitigation scenarios do not substantively diverge until at least 25 years into the future (Overland et al. 2014). Therefore, the trend in sea ice loss appears fixed for several decades regardless of global efforts to mitigate greenhouse‐gas emissions. Although reduction of emissions is the primary solution for mitigating long‐term future warming, scientists, managers, conservationists, industry, and local communities dependent on AMMs must prepare to deal with unprecedented environmental change. Accordingly, we make the following recommendations for AMM conservation.
Effective Comanagement
Maintaining and increasing comanagement by local and governmental entities is a key component of AMM conservation in the face of climate‐induced changes in population viability (see “Human Use” section). AMM populations provide resources that are important to the culture and well‐being of local and indigenous people throughout the Arctic (Born et al. 2011). Comanagement agreements directly involve people who use a resource and have a vested interest in it. They can lead to increased community participation in conservation efforts (e.g., minimizing human–polar bear conflicts), collection of traditional ecological knowledge, compliance with harvest monitoring and local harvest restrictions, identification of science priorities important to local communities, and opportunities for scientific sampling. In the Arctic, many of these activities and benefits are prohibitively expensive or otherwise infeasible without local involvement.
Future management of some AMMs will require balancing subsistence needs with declines in environmental carrying capacity due to sea ice loss. There will be a need to scientifically quantify the incremental effects of harvest on population viability and to balance such effects with the social and conservation benefits of continued use. If carrying capacity is declining due to habitat loss, responsible harvest management (e.g., harvest at an appropriate percentage of current abundance [Runge et al. 2009; Regehr et al. 2015]) will hold populations below carrying capacity and is unlikely to accelerate environmentally driven declines. Thus, the effects on population viability may be small if the estimates of harvest percentage and current abundance are either updated periodically based on scientific monitoring or are set at conservative levels. A precautionary approach may be warranted for some small or declining AMM populations, whereby increasingly risk‐averse management is applied if populations decline in size or resilience.
Given the fast pace of ecological change in the Arctic and uncertainty in how populations will respond, flexible and adaptive management will be critical. This requires clear articulation of conservation goals and population targets. It also requires an understanding with local resource users and managers that reducing direct human‐caused disturbance and removals, including subsistence harvests, may be one of the only available mechanisms to influence (but not necessarily offset) declines in AMM populations due to sea ice loss.
Variability in AMM Responses to Climate Change
Species and subpopulations exhibit variable responses to climate change over time and space (Post et al. 2009, 2013; Moritz & Agudo 2013). This can be due to variation in environmental characteristics (e.g., sea ice), species’ ability to move to favorable habitats, phenotypic and behavioral plasticity, or genetic traits that bolster resilience. Such variability is important to managing the potential effects of climate change at regional scales (e.g., Post et al. 2009; Gilg et al. 2012) and should be incorporated into predictive models and management plans.
There are several examples of contrasting population responses to changes in climate. Chukchi Sea and southern Beaufort Sea polar bears live in regions with similar rates of sea ice loss (Table 2), but Chukchi bears have body condition and reproductive parameters that are similar to historic values in the same region, whereas values for southern Beaufort bears have declined (Rode et al. 2014). Samples from subsistence‐harvested ringed seals in the northern Bering and Chukchi Seas in the 2000s indicate that several vital rates are similar to or better than during the 1960s and 1970s (Quakenbush et al. 2011), whereas in the Beaufort Sea, ringed seal body condition and productivity declined from 1992 to 2011 (Harwood et al. 2012). For both polar bears and ringed seals, the observed variation has been attributed at least in part to regional differences in biological productivity.
Some bowhead whale subpopulations have also shown positive population growth concurrent with regional sea ice loss. Both the Bering‐Chukchi‐Beaufort (BCB) and the Canada–Greenland subpopulations were previously reduced to low levels by commercial whaling and have increased at >3%/year, close to their theoretical maximum (Wiig et al. 2011; Givens et al. 2013). At the same time, body condition of harvested BCB bowhead whales improved from 1989 to 2011 during a period of reduction in sea ice (George et al. 2015), likely due to more extensive and productive feeding opportunities (Moore & Laidre 2006).
Though some species are showing positive responses to recent environmental changes, sea ice forecasts for the next 50–100 years (Wang & Overland 2012) indicate serious threats to many AMMs. Models that forecast population declines in several decades or a century (e.g., Amstrup et al. 2010; Udevitz et al. 2013) may inform the long‐term conservation status of AMMs but do not necessarily prescribe effective management in the near term. Part of the challenge is that climate models use broad spatial resolutions (Overland & Wang 2013) and exhibit uncertainty in sea ice trends over long time frames. Pitfalls associated with making local decisions based on coarse‐resolution forecasts include missed opportunities, both for conservation and sustainable use, the latter of which may risk alienating stakeholders and compromising future conservation efforts. We recommend that, to the extent possible, managers consider relative risks and benefits based on scientific information at both coarse and fine spatial and temporal scales.
Improved AMM Monitoring
Some populations of AMMs are monitored for harvest management purposes or to determine the effects of anthropogenic activities on a regional scale. Measuring trend, which is an important indicator of population status, requires abundance data over many years or a demographic analysis of vital rates (e.g., reproduction and survival), which are available for relatively few populations (Table 1). Given the elusive behavior and wide distribution of AMMs, acquiring data on trends in abundance for all AMMs is not realistic; thus, it is important to develop other information sources. The availability of subsistence harvest samples provides an opportunity, in collaboration with local communities, to obtain a suite of metrics (e.g., age at maturity, pregnancy rate, growth rate, body condition, pollution, and contaminant loads) that could serve as broader ecological indicators. Other metrics that are relatively feasible to monitor include habitat quality through remote sensing or ocean observation (Moore & Gulland 2014). Such data, analyzed together with regular surveys of subpopulation abundance in key areas, may be a reasonable monitoring strategy.
Long‐term monitoring of specific subpopulations has provided a foundation of knowledge for several species, including polar bears (e.g., Western Hudson Bay and Southern Beaufort Sea [Stirling et al. 1999; Regehr et al. 2010]) and ringed seals (Harwood et al. 2012). Scientific monitoring programs working in conjunction with local communities, including subsistence hunters, have the potential to provide large amounts of data at relatively low cost. Successful examples in Alaska include the Department of Fish and Game's Ice Seal Bio‐monitoring Program (Quakenbush et al. 2011) and the U.S. North Slope Borough's bowhead whale harvest sampling program (George et al. 2015). Harvest biosampling for polar bears is also successfully conducted in Greenland and Canada.
Circumpolar scientific monitoring plans have been drafted for beluga whales, ringed seals, and polar bears (e.g., Kovacs 2008; Laidre et al. 2008 b; Simpkins et al. 2009; Vongraven et al. 2012; Kovacs et al. 2014), but they have not been systematically implemented. This may be due to a number of factors including insufficient funding, a lack of political organization or will, lack of awareness of the plans, or the absence of major economic incentives for international cooperation on biodiversity monitoring and conservation. Circumpolar monitoring plans often recommend broad projects that span agency interests and authority. This may be scientifically rigorous, but it can make it difficult for any single agency to support all the work outlined in a plan while concurrently performing the focused studies necessary to meet regional needs.
With sea ice loss, a physical barrier that previously limited interchange between some subpopulations is disappearing (Heide‐Jørgensen et al. 2011), which complicates how changes are monitored and emphasizes the importance of circumpolar analyses. On the international level, the Arctic Council has been moving toward circumpolar monitoring for AMMs through the Arctic Monitoring and Assessment Program and the CAFF Circumpolar Biodiversity Monitoring Program. The polar bear range states are preparing a circumpolar action plan under the auspices of the 1973 International Agreement for the Conservation of Polar Bears.
An example of successful circumpolar monitoring in the Southern Ocean is provided by the Commission for the Conservation of Antarctic Marine Living Resources, a multinational program supported by the International Antarctic Treaty (Miller & Slicer 2014). In this case, national governments support monitoring because of a mutual need to understand the effects of commercial harvest of a broad‐scale shared resource, the multinational krill fishery. Emerging fisheries, shipping, expanding oil and gas development, and other types of development in the Arctic may provide future economic incentives for AMM circumpolar monitoring.
Overall, future monitoring of AMMs will benefit from careful consideration of objectives. These should include scientific monitoring to understand how ecological mechanisms such as trophic interactions cause change and monitoring for applied conservation and management (e.g., noise, oil risk, pollution, subsistence harvest), prioritization of which AMMs to monitor, a priori scientific study design to guide sampling efforts, and tasking international forums to standardize data collection, facilitate scientific analyses, and evaluate patterns across population and political boundaries.
Impacts of Industrial Activities
It is important to understand and mitigate cumulative impacts from industrial activities. The longer open‐water season has contributed to industrial interest and development in the Arctic and to increased use of shorter international shipping routes (Reeves et al. 2012). More than half the circumpolar range of Arctic cetaceans overlaps known or suspected offshore oil and gas deposits (Reeves et al. 2014). Potential threats associated with oil and gas development include human‐generated underwater sound, ship strikes, displacement from critical habitat, and the risk of accidental or illegal discharge of oil (AMSA 2009). Arctic countries currently have little or no capacity to contain an oil spill in the sea ice and little is known about the impacts of dispersants on Arctic biota or AMMs. Similarly, little is known about the cumulative effects of multiple stressors on AMMs, and monitoring is difficult due to low levels of formal circumpolar oversight for industry. The Arctic Council's Emergency Prevention, Preparedness and Response Working Group, as well as the Protection of the Arctic Marine Environment Working Group, are currently developing international guidelines and agreements. However, the regulations in place and capacity to mitigate threats vary among regions, and linking planning efforts to actual infrastructure development and capacity remains an emerging need.
In the United States, local groups including the Arctic Marine Mammal Coalition are working to facilitate communication between coastal communities and the U.S. Coast Guard, National Oceanic and Atmospheric Administration (NOAA), and other regulatory agencies on the potential impacts of increased ship traffic. In Alaska annually negotiated conflict avoidance agreements between the Alaska Eskimo Whaling Commission and offshore oil and gas operators mitigate potential conflicts between subsistence hunters and industrial development.
In the Arctic, the protection of species and habitats is intertwined with rural development, resource extraction, and climate change. Understanding and mitigating anthropogenic impacts is of essential importance for setting future conservation priorities. Addressing those priorities will require effective partnerships and collaborations among local people, industry, nongovernmental organizations, and government agencies.
Protected Species Legislation
It is important to recognize both the utility and limitations of protected species legislation in a changing Arctic. Existing frameworks for international and national protected species legislation have only recently begun to include climate change. For example, climate change was not incorporated as an explicit threat into the 2008 IUCN mammalian Red List process (Schipper et al. 2008), although it is expected to be considered in the future. After delays associated with the creation of memorandums of understanding between the federal government and Aboriginal land claim comanagement boards, Canada is moving forward with listing some AMMs under the Species at Risk Act. In the United States, the polar bear was listed as threatened in 2008 (USFWS 2013) under the Endangered Species Act (ESA) and some populations of bearded seals and ringed seals were listed as threatened in 2012 (National Marine Fisheries Service 2012).
Until recently ESA listings were largely made on the basis of immediate or shorter term anthropogenic threats that could be mitigated by regulatory or other action (Ruhl 2009). In contrast, reducing emissions of greenhouse gases is a long‐term and global challenge that cannot be addressed by regional actions. Furthermore, the causal connections between sea ice loss and impacts on the habitat or status of AMMs are not uniform. The ESA listings have been made on the basis of long‐term predictive models (Schliebe et al. 2006; Cameron et al. 2010; Kelly et al. 2010) rather than on observed population declines. The contrast between current AMM abundances and predicted declines can make setting near‐term conservation priorities difficult. For example, an ESA threat analysis predicted that ringed seal populations will decline during the 21st century from the present‐day level of several million animals to a level that would place them in danger of extinction (Kelly et al. 2010; National Marine Fisheries Service 2012).
Agencies tasked with recovery planning under the ESA (U.S. Fish and Wildlife Service and NOAA) do not have the authority to regulate greenhouse gases (e.g., USFWS 2013), the primary threat that justified the listings. It is not yet clear whether species legislation will influence national or global level decision making about greenhouse‐gas emissions. In some cases, listings may provide the basis to mitigate secondary factors (e.g., industrial development or subsistence use) and afford species an increased chance of persistence until global action results in a stabilized climate (Ruhl 2009; Seney et al. 2013). Agencies will need to formulate a balanced approach with regard to secondary factors, which can affect persistence of AMMs but may also provide benefits such as economic growth and maintenance of nutritional and cultural traditions (Regehr et al. 2015).
International agreements may be needed to protect key AMM habitats in some regions including the Northwest and Northeast Passages and Bering Strait (key areas of interest for shipping). Protection of Ellesmere Island and North Greenland, areas projected to become AMMs’ last ice refuge, will require cooperation between Greenland and Canada, and possibly other nations (ACIA 2005). The Arctic Basin, an area now largely free of human activity, may also become an important habitat for some AMMs.
Forward‐Looking Conservation
AMMs range across international borders, serve as biological indicators, occupy areas rich in natural resources, are important for traditional subsistence and ecosystem health, and have become icons of the consequences of climate change. As concerns about the effects of climate change on AMMs increase, there will be a need for those involved in AMM conservation and management to mitigate new resource and management conflicts (e.g., Derocher et al. 2013) by incorporating scientific evidence on species status with value‐based conservation. This will include communicating accurate information to the public (Brulle 2014) on topics such as regional variability in AMM responses to reduced ice, lack of baselines, scientific uncertainty in data, and a future Arctic ecosystem that may contain a different marine mammal species assemblage.
Success in AMM conservation over the near‐ and mid‐term will require engagement of regional governments and indigenous organizations that influence the direct interface between humans and AMMs. Success over the longer term will require the influence of federal governments that can implement new policies and address the global causes of climate change, particularly greenhouse‐gas emissions. Nongovernmental organizations committed to cooperative conservation can play a role in all the steps of the process. We suggest that common ground can be found if all stakeholders recognize AMMs as iconic species with inherent value and as resources connected to the well‐being of humans who harvest, interact, and live with them. Accurate scientific data—currently lacking for many species—will be key to making informed and effective decisions about the conservation challenges and tradeoffs facing AMMs in the 21st century.
Supporting information
Disclaimer: Supplementary materials have been peer‐reviewed but not copyedited.
The methods used to analyze sea ice retreat and advance in 12 regions (Appendix S1) and the citations to surveys from which abundance estimates and trends (Table 1) were obtained (Appendix S2) are available online. A Russian abstract is available in Appendix S3. The authors are solely responsible for the content and functionality of these materials. Queries (other than absence of material) should be directed to the corresponding author.
Acknowledgments
Thanks to CAFF Arctic Biodiversity Assessment mammal chapter coleaders, D. Reid and D. Berteaux, and authors J. Garlich‐Miller, E. Peacock, and M. Simpkins who contributed to that work. K.L.L. was supported by the Danish Ministry of the Environment, the Greenland Institute of Natural Resources, and, together with H.S., supported by NASA grants NNX13AN28G and NNX11AO63G. J. Estes, M. Robards, and 2 anonymous reviewers provided useful input. The findings and conclusions in this article are those of the authors and do not necessarily represent the views of the U.S. Fish and Wildlife Service or NOAA.
Literature Cited
- AMSA (Arctic Marine Shipping Assessment) . 2009. Arctic Marine Shipping Assessment 2009 Report. Arctic Council, April 2009. [Google Scholar]
- AMAP (Arctic Monitoring and Assessment Programme) . 2009. Arctic pollution 2009. Arctic Monitoring and Assessment Programme, Oslo. [Google Scholar]
- Amstrup SC, DeWeaver ET, Douglas DC, Marcot BG, Durner GM, Bitz CM, Bailey DA. 2010. Greenhouse gas mitigation can reduce sea‐ice loss and increase polar bear persistence. Nature 468:955–958. [DOI] [PubMed] [Google Scholar]
- ACIA (Arctic Council, Arctic Climate Impact Assessment, Arctic Monitoring and Assessment Programme, CAFF & International Arctic Science Committee) . 2005. Arctic climate impact assessment. Cambridge University Press, Cambridge, United Kingdom. [Google Scholar]
- Born EW, Heilmann A, Holm LK, Laidre KL. 2011. Polar bears in northwest Greenland: an interview survey about the catch and the climate. Meddelelser om Grønland 351:1–250. [Google Scholar]
- Brulle RJ. 2014. Institutionalizing delay: foundation funding and the creation of U.S. climate change counter‐movement organizations. Climatic Change 122:681–694. [Google Scholar]
- CAFF (Conservation of Arctic Flora and Fauna) . 2011. Arctic marine biodiversity monitoring plan (CBMP‐MARINE PLAN) Monitoring series report 3. CAFF International Secretariat, Akureyri, Iceland. [Google Scholar]
- Cameron MF, et al. 2010. Status review of the bearded seal (Erignathus barbatus). Technical memorandum NMFS‐AFSC‐211. U.S. Department of Commerce, National Oceanic and Atmospheric Administration, Washington, DC. [Google Scholar]
- Carmack E, Wassmann P. 2006. Food webs and physical–biological coupling on pan‐Arctic shelves: unifying concepts and comprehensive perspectives. Progress in Oceanography 71:446–477. [Google Scholar]
- Cavalieri D, Parkinson C, Gloersen P, Zwally HJ. 1996. Sea ice concentrations from Nimbus‐7 SMMR and DMSP SSM/I‐SSMIS passive microwave data, 1979‐2010. National Snow and Ice Data Center, Boulder, Colorado (digital media, updated yearly).
- Derocher AE, et al. 2013. Rapid ecosystem change and polar bear conservation. Conservation Letters 6:368–375. [Google Scholar]
- Furgal CM, Kovacs KM, Innes S. 1996. Characteristics of ringed seal (Phoca hispida) subnivian lairs and habitat and their effects on predation. Canadian Journal of Zoology 74:858–874. [Google Scholar]
- George JC, Druckenmiller M, Laidre KL, Suydam R, Person B. 2015. Western Arctic bowhead whale body condition and links to summer sea ice and upwelling in the Beaufort Sea Progress in Oceanography: in press. [Google Scholar]
- Gilg O, et al. 2012. Climate change and the ecology and evolution of Arctic vertebrates. Annals NY Academy of Science 1249:166–190. [DOI] [PubMed] [Google Scholar]
- Givens GH, Edmondson SL, George JC, Suydam R, Charif RA, Rahaman A, Hawthorne D, Tudor B, DeLong RA, Clark CW. 2013. Estimate of 2011 Abundance of the Bering‐Chukchi‐Beaufort Seas bowhead whale population. Paper SC/65a/BRG01. International Whaling Commission, Cambridge, United Kingdom. [Google Scholar]
- Harwood LA, Smith TG, Melling H, Alikamik J, Kingsley MCS. 2012. Ringed seals and sea ice in Canada's western Arctic: harvest based monitoring 1992‐2011. Arctic 65:377–390. [Google Scholar]
- Heide‐Jørgensen MP, Laidre KL, Quakenbush LT, Citta J. 2011. Northwest Passage opens for bowhead whales. Biology Letters 8:270–273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iacozza J, Ferguson SH. 2014. Spatio‐temporal variability of snow cover over sea ice in western Hudson Bay, with reference to ringed seal pup survival. Polar Biology 37:817–832. [Google Scholar]
- IPCC (Intergovernmental Panel on Climate Change) . 2013. Climate change 2013: the physical science basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, United Kingdom. [Google Scholar]
- Jay CV, Fischbach AS, Kochnev AA. 2012. Walrus areas of use in the Chukchi Sea during sparse sea ice cover. Marine Ecology Progress Series 468:1–13. [Google Scholar]
- Kelly BP, et al. 2010. Status review of the ringed seal (Phoca hispida). U.S. Department of Commerce, National Oceanic and Atmospheric Administration, Technical Memorandum NMFS‐AFSC‐212. 250 p.
- Kovacs KM. 2008. Background document for development of a circumpolar ringed seal (Phoca hispida) monitoring plan. Report to the U.S. Marine Mammal Commission, Bethesda. [Google Scholar]
- Kovacs KM, et al. 2012. Global threats to pinnipeds. Marine Mammal Science 28:414–436. [Google Scholar]
- Kovacs KM, et al. 2014. Circumpolar ringed seal (Pusa hispida) monitoring. Report series 143. Norwegian Polar Institute, Tromsø, Norway. [Google Scholar]
- Kovacs KM, Moore S, Overland JE, Lydersen C. 2011. Impacts of changing sea ice conditions on Arctic marine mammals. Marine Biodiversity 41:181–194. [Google Scholar]
- Laidre KL, Stirling I, Lowry L, Wiig, Heide‐Jørgensen MP, Ferguson S. 2008. a Quantifying the sensitivity of arctic marine mammals to climate‐induced habitat change. Ecological Applications 18:S97–S125. [DOI] [PubMed] [Google Scholar]
- Laidre KL, Simpkins M, Lowry L, Heide‐Jørgensen MP, Reeves R, Moore S. 2008. b Background document for development of a circumpolar beluga (Delphinapterus leucas) monitoring plan. Report to the U.S. Marine Mammal Commission, Bethesda. [Google Scholar]
- Miller D, Slicer NM. 2014. CCAMLR and Antarctic conservation, in Governance of Marine Fisheries and Biodiversity Conservation: Interaction and Coevolution. Garcia SM, Rice J, Charles A, editors. John Wiley & Sons, Ltd, Chichester, United Kingdom. [Google Scholar]
- Moore SE, Gulland FMD. 2014. Linking marine mammal health and ecology in the‘new normal’ Arctic. Ocean & Coastal Management; 122:55–57. [Google Scholar]
- Moore SE, Laidre KL. 2006. Trends in sea ice cover within habitats used by bowhead whales in the western Arctic. Ecological Applications 16:932–944. [DOI] [PubMed] [Google Scholar]
- Mortiz C, Agudo R. 2013. The future of species under climate change: Resilience or decline? Science 341:504–508. [DOI] [PubMed] [Google Scholar]
- National Marine Fisheries Service . 2012. Threatened status for the Arctic, Okhotsk, and Baltic subspecies of the ringed seal and endangered status for the Ladoga subspecies of the ringed seal. Federal Register 77:249 (December 28, 2012) 76706–76738. [Google Scholar]
- Notz D. 2009. The future of ice sheets and sea ice: between reversible retreat and unstoppable loss. Proceedings of the National Academy of Sciences 106:20590–20595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Overland JE Wang M. 2013. When will the summer Arctic be nearly sea ice free? Geophysical Research Letters 40:2097–2101. [Google Scholar]
- Overland JE, Wang M, Walsh JE, Stroeve JC. 2014. Future Arctic climate changes: adaptation and mitigation time scales. Earth's Future 2:68–74. [Google Scholar]
- Øigård TA, Haug T, Nilssen KT. 2010. Estimation of pup production of hooded seals and harp seals in the Greenland Sea in 2007: Reducing uncertainty using generalized additive models. Journal of Northwest Fisheries Science 42:103–123. [Google Scholar]
- Øigård TA, Lindstrom U, Haug T, Nilssen KT, Smout, S. 2013. Functional relationship between harp seal body condition and available prey in the Barents Sea. Marine Ecology Progress Series 484:287–301. [Google Scholar]
- Post E, et al. 2009. Ecological dynamics across the Arctic associated with recent climate change. Science 325:1355–1358. [DOI] [PubMed] [Google Scholar]
- Post E, Bhatt US, Bitz CM, Brodie JF, Fulton TL, Hebblewhite M, Kerby J, Kutz SJ, Stirling I, Walker DA. 2013. Ecological consequences of sea ice decline. Science 341:519–524. [DOI] [PubMed] [Google Scholar]
- Quakenbush L, Citta J, Crawford J. 2011. Biology of the ringed seal (Phoca hispida) in Alaska, 1960–2010. Final Report to the National Marine Fisheries Service. Available from Alaska Department of Fish and Game, Fairbanks. [Google Scholar]
- Reeves RR, et al. 2014. Distribution of endemic cetaceans in relation to hydrocarbon development and commercial shipping in a warming Arctic. Marine Policy 44:375–389. [Google Scholar]
- Reeves R, Rosa C, George JC, Sheffield G, Moore M. 2012. Implications of Arctic industrial growth and strategies to mitigate future vessel and fishing gear impacts on bowhead whales. Marine Policy 36:454–462. [Google Scholar]
- Regehr EV, Hunter CM, Caswell H, Amstrup SC, Stirling I. 2010. Survival and breeding of polar bears in the southern Beaufort Sea in relation to sea‐ice. Journal of Animal Ecology 79:117–127. [DOI] [PubMed] [Google Scholar]
- Regehr EV, Lunn NJ, Amstrup SC, Stirling I. 2007. Effects of earlier sea‐ice breakup on survival and population size of polar bears in western Hudson Bay. Journal of Wildlife Management 71:2673–2683. [Google Scholar]
- Regehr EV, Wilson RR, Rode KD, Runge MC. 2015. Resilience and risk: a demographic model to inform conservation planning for polar bears: U.S. Geological Survey Open–File Report 2015–1029, 56 p., http://dx.doi.org/10.3133/ofr20151029. [Google Scholar]
- Reid DG, Berteaux D, Laidre KL. 2013. Chapter 3: Mammals Arctic biodiversity assessment: status and trends in Arctic biodiversity. Pages 78‐141 in Meltofte H, editor. Conservation of Arctic Flora and Fauna, Akureyri. Arctic Biodiversity Assessment. [Google Scholar]
- Rode KD, Regehr EV, Douglas DC, Durner G, Derocher AE, Thiemann GW, Budge SM. 2014. Variation in the response of an Arctic top predator experiencing habitat loss: feeding and reproductive ecology of two polar bear populations. Global Change Biology 20:76–88. [DOI] [PubMed] [Google Scholar]
- Rosenzweig C, et al. 2008. Attributing physical and biological impacts to anthropogenic climate change. Nature 453:353–357. [DOI] [PubMed] [Google Scholar]
- Ruhl JB. 2009. Climate change and the Endangered Species Act: building bridges to the no‐analog future. Environmental Law and Policy Annual Review 39:10735–10745. [Google Scholar]
- Runge MC, Sauer JR, Avery ML, Blackwell BF, Koneff MD. 2009. Assessing allowable take of migratory birds. Journal of Wildlife Management 73:556–565. [Google Scholar]
- Schipper J, et al. 2008. The status of the world's land and marine mammals: diversity, threat, and knowledge. Science 322:225–230. [DOI] [PubMed] [Google Scholar]
- Schliebe S, Evans T, Johnson K, Roy M, Miller S, Hamilton C, Meehan R, Jahrsdoerfer S. 2006. Range‐wide status review of the polar bear (Ursus maritimus). U.S. Fish and Wildlife Service, Anchorage, Alaska. [Google Scholar]
- Schweiger A, Lindsay R, Zhang J, Steele M, Stern H, Kwok R. 2011. Uncertainty in modeled Arctic sea ice volume, Journal of Geophysical Research 116:C00D06. DOI: 10.1029/2011JC007084. [Google Scholar]
- Seney EE, Rowland MJ, Lowery RA, Griffis RB, McClure MM. 2013. Climate change, marine environments, and the U.S. Endangered Species Act. Conservation Biology 27:1138–1146. [DOI] [PubMed] [Google Scholar]
- Simpkins M, Kovacs KM, Laidre K, Lowry L. 2009. A framework for monitoring arctic marine mammals. Conservation of Arctic flora and fauna (CAFF) circumpolar biodiversity monitoring plan. Report 16. CAFF, Akureyri, Iceland. [Google Scholar]
- Stammerjohn S, Massom R, Rind D, Martinson D. 2012. Regions of rapid sea ice change: an inter‐hemispheric seasonal comparison. Geophysical Research Letters 39:L06501. DOI: 10.1029/2012GL050874. [Google Scholar]
- Stirling I, Lunn NJ, Iacozza J. 1999. Long‐term trends in the population ecology of polar bears in western Hudson Bay in relation to climatic change. Arctic 52:294–306. [Google Scholar]
- Stirling I, McDonald TL, Richardson ES, Regehr EV, Amstrup SC. 2011. Polar bear population status in the Northern Beaufort Sea, Canada, 1971–2006. Ecological Applications 21:859–876. [DOI] [PubMed] [Google Scholar]
- Stroeve JC, Markus T, Boisvert L, Miller J, Barrett A. 2014. Changes in Arctic melt season and implications for sea ice loss. Geophysical Research Letters 41 DOI:10.1002/2013GL058951. [Google Scholar]
- Stroeve JC, Serreze MC, Holland MM, Kay JE, Malanik J, Barrett AP. 2012. The Arctic's rapidly shrinking sea ice cover: a research synthesis. Climatic Change 110:1005–1027. [Google Scholar]
- Udevitz MS, Taylor RT, Garlich‐Miller JL, Quakenbush LT, Snyder J. 2013. Potential population‐level effects of increased haulout‐related mortality of Pacific walrus calves. Polar Biology 36:291–298. [Google Scholar]
- U.S. Fish and Wildlife Service (USFWS) . 2013. Endangered and threatened wildlife and plants; special rule for the polar bear under section 4(d) of the Endangered Species Act. Federal Register 78:34:11766–11788. [Google Scholar]
- Vongraven D, et al. 2012. A circumpolar monitoring framework for polar bears. Ursus 23(sp2):1–66. [Google Scholar]
- Wade PR, Reeves RR, Mesnick SL. 2012. Social and behavioural factors in cetacean responses to overexploitation: Are odontocetes less “resilient” than mysticetes? Journal of Marine Biology 15: (e 567276) DOI: 10.1155/2012/567276. [Google Scholar]
- Wang M, Overland JE. 2012. A sea ice free summer Arctic within 30 years‐an update from CMIP5 models. Geophysical Research Letters 39:DOI: 10.1029/2012GL052868. [Google Scholar]
- Webster MA, Rigor IG, Nghiem SV, Kurtz NT, Farrell SL, Perovich DK, Sturm M. 2014. Interdecadal changes in snow depth on Arctic sea ice. Journal of Geophysical Research, Oceans 119:5395–5406. [Google Scholar]
- Wiig Ø, Bachmann L, Heide‐Jørgensen MP, Lindquist C, Laidre KL, Postma L, Dueck L, Palsbøll P, Bachmann L. 2011. Recaptures of genotyped bowhead whales (Balaena mysticetus) in Eastern Canada and West Greenland. Endangered Species Research 14:235–242. [Google Scholar]
- Witting L, Born EW. 2013. Population dynamics of walrus in Greenland. Scientific publications 9. North Atlantic Marine Mammal Commission, Polar Environmental Centre N‐9296, Tromsø, Norway. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Disclaimer: Supplementary materials have been peer‐reviewed but not copyedited.
The methods used to analyze sea ice retreat and advance in 12 regions (Appendix S1) and the citations to surveys from which abundance estimates and trends (Table 1) were obtained (Appendix S2) are available online. A Russian abstract is available in Appendix S3. The authors are solely responsible for the content and functionality of these materials. Queries (other than absence of material) should be directed to the corresponding author.