

Abstract
APETALA2/Ethylene-Responsive Factor (AP2/ERF) gene family is plant specific transcription factor. It plays critical roles in development process, tolerance to biotic and abiotic stresses, and responses to plant hormones. However, limited data are available on the contributions of AP2/ERF gene family in barley (Hordeum vulgare L.). In the present study, 121 HvAP2/ERF genes in barley were identified by using bioinformatics methods. A total of 118 HvAP2/ERF (97.5%) genes were located on seven chromosomes. According to phylogenetic classification of AP2/ERF family in Arabidopsis, HvAP2/ERF proteins were divided into AP2 (APETALA2), RAV (Related to ABI3/VP), DREB (dehydration responsive element binding), ERF (ethylene responsive factors) and soloist sub families. The analysis of duplication events indicated that tandem repeat and segmental duplication contributed to the expansion of the AP2/ERF family in barley. HvDREB1s/2s genes displayed various expression patterns under abiotic stress and phytohormone. Taken together, the data generated in this study will be useful for genome-wide analysis to determine the precise role of the HvAP2/ERF gene during barley development, abiotic stress and phytohormone responses with the ultimate goal of improving crop production.
Introduction
The APETALA2/Ethylene-Responsive Factor (AP2/ERF) gene family is one of largest gene families, encoding plant-specific transcription factors. The AP2/ERF superfamily is defined by the AP2/ERF domain, which consists of about 60 to 70 amino acids and is involved in DNA binding. The AP2/ERF superfamily is divided into AP2 (APETALA2), RAV (Related to ABI3/VP), DREB (dehydration responsive element binding), ERF (ethylene responsive factors) and soloist sub families [1–4]. The AP2 subfamily members contain multiple AP2/ERF domains or lacking a conserved WLG motif within AP2/ERF domain, the RAV subfamily transcription factors include a single AP2/ERF domain and B3 domain, DREB and ERF subfamily possess only one AP2/ERF domain, the remaining genes are defined as soloist [1]. The AP2/ERF gene members play an important role in the regulation of plant development and tolerance to biotic and abiotic stresses [4–12]. With more extensive plant genome sequences, AP2/ERF gene family have been identified in various plants, such as Arabidopsis [3], rice [3], maize [4], sorghum [13], soybean [14], foxtail millet [15]. However, no research has been performed for the identification and characterization of the AP2/ERF family in barley.
Salinity and drought are two of the most serious abiotic stress factors. Plants have evolved on the molecular level to survive these environmental stresses. Plant hormone ABA also plays an important role in improving the tolerance of plants to drought and salinity [16–19]. The DREBs/CBFs (Dehydration Responsive Element Binding proteins/C-repeat CRT binding transcription factors), hereafter referred as DREBs is a major member of AP2/ERF gene family, which binds to DRE (A/GCCGAC) and/or the CRT (TGGCCGAC) core cis-acting sequences in promoters of drought and salt responsive genes and regulates stress-responsive genes [20]. The ectopic overexpression of DREB genes in plant systems resulted in improved salt stress tolerance as positive regulator [20–22]. Enhanced expression of OsDREB2A and OsDREB1F results in improved drought and salt stress tolerance in rice and Arabidopsis [22]. In rice, OsDREB1A and OsDREB1F were induced by cold stress. OsDREB1F was also induced by drought, salt and ABA treatment. Over-expression of OsDREB1A and OsDREB1F resulted in with higher tolerance to drought, high-salt stress in Arabidopsis [20–21]. In Arabidopsis, a cbf2 mutant was identified by using a reverse genetic approach, in which the CBF2/DREB1C gene was abnormal. cbf2 mutant showed increased tolerance to drought and salt stress. Expression analysis indicated that CBF2/DREB1C negatively regulated the expression of CBF1/DREB1B and CBF3/DREB1A [23]. Remarkably, DREB1/2 genes were induced under drought and salt stress, indicating cross-talk between them [17, 21, 24–26]. These results suggest that the functions of DREB1s and DREB2s genes in combination with ABA are conserved both in monocot and dicot plants, and play an important role in plant drought and salt stress responses [20].
Barley is considered as the most salt and drought tolerant among cereal crops but cultivars show considerable variation to different tolerances [27]. With growing world population, global food production should be meet the demands by improving abiotic tolerance of crops, and has become the focus for enhancing breeding efforts [27]. In the present study, we identified 121 HvAP2/ERF genes in barley by using bioinformatics methods and constructed a phylogenetic tree. Most of the HvAP2/ERF genes were localized to chromosomes using drawing tools and duplication events were also analyzed. The expression patterns of one hundred and seven AP2/ERF genes were detected using published RNA sequencing. Finally, the expression level of twenty one genes in response to drought, high-salt stress and exogenous ABA was performed by Quantitative real-time PCR. These results will be useful in further investigation of the AP2/ERF family in plants.
Materials and Methods
Sequence database searches
Multiple database searches were performed to collect all members of the barley HvAP2/ERF gene. Barley sequence data were sourced from the Morex assembly (http://webblast.ipk-gatersleben.de/barley/) [28], Gramene (http://ensembl.gramene.org/Hordeum_vulgare/Info/Index) and NCBI database (http://www.ncbi.nlm.nih.gov/). We used the BLAST programs (TBLASTN and BLASTN) which is available on the IPK barley genome database and NCBI barely EST database. We used the amino acid sequence of the AP2/ERF domain from Arabidopsis (Arabidopsis thaliana), rice (Oryza sativa L.) and maize (Zea mays L.) as a query sequences [3, 4]. To increase the extent of the database search results, we also performed the database searches using amino acid sequences of the AP2/ERF genes of some members of the Transcription Factor Database (http://planttfdb.cbi.pku.edu.cn/) [29]. Barley HvAP2/ERF candidate genes with expected (E) values less than 1.0 were retrieved and the non-redundant sequences were examined for the presence of conserved AP2/ERF domain using the domain analysis programs Pfam (Protein family: http://pfam.sanger.ac.uk/) [30], HMMSCAN (https://www.ebi.ac.uk/Tools/hmmer/search/hmmscan) and SMART (Simple Modular Architecture Research Tool: http://smart.embl-heidelberg.de/) with the default cutoff parameters [31]. The isoelectric points and protein molecular weights were obtained with the help of the proteomics and sequence analysis tools on the ExPASy proteomics server (http://expasy.org/) [32]. The gene names of HvDREB and HvERF was given according to the ascending order of AP2/ERF domain of analysis in DREB and ERF subfamily. The gene names of HvAP2 and HvRAV was given according to the ascending order of the phylogenetic tree in the AP2 and RAV subfamily.
Chromosomal location, gene structure, promoter and duplication events of HvAP2/ERF genes
The chromosomal locations were retrieved from the Gramene (http://ensembl.gramene.org/Hordeum_vulgare/Info/Index). All genes were mapped to the chromosomes with MapDraw software [33]. The exon/intron structures were constructed using GSDS (http://gsds.cbi.pku.edu.cn/) [34]. Maximum 2,000 bp promoter regions were examined to identify cis-regulatory element using PlantCARE (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/). Tandem duplication genes were identified manually if they were within 10 predicted genes or within 30 kb apart of each other and marked on the barley physical map [35]. Segmental duplications were identified by BLASTP ten predicted proteins upstream and downstream of each HvAP2/ERF [36].
Phylogenetic tree analysis
Full-length amino acid sequences of HvAP2/ERF genes identified in barley were aligned using the Clustal X 1.83 program with default pairwise and multiple alignment parameters. The phylogenetic tree was constructed based on this alignment result using the neighbor joining (NJ) method in MEGA version 6 with the following parameters [37]: Poisson correction, pairwise deletion, uniform rates and bootstrap (1000 replicates). Conserved motifs were investigated by multiple alignment analyses using MEME version 3.0 [38].
Comparative genomics analysis among barley, maize, rice, Brachypodium and foxtail millet
Comparative genomics analysis was performed according to Lata et al [15]. The protein sequences of HvAP2/ERF proteins were carried out BLASTP search against protein sequences of maize, rice, Brachypodium and foxtail millet (http://gramene.org/; www.phytozome.net), orthologous genes was also performed reciprocal BLASTP search to conform relationship among them. Cutoff with E-value≤1e-05 and at least 80% similarity were considered significant. The amino-acid sequence of paralogous and orthologous AP2/ERF proteins combined the corresponding CDS sequences among barley, maize, rice, Brachypodium and foxtail millet were aligned using Clustal W, and then analyzed by using PAL2NAL (http://www.bork.embl.de/pal2nal/) for calculating the synonymous (Ks) and non-synonymous (Ka) substitution rates [39].
Expression profiling analysis of the HvAP2/ERF genes
Gene expression data from eight tissues of the cultivar ‘Morex’ were obtained by making use of the barley genome database (http://apex.ipk-gatersleben.de/apex/f?p=284:10: 6281639160219::NO). Eight tissues of the cultivar ‘Morex’ earmarking stages of the barley life cycle, including 4-day embryos dissected from germinating grains, roots and shoot from the seedlings (10 cm shoot stage), young developing inflorescences (5 mm), developing inflorescences (1–1.5 cm), developing tillers at six leaf stage (the third internode), 5 and 15 days post-anthesis developing grain (DPA) (bracts removed), which were selected for deep RNA sequencing (RNA-seq) [28]. The expression patterns are presented as heat maps in green/yellow/red/ coding, which reflected the FPKM (Fragments Per Kilobase of transcript per Million mapped reads) with red indicating high expression level, yellow indicating middle expression level, and green indicating low expression level.
Plant materials and treatments
Barley Gairdner was a salt-sensitive variety and CM72 was a salt-tolerant variety [40–41]. Whole-plant responses to salinity, polyethylene glycol (PEG) and abscisic acid (ABA) were studied in glasshouse by using a hydroponic culture technique. Seeds of Gairdner and CM72 were sterilized with 5% sodium hypochlorite for 10 min and rinsed with distilled water, then germinated on wet filter paper at 20°Cfor 3 days. The germinated seeds were transferred into 60-well plastic containers (25 L) with aerated hydroponic solution similar to that used by Wu et al [42]. The pH of the hydroponic solution was adjusted to 6.8 using 1 M HCl as required. All solutions were renewed weekly. Plants were grown in a greenhouse, and a temperature of 20°C/day and 15°C/night. Three weeks old seedlings were exposed to 250 mM NaCl, 20% PEG and 100 μM ABA for 0 h, 1 h, 6 h, 12 h, 24 h and 48 h. After treatment, root and leaf were collected and immediately frozen in liquid nitrogen for RNA extraction with three biological replicates. For each replicate, ten plants of each genotype were used for RNA analysis.
Quantitative real-time PCR analysis of barley HvDREB1s/2s genes under salt, drought and phytohormone treatment
Total RNA of each sample was isolated using an RNA extraction kit (TRIzol reagent, Invitrogen, USA) and incubated with RNase-free DNase I (TaKaRa, Japan) for removing DNA contamination. Quality and yields of RNA were analyzed by agarose gels electrophoresis and NanoDrop 1000 Spectrophotometer V 3.7. First strand cDNA was generated from 2 μg total RNA with M-MLV reverse transcriptase (TaKaRa, Japan) by using random primers. Specific primers for quantitative real-time PCR analysis were listed in S1 Table. Reaction was carried out in 20 μl reaction system containing 10 mM Tris-HCl (pH 8.5), 50 mM KCl, 2 mM MgCl2, 0.4 μl DMSO, 200 mM dNTPs, specific PCR primers 10 pmol, Taq DNA polymerase 1 U, SYBR GREEN I fluorescence dye 0.5 μl. Quantitative real-time PCR was performed using a ViiA™ 7 Real-Time PCR System (Applied Biosystems, USA). The running protocol was as follows: 94°C for 3 min, followed by 40 cycles at 94°C for 30 s, 58°C for 30 s, 72°C for 30 s, and a final extension of 72°C for 5 min. ADP-ribosylation factor 1-like protein(ADP) was used as an internal control [43]. All reactions were run in triplicate. Ct values were determined by the ViiA™ 7 software with default settings. The relative expression levels of target genes were determined using 2-ΔΔCt method [44]. For each sample, PCR was performed with three biological replicates.
Results
Identification of the AP2/ERF family genes in barley genome
To identify the AP2/ERF genes in barley, BLAST searches of the barley databases were performed using the AP2/ERF domains of the Arabidopsis, rice and maize protein as a query sequences. A total of 121 HvAP2/ERF genes from the entire barley genome were identified as potential ones encoding AP2/ERF domain (S2 and S3 Tables). Among which 33 HvAP2/ERF genes (33/121, 27.3%) were found splice variants of primary transcripts. The number of alternate transcript ranged from 2 to 7 (S3 Table). Based upon the phylogenetic classification of AP2/ERF family in Arabidopsis, 19 HvAP2/ERF proteins were classified into AP2 subfamily (S4 Table), with containing one AP2/ERF domain without WLG motif, two (HvAP2-1/3/5/6/7/8/9/11/12/14/17) or three (HvAP2-18) AP2/ERF domain, the remaining members only contained (S4 Table). Six of the proteins, containing both AP2 and B3 domains, were classified into the RAV subfamily (S4 Table). Of the remaining 95 proteins that have only one AP2/ERF domain, 41 members were classified into the DREB subfamily and 54 members were placed in the ERF subfamily based on the conserved motif (S4 Table). A soloist gene including an AP2/ERF like domain sequence was assigned due to a lack of homology with the other HvAP2/ERF genes (S4 Table). All the identified HvAP2/ERF genes encode proteins ranging from 123 (HvERF3.5) to 659 (HvAP2-1) amino acids along with a protein mass from 13.38 kD to 69.73 kD and protein pI ranging from 3.98 (HvDREB2.11) to 11.87 (HvERF6.5) (S3 Table).
Table 1 shows the comparison of the AP2/ERF genes from Arabidopsis, maize, rice, foxtail millet and barley. In barley, 95 HvAP2/ERF genes were classified into the DREB/ERF subfamily. The number of the genes in Arabidopsis, maize, rice and foxtail millet were 122, 163, 131 and 138, respectively [3–4, 15]. In AP2 subfamily barley, Arabidopsis, maize, rice and foxtail millet had 19, 18, 44, 36, and 28 HvAP2/ERF genes, respectively. Six barley HvAP2/ERF genes were predicted to encode proteins with an AP2/ERF domains and B3 domains, which were classified into the RAV subfamily. Similar numbers of genes were also found in Arabidopsis, maize, rice and foxtail millet in RAV subfamily (Table 1).
Table 1. Summary of the AP2/ERF superfamily in Arabidopsis, rice, maize, barley and foxtail millet.
| Classification | Group | Barleya | Arabidopsisb | Riceb | Maizec | Foxtail Milletd |
|---|---|---|---|---|---|---|
| AP2 sub family | 19 | 18 | 36 | 44 | 28 | |
| Multiple AP2/ERF domain | 13 | 14 | 22 | |||
| Single AP2/ERF domain (without WLG motif) | 6 | 4 | 22 | |||
| DREB/ERF sub family | 95 | 122 | 131 | 163 | 138 | |
| DREB sub family | 41 | 57 | 65 | 48 | ||
| ERF sub family | 54 | 65 | 98 | 90 | ||
| RAV | 6 | 6 | 7 | 3 | 5 | |
| Soloist | 1 | 1 | 0 | 0 | 0 | |
| Total | 121 | 147 | 164 | 210 | 171 | |
a) Some members of HvAP2/ERF genes were derived from the Transcription Factor Database (http://planttfdb.cbi.pku.edu.cn/) [29].
b) Nakano et al., 2006 [3];
c) Liu et al., 2013 [4].
d) Lata et al., 2014[15]
Chromosomal location and structure of HvAP2/ERF genes
One hundred and eighteen HvAP2/ERF genes (97.5%, 118/121) were located on 7 chromosomes; three genes (HvERF2.15, HvERF2.17 and HvERF4.4) were not found the precise chromosome and physical locations on barley genome (Fig 1, S3 Table). Chromosome 6 had the largest number of HvAP2/ERF genes (29 genes) while only eight genes were located on 4H. Other chromosomes contained 13–22 HvAP2/ERF genes. Gene structural analyses showed that most HvAP2/ERF genes contain one exon (87/121, 71.9%), the remaining HvAP2/ERF genes shared 2 (19 genes), 4 (3 genes), 5 (1 gene), 6 (1 gene), 7 (4 genes), 8 (2 genes), 9 (3 genes) and 10 exons (1 gene) (S1 Fig). Duplication events analysis indicated that 22 (22/121, 18.2%) HvAP2/ERF genes were tandem repeated (S5 Table) and 6 (6/121, 5.0%) HvAP2/ERF genes were segmentally duplicated (S6 Table). The tandem duplicated genes contained six clusters, the largest cluster located on chromosome 5, which including six genes, chromosome 2 and 7 contained one gene pairs, respectively, chromosome 6 including 3 clusters. Remarkably, three HvAP2/ERF gene pairs displayed segmentally duplicated, including HvAP2-5 and HvAP2-8, HvRAV-1 and HvRAV-2, HvERF3.6 and HvERF3.9 gene pairs.
Fig 1. The chromosome location of the HvAP2/ERF genes in barley.

The red lines represent tandemly duplicated gene pairs.
Phylogenetic tree of the HvAP2/ERF proteins in barley
The AP2/ERF family genes are plant-specific transcription factors. Based on alignment of the HvAP2/ERF domain of barley, 121 barley HvAP2/ERF genes were classified into the DREB, ERF, AP2 and RAV subfamilies, and one soloist (Fig 2, S3 Table). Phylogenetic trees of the AP2, RAV, DREB and ERF subfamilies in barley were constructed (Fig 2). A total of 41 DREB subfamily genes distributed into the A1, A2, A4, A5 and A6 groups in barley, including 11, 12, 6, 5 and 7 genes, respectively (Fig 2, S2 Fig). Additionally, 54 genes belonging to the ERF subfamily in barley distributed into the B1-B6 groups, including 6, 18, 9, 10, 6 and 5 genes, respectively (Fig 2, S2 Fig). In addition, 19 genes were classified into the AP2 subfamily and six genes into the RAV subfamily (Fig 2).
Fig 2. The phylogenetic analysis of HvAP2/ERF genes in barley.

Conserved motifs of the HvAP2/ERF proteins
Conserved motifs can provide evidence for further classification as it is likely that identical motifs exhibit similar functions. Five conserved motif (Motif 1–5) were analyzed by using MEME software to identify conserved motifs among proteins in the families and subfamilies. The MEME motif analysis revealed that different HvAP2/ERF proteins had different conserved motifs (Fig 3, S3 Fig). All of the HvAP2/ERF genes had the full AP2/ERF domain and Motif 1 (S3 Fig, S4 Table). In addition, motif 2, 3 and 4 were detected in most of the HvAP2/ERF genes (S2 Fig). Remarkably, motif 5 only observed in HvAP2-16 and HvERF2.1–2.10 (S2 Fig). Two conserved amino acids in the AP2/ERF domains differ between DREB and ERF [45]. In DREB/ERF subfamily, the 14th valine (V14) and the 19th glutamic acid (E19) are conserved in AP2/ERF domain of HvDREB protein, whereas alanine (A) and aspartic acid (D) are conserved in the corresponding positions of the HvERF proteins (S3 Fig).
Fig 3. Conserved domains of HvAP2/ERF proteins in barley.

Orthologous relationships of HvAP2/ERF genes among grass species
To investigate the relationship of HvAP2/ERF genes among grass species, comparative genomics was carried out between barley, maize, rice, Brachypodium and foxtail millet (S7–S10 Tables). The largest number of orthology of genes was 18 (18/121, 15%) between barley and Brachypodium (S10 Table), followed by rice (12/121, 10%) (S8 Table), foxtail millet (9/121, 7%) (S10 Table) and maize (6/121; 5%) (S9 Table). For HvAP2-10, barley shared a greater orthology with maize (81%), rice (83%), Brachypodium (86%) and foxtail millet (88%), suggesting that HvAP2-10 shared the similar functions in different species.
Expression pattern analysis of barley HvAP2/ERF genes under different development stage
Expression profiling analysis of the gene family can provide important clues regarding their functions [46]. One hundred and seven HvAP2/ERF genes were detected by RNA sequencing, 14 HvAP2/ERF genes had no transcript in any tissue (Fig 4). The transcripts of 49 HvAP2/ERF genes (49/107, 45.8%) were detected in four days embryo after germinating. 40 (40/107, 37.4%) and 68 (68/107, 63.6%) HvAP2/ERF genes were detected in root and shoot from seedling, respectively. 66 (66/107, 61.7%) HvAP2/ERF genes were found expressed in developing tillers (six-leaf stage). In addition, 45 (45/107, 42.1%) and 59 (59/107, 55.1%) HvAP2/ERF genes were expressed in young fluoresce (5 mm and 1–1.5 cm length) and developing grains (5 and 15 DPA), among which HvERF2.11 displayed extremely higher expressed in 15 DPA than 5 DPA developing grains. Remarkably, HvERF2.3 and HvERF2.4 displayed higher expression level in developing tillers than other tissues. Indicating that expression analysis could be contributed to functional analysis of HvAP2/ERF genes in barley.
Fig 4. The expression profile of HvAP2/ERF genes in eight tissues of barley.

The expression patterns are presented as heat maps in green/yellow/red/ coding, which reflected the FPKM with red indicating high expression level, yellow indicating middle expression level, and green indicating low expression level.
Expression analysis of HvDREB1s/2s genes under abiotic stresses and phytohormone treatments
Plant growth is affected by various abiotic stresses, such as drought, high salinity, and low temperature [21]. Moreover, many HvDREB1s/2s genes are induced under stress conditions and phytohormone [4, 11, 17, 21–22, 26]. As shown in Fig 5, the quantitative real-time PCR was performed, including 11 HvDREB1s genes and 10 HvDREB2s genes. Most of HvDREB1s and HvDREB2s displayed similar expression patterns between Gairdner and CM72 in root and leaf tissue. Under salt stress, HvDREB1.11 was up-regulated in Gairdner and down-regulated in CM72 in root tissue, the fold change value ranged from 0.41 to 1.00 and 0.79 to 1.35 in Gairdner and CM72, respectively (S11 Table). However, HvDREB2.6 showed the opposite expression pattern. HvDREB1.3 and HvDREB2.8 were up-regulated in CM72 but down-regulated in Gairdner in leaf tissue after 12 h treatment. In contrast, HvDREB1.8, HvDREB1.12, and HvDREB2.10 were down-regulated in CM72 and up-regulated in Gairdner in leaf tissue (Fig 5A). Remarkably, HvDREB2.8 exhibiting more than a 1-fold change value in Gairdner and CM72 under high salt stress in root tissue (S11 Table). Under drought stress, HvDREB1.8 showed higher expression in CM72 than Gairdner, while HvDREB1.9 showed less expression in CM72 than Gairdner in root tissue. Both genes displayed down-regulated after early treatment (1 h time point) in leaf tissue (Fig 5B). HvDREB2.8 displayed up-regulated expression in both root and leaf tissue in both varieties with the expression level in leaf being higher than that in root tissue (Fig 5B), the average fold change value was 1.72 and 3.00 in root and leaf tissue, respectively (S11 Table).
Fig 5. The expression analysis of HvDREB1s and HvDREB2s genes in response to salinity, dehydration and Abscisic acid (ABA).
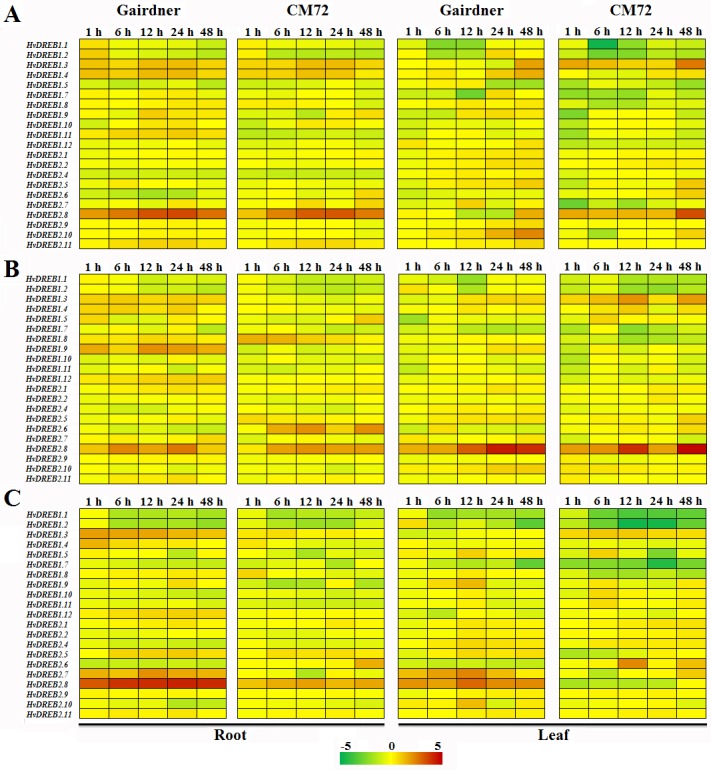
A, B, and C represent heat maps showing differential gene expression patterns in response to salinity, dehydration and ABA at 0 h, 1 h, 6 h, 12 h, 24 h and 48 h time point.
The plant hormone ABA has important roles in improving the tolerance of plants to drought and salinity stresses [16]. The Quantitative real-time PCR was also carried out to analyze the expression level of selected 21 HvDREB1s and HvDREB2s genes after ABA treatment (Fig 5C). The heat maps indicated that HvDREB1.3 exhibited higher expressed in Gairdner than in CM72 in root tissue with the average fold change value of 1.61 and 0.37, respectively (S11 Table). On the contrary, HvDREB1.3 was up-regulated expressed in CM72 and down-regulated expressed in Gairdner in leaf tissue. The expression level of HvDREB2.8 was dramatically increased in Gairdner with 3.99-fold change value than CM72 with 1.56-fold change value in both tissues, however, HvDREB2.6 displayed the opposite expression pattern (S11 Table).
Promoter analysis is a vital method to identify regulatory networks between environment stimulation and gene expression. Therefore, a cis-element scan was carried out to identify the potential regulatory elements response to abiotic stress and hormones. A total of 82 cis-regulatory elements were observed at least in one HvDREB1s/2s genes (S12 Table). Several cis-regulatory elements were involved in abiotic stress and phytohormone response. For example, ABRE (abscisic acid responsiveness element) and DRE (Dehydration-responsive element) involved in 18 and 1 HvDREB1s/2s genes, respectively, both of them were identified as cis-regulatory elements that participated in ABA-dependent and ABA-independent response to dehydration-inducible genes [16] (S12 Table). In addition, GARE-motif (gibberellin-responsive element), AuxRR-core (cis-acting regulatory element involved in auxin responsiveness) and MBS (MYB binding site involved in drought-inducibility) were also detected in promoter of regions (S12 Table).
Discussions
Characterization of the barely AP2/ERF superfamily
AP2/ERF superfamily is one of the largest groups of transcription factor family in plants, which also plays an important role in the transcriptional regulation involving in complicated developmental processes, biotic and abiotic stress, including seed germination, flower development and leaf senescence, fruit ripening, and responses to salt, drought, low temperature and pathogen attack [10, 12, 20, 47–51]. Based on the sequencing of plant genome, the AP2/ERF gene family was widely analyzed in plants [3–4, 13–15, 52–54]. However, there is still little information about barley HvAP2/ERF genes. To further investigate the AP2/ERF family in barley, 121 HvAP2/ERF genes were identified from 79379 (high-confidence and low-confidence genes) annotated genes and genome DNA database [28]. Each of them has notable features with at least one conserved AP2/ERF domain. Remarkably, the number of HvAP2/ERF gene in barley was less than Arabidopsis (147 genes), maize (210 genes) and rice (164 genes) [3, 4]. In addition, the numbers of some subfamilies were similar. For example, the numbers of AP2 subfamily in barley and Arabidopsis were 19 and 18, respectively, being half of the number in rice and maize. The numbers of RAV subfamily in barley, Arabidopsis, rice and foxtail millet were 6, 6, 7 and 5, respectively. On the contrary, the numbers of other subfamilies were significantly different. For example, 95 genes were identified in the DREB/ERF subfamily in barley. The numbers of genes were 122, 131, 163 and 138 in Arabidopsis, rice, maize and foxtail millet, respectively. Recently, it has been reported that segmental duplication events and tandem duplication events in plants were contributed to the expansion of the AP2/ERF family, suggesting that pressure was the predominant force acting on the evolution of the AP2/ERF family [3–4, 15]. In the present study, the Ka/Ks ratio for tandem duplicated and segmental duplication gene-pairs in barley HvAP2/ERF genes less than 1.0 expect HvERF2.8 and HvERF2.9 gene pairs, indicating that HvAP2/ERF genes undergoing purifying selection in gene expansion (S5 and S6 Tables). The smaller number of genes in AP2/ERF family in barley suggested that there may be more other HvAP2/ERF genes existing in the unknown genomic regions or chromosome duplication events was restricted in barley evolutionarily expansion.
Conserved motifs in transcription factors play an important role in gene function [1]. A total of fifty conserved motifs outside of the AP2/ERF domain were detected in Arabidopsis [3]. In the present study, we analyzed five motifs of AP2/ERF proteins, motif 1 (partial of AP2/ERF domain) was observed in all gene members, other motifs were outside the AP2/ERF domain. Previous studies revealed that DREB and ERF subfamily contained conserved WLG motif in AP2/ERF domain [3–4]. In the present study, WLG motif was highly conserved in DREB and ERF subfamily, as well as in RAV and Solosist subfamily (Table 1, Fig 2). Sequence alignment of ERF subfamily has revealed 14th alanine and the 19th aspartic acid of AP2/ERF domain are conserved, whereas valine and glutamic acid residues are conserved at the corresponding positions of DREB subfamily [3]. The two conserved amino acids are located on the β-sheet in the AP2/ERF domain, which is important for binding to the target DNA sequences [1]. The AP2/ERF domain of DREB and ERF sub family were well-conserved among Arabidopsis, rice and maize [3–4]. Remarkably, all DREB and ERF subgroups were completely conserved in Val-14 and Ala-14 acid residues, respectively. However, in the present study, seventeen DREB gene members were conserved in Glu-19, meanwhile fifty ERF subfamily gene members were completely conserved in Asp-19, only HvERF1.6/2.12 was not conserved in Asp-19 amino acid residues. These conserved amino acid residues probably indicate crucial roles for DREB/ERF sub family genes involved in different forms of physical interaction with DNA [1].
Expression analysis indicated HvAP2/ERF genes may play important roles during plant growth, abiotic stress and hormone response
Tissue-specific expression data at a given developmental stage is useful for identifying genes involved in defining precise nature of individual tissues. In the present study, the expression pattern of one hundred and seven HvAP2/ERF genes were detected by RNA sequencing, it was contribute to investigate the function of the HvAP2/ERF genes in barley. Remarkably, HvAP2-12 gene also named as Cleistogamy 1 (Cly1)/HvAPETALA2 (HvAP2), which was an ortholog of AP2 (AT4G36920.1), TOE3 (AT5G67180.1) and rice AP2-like gene Os04g0649100 [55–56]. In situ RNA hybridization indicated that the Cly1 transcript was detected in the lodicule up to the stamen primordium stage [55]. Another gene HvDREB2.2 also named Nud (Nudum), which control covered/naked caryopsis in barley and was expressed in the caryopsis at two weeks after anthesis rather than in hulls or leaves [57]. In the present study, HvAP2-12 displayed relative high expression level in developing tillers at six leaf stage, the transcript of HvDREB2.2 gene was only detected in developing grains (5 DPA). Therefore, more sophisticated specificity expression analysis is helpful to parse the function of HvAP2/ERF genes.
Plants were involved in adverse environmental stresses in their natural environments. On the molecular levels, they have evolved a wide range of mechanisms to cope with them. In plants, the genes respond to drought and high-salt stress involved in two ABA dependent and two ABA-independent signal transduction pathways [16–17]. A phylogenetic tree of the HvDREB1s and HvDREB2s proteins and their orthologs from Arabidopsis, maize and rice was constructed (S4 Fig). HvDREB2.9 was an orthology gene of OsDREB2A with improving drought and salt stress tolerance in rice and Arabidopsis, which had no obvious difference after drought, high-salt and ABA treatment although promoter contained multiple ABREs in barley. The expression level of HvDREB1.1 and HvDREB1.2 were decreased after 6 h, on the contrary, OsDREB1F as an orthology gene of HvDREB1.1 and HvDREB1.2 was increased after drought, high-salt and ABA treatment [20]. Remarkably, some HvDREB1s and HvDREB2s may be displayed different functions in plants. HvDREB1.8 was orthology gene of OsDREB1A which was induced by drought, high-salt stress and also ABA treatment in root, the expression level of HvDREB1.8 was higher in CM72 than Gairdner. In addition, HvDREB1.3/1.4/2.8 was increased under drought and salt stress, as well as HvDREB2.8 response to ABA treatment. Further analysis revealed that HvDREB1.3 and HvDREB1.4 were identified as homologous to OsDREB1C genes in rice whereas OsDREB1C showed constitutive expression [21]. HvDREB2.8 was a homologous of maize ZmDREB2.7 which enhanced tolerance to drought stress by overexpressing ZmDREB2.7 in transgenic Arabidopsis [4] (S4 Fig). However, HvDREB2.8 displayed higher expression level in Gairdner than CM72 except in leaf under salt stress. An ABRE functions as a cis-acting DNA element involved in ABA-regulated gene expression, which was observed in promoter regions of dehydration-inducible genes. In the present study, promoter analysis revealed that HvDREB1.3/1.4/1.8/2.8 contained multiple ABREs, suggesting that these genes are involved in ABA-dependent response under drought and high-salt conditions (S12 Table). Therefore, gene expression analysis of HvDREB1s and HvDREB2s should help us to investigation the molecular mechanisms of environment adaptability of plant under abiotic stress and hormone response, the function of differentially expressed genes between Gairdner and CM72 should be further investigated in relation to abiotic stress and hormone response.
Supporting Information
(TIF)
(TIF)
(TIF)
The phylogenetic tree was constructed based on the sequence alignments of seventy-four full-length DREB 1 and DREB2 genes from four species. The gene ID and names are illustrated in red for barley; black for rice; blue for Arabidopsis; and green for maize. The gene names were used in the present study according to published data [1, 4, 46, 58]. Bootstrap values from 1,000 replicates were indicated at each node and the scale represents branch lengths.
(TIF)
(XLS)
(DOCX)
(XLS)
(XLS)
(XLS)
(XLS)
(XLSX)
(XLSX)
(XLSX)
(XLSX)
(XLSX)
(XLSX)
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This research was supported from the National Natural Science Foundation of China (31401370 and 31571648), National Barley and Highland Barley Industrial Technology Specially Constructive Foundation of China (CARS-05), Jiangsu Agriculture Science and Technology Innovation Fund (CX(14)2123) and a Project Funded by the Priority Academic Program Development of Jiangsu Higher Education Institutions. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Sakuma Y, Liu Q, Dubouzet JG, Abe H, Shinozaki K, Yamaguchi-Shinozaki K. DNA-binding specificity of the ERF/AP2 domain of Arabidopsis DREBs, transcription factors involved in dehydration- and cold-inducible gene expression. Bioch Bioph Res Co 2002; 290: 998–1009. 10.1006/bbrc.2001.6299 [DOI] [PubMed] [Google Scholar]
- 2.Wessler SR. Homing into the origin of the AP2 DNA binding domain. Trends Plant Sci 2005; 10: 54–56. 10.1016/j.tplants.2004.12.007 [DOI] [PubMed] [Google Scholar]
- 3.Nakano T, Suzuki K, Fujimura T, Shinshi H. Genome-wide analysis of the ERF gene family in Arabidopsis and rice. Plant Physiol 2006; 140:411–432. 10.1104/pp.105.073783 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Liu S, Wang X, Wang H, Xin H, Yang X, Yan J, et al. Genome-Wide analysis of ZmDREB genes and their association with natural variation in drought tolerance at seedling stage of Zea mays L. Plos Genetics 2013; 9(9):119–129. 10.1371/journal.pgen.1003790 PMC3784558 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Moose SP, Sisco PH. Glossy15, an APETALA2-like gene from maize that regulates leaf epidermal cell identity. Genes Dev 1996; 10: 3018–3027. 10.1101/gad.10.23.3018 [DOI] [PubMed] [Google Scholar]
- 6.Aukerman MJ, Sakai H. Regulation of flowering time and floral organ identity by a microRNA and its APETALA2-like target genes. Plant Cell 2003; 15: 2730–2741. 10.1105/tpc.016238 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Jofuku KD, Omidyar PK, Gee Z, Okamuro JK. Control of seed mass and seed yield by the floral homeotic gene APETALA2. Proc Natl Acad Sci USA. 2005; 102: 3117–3122. 10.1073/pnas.0409893102 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Ohto MA, Floyd SK, Fischer RL, Goldberg RB, Harada JJ. Effects of APETALA2 on embryo, endosperm, and seed coat development determine seed size in Arabidopsis. Sex Plant Reprod 2009; 22: 277–289. 10.1007/s00497-009-0116-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Lata C, Prasad M. Role of DREBs in regulation of abiotic stress responses in plants. Journal of Experimental Botany 2011; 62(14):4731–4748. 10.1093/jxb/err210 [DOI] [PubMed] [Google Scholar]
- 10.Schmidt R, Mieulet D, Hubberten HM, Obata T, Hoefgen R, Fernie AR, et al. SALT-RESPONSIVE ERF1 regulates reactive oxygen species-dependent signaling during the initial response to salt stress in rice. Plant Cell 2013; 25(6):2115–2131. 10.1105/tpc.113.113068 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Jiang QY, Hu Z, Zhang H, Ma YZ. Overexpression of GmDREB1 improves salt tolerance in transgenic wheat and leaf protein response to high salinity. Crop Journal 2014; 2:120–131. 10.1016/j.cj.2014.02.003 [DOI] [Google Scholar]
- 12.Zhu XL, Qi L, Liu X, Cai S, Xu H, Huang R, et al. The wheat ethylene response factor transcription factor pathogen-induced ERF1 mediates host responses to both the necrotrophic pathogen Rhizoctonia cerealis and freezing stresses. Plant Physiology 2014; 164(3):1499–1514. 10.1104/pp.113.229575 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Yan HW, Hong L, Zhou YQ, Jiang HY, Zhu SW, Fan J, et al. A genome-wide analysis of the ERF gene family in sorghum. Genetics & Molecular Research 2013; 12(2):2038–2055. [DOI] [PubMed] [Google Scholar]
- 14.Zhang G, Ming C, Chen X, Xu ZS, Guan S, Li LC, et al. Phylogeny, gene structures, and expression patterns of the ERF gene family in soybean (Glycine max L.). Journal of Experimental Botany 2008; 59(15): 4095–4107. 10.1093/jxb/ern248 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Lata C, Mishra AK, Muthamilarasan M, Bonthala VS, Khan Y, Prasad M. Genome-wide investigation and expression profiling of AP2/ERF transcription factor superfamily in foxtail millet (Setaria italica L.). PLoS One 2014; 9(11): e113092 10.1371/journal.pone.0113092 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Shinozaki K, Yamaguchi-Shinozaki K. Gene expression and signal transduction in water-stress response. Plant physiology 1997; 115(2): 327 10.1104/pp.115.2.327 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Nakashima K, Shinwari ZK, Sakuma Y, Seki M, Miura S, Shinozaki K, et al. Organization and expression of two Arabidopsis DREB2 genes encoding DRE-binding proteins involved in dehydration- and high-salinity-responsive gene expression. Plant Molecular Biology 2000; 42(4):657–665. 10.1023/A:1006321900483 [DOI] [PubMed] [Google Scholar]
- 18.Zhu JK. Salt and drought stress signal transduction in plants. Annu Rev Plant Biol 2002; 53:247–273. 10.1146/annurev.arplant.53.091401.143329 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Golldack D, Li C, Mohan H, Probst N. Tolerance to drought and salt stress in plants: unraveling the signaling networks. Front Plant Sci 2014; 5:151 10.3389/fpls.2014.00151 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Wang Q, Guan Y, Wu Y, Chen H, Chen F, Chu C. Overexpression of a rice OsDREB1F gene increases salt, drought, and low temperature tolerance in both Arabidopsis and rice. Plant Molecular Biology 2008; 67,589–602. 10.1007/s11103-008-9340-6 [DOI] [PubMed] [Google Scholar]
- 21.Dubouzet JG, Sakuma Y, Ito Y, Kasuga M, Dubouzet EG, Miura S, et al. OsDREB genes in rice, Oryza sativa L., encode transcription activators that function in drought-, high-salt- and cold-responsive gene expression. Plant Journal for Cell & Molecular Biology 2003; 33(4):751–763. 10.1046/j.1365-313X.2003.01661.x [DOI] [PubMed] [Google Scholar]
- 22.Mallikarjuna G, Mallikarjuna K, Reddy MK, Kaul T. Expression of OsDREB2A transcription factor confers enhanced dehydration and salt stress tolerance in rice (Oryza sativa L.). Biotechnology Letters, 2011; 33(8):1689–1697. [DOI] [PubMed] [Google Scholar]
- 23.Novillo F, Alonso JM, Ecker JR, Salinas J. CBF2/DREB1C is a negative regulator of CBF1/DREB1B and CBF3/DREB1A expression and plays a central role in stress tolerance in Arabidopsis. Proc Natl Acad Sci USA 2004; 101:3985–3990. 10.1073/pnas.0303029101 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Liu L, Zhu K, Yang Y, Wu J, Chen F, Yu D. Molecular cloning, expression profiling and trans-activation property studies of a DREB2-like gene from chrysanthemum (Dendranthema vestitum). Journal of Plant Research 2008; 121:215–226. 10.1007/s10265-007-0140-x [DOI] [PubMed] [Google Scholar]
- 25.Chen J, Xia X, Yin W. Expression profiling and functional characterization of a DREB2-type gene from Populus euphratica. Biochemical and Biophysical Research Communications 2009; 378:483–487. 10.1016/j.bbrc.2008.11.071 [DOI] [PubMed] [Google Scholar]
- 26.Gupta K, Agarwal PK, Reddy MK, Jha B. SbDREB2A, an A-2 type DREB transcription factor from extreme halophyte Salicornia brachiata confers abiotic stress tolerance in Escherichia coli. Plant Cell Reports 2010; 29:1131–1137. 10.1007/s00299-010-0896-7 [DOI] [PubMed] [Google Scholar]
- 27.Witzel K, Weidner A, Surabhi GK, Börner A, Mock HP, et al. Salt stress-induced alterations in the root proteome of barley genotypes with contrasting response towards salinity. Journal of Experimental Botany 2009; 60(12):3545–3557. 10.1093/jxb/erp198 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.The International Barley Genome Sequencing Consortium. A physical, genetic and functional sequence assembly of the barley genome. Nature 2012; 491:711–716. 10.1038/nature11543 [DOI] [PubMed] [Google Scholar]
- 29.Jin JP, Zhang H, Kong L, Gao G and Luo JC. PlantTFDB 3.0: a portal for the functional and evolutionary study of plant transcription factors. Nucleic Acids Research 2014; 42(D1):D1182–D1187. 10.1093/nar/gkt1016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Punta M, Coggill PC, Eberhardt RY, Mistry J, Tate J, Boursnell C, et al. The Pfam protein families database. Nucleic Acids Res 2012; 40, D290–301. 10.1093/nar/gkr1065 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Letunic I, Doerks T, Bork P. SMART 7: recent updates to the protein domain annotation resource. Nucleic acids research 2012; 40:D302–305. 10.1093/nar/gkr931 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Artimo P, Jonnalagedda M, Arnold K, Baratin D, Csardi G, de Castro E, et al. ExPASy: SIB bioinformatics resource portal. Nucleic Acids Res 2012; 40:W597–603. 10.1093/nar/gks400 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Liu RH, Meng JL. MapDraw: a microsoft excel macro for drawing genetic linkage maps based on given genetic linkage data. Yi Chuan 2003; 25:317–321. 10.3321/j.issn:0253-9772.2003.03.019 [DOI] [PubMed] [Google Scholar]
- 34.Hu B, Jin JP, Guo AY, Zhang H, Luo JC, Gao G. GSDS 2.0: an upgraded gene feature visualization server. Bioinformatics 2015; 31(8):1296–1297. 10.1093/bioinformatics/btu817 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Shiu SH, Bleecker AB. Expansion of the receptor-like kinase/Pelle gene family and receptor-like proteins in Arabidopsis. Plant Physiology 2003; 132(2): 530–543. 10.1104/pp.103.021964 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Maher C, Stein L, Ware D. Evolution of Arabidopsis microRNA families through duplication events. Genome research 2006; 16(4): 510–519. 10.1101/gr.4680506 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Molecular Biology and Evolution 2013; 30, 2725–2729. 10.1093/molbev/mst197 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Bailey T L, Elkan C. 1994. Fitting a mixture model by expectation maximization to discover motifs in biopolymers. InRAltman, DBrutlog P Karp, R Lathrop, D Searls, eds, Proceedings of the Second International Conference on Intelligent Systems for Molecular Biology. American Association for Artificial Intelligence Press, Menlo Park, CA, pp 28–36. [PubMed]
- 39.Suyama M, Torrents D, Bork P. PAL2NAL: robust conversion of protein sequence alignments into the corresponding codon alignments. Nucleic Acids Res 2006; 34: W609–W612. 10.1093/nar/gkl315 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Chen ZH, Newman I, Zhou MX, Mendham N, Zhang G, Shabala S. Screening plants for salt tolerance by measuring K+ flux: a case study for barley. Plant Cell & Environment 2005; 28(10):1230–1246. 10.1111/j.1365-3040.2005.01364.x [DOI] [Google Scholar]
- 41.Chen ZH, Cuin TA, Zhou MX, Twomey A, Naidu BP, Shabala S. Compatible solute accumulation and stress-mitigating effects in barley genotypes contrasting in their salt tolerance. Journal of Experimental Botany 2007; 58(15–16):4245–4255. 10.1093/jxb/erm284 [DOI] [PubMed] [Google Scholar]
- 42.Wu DZ, Qiu L, Xu L, Ye LZ, Chen MX, Sun DF, et al. Genetic variation of HvCBF genes and their association with salinity tolerance in Tibetan annual wild barley. Plos One 2011; 6: e22938 10.1371/journal.pone.0022938 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Ferdous J, Li Y, Reid N, Langridge P, Shi BJ, Tricker PJ. Identification of reference genes for quantitative expression analysis of MicroRNAs and mRNAs in barley under various stress conditions. Plos One 2015; 10(3). 10.1371/journal.pone.0118503 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real time quantitative PCR and the 2ΔΔCt method. Methods 2001; 25: 402–408. 10.1006/meth.2001.1262 [DOI] [PubMed] [Google Scholar]
- 45.Liu Q, Kasuga M, Sakuma Y, Abe H, Miura S, Yamaguchi S, hinozaki K, Shinozaki K.Two transcription factors, DREB1 and DREB2, with an EREBP/AP2 DNA binding domain separate two cellular signal transduction pathways in drought- and low-temperature-responsive gene expression, respectively, in Arabidopsis. Plant Cell 1998; 10, 1391–1406. 10.1105/tpc.10.8.1391 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Mao DH, Chen CY. Colinearity and similar expression pattern of rice DREB1s reveal their functional conservation in the cold-responsive pathway. Plos One 2012; 7(10):e47275–e47275. 10.1371/journal.pone.0047275 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Klucher KM, Chow H, Reiser L, Fischer RL. The AINTEGUMENTA gene of Arabidopsis required for ovule and female gametophyte development is related to the floral homeotic gene APETALA2. Plant Cell 1996; 8(2):137–153. 10.1105/tpc.8.2.137 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Wang A, Tan D, Takahashi A, Li TZ, Harada T. MdERFs, two ethylene-response factors involved in apple fruit ripening. J Exp Bot 2007; 58:3743–3748. 10.1093/jxb/erm224 [DOI] [PubMed] [Google Scholar]
- 49.Hattori Y, Nagai K, Furukawa S, Song XJ, Kawano R, Sakakibara H. The ethylene response factors SNORKEL1 and SNORKEL2 allow rice to adapt to deep water. Nature 2009; 460(7258): 1026–1030. 10.1038/nature08258 [DOI] [PubMed] [Google Scholar]
- 50.Woo H R, Kim J H, Kim J, Kim J, Lee U, Song IJ, et al. The RAV1 transcription factor positively regulates leaf senescence in Arabidopsis. Journal of Experimental Botany 2010; 61(14):3947–3957. 10.1093/jxb/erq206 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Yang CY, Hsu FC, Li JP, Wang NN, Shih MC. The AP2/ERF transcription factor AtERF73/HRE1 modulates ethylene responses during hypoxia in Arabidopsis. Plant Physiology 2011; 156(1):202–212. 10.1104/pp.111.172486 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Thamilarasan SK, Park JI, Jung HJ, Nou IS. Genome-wide analysis of the distribution of AP2/ERF transcription factors reveals duplication and CBFs genes elucidate their potential function in Brassica oleracea. BMC Genomics 2014; 15(12):71–71. 10.1186/1471-2164-15-422 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Sharma MK, Kumar R, Solanke AU, Sharma R, Tyagi AK, Sharma AK. Identification, phylogeny, and transcript profiling of ERF family genes during development and abiotic stress treatments in tomato. Mol Genet Genomics 2010; 284:455–475. 10.1007/s00438-010-0580-1 [DOI] [PubMed] [Google Scholar]
- 54.Sun ZM, Zhou ML, Xiao XG, Tang YX, Wu YM. Genome-wide analysis of AP2/ERF family genes from Lotus corniculatus shows LcERF054 enhances salt tolerance. Functional & Integrative Genomics 2014; 14(3):453–466. 10.1007/s10142-014-0372-5 [DOI] [PubMed] [Google Scholar]
- 55.Nair SK, Phillips RL. Cleistogamous flowering in barley arises from the suppression of microRNA-guided HvAP2 mRNA cleavage. Proc Natl Acad Sci USA 2010; 107(1):490–495. 10.1073/pnas.0909097107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Houston K, McKim S M, Comadran J, Bonar N, Druka I, Uzrek N, et al. Variation in the interaction between alleles of HvAPETALA2 and microRNA172 determines the density of grains on the barley inflorescence. Proceedings of the National Academy of Sciences 2013; 110(41): 16675–16680. 10.1073/pnas.1311681110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Taketa S, Amano S, Tsujino Y, Sato T, Saisho D, Kakeda K, et al. Barley grain with adhering hulls is controlled by an ERF family transcription factor gene regulating a lipid biosynthesis pathway. Proc Natl Acad Sci USA 2008; 105(10): 4062–4067. 10.1073/pnas.0711034105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Matsukura S, Mizoi J, Yoshida T, Todaka D, Ito Y, Maruyama K, et al. Comprehensive analysis of rice DREB2-type genes that encode transcription factors involved in the expression of abiotic stress-responsive genes. Mol Genet Genomics 2010; 283: 185–196. 10.1007/s00438-009-0506-y [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(TIF)
(TIF)
(TIF)
The phylogenetic tree was constructed based on the sequence alignments of seventy-four full-length DREB 1 and DREB2 genes from four species. The gene ID and names are illustrated in red for barley; black for rice; blue for Arabidopsis; and green for maize. The gene names were used in the present study according to published data [1, 4, 46, 58]. Bootstrap values from 1,000 replicates were indicated at each node and the scale represents branch lengths.
(TIF)
(XLS)
(DOCX)
(XLS)
(XLS)
(XLS)
(XLS)
(XLSX)
(XLSX)
(XLSX)
(XLSX)
(XLSX)
(XLSX)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.