

Abstract
The cytochrome P450 monooxygenases (P450s) represent a large and important enzyme superfamily in plants. They catalyze numerous monooxygenation/hydroxylation reactions in biochemical pathways, P450s are involved in a variety of metabolic pathways and participate in the homeostasis of phytohormones. The CYP82 family genes specifically reside in dicots and are usually induced by distinct environmental stresses. However, their functions are largely unknown, especially in soybean (Glycine max L.). Here, we report the function of GmCYP82A3, a gene from soybean CYP82 family. Its expression was induced by Phytophthora sojae infection, salinity and drought stresses, and treatment with methyl jasmonate (MeJA) or ethephon (ETH). Its expression levels were consistently high in resistant cultivars. Transgenic Nicotiana benthamiana plants overexpressing GmCYP82A3 exhibited strong resistance to Botrytis cinerea and Phytophthora parasitica, and enhanced tolerance to salinity and drought stresses. Furthermore, transgenic plants were less sensitive to jasmonic acid (JA), and the enhanced resistance was accompanied with increased expression of the JA/ET signaling pathway-related genes.
Introduction
Soybean (Glycine max L.) is an agronomic crop grown throughout the world. It is not only an important source of vegetable protein and oil for humans and animals, but it is also a source of biofuels. In addition, soybean seeds contain a number of high-value secondary compounds with nutraceutical properties such as isoflavones, saponins, and tocopherols [1–3]. Phytophthora root and stem rot caused by Phytophthora sojae is one of the most destructive soybean diseases, results in annual losses of $200 million in the USA and $1–2 billion globally [4]. The effective way to protect soybean against P. sojae infection is breeding with dominant Rps (Resistance to Phytophthora sojae) genes. But continuous utilization of a single Rps gene may result in selection pressure and promote the evolution of more pathogenic races of P. sojae. Thus, a particular Rps gene is effective for only 8 to 15 years. Partial resistance is another type of resistance which has been described as the relative ability of susceptible plants to survive infection without showing severe symptoms like death, stunting, or yield loss. It is conferred by multiple genes or quantitative trait loci (QTL), sometimes referred as quantitative, rate-reducing, or field resistance [5–7]. It is durable against all races of P. sojae and highly heritable [8, 9]. Therefore, partial resistance provides an effective way to develop Phytophthora resistant cultivars.
Till now, more than twenty QTL for partial resistance to P. sojae have been mapped in recombinant inbred line (RIL) populations. Most of the alleles of the QTL for partial resistance originating from Conrad, which is a proverbial cultivar with highly partial resistance to P. sojae [10–14]. At the same time, additional QTL were identified in V71-370 × PI 407162, Su88-M21 × Xinyixiaoheidou, S99-2281 × PI 408105A and OX20-8 × PI 398841 populations [15–19]. Identifying the key genes controlling these QTL and characterizing their functions will facilitate to understand the mechanisms that contribute to partial resistance. Generally, the genes that encode pathogenesis-related (PR) protein PR1a, PR2, basic peroxidase, and matrix metalloproteinase were present at higher abundances in partial resistant cultivars during infections [20]. The amount of preformed suberin was also found contribute to the partial resistance [21, 22]. Using soybean Aymetrix gene chips, whole-genome transcription profiles were analyzed in soybean genotypes with differential levels of partial resistance [8, 13, 23]. Appraising the differential transcript genes in Conrad and Sloan underlying the QTL found that most of them encompassed putative physiological trait genes, defense-related genes, and disease resistance-like genes [13, 14]. But the evidence of these candidate genes conferring resistance to P. sojae are still absent.
The cytochrome P450 (CYP) monooxygenases represent a large and important superfamily in plants. The enzymes catalyze a wide variety of monooxygenation/hydroxylation reactions in biochemical pathways involved in primary and secondary metabolism pathways [24, 25]. P450 genes also participate in the homeostasis of phytohormones [25]. In soybean, the cytochrome P450 gene family is comprised of 322 genes and 378 pseudogenes, but the biological functions most of them have not been elucidated [26]. The CYP82 family, which belongs to the CYP71 clan, is only present in dicots [27]. Some CYP82 members are reported to be highly induced by environmental stress in tobacco (Nicotiana tabacum L.), pea (Pisum sativum L.), soybean, and Arabidopsis [28–31]. A few CYP82 family members are involved in a variety of metabolic pathways. CYP82E4v1, from N. tabacum, was identified as a nicotine N-demethylase and the synthesis of nornicotine was suppressed in plants when the gene was silenced [32]. CYP82N2v2, from Eschscholzia californica, is involved in sanguinarine biosynthesis by catalyzing the conversion of protopine to dihydrosanguinarine by the P450 reaction [33]. CYP82D is reported to be involved in lipophilic flavone biosynthesis in sweet basil (Ocimum bacilicum L.) [34]. In Arabidopsis, CYP82G1 catalyzes the final step in the synthesis of the common plant homoterpene volatiles TMTT/DMNT, CYP82C2 and CYP82C4 hydroxylate the therapeutic compound 8-methoxypsoralen [35, 36]. The CYP82 family genes also participate in the interaction between plants and pathogens. For example, cotton (Gossypium hirsutum L.) CYP82D can regulate systemic cell death by modulating the octadecanoid pathway and negatively regulate disease resistance to Verticillium dahliae by controlling JA biosynthesis [37]. AtCYP82C2 can increase expressions of the JA-induced defense-related genes and contents of JA-induced IGs, and enhance resistance to Botrytis cinerea [38].
GmCYP82A3 was highly stress responsive and located in the P. sojae resistant QTL region [13, 28]. But the accurate function remains unknown. In this study, we characterized the functions of GmCYP82A3 in response to biotic and abiotic stresses. Ectopic expression of GmCYP82A3 in N. benthamiana enhanced resistance to the pathogen Botrytis cinerea and Phytophthora parasitica, tolerance to the abiotic salinity and drought stresses. Furthermore, we demonstrated that JA/ET signaling pathway was altered in the GmCYP82A3 overexpression plants. These findings provide valuable information on soybean partial resistance mechanisms.
Materials and Methods
Plant materials and growth conditions
The soybean cultivars Sloan (highly susceptible to P. sojae), Williams (moderately partial resistance to P. sojae) and Conrad (highly partial resistance) [8] were grown in a greenhouse. The greenhouse was maintained at 25°C and the photoperiod was set to 8D:16L. The N. benthamiana plants used were cultivated in the greenhouse under the same conditions.
Gene induction assays
The germplasm Conrad was used for gene expression assay. The detached leaves from 3-week-old seedlings were used for P. sojae infection. Hyphae disks were cut from the edges of newly cultured P. sojae P6497 isolate on 10% V8 medium [39]. The inoculated leaves were placed in Petri dishes, which contained a layer of filter paper to retain moisture. Then the Petri dishes were placed in a climate chamber and samples were taken from the inoculated site at 0, 3, 6, 12, and 24 hours post-infection (hpi).
For salinity and drought stress treatments, three-week-old seedlings were uprooted and washed to remove vermiculite. The plants were cultured in Hoagland liquid medium [40] for 2 days, then transferred into the same medium containing 200 mM NaCl or 20% PEG6000. The roots were sampled at 0, 6, 12, and 24 hours post-treatment (hpt).
For the different phytohormone treatments, three-week-old seedlings were uprooted and cultured in Hoagland liquid medium for 2 days, and then transferred into the same medium containing 100 μM methyl jasmonate (MeJA), 100 μM ethephon (ETH), 2 mM salicylic acid (SA) and 100 μM abscisic acid (ABA). The roots were sampled at 0, 6, 12, and 24 hpt. All samples were rapidly frozen in liquid nitrogen and stored at -70°C.
Plasmid construction and N. benthamiana stable transformation
To overexpress GmCYP82A3, the coding DNA sequence (CDS) of GmCYP82A3 (GenBank: NM_001254043.1) was amplified using the primers CYP82A3-F and CYP82A3-R from Conrad cDNA (S1 Table). The 1584 bp gene fragment was first cloned into the Gateway entry vector pDONR221, then cloned into pEarlyGate202 through an LR recombination reaction between the entry clone and the destination vector (Invitrogen, USA) [41]. The constructed vector was validated by sequencing, subsequently transformed into Agrobacterium tumefaciens (strain EHA105) by electroporation for further transformation.
The transgenic N. benthamiana plants were generated by A. tumefaciens mediated transformation from leaf discs, as described by Horsch [42]. The T1 seeds collected from self-pollinated T0 plants were germinated on MS medium with 50 mg/L Glufosinate ammonium (Sigma, USA) to produce T1 transgenic plants. T2 seeds were collected and the plants were cultured for functional characterizations. The transgenic plants were confirmed by PCR screening of both genomic DNA and cDNA using gene specific primers.
Pathogen inoculation assay
We used several approaches to evaluate the effect of GmCYP82A3 on plant resistance to pathogens. The uniform leaves cut from 7-week-old WT (wild type plant), EV (a transgenic line expressing empty vector as a negative control), and two GmCYP82A3 overexpressing lines (2–3 and 4–1) were placed in Petri dishes, as described above for the soybean leaves. The detached leaves were infected with both necrotrophic B. cinerea and semi-biotrophic P. parasitica. For B. cinerea inoculation, we placed a 5 mm circular potato dextrose agar (PDA) agar containing mycelia cut from the edge of fresh cultured B. cinerea on the leaves. The diameters of the disease lesions were measured and the infected leaves were photographed at 4 days post-inoculation (dpi). For P. parasitica inoculation, the detached leaves were inoculated with 20 μl (approximately 5×104 zoospores ml-1) of zoospore suspension. The WT and EV leaves were used as controls. The diameters of disease lesions were measured at 24 and 48 hpi, and the lesion area was calculated. Statistical significance was determined according to the Dunnett t-test method.
Two-week-old plants (T2 transgenic and WT) hydroponically grown in Hoagland liquid medium were inoculated with P. parasitica zoospore suspension. The roots were immersed in an approximately 5000 ml-1 zoospore suspension. The infected plants were then stored in the growth chamber and photographed at 2 and 5 dpi.
Seven-week-old plants were also used to determine the resistance level. One ml zoospore suspension (approximately 5×104 zoospores ml-1) was dripped in a 1 ml deep hole close to the plant root for inoculation. Twenty plants from each line (WT, EV, 2–3 and 4–1) were infected. Plants with susceptible phenotypes were photographed at 2 and 5 dpi.
Staining with trypan blue
To monitor cell death of N. benthamiana leaves and hypha growth of P. parasitica, the inoculated leaves were stained with lactophenol-trypan blue (10 ml of lactic acid, 10 ml of glycerol, 10 g of phenol, and 10 mg of trypan blue, dissolved in 10 ml of distilled water) [43]. After boiling for 5 min in the staining solution and de-staining in 2.5 g/ml chloral hydrate, the samples were mounted in 70% glycerol for microscopic observation.
RNA isolation, semi-quantitative and quantitative real-time PCR
Total RNA was extracted using the Total RNA kit (Tiangen, CHINA), gDNA elimination and reverse transcription were performed with the PrimeScript™ RT reagent kit (TaKaRa, JAPAN). Quantitative real-time PCR was performed with the ABI PRISM 7500 real-time PCR system (Applied Biosystems, USA) using the AceQ® qPCR SYBR Green Master Mix (Vazyme, CHINA) according to the manufacturer's instructions. The relative expression levels of target genes were calculated using the 2–ΔΔCT method [44]. The significant difference of the genes expression was determined according to the Dunnett t-test method compared to WT. The genes encoded soybean GmTubulin alpha-3 (GmTUA) (GenBank: XM_006584532.1) [45] and N. benthamiana NbEF1α (GenBank: AY206004.1) [46] were selected as the references for soybean and N. benthamiana, respectively. A 476-bp specific sequence of GmCYP82A3 was used to test transcription induction by P. sojae in soybean and transgenic N. benthamiana plants screened by RT-PCR. All the nucleic acid sequence of the primers list in S1 Table.
Tolerance of transgenic N. benthamiana plants to abiotic stresses and JA
The seeds of WT, EV and two overexpression lines (2–3 and 4–1) were surface sterilized in 70% ethanol for 30 s, followed by 30% NaClO for 5 min, then washed at least five times with sterile distilled water. The seeds were placed in solid Murashige and Skoog (MS) medium with or without 100 mM NaCl or 8% PEG6000.
Germination rates were calculated from the percentage of seeds with radicles protruding through the seed coat. The assays were replicated at least three times, using 40 seeds each time. Seeds were geminated in the growth chamber with an 8D:16L photoperiod, 25°C, and 60% relative humidity.
For JA tolerance assays, the WT and transgenic tobacco plants (EV, 2–3, and 4–1) were germinated on MS medium containing 10 μM JA (Sigma, USA). The plates were placed in the chamber for 2 weeks to measure root elongation.
Results
GmCYP82A3 highly expressed in resistant soybean cultivars and responds to P. sojae infection
Gma.3136.2.A1_s_at was highly induced by P. sojae in cultivars with high level of partial resistance based on an Affymetrix GeneChip microarray data analysis [23]. This gene was up-regulated about 5-fold at both 3 and 5 dpi in Conrad, a cultivar with high level of partial resistance, and about 10-fold at 5 dpi in General, another highly partial resistant cultivar. However, the gene was weakly expressed in the moderately resistant cultivar Williams and the susceptible cultivar Sloan (Fig 1A).
Fig 1. The expression pattern of GmCYP82A3 during P. sojae infection.
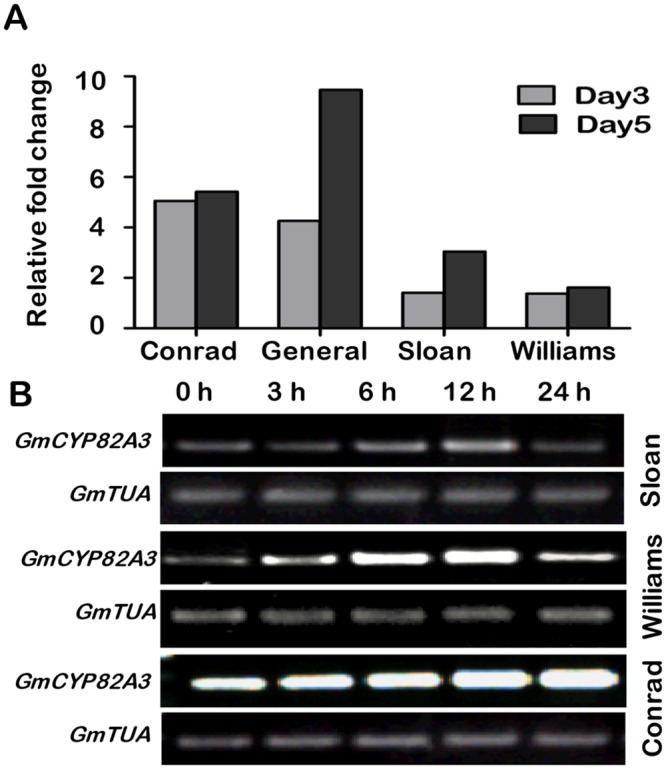
(A) The microarray data of relative expression levels of GmCYP82A3. The expression levels in four soybean cultivars with different resistance level (Conrad and General with highly partial resistance, Williams with moderately partial resistance and Sloan is highly susceptible). The relative expression levels were normalized to the mock samples. (B) Expression pattern of GmCYP82A3 induced by P. sojae determined by RT-PCR. Soybean leaves of Sloan, Williams and Conrad were inoculated with P. sojae and samples were taken at 0, 3, 6, 12 and 24 hpi. The GmTUA gene serves as a control.
Sequence analysis of this soybean EST showed that this gene is GmCYP82A3 (GenBank: 359806337) and encodes a putative protein with 527-amino acids. GmCYP82A3 is a eukaryotic P450 protein, and shares 52% identity with cotton GhCYP82D1 [37], 49% and 48% identity with Arabidopsis AtCYP82C4 and AtCYP82C2 [29, 38] (S1 Fig).
To validate the response of GmCYP82A3 to P. sojae infection, RT-PCR analysis was used to determine its expression profile in soybean leaves at 0, 3, 6, 12, and 24 hpi. GmCYP82A3 was induced in Sloan, Williams and Conrad. In Sloan, GmCYP82A3 was up-regulated after P. sojae infection, and reached the highest level at 12 hpi. In Williams, GmCYP82A3 showed a similar expression pattern as in Sloan, but the expression level was much stronger. The transcription level was obviously up-regulated at 3 hpi, reached the highest level at 12 hpi, and was subsequently reduced at 24 hpi. The initial level in Conrad was much higher than in Williams, and continuously increased at later time points (Fig 1B).
Expression of GmCYP82A3 is induced by abiotic stresses and phytohormones
Then, we characterized the expression patterns of GmCYP82A3 response to abiotic stresses in Conrad, using qRT-PCR. In general, the expression of GmCYP82A3 was dramatically up-regulated in both PEG6000 and NaCl treatments, but with different trends. When treated with NaCl, the expression level was up-regulated 10-fold at 6 hpt and 64-fold at 12 hpt, then maintained high levels at subsequent time points. When treated with PEG6000, expression was up-regulated 164-fold at 6 hpt and then dramatically reduced at later time points (Fig 2).
Fig 2. The expression profiles of GmCYP82A3 response to various abiotic stress and phytohormones.
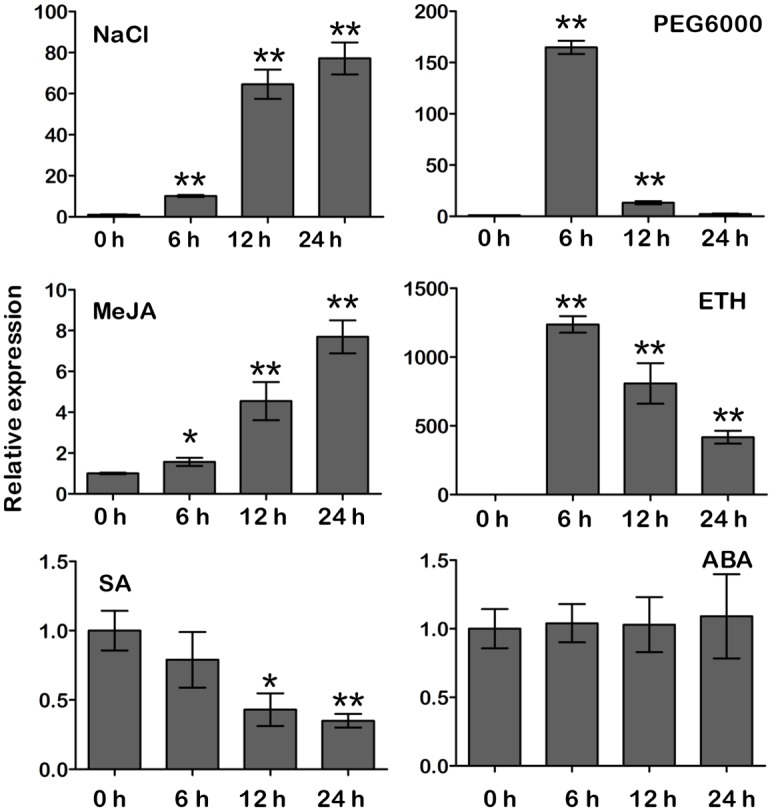
Three-week-old seedlings were treated with the indicated compounds, and then the samples were collected at 0, 6, 12 and 24 hpt. The relative expression level was normalized to soybean GmTUA. Mean and standard deviation (SD) were calculated from three independent biological replicates. The asterisk at the top of the columns indicate significant differences (Dunnett-t test, * P<0.05, ** P<0.01).
Previous studies showed that some CYP82 family genes that share high identity with GmCYP82A3 could be induced by phytohormones [37, 38]. To examine responses to phytohormones, soybean seedlings of Conrad were treated with methyl jasmonate (MeJA), ethephon (ETH), salicylic acid (SA), and abscisic acid (ABA). As shown in Fig 2, expression levels were enhanced at all time points after MeJA treatment, and up-regulated about 8-fold at 24 hpt. However, it was dramatically up-regulated by more than 1,000-fold at 6 hpt, and then attenuated, but still maintained a high level with ETH treatment. In contrast, a decrease in GmCYP82A3 expression was observed when treated with SA, and there was no response to ABA treatment.
GmCYP82A3 overexpression enhances resistance of N. benthamiana to B. cinerea and P. parasitica
To explore the role of this P450 family gene, GmCYP82A3 was overexpressed in N. benthamiana plants driven by the 35S promoter and two independent T2 generations were selected for functional characterizations. Transgene integration and expression were confirmed by genomic PCR and RT-PCR, respectively (S2 Fig). No obvious phenotypic differences were observed between the WT, EV, and GmCYP82A3 overexpressing (2–3 and 4–1) seedlings. We first challenged the detached leaves with B. cinerea, a necrotrophic pathogen and causes necrotic symptoms on N. benthamiana (Fig 3A, S3 Fig). The average sizes of the lesion areas were ~ 357.15 and ~ 344.54 mm2 at 4 dpi in WT and EV leaves, respectively. However, the necrotic areas were significantly reduced in the two independent overexpression lines. The average sizes of the lesion areas were ~ 201.97 and ~ 250.12 mm2 (Fig 3B). The results showed that GmCYP82A3 could enhance resistance to B. cinerea.
Fig 3. Enhanced resistance to B. cinerea of transgenic N. benthamiana lines.
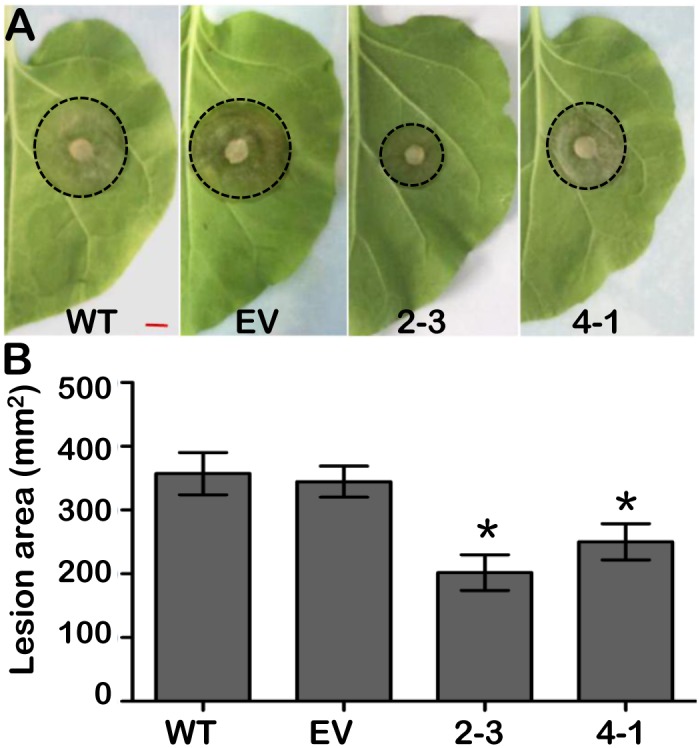
(A) Phenotypes of the N. benthamiana leaves from WT, EV and overexpression lines (2–3 and 4–1) inoculated with B. cinerea. The mycelia growing on PDA medium was used to infect the transgenic and wild type leaves. Photographs were taken 4 dpi. Bar = 5 mm. (B) Lesion area of inoculated leaves. Lesion diameters were measured at 4 dpi and then the lesion area was calculated. Similar results were observed at least three duplications. SD represented with the bars (Dunnett-t test: * P<0.05).
To further study the disease resistance function of GmCYP82A3, we also characterized the resistance level of transgenic plants to P. parasitica, a hemibiotrophic pathogen that is similar to P. sojae. When the hydroponically cultured 2-week-old plants were inoculated with P. parasitica zoospores, the WT and EV plants withered at 2 dpi, then exhibited stem rot and collapse of the whole plant at 5 dpi. In contrast, the two overexpression lines were more resistant and the symptoms were much weaker (Fig 4A).
Fig 4. Increased resistance to P. parasitica of transgenic N. benthamiana lines.

(A) Phenotypes of the 2-week-old GmCYP82A3 transgenic plants inoculated with P. parasitica zoospores. Photographs were taken at 2 and 5 dpi, respectively. (B) Phenotypes of the 7-week-old GmCYP82A3 transgenic plants inoculated with P. parasitica zoospores. Photographs were taken at 2 and 5 dpi, respectively. The photos of the bottom row are high magnification views of the red pane marked in the upper row. (C) Lesion area of the inoculated leaves measured at 24 and 48 hpi. The lesion area was calculated from over three repeats. SD represented with the bars (Dunnett-t test: * P<0.05, ** P<0.01). (D) Detached leaves from WT, EV and GmCYP82A3 overexpression plants were inoculated with P. parasitica zoospores. Photographs were taken at 48 hpi under a UV lamp. Bar = 5 mm. (E) Trypan blue staining of the P. parasitica inoculated sites of N. benthamiana leaves. The hypha accumulation in the WT, EV and GmCYP82A3 overexpression leaves were detected using trypan blue staining at 12 and 24 hpi. Bar = 100 μm.
The root inoculation assay performed on 7-week-old plants to confirm the resistant phenotype. Inoculated seedlings displayed wilting symptoms at 2 dpi. However, the brown and necrotic areas on stems progressively expanded from the roots, causing stem wither and lodge in WT and EV plants at 5 dpi. Symptoms were restricted or expanded slightly in the two overexpression plant lines during the same period (Fig 4B).
The detached transgenic leaves were also used to test for resistance. The average lesion areas caused by P. parasitica zoospores infection were significantly smaller on GmCYP82A3 overexpression leaves than those on WT and EV leaves, at both 24 and 48 hpi (Fig 4C and 4D). Trypan blue staining showed fewer hyphae on GmCYP82A3 overexpression leaves (Fig 4E). These results imply that GmCYP82A3 contributes to resistance to the two tested pathogens.
GmCYP82A3 overexpression N. benthamiana plants are insensitive to JA
JA plays an important role in plant defense signaling pathway and it also inhibits root growth [47]. Both GhCYP82D1-silenced cotton and AtCYP82C2 mutant Arabidopsis seedlings were hypersensitive to exogenous JA treatment [37, 38]. We evaluated whether GmCYP82A3 could alter the sensitivity of plants to JA. Under normal growth conditions, the overall growth rate and morphology of GmCYP82A3 overexpression plants were similar to WT and EV plants. However, when 10 μM JA were added to the medium, roots of the two selected lines were significantly longer than those of WT and EV roots (Fig 5), suggesting that the plants expressing GmCYP82A3 were less sensitive to JA-mediated inhibition of root growth. The results above imply that GmCYP82A3 might be involved in the JA signaling pathway.
Fig 5. The effect of GmCYP82A3 on root growth under JA treatment.
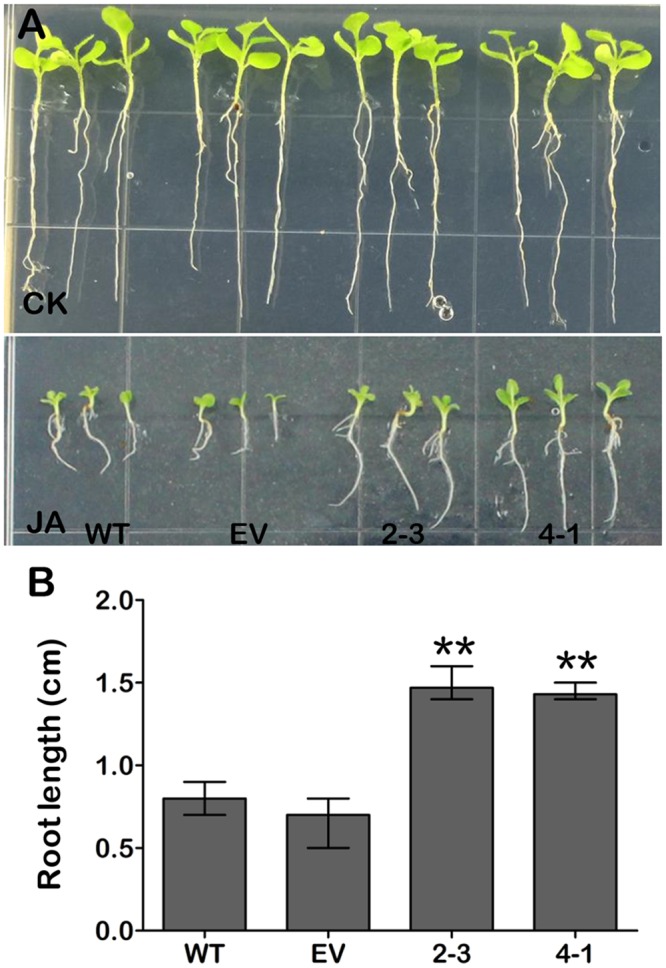
(A) The phenotypes of 2-week-old seedlings of indicated genotypes grown on MS medium without (Control) or with 10 μΜ JA. (B) Root length of 2-week-old WT, EV and GmCYP82A3 overexpression seedlings grown on MS medium containing 10 μΜ JA. The experiments were repeated three times with similar results. SD represented with the bars. The asterisk at the top of the columns indicate significant differences (Dunnett-t test, ** P<0.01).
GmCYP82A3 affects the expression of defense related marker genes
Because of changes of resistance levels to P. parasitica infection, we investigated GmCYP82A3 overexpression plants for the expression pattern of defense related marker genes in the defense cascade, and to gain further insight into possible regulation pathways. Four well known PR (pathogenesis related) genes were firstly selected for comparison. The SA marker PR1 (Pathogenesis-related protein 1) and PR2 (Pathogenesis-related protein 2) [48] were significantly lower in GmCYP82A3 overexpression plants than those in WT and EV plant before and after infection, except PR1 showed no different at 12 hpi. In contrast, the basal expression levels of another two genes response to JA/ET, PR3 (B-chitinase) and PR4 (Hevein-like protein) [49], were significantly higher in the two overexpression lines than in EV and WT plants. Both PR3 and PR4 greatly increased after infection with P. parasitica, and also had a higher expression level in overexpression plants. JA and ET regulated defense related marker gene PDF1.2 (Plant defensing 1.2) [49] significantly increased in overexpression plants and the basal expression levels were about 11 and 8-fold higher in 2–3 and 4–1 lines than that in WT. Meanwhile, PDF1.2 expression induced by P. parasitica infection was also significantly higher in overexpression plants at 12 and 24 hpi (Fig 6).
Fig 6. Expression profiles of defense marker genes in transgenic N. benthamiana plants.

Total RNA was extracted from detached leaves of WT, EV and GmCYP82A3 overexpression plants at 0, 12, 24 hpi by P. parasitica zoospores. Expression levels were determined by qRT-PCR using gene specific primers and normalized to NbEF1a with three replicate experiments. Data are the means of three replications, error bars indicate SD. The significant differences between WT and transgenic plants are indicated by asterisk (Dunnett-t test, * P<0.05, ** P<0.01).
GmCYP82A3 is involved in the JA/ET signaling pathway
For further detecting the signaling transduction effected by GmCYP82A3, several key regulators in the JA/ET signaling pathways were selected for expressional analysis. LOX1 (Lipoxygenase 1) is involved in JA biosynthesis and signaling pathway [50]. Its basal expression was not effected between WT and transgenic plants, but the expression exhibited more intense inductive effect after infection in overexpression plants. JAR1 (Jasmonate resistant 1) catalyzes JA conjugated to isoleucine, resulting in biologically highly active specific enantiomer of jasmonoyl-isoleucine (JA-Ile) [51]. COI1 (Coronatine insensitive 1) encodes an F-box protein to assemble SCFCOI1 protein complex, act as JA-Ile receptor in JA signaling [52]. In the expressional analysis, JAR1 was induced at 12 hpi and then reduced at 24 hpi, although, the expression levels were significantly higher in overexpression plants both before and after infection. The expression of COI1 down-regulated after infection, and there was no difference between the lines at both 0 and 12 hpi, but up-regulated and significantly higher in overexpression lines at 24 hpi.
The downstream of JA signaling occurs via two different branches, which regulated by MYC2 (Myelocytomatosis protein 2) or ERF1 (Ethylene response factor 1) respectively [49]. In this study, MYC2 and VSP2 (Vegetative storage protein 2) [53] selected as the MYC branch marker genes. The results indicated that the basal expression of MYC2 and VSP2 were suppressed in GmCYP82A3 overexpression plants. After infection, both genes were decreased, and MYC2 was no difference between overexpression and control plants, but VSP2 was significantly lower in overexpression plants at 24 hpi. In ERF branch, ERF1 plays a crucial role in the cross talk between JA and ET signaling pathway [54, 55], the expression was induced by P. parasitica infection. In overexpression plants, the expression of ERF1 showed significantly higher both before and after infection (Fig 7).
Fig 7. Expression profiles of key JA/ET signaling pathway genes in transgenic N. benthamiana plants.

Data are the means of three replications, error bars indicate SD (Dunnett-t test, * P<0.05, ** P<0.01).
In ET signaling pathway, EIN3 (Ethylene insensitive 3) positive regulate downstream transcription of ethylene response [56], the expression was induced at 12 hpi and then reduced at 24 hpi, and exhibited significantly higher in overexpression plants than that in EV and WT plants. EBF2 (Ein3-binding F box protein 2) which regulate EIN3 protein degradation [56], the expression was suppressed both before and after infection compared with WT and EV. (Fig 7). These results indicated that overexpression GmCYP82A3 disturbed the key regulate genes expression, suggesting that GmCYP82A3 might be involved in the JA/ET-mediated signaling pathway and contributes to delayed disease development when challenged with P. parasitica and B. cinerea on detached leaves.
GmCYP82A3 overexpression enhances seed germination under salt and osmotic stress
Given that the expression of GmCYP82A3 was significantly induced by salt and drought stress (Fig 2), we conducted tests to determine whether GmCYP82A3 could regulate the response of the transgenic plants to abiotic stresses. Germination rates of the transgenic and WT seeds were calculated under abiotic stress. The germination of WT, EV, and overexpression seeds showed no obvious differences on the MS medium without NaCl or PEG6000 (Fig 8A). Under treatment with 100 mM NaCl and 8% PEG 6000, germination rates of the seeds showed significant difference. The germination of WT and EV seeds were severely suppressed under the stresses. However, the germination rates of GmCYP82A3 overexpression seeds were higher compared with the WT and EV seeds at 4–7 and 4–5 days after sowing on MS medium containing 100 mM NaCl and 8% PEG 6000 respectively (Fig 8B and 8C). Imply that enhanced tolerance to salt and drought stress of GmCYP82A3 overexpression seeds.
Fig 8. Seed germination rates of transgenic N. benthamiana under abiotic stress.

The seeds as indicated were cultured on MS medium (A) or containing NaCl (B) or PEG6000 (C). The germination rates were calculated from the percentage of seeds with radicle protruded through the seed coat after cultivation every day. Mean values and SD were obtained from five independent experiments (n = 40). The significant differences between WT and transgenic plants are indicated by asterisk (Dunnett-t test, * P<0.05, ** P<0.01).
Discussion
Since partial resistance is important for soybean breeding resistance to Phytophthora root and stem rot, attempts have been made to identify the functional candidate genes conferring this resistance. A wide list of candidate genes underlying the soybean QTL conferring resistance to P. sojae were identified by whole-genome transcription profiling analysis of cultivars with different levels of resistance [13, 14, 20]. In this study, we performed a functional analysis of the soybean P450 gene, called GmCYP82A3, which is highly expressed in partial resistant soybean cultivars during P. sojae infection and underlying the QTL conferring resistance to P. sojae [14]. Our results indicated that the expression level of GmCYP82A3 can be induced by various abiotic stresses and phytohormones molecules. The overexpressed transgenic N. benthamiana plants showed enhanced resistance to the necrotrophic pathogen B. cinerea and the hemibiotrophic pathogen P. parasitica, implying that this gene underlying the QTL region may contribute to plant resistance. Further examination found the transgenic plants were less sensitive to jasmonic acid (JA), and the enhanced resistance accompanied with increased expression of the JA/ET signaling pathway genes.
Previous studies revealed that members of CYP82 family are highly stress responsive in tobacco, pea, soybean, and Arabidopsis [28–31]. In this study, GmCYP82A3 was induced by P. sojae infection and showed a different transcription profiles in soybean cultivars with diverse partial resistant levels, which consists with that from the candidate genes identification of soybean partial resistant QTL [13]. Besides the pathogen induction, GmCYP82A3 was remarkably up-regulated by salt and drought treatment. Phytohormones are widely believed to play key roles in signaling transduction involved in plant responses biotic and abiotic stresses [57–59]. GmCYP82A3 was highly induced by MeJA and ETH, but reduced in some degree when treated with SA, and did not respond to ABA treatment. This was consistent with previous reports, in which cotton and Arabidopsis CYP82 genes were both highly induced by JA/MeJA treatment [37, 38]. Thus, we speculated that this soybean P450 gene may play important role in abiotic stress and defense processes.
JA, ET, SA signaling and the cross-talk between them play a role in plant defense response activation upon pathogen infection. For example, SA mediates defense responses to biotrophic pathogens, JA/ET mediates defense against necrotrophic or hemibiotrophic pathogens and usually antagonism with SA [58, 59]. Transcriptional analysis of phytohormone signaling marker genes revealed that GmCYP82A3 overexpression affected the phytohormone signaling transduction. Two SA signaling marker genes, PR1 and PR2 were suppressed in transgenic plants both before and after pathogen infection compare with control, especially PR2. The results proved that GmCYP82A3 partly suppressed SA signaling pathway. PR3, PR4 and PDF1.2, which involved in JA and ET dependent resistance were significantly higher expression levels in overexpression plants [49]. We speculated that GmCYP82A3 might be involved in the JA/ET signaling pathway. In this study, the root growth in response to JA of GmCYP82A3 overexpression plants showed a similar phenotype of AtCYP82C2, which the Arabidopsis overexpression plants also showed less sensitive to JA-mediated root growth inhibition [38], implying that GmCYP82A3 is involved in JA signaling. LOX1 has been widely demonstrated to be involved in JA biosynthesis and signaling pathway [50], JAR1 catalyzes the formation of a biologically active JA-Ile conjugate [51]. COI1 encodes an F-box protein to assemble SCFCOI1 protein complex, act as JA-Ile receptor in JA signaling. The binding of JA-Ile to COI1 leads degradation of JASMONATE ZIM-domain (JAZ) transcriptional repressor proteins, and results in the activation of JA responsive genes [52]. The higher expression levels of LOX1, JAR1, and COI1 in GmCYP82A3 overexpression plants after P. parasitica infection suggest that the JA signaling transduction may be enhanced. At least two distinct branches lie in JA signaling pathway to regulates downstream genes expression [49, 53]. ERF branch confer resistance to necrotrophic pathogens and MYC2 branch is associated with wound response and insect herbivores resistance, but MYC2 branch also has been demonstrated to play a role for enhance pathogen defense [49, 60, 61]. Alternatively, the MYC branch has an antagonistic effect with ERF branch, AtMYC2 represses the expression of ERF branch genes while activating the wound responsive genes such as VSP2 in MYC branch [53]. The ERF branch and downstream defense relate genes ERF1, PDF1.2, PR3 and PR4 showed significant up-regulated in GmCYP82A3 overexpression plants. In contrast, the expression of MYC2 and VSP2 which involved in the MYC branch were suppressed. The results demonstrated that ectopic expression GmCYP82A3 enhanced the signaling transduction of JA ERF branch during the transgenic N. benthamiana plants and P. parasitica interaction.
Ethylene regulates wide physiological responses in plants. Ethylene is the first plant hormone, the signaling pathway has provided a framework in Arabidopsis. EIN3 is a plant specific nuclear transcription factor and positive regulate downstream transcription of ethylene response, such as the target gene of ERF1 [55, 56]. The critical regulatory mechanism of ethylene signaling in the nucleus is controlled by EIN3 protein levels. In the absence of ethylene, EIN3 is rapidly degraded by 26S proteasomal under the regulation of two F-box proteins EBF1 and EBF2. In the presence of ethylene, EBF1 and EBF2 are degraded, thus allowing EIN3 protein accumulation and activated ethylene responsive gene expression [56]. In this study, transcription profiles of EIN3 during the pathogen infection was elevated in GmCYP82A3 overexpression plants, whereas, EBF2 showed suppression effect. In Arabidopsis, the ET and ERF branch of JA signaling act synergistically on the defense related genes expression, such as PDF1.2, ERF1, PR3 and PR4 [54, 62–64]. ERF1 transcript was induced by B. cinerea infection, and the overexpressed Arabidopsis plants showed enhanced resistance to necrotrophic fungi such as B. cinerea and Plectospaerella cucumerina, but reduced tolerance to biotrophic Pst DC3000 [65]. We believed that the ET signaling transduction was activated and contributed to the enhanced resistance.
Besides play essential roles in regulating plant defense against pathogens as discussed above, JA/ET signaling also contributed to plant abiotic stress tolerance such as salt and drought [57]. Transgenic expression of several genes involved in JA biosynthesis and signaling pathway showed enhanced salt tolerance [66–68]. The pathway component EIN3 performed as a positive regulator on salt stress tolerance, further studies found the downstream ERF1 selectively actives salt tolerance genes by binding to the DRE-box of these genes promoter [69, 70]. Drought tolerance can be mediated through a wide range of mechanisms. Till now, JA and ET signaling pathways are also implicated in drought tolerance [57].
In this study, ectopic expression soybean GmCYP82A3 in N. benthamiana enhanced resistance to two different kinds of pathogens and salt, drought stress tolerance. The transcription analysis of phytohormones pathway genes revealed that PR1 and PR2 involved in SA signaling was suppressed, on the other hand, the ERF branch of JA and ET signaling genes were upregulated in the transgenic plants, indicating that the ERF branch of JA and ET signaling pathways were activated by GmCYP82A3. But the potential mechanism is still needed to be explored. Since several CYP82 family members participate in a variety of metabolic pathways [32–38], so further characterization of the biochemical function is needed.
As a form of incomplete resistance in the P. sojae-soybean system and non-race-specific resistance, partial resistance has been proposed as a way to improve the breeding efforts for soybean resistance to P. sojae. Previous studies showed that some cultivars with high levels of partial resistance were difficult to distinguish from the Rps genotype when inoculated with the simple isolate [71]. In this study, we found that GmCYP82A3 affected defense related genes expression associated with the JA/ET signaling pathway, conferring transgenic plants resistant to two different types of pathogens. From our understanding of the mechanism of soybean partial resistance, we believe that the strategy of generate highly partial resistant soybean cultivars offer a promising approach for Phytophthora root rot control.
Supporting Information
(DOCX)
Sequence alignment was done by ClustalW. Black and gray backgrounds indicate identical and similar residues, respectively; dotted lines indicate gaps.
(TIF)
Electrophoresis pattern of PCR from genomic DNA (A) and cDNA (B) corresponding to GmCYP82A3 (upper panel) of wild-type (WT), empty vector (EV) transformants (EV-1, EV-2) and the six T2 transgenic lines expressing GmCYP82A3 (2–3, 3–3, 4–1, 10–1, 15–2, and 9–1). The NbEF1a (lower panel) was used as an internal control. M, DNA Marker DL2000 PLUS.
(TIF)
(A) Phenotypes of the N. benthamiana leaves from WT, EV and overexpression lines (2–3 and 4–1) inoculated with B. cinerea at 4 dpi. Bar = 5 mm. (B) Lesion area of inoculated leaves. Lesion diameters were measured at 4 dpi and then the lesion area was calculated. SD represented with the bars (Dunnett-t test: ** P<0.01).
(TIF)
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This work was supported by grants from National Science and Technology Major Projects (2014ZX0800910B), Daolong Dou, http://www.nmp.gov.cn/; National Natural Science Foundation of China (31171831), Daolong Dou, http://www.nsfc.gov.cn/nsfc/cen/2016sqsl/index.html; Jiangsu Planned Projects for Postdoctoral Research Funds (1501046A), Qiang Yan, http://bsh.jshrss.gov.cn/. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Rajcan I, Hou G, Weir AD. Advances in breeding of seed-quality traits in soybean. J Crop Improv. 2005;14(1–2):145–74. 10.1300/J411v14n01_07 [DOI] [Google Scholar]
- 2.Clemente TE, Cahoon EB. Soybean oil: genetic approaches for modification of functionality and total content. Plant Physiol. 2009;151(3):1030–40. 10.1104/pp.109.146282 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Mushrush GW, Wynne JH, Willauer HD, Lloyd CL. Soybean-derived biofuels and home heating fuels. J Environ Sci Heal A. 2006;41(11):2495–502. 10.1080/10934520600927591 . [DOI] [PubMed] [Google Scholar]
- 4.Tyler BM. Phytophthora sojae: root rot pathogen of soybean and model oomycete. Mol Plant Pathol. 2007;8(1):1–8. 10.1111/j.1364-3703.2006.00373.x . [DOI] [PubMed] [Google Scholar]
- 5.Sugimoto T, Kato M, Yoshida S, Matsumoto I, Kobayashi T, Kaga A, et al. Pathogenic diversity of Phytophthora sojae and breeding strategies to develop Phytophthora-resistant soybeans. Breeding Sci. 2012;61(5):511–22. 10.1270/jsbbs.61.511 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Walker AK, Schmitthenner AF. Heritability of tolerance to Phytophthora rot in Soybean. Crop Sci. 1984;24(3):490–1. 10.2135/cropsci1984.0011183X002400030014x [DOI] [Google Scholar]
- 7.Tooley PW, Grau CR. The relationship between rate-reducing resistance to Phytophthora megasperma f.sp.glycinea and yield of soybean. Phytopathology. 1984;74(10):1209–16. GUA85007882. [Google Scholar]
- 8.Tyler BM, Jiang RHY, Zhou LC, Tripathy S, Dou DL, Torto-Alalibo T, et al. Functional genomics and bioinformatics of the Phytophthora sojae soybean interaction In: Gustafson JP, Taylor J, Stacey G, editors. Genomics of disease. stadler genetics symposia series: Springer; New York; 2008. pp. 67–78. 10.1007/978-0-387-76723-9_6. WOS:000254891400006. [DOI] [Google Scholar]
- 9.Dorrance AE, McClure SA, St. Martin SK. Effect of partial resistance on Phytophthora stem rot incidence and yield of soybean in Ohio. Plant Dis. 2003;87(3):308–12. 10.1094/PDIS.2003.87.3.308 [DOI] [PubMed] [Google Scholar]
- 10.Burnham KD, Dorrance AE, VanToai TT, St. Martin SK. Quantitative trait loci for partial resistance to Phytophthora sojae in soybean. Crop Sci. 2003;43(5):1610–7. [Google Scholar]
- 11.Han Y, Teng W, Yu K, Poysa V, Anderson T, Qiu L, et al. Mapping QTL tolerance to Phytophthora root rot in soybean using microsatellite and RAPD/SCAR derived markers. Euphytica. 2008;162(2):231–9. 10.1007/s10681-007-9558-4. IND44065363. [DOI] [Google Scholar]
- 12.Weng C, Yu K, Anderson TR, Poysa V. A quantitative trait locus influencing tolerance to Phytophthora root rot in the soybean cultivar 'Conrad'. Euphytica. 2007;158(1–2):81–6. [Google Scholar]
- 13.Wang H, Waller L, Tripathy S, St. Martin SK, Zhou L, Krampis K, et al. Analysis of genes underlying soybean quantitative trait loci conferring partial resistance to Phytophthora sojae. Plant Genome. 2010;3(1):23–40. 10.3835/plantgenome2009.12.0029 [DOI] [Google Scholar]
- 14.Wang H, St. Martin SK, Dorrance AE. Comparison of phenotypic methods and yield contributions of quantitative trait loci for partial resistance to Phytophthora sojae in soybean. Crop Sci. 2012;52(2):609–22. [Google Scholar]
- 15.Wu X, Zhou B, Zhao J, Guo N, Zhang B, Yang F, et al. Identification of quantitative trait loci for partial resistance to Phytophthora sojae in soybean. Plant Breed. 2011;130(2):144–9. 10.1111/j.1439-0523.2010.01799.x. IND44518486. [DOI] [Google Scholar]
- 16.Lee S, Mian MAR, McHale LK, Wang H, Wijeratne AJ, Sneller CH, et al. Novel quantitative trait loci for partial resistance to Phytophthora sojae in soybean PI 398841. Theor appl genet. 2013;126(4):1121–32. 10.1007/s00122-013-2040-x . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Tucker DM, Saghai Maroof MA, Mideros S, Skoneczka JA, Nabati DA, Buss GR, et al. Mapping quantitative trait loci for partial resistance to Phytophthora sojae in a soybean interspecific cross. Crop Sci. 2010;50(2):628–35. 10.2135/cropsci2009.03.0161 [DOI] [Google Scholar]
- 18.Nguyen VT, Vuong TD, VanToai T, Lee JD, Wu X, Mian MAR, et al. Mapping of quantitative trait loci associated with resistance to Phytophthora sojae and flooding tolerance in soybean. Crop Sci. 2012;52(6):2481–93. 10.2135/cropsci2011.09.0466 [DOI] [Google Scholar]
- 19.Lee S, Mian MAR, Sneller CH, Wang H, Dorrance AE, McHale LK. Joint linkage QTL analyses for partial resistance to Phytophthora sojae in soybean using six nested inbred populations with heterogeneous conditions. Theor appl genet. 2014;127(2):429–44. 10.1007/s00122-013-2229-z . [DOI] [PubMed] [Google Scholar]
- 20.Vega-Sánchez ME, Redinbaugh MG, Costanzo S, Dorrance AE. Spatial and temporal expression analysis of defense-related genes in soybean cultivars with different levels of partial resistance to Phytophthora sojae. Physiol Mol Plant P. 2005;66(5):175–82. 10.1016/j.pmpp.2005.07.001 [DOI] [Google Scholar]
- 21.Ranathunge K, Thomas RH, Fang X, Peterson CA, Gijzen M, Bernards MA. Soybean root suberin and partial resistance to root rot caused by Phytophthora sojae. Phytopathology. 2008;98(11):1179–89. 10.1094/PHYTO-98-11-1179 . [DOI] [PubMed] [Google Scholar]
- 22.Thomas R, Fang X, Ranathunge K, Anderson TR, Peterson CA, Bernards MA. Soybean root suberin: anatomical distribution, chemical composition, and relationship to partial resistance to Phytophthora sojae. Plant Physiol. 2007;144(1):299–311. 10.1104/pp.106.091090 ; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Zhou L, Mideros SX, Bao L, Hanlon R, Arredondo FD, Tripathy S, et al. Infection and genotype remodel the entire soybean transcriptome. BMC Genomics. 2009;10:49 10.1186/1471-2164-10-49 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Mizutani M, Ohta D. Diversification of P450 genes during land plant evolution. Annu Rev Plant Biol. 2010;61:291–315. 10.1146/annurev-arplant-042809-112305 . [DOI] [PubMed] [Google Scholar]
- 25.Mizutani M. Impacts of diversification of cytochrome P450 on plant metabolism. Biol Pharm Bull. 2012;35(6):824–32. . [DOI] [PubMed] [Google Scholar]
- 26.Guttikonda SK, Trupti J, Bisht NC, Chen H, An YQ, Pandey S, et al. Whole genome co-expression analysis of soybean cytochrome P450 genes identifies nodulation-specific P450 monooxygenases. BMC Plant Biol. 2010;10:243 10.1186/1471-2229-10-243 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Nelson DR, Schuler MA, Paquette SM, Werck-Reichhart D, Bak S. Comparative genomics of rice and Arabidopsis. Analysis of 727 cytochrome P450 genes and pseudogenes from a monocot and a dicot. Plant Physiol. 2004;135(2):756–72. 10.1104/pp.104.039826 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Schopfer CR, Ebel J. Identification of elicitor-induced cytochrome P450s of soybean (Glycine max L.) using differential display of mRNA. Mol Gen Genet. 1998;258(4):315–22. . [DOI] [PubMed] [Google Scholar]
- 29.Murgia I, Tarantino D, Soave C, Morandini P. Arabidopsis CYP82C4 expression is dependent on Fe availability and circadian rhythm, and correlates with genes involved in the early Fe deficiency response. J Plant Physiol. 2011;168(9):894–902. 10.1016/j.jplph.2010.11.020 . [DOI] [PubMed] [Google Scholar]
- 30.Ralston L, Kwon ST, Schoenbeck M, Ralston J, Schenk DJ, Coates RM, et al. Cloning, heterologous expression, and functional characterization of 5-epi-aristolochene-1,3-dihydroxylase from tobacco (Nicotiana tabacum). Arch Biochem Biophys. 2001;393(2):222–35. 10.1006/abbi.2001.2483 . [DOI] [PubMed] [Google Scholar]
- 31.Frank MR, Deyneka JM, Schuler MA. Cloning of wound-induced cytochrome P450 monooxygenases expressed in pea. Plant Physiol. 1996;110(3):1035–46. 10.1104/pp.110.3.1035 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Siminszky B, Gavilano L, Bowen SW, Dewey RE. Conversion of nicotine to nornicotine in Nicotiana tabacum is mediated by CYP82E4, a cytochrome P450 monooxygenase. P Natl Acad Sci USA. 2005;102(41):14919–24. 10.1073/pnas.0506581102 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Takemura T, Ikezawa N, Iwasa K, Sato F. Molecular cloning and characterization of a cytochrome P450 in sanguinarine biosynthesis from Eschscholzia californica cells. Phytochemistry. 2013;91:100–8. 10.1016/j.phytochem.2012.02.013 . [DOI] [PubMed] [Google Scholar]
- 34.Berim A, Gang DR. The roles of a flavone-6-hydroxylase and 7-O-demethylation in the flavone biosynthetic network of sweet basil. J Biol Chem. 2013;288(3):1795–805. 10.1074/jbc.M112.420448 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Lee S, Badieyan S, Bevan DR, Herde M, Gatz C, Tholl D. Herbivore-induced and floral homoterpene volatiles are biosynthesized by a single P450 enzyme (CYP82G1) in Arabidopsis. P Natl Acad Sci USA. 2010;107(49):21205–10. 10.1073/pnas.1009975107 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Kruse T, Ho K, Yoo HD, Johnson T, Hippely M, Park JH, et al. In planta biocatalysis screen of P450s identifies 8-methoxypsoralen as a substrate for the CYP82C subfamily, yielding original chemical structures. Chem Biol. 2008;15(2):149–56. 10.1016/j.chembiol.2008.01.008 . [DOI] [PubMed] [Google Scholar]
- 37.Sun L, Zhu L, Xu L, Yuan D, Min L, Zhang X. Cotton cytochrome P450 CYP82D regulates systemic cell death by modulating the octadecanoid pathway. Nat Commun. 2014;5:5372 10.1038/ncomms6372 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Liu F, Jiang H, Ye S, Chen WP, Liang W, Xu Y, et al. The Arabidopsis P450 protein CYP82C2 modulates jasmonate-induced root growth inhibition, defense gene expression and indole glucosinolate biosynthesis. Cell Res. 2010;20(5):539–52. 10.1038/cr.2010.36 . [DOI] [PubMed] [Google Scholar]
- 39.Schmitthenner AF, Bhat RG. Useful methods for studying Phytophthora in the laboratory. Special circular (Ohio Agricultural Research and Development Center) (USA) no 143. 1994. Available: http://kb.osu.edu/dspace/handle/1811/71884
- 40.Hoagland DR, Arnon DI. The water-culture method for growing plants without soil. Calif Agric Exp Stn Circ. 1950;347(5406):357–9. [Google Scholar]
- 41.Earley KW, Haag JR, Pontes O, Opper K, Juehne T, Song K, et al. Gateway-compatible vectors for plant functional genomics and proteomics. Plant J. 2006;45(45):616–29. 10.1111/j.1365-313X.2005.02617.x . [DOI] [PubMed] [Google Scholar]
- 42.Horsch RB, Rogers SG, Fraley RT. Transgenic plants. Cold Spring Harbor Symposia on Quantitative Biology. 1985;50(6):433–7. 10.1101/SQB.1985.050.01.054 . [DOI] [PubMed] [Google Scholar]
- 43.Hwang IS, Hwang BK. The pepper 9-lipoxygenase gene CaLOX1 functions in defense and cell death responses to microbial pathogens. Plant Physiol. 2010;152(2):948–67. 10.1104/pp.109.147827 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods. 2001;25(4):402–8. 10.1006/meth.2001.1262 . [DOI] [PubMed] [Google Scholar]
- 45.Zeng WT, Chai CY, Dou DL. Selection and validation of reference genes for quantitative RT-PCR analysis in soybean. Journal of Nanjing Agricultural University. 2015;38(5):787–95. [Google Scholar]
- 46.Chai C, Lin Y, Shen D, Wu Y, Li H, Dou D. Identification and functional characterization of the soybean GmaPPO12 promoter conferring Phytophthora sojae induced expression. PLoS One. 2013;8(6):e67670 10.1371/journal.pone.0067670 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Corbineau F, Rudnicki RM, Côme D. The effects of methyl jasmonate on sunflower (Helianthus annuus L.) seed germination and seedling development. Plant Growth Regul. 1988;7(3):157–69. [Google Scholar]
- 48.Lee S, Dong SY, Uppalapati SR, Sumner LW, Mysore KS. Suppression of plant defense responses by extracellular metabolites from Pseudomonas syringae pv. tabaci in Nicotiana benthamiana. BMC Plant Biol. 2013;13(1):1–13. 10.1186/1471-2229-13-65 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Lorenzo O, Solano R. Molecular players regulating the jasmonate signalling network. Curr Opin Plant Biol. 2005;8(5):532–40. 10.1016/j.pbi.2005.07.003 . [DOI] [PubMed] [Google Scholar]
- 50.Porta H, Rocha-Sosa M. Plant lipoxygenases. Physiological and molecular features. Plant Physiol. 2002;130(1):15–21. 10.1104/pp.010787 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Staswick PE, Tiryaki I. The oxylipin signal jasmonic acid is activated by an enzyme that conjugates it to isoleucine in Arabidopsis. Plant Cell. 2004;16(8):2117–27. 10.1105/tpc.104.023549 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Thines B, Katsir L, Melotto M, Niu Y, Mandaokar A, Liu G, et al. JAZ repressor proteins are targets of the SCF(COI1) complex during jasmonate signalling. Nature. 2007;448(7154):661–5. 10.1038/nature05960 . [DOI] [PubMed] [Google Scholar]
- 53.Lorenzo O, Chico JM, Sanchez-Serrano JJ, Solano R. JASMONATE-INSENSITIVE1 encodes a MYC transcription factor essential to discriminate between different jasmonate-regulated defense responses in Arabidopsis. Plant Cell. 2004;16(7):1938–50. 10.1105/tpc.022319 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Lorenzo O, Piqueras R, Sanchez-Serrano JJ, Solano R. ETHYLENE RESPONSE FACTOR1 integrates signals from ethylene and jasmonate pathways in plant defense. Plant Cell. 2003;15(1):165–78. 10.1105/tpc.007468 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Solano R, Stepanova A, Chao Q, Ecker JR. Nuclear events in ethylene signaling: a transcriptional cascade mediated by ETHYLENE-INSENSITIVE3 and ETHYLENE-RESPONSE-FACTOR1. Genes Dev. 1998;12(23):3703–14. 10.1101/gad.12.23.3703 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Wen CK, Li W, Guo H. Regulatory components of ethylene signal transduction In: Wen CK, editors. Ethylene in plants: Springer; Netherlands; 2015. Pp. 73–92. 10.1007/978-94-017-9484-8_5 [DOI] [Google Scholar]
- 57.Kazan K. Diverse roles of jasmonates and ethylene in abiotic stress tolerance. Trends Plant Sci. 2015;20(4):219–29. 10.1016/j.tplants.2015.02.001 . [DOI] [PubMed] [Google Scholar]
- 58.Pieterse CM, Leon-Reyes A, Van der Ent S, Van Wees SC. Networking by small-molecule hormones in plant immunity. Nat Chem Biol. 2009;5(5):308–16. 10.1038/nchembio.164 . [DOI] [PubMed] [Google Scholar]
- 59.Pieterse CM, Van der Does D, Zamioudis C, Leon-Reyes A, Van Wees SC. Hormonal modulation of plant immunity. Annu Rev Cell Dev Biol. 2012;28:489–521. 10.1146/annurev-cellbio-092910-154055 . [DOI] [PubMed] [Google Scholar]
- 60.Pozo MJ, Van Der Ent S, Van Loon LC, Pieterse CM. Transcription factor MYC2 is involved in priming for enhanced defense during rhizobacteria-induced systemic resistance in Arabidopsis thaliana. New Phytol. 2008;180(2):511–23. 10.1111/j.1469-8137.2008.02578.x . [DOI] [PubMed] [Google Scholar]
- 61.Van der Ent S, Van Hulten M, Pozo MJ, Czechowski T, Udvardi MK, Pieterse CMJ, et al. Priming of plant innate immunity by rhizobacteria and beta-aminobutyric acid: differences and similarities in regulation. New Phytol. 2009;183(2):419–31. 10.1111/j.1469-8137.2009.02851.x . [DOI] [PubMed] [Google Scholar]
- 62.Pre M, Atallah M, Champion A, De Vos M, Pieterse CM, Memelink J. The AP2/ERF domain transcription factor ORA59 integrates jasmonic acid and ethylene signals in plant defense. Plant Physiol. 2008;147(3):1347–57. 10.1104/pp.108.117523 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Penninckx IA, Thomma BP, Buchala A, Métraux JP, Broekaert WF. Concomitant activation of jasmonate and ethylene response pathways is required for induction of a plant defensin gene in Arabidopsis. Plant Cell. 1998;10(12):2103–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Fujimoto SY, Ohta M, Usui A, Shinshi H, Ohme-Takagi M. Arabidopsis ethylene-responsive element binding factors act as transcriptional activators or repressors of GCC box-mediated gene expression. Plant Cell. 2000;12(3):393–404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Berrocal-Lobo M, Molina A, Solano R. Constitutive expression of ETHYLENE-RESPONSE-FACTOR1 in Arabidopsis confers resistance to several necrotrophic fungi. Plant J. 2002;29(1):23–32. 10.1046/j.1365-313x.2002.01191.x . [DOI] [PubMed] [Google Scholar]
- 66.Zhao Y, Dong W, Zhang N, Ai X, Wang M, Huang Z, et al. A wheat allene oxide cyclase gene enhances salinity tolerance via jasmonate signaling. Plant Physiol. 2013;164(2):1068–76. 10.1104/pp.113.227595 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Garcia-Abellan JO, Fernandez-Garcia N, Lopez-Berenguer C, Egea I, Flores FB, Angosto T, et al. The tomato res mutant which accumulates JA in roots in non-stressed conditions restores cell structure alterations under salinity. Physiol Plant. 2015;155(3):296–314. 10.1111/ppl.12320 . [DOI] [PubMed] [Google Scholar]
- 68.Hua W, Ye H, Yao R, Tao Z, Xiong L. OsJAZ9 acts as a transcriptional regulator in jasmonate signaling and modulates salt stress tolerance in rice. Plant Sci. 2014;232:1–12. 10.1016/j.plantsci.2014.12.010 . [DOI] [PubMed] [Google Scholar]
- 69.Cheng MC, Liao PM, Kuo WW, Lin TP. The Arabidopsis ETHYLENE RESPONSE FACTOR1 regulates abiotic stress-responsive gene expression by binding to different cis-acting elements in response to different stress signals. Plant Physiol. 2013;162(3):1566–82. 10.1104/pp.113.221911 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Peng J, Li Z, Wen X, Li W, Shi H, Yang L, et al. Salt-induced stabilization of EIN3/EIL1 confers salinity tolerance by deterring ROS accumulation in Arabidopsis. Plos Genet. 2014;10(10):e1004664–e. 10.1371/journal.pgen.1004664 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Mideros S, Nita M, Dorrance AE. Characterization of components of partial resistance, Rps2, and root resistance to Phytophthora sojae in Soybean. Phytopathology. 2007;97(5):655–62. 10.1094/PHYTO-97-5-0655 . [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(DOCX)
Sequence alignment was done by ClustalW. Black and gray backgrounds indicate identical and similar residues, respectively; dotted lines indicate gaps.
(TIF)
Electrophoresis pattern of PCR from genomic DNA (A) and cDNA (B) corresponding to GmCYP82A3 (upper panel) of wild-type (WT), empty vector (EV) transformants (EV-1, EV-2) and the six T2 transgenic lines expressing GmCYP82A3 (2–3, 3–3, 4–1, 10–1, 15–2, and 9–1). The NbEF1a (lower panel) was used as an internal control. M, DNA Marker DL2000 PLUS.
(TIF)
(A) Phenotypes of the N. benthamiana leaves from WT, EV and overexpression lines (2–3 and 4–1) inoculated with B. cinerea at 4 dpi. Bar = 5 mm. (B) Lesion area of inoculated leaves. Lesion diameters were measured at 4 dpi and then the lesion area was calculated. SD represented with the bars (Dunnett-t test: ** P<0.01).
(TIF)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.