

Abstract
Platelet contractile forces play a major role in clot retraction and help to hold hemostatic clots against the vessel wall. Platelet forces are produced by its cytoskeleton, which is composed of actin and nonmuscle myosin filaments. In this work, we studied the role of Rho kinase, myosin light-chain kinase, and myosin in the generation of contractile forces by using pharmacological inhibitors and arrays of flexible microposts to measure platelet forces. When platelets were seeded onto microposts, they formed aggregates on the tips of the microposts. Forces produced by the platelets in the aggregates were measured by quantifying the deflection of the microposts, which bent in proportion to the force of the platelets. Platelets were treated with small molecule inhibitors of myosin activity: Y-27632 to inhibit the Rho kinase (ROCK), ML-7 to inhibit myosin light-chain kinase (MLCK), and blebbistatin to inhibit myosin ATPase activity. ROCK inhibition reduced platelet forces, demonstrating the importance of the assembly of actin and myosin phosphorylation in generating contractile forces. Similarly, MLCK inhibition caused weaker platelet forces, which verifies that myosin phosphorylation is needed for force generation in platelets. Platelets treated with blebbistatin also had weaker forces, which indicates that myosin's ATPase activity is necessary for platelet forces. Our studies demonstrate that myosin ATPase activity and the regulation of actin–myosin assembly by ROCK and MLCK are needed for the generation of platelet forces. Our findings illustrate and explain the importance of myosin for clot compaction in hemostasis and thrombosis.
Introduction
In hemostasis, platelets produce forces that help to maintain structural integrity of a hemostatic clot. Platelet forces lead to clot compaction by pulling together fibrin strands and nearby platelets into tight proximity with one another [1]. Reducing the size of the clot alleviates the fluid drag forces acting on it and lessens the risk of its detachment and embolization [2–5]. Platelet forces are transmitted through integrin receptors, such as αIIbβ3. These forces maintain the engagement of integrins to their matrix ligands through mechanotransduction pathways [6]. Since platelet forces help to pull adjacent platelets closer together, it can be inferred that these forces reduce the paracellular space within a clot and thereby increase the concentration of agonists that activate platelets [1]. The ability of platelets to generate contractile forces has been well known [7], but the mechanisms that regulate platelet forces and their direct contribution to clot compaction have not been fully established.
Nonmuscle myosin IIA is the predominate isoform of the myosin family that is present in platelets and is encoded by the MYH9 gene [8,9]. When the regulatory light chain of myosin is phosphorylated by MLCK or ROCK, myosin is able to form bipolar filaments, which are needed for the assembly of actin–myosin filaments [8]. Myosin relies on its ATPase activity in order to move along an actin filament. The procession of myosin causes actin filaments to slide past each other, creating tension in the cytoskeleton. The tension is transmitted to the adhesions of a cell through its integrin receptors as a contractile force [8].
In platelets, actin–myosin interactions are known to play a major role in shape change [10,11] and clot retraction [12]. In particular, ROCK is involved in forming focal adhesions [13] and actin–myosin filaments [14], which are needed for the stable attachment of platelets to a surface. It is likely that MLCK, ROCK, and myosin's ATPase activity play a role in the generation of platelet forces, but these relationships have not been examined due to lack of techniques for the direct measurement of platelet force.
Platelet forces have been quantified in blood or platelet–plasma samples by using isometric testing, clot retractometry, and rheological techniques [15–17]. These techniques have demonstrated the importance of platelet forces in hemostasis, but the measurements of platelet force are obscured by the degree of fibrin formation and clot lysis [18]. Recently, new techniques like atomic force microscopy [19], polyacrylamide gels [19,20], and micropost arrays [21] have been developed to measure platelet forces at the microscale. In particular, micropost arrays have been used to measure the forces generated by platelet aggregates, which are similar to the platelet-rich plugs that form during primary hemostasis [21].
In this study, we used micropost arrays to investigate the role of myosin regulation in the generation of platelet forces. We treated platelets with small molecule inhibitors of myosin activity. Specifically, Y-27632 [22], ML-7 [23], and blebbistatin [24] were used to inhibit ROCK, MLCK, and ATPase activity of myosin, respectively. We find that platelet forces are significantly reduced with each of the inhibitions, which demonstrates the important role of myosin in hemostasis.
Materials and Methods
Subjects.
Human whole blood samples were collected from donors by licensed professionals at the University of Washington, Hall Health Center, Seattle, WA. Blood samples were collected from donors who had not taken any antiplatelet medications within 2 weeks prior to the donation. Written informed consent was obtained from all the donors under a protocol approved by the Human Subject Division at the University of Washington.
Sample Preparation.
Tubes containing acid–citrate–dextrose (ACD; BD Medical, Franklin Lakes, NJ) solution was used to collect blood in order to prevent coagulation. To separate platelets from whole blood, two steps of centrifugation were performed. First, platelet-rich plasma (PRP) was collected after 20 min of centrifugation at 120×g. The PRP was then transferred to a new tube with an equal volume of CGS washing buffer (120 mM NaCl, 13 mM sodium citrate, and 30 mM glucose at pH 7) and centrifuged at 1200×g for 10 min. Platelet poor plasma was removed from the tube and Tyrode buffer (10 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid, 138 mM NaCl, 5.5 mM glucose, 12 mM NaHCO3, 0.36 mM Na2HPO4, 2.9 mM KCl, 0.4 mM MgCl2, and 0.8 mM CaCl2 at pH 7.5) was added. The pellet at the bottom of the tube was aspirated gently to put the platelets back into suspension in Tyrode buffer.
Sensor Fabrication and Preparation.
We used micropost arrays to measure platelet forces, as previously reported [21]. Briefly, arrays of microposts were replicated in polydimethylsiloxane (PDMS, Sylgard 184, Dow Corning) [25,26]. The arrays contain vertical cantilevers with microscale dimensions: 2.2 μm in diameter, 6.3 μm in height, and 9 μm in center-to-center spacing (Fig. 1(a)). Microcontact printing was used to transfer a coating of fibronectin (50 μg/ml; BD Biosciences) onto the tips of the microposts. Microposts were then fluorescently labeled with Alexa Fluor® 594-conjugated bovine serum albumin (BSA) (5 μg/ml; Invitrogen). The BSA coating on the base and sides of the microposts partially blocked platelet adhesion. Additionally, the arrays of microposts were treated with 0.2% Pluronic F-127 (BASF, Florham Park, NJ) for 1 h to completely block the nonspecific adhesion of platelets.
Fig. 1.
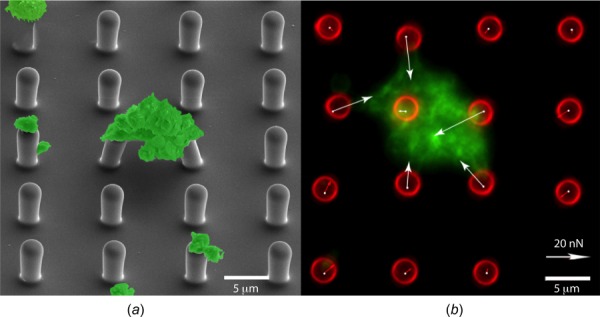
Platelet aggregates on arrays of microposts. (a) Scanning electron microscopy image of platelets on an array of microposts with pseudocoloring (green). (b) Fluorescent image of a platelet aggregate on microposts. Forces are measured by analyzing the deflection of the microposts. Green: actin and red: microposts.
Inhibitor Assay.
Substrates containing the arrays of microposts were submerged in Tyrode buffer. Platelets were incubated with micropost arrays for 30 min. Upon contact with the tips of the microposts, platelets adhered and formed aggregates. Unbound platelets were then washed, and the remaining platelets aggregated on the microposts were incubated with human α-thrombin (1 U/ml; Enzyme Research Laboratories) for 15 min. We then treated the samples with inhibitors—Y-27632 (10 or 50 μM; Sigma, St. Louis, MO), ML-7 (5 or 20 μM; Sigma), or blebbistatin (10 or 50 μM; Sigma)—for 5 min before fixation with 4% paraformaldehyde. Samples were permeabilized with 0.1% Triton X-100 (Sigma), and actin filaments were fluorescently labeled with Alexa Fluor® 488-conjugated phalloidin (Invitrogen). Samples of platelet aggregates treated with thrombin but not with any inhibitor were used as a control.
Force Measurement.
A Nikon TiE inverted microscope with a 60× oil objective was used to obtain fluorescent images of platelet aggregates that attached to the microposts (Fig. 1(b)). These images were processed using an analysis code developed in matlab to measure the deflection of the microposts and thereby calculate the forces of the platelet aggregates.
The contractile force at each of these microposts was calculated using Euler–Bernoulli beam theory: F = (3πED4/64H3)δ, where E is the Young's modulus of PDMS (2.5 MPa), D is the diameter of the micropost (2.2 μm), H is the height of the micropost (6.3 μm), and δ is the deflection of the tip of microposts. From this measurement, the total force was calculated, which is the sum of the magnitude of contractile forces acting on all the posts underneath a platelet aggregate. Since the calculation of total force depends on the number of posts underneath an aggregate, its value increases with the size of an aggregate. To compare the contractile force for platelet aggregates with different sizes, the average force per post was quantified by dividing the total force by the number of posts on which an aggregate was attached. Thus, force per post was used to normalize the data by the size of a platelet aggregate.
Statistics.
Samples were analyzed for significance using ANOVA with Bonferroni's post-hoc adjustment. Comparisons were considered significant for p-values less than 0.05 and are marked with an asterisk in the graphs. Error bars in the graphs denote the standard error of the mean.
Results and Discussion
We first investigated the role of the Rho kinase (ROCK) in the generation of contractile forces by platelet aggregates on the microposts. ROCK phosphorylates myosin and prevents myosin phosphatase activity, both of which lead to a greater amount of myosin bipolar filaments [22,27]. We used a ROCK inhibitor, Y-27632, to assess the role of ROCK activity in regulating platelet forces (Fig. 2(a)). Our results indicate that platelet forces are decreased by approximately 50% with the addition of Y-27632 (Fig. 2(b)). Previous studies have shown that ROCK is needed for the formation of tightly packed platelet aggregates and its inhibition causes thrombi to be unstable [12]. Normally, there is a 30–40% reduction in the net volume of a thrombus, but this compaction is significantly reduced when platelets received ROCK inhibitors such as H-1152 or HA-1077 [12]. The partial loss of platelet forces due to inhibition of ROCK may be the reason for the loss of clot compaction observed in these previous studies. Our results complement these findings and what is known about the role of ROCK activity in hemostasis.
Fig. 2.
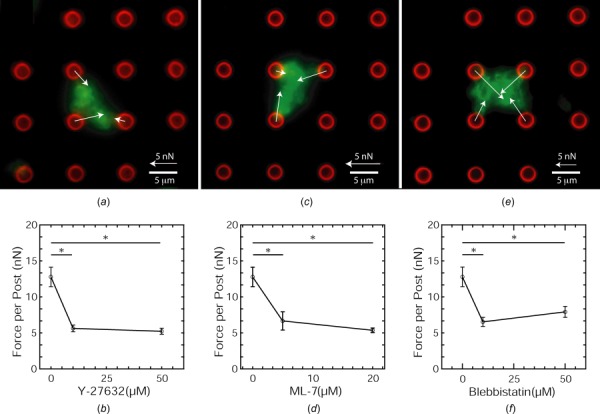
Platelet contractile forces are myosin-dependent. (a) Fluorescent image of a platelet aggregate on microposts treated with 10 μM Y-2732. (b) Platelets forces are reduced by Y-27632 concentration. (c) Fluorescent image of a platelet aggregate on microposts treated with 5 μM ML-7. (d) Platelets forces are reduced by ML-7 concentration. (e) Fluorescent image of a platelet aggregate on microposts treated with 10 μM blebbistatin. (f) Platelets forces are reduced by blebbistatin concentration. Green: actin and red: microposts.
MLCK can be considered to have a more direct role on nonmuscle myosin than ROCK. MLCK phosphorylates the regulatory light chain of myosin through a Ca2+/calmodulin-dependent pathway and does not affect myosin phosphatase or actin polymerization [23]. Assaying the level of myosin phosphorylation has been used previously to measure the activity of MLCK in platelets, but quantifying platelet forces can be another way to measure MLCK activity. Here, we used small molecule inhibitor ML-7 to inhibit MLCK activity in platelets on microposts (Figs. 2(c) and 2(d)). The use of ML-7 leads to a decrease in platelet forces. Previously, it was found that MLCK inhibition by chelating extracellular calcium with ethylene glycol-bis(β-aminoethyl ether)-N,N,N',N'-tetraacetic acid reduced clot compaction up to 52% [12]. Our results further suggest that the loss of clot compaction may be due to a loss in contractility of platelets within those clots.
Tension is generated in a platelet's cytoskeleton by myosin's hydrolysis of adenosine triphosphate [24]. To inhibit myosin's ATPase activity, we treated platelets with a small molecule inhibitor, blebbistatin. Blebbistatin stabilizes the binding of adenosine diphosphate to myosin, which thereby inhibits its ATPase activity while not affecting the assembly of its bipolar filaments [28]. We found that platelet forces decrease with the addition of blebbistatin (Figs. 2(e) and 2(f)). Inhibition of myosin activity with blebbistatin has been shown to prevent the tight packing of aggregated platelets in vitro, which undermines the stability of a thrombus [12]. In vivo studies also report that blebbistatin causes a 38% reduction in clot compaction [12]. Our results show similar reduction of contractile forces when myosin's ATPase activity is inhibited in actin–myosin filaments as the percent loss of reduction observed previously, which supports the role of platelet forces in clot compaction and thrombus stability.
The loss of nonmuscle myosin is present in a class of rare hereditary disorders that are marked by a mutation in the MYH9 gene, which encodes nonmuscle myosin heavy chain IIA. Patients with MYH9 disorders have thrombocytopenia, large platelets, and suffer from severe bleeding, even though platelet aggregation and secretion are normal [9]. It is likely that the increase bleeding in patients with MYH9 disorder is due to poor clot compaction [11,14]. ROCK, MLCK, and ATPase activity inhibition all lead to a reduction in the contractile forces of platelet aggregates. Taken together, we propose that platelet forces play an essential role in the stability of hemostatic clots through clot compaction, which is underscored by the importance of myosin phosphorylation and ATPase activity in generating these contractile forces.
Acknowledgment
This work was supported in part by grants from the NIH (HL097284 and EB001650), a National Science Foundation CAREER Award and Nanotechnology Undergraduate Education Program, and a Defense Advanced Research Program Agency (DARPA) Young Faculty Award.
The authors declare that N. J. Sniadecki is a cofounder and N. J. Sniadecki and S. Feghhi have equity in Stasys Medical Corporation.
Contributor Information
Shirin Feghhi, Department of Mechanical Engineering, , University of Washington, , Seattle, WA 98195 , e-mail: shfeghhi@uw.edu.
Wes W. Tooley, Department of Mechanical Engineering, , University of Washington, , Seattle, WA 98195 , e-mail: westooley@gmail.com
Nathan J. Sniadecki, Department of Mechanical Engineering, , University of Washington, , Seattle, WA 98195 , e-mail: nsniadec@uw.edu
References
- [1]. Brass, L. F. , Zhu, L. , and Stalker, T. J. , 2005, “ Minding the Gaps to Promote Thrombus Growth and Stability,” J. Clin. Invest., 115(12), pp. 3385–3392. 10.1172/JCI26869 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [2]. Basmadjian, D. , 1984, “ The Hemodynamic Forces Acting on Thrombi, From Incipient Attachment of Single Cells to Maturity and Embolization,” J. Biomech., 17(4), pp. 287–298. 10.1016/0021-9290(84)90139-8 [DOI] [PubMed] [Google Scholar]
- [3]. Basmadjian, D. , 1986, “ The Hemodynamic and Embolizing Forces Acting on Thrombi-II. The Effect of Pulsatile Blood Flow,” J. Biomech., 19(10), pp. 837–845. 10.1016/0021-9290(86)90134-X [DOI] [PubMed] [Google Scholar]
- [4]. Basmadjian, D. , 1989, “ Embolization: Critical Thrombus Height, Shear Rates, and Pulsatility. Patency of Blood Vessels,” J. Biomed. Mater. Res., 23(11), pp. 1315–1326. 10.1002/jbm.820231108 [DOI] [PubMed] [Google Scholar]
- [5]. Stalker, T. J. , Traxler, E. A. , Wu, J. , Wannemacher, K. M. , Cermignano, S. L. , Voronov, R. , Diamond, S. L. , and Brass, L. F. , 2013, “ Hierarchical Organization in the Hemostatic Response and Its Relationship to the Platelet-Signaling Network,” Blood, 121(10), pp. 1875–1885. 10.1182/blood-2012-09-457739 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [6]. Geiger, B. , Bershadsky, A. , Pankov, R. , and Yamada, K. M. , 2001, “ Transmembrane Extracellular Matrix-Cytoskeleton Crosstalk,” Nat. Rev. Mol. Cell Biol., 2(11), pp. 793–805. 10.1038/35099066 [DOI] [PubMed] [Google Scholar]
- [7]. Cohen, I. , 1979, “ The Contractile System of Blood Platelets and Its Function,” Methods Achiev. Exp. Pathol., 9, pp. 40–86.http://europepmc.org/abstract/med/368520 [PubMed] [Google Scholar]
- [8]. Vicente-Manzanares, M. , Ma, X. , Adelstein, R. S. , and Horwitz, A. R. , 2009, “ Non-Muscle Myosin II Takes Centre Stage in Cell Adhesion and Migration,” Nat. Rev. Mol. Cell Biol., 10(11), pp. 778–790. 10.1038/nrm2786 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [9]. Noris, P. , Spedini, P. , Belletti, S. , Magrini, U. , and Balduini, C. L. , 1998, “ Thrombocytopenia, Giant Platelets, and Leukocyte Inclusion Bodies (May-Hegglin Anomaly): Clinical and Laboratory Findings,” Am. J. Med., 104(4), pp. 355–360. 10.1016/S0002-9343(98)00062-X [DOI] [PubMed] [Google Scholar]
- [10]. Daniel, J. L. , Molish, I. R. , Rigmaiden, M. , and Stewart, G. , 1984, “ Evidence for a Role of Myosin Phosphorylation in the Initiation of the Platelet Shape Change Response,” J. Biol. Chem., 259(15), pp. 9826–9831.http://www.jbc.org/content/259/15/9826 [PubMed] [Google Scholar]
- [11]. Leon, C. , Eckly, A. , Hechler, B. , Aleil, B. , Freund, M. , Ravanat, C. , Jourdain, M. , Nonne, C. , Weber, J. , Tiedt, R. , Gratacap, M. P. , Severin, S. , Cazenave, J. P. , Lanza, F. , Skoda, R. , and Gachet, C. , 2007, “ Megakaryocyte-Restricted MYH9 Inactivation Dramatically Affects Hemostasis While Preserving Platelet Aggregation and Secretion,” Blood, 110(9), pp. 3183–3191. 10.1182/blood-2007-03-080184 [DOI] [PubMed] [Google Scholar]
- [12]. Ono, A. , Westein, E. , Hsiao, S. , Nesbitt, W. S. , Hamilton, J. R. , Schoenwaelder, S. M. , and Jackson, S. P. , 2008, “ Identification of a Fibrin-Independent Platelet Contractile Mechanism Regulating Primary Hemostasis and Thrombus Growth,” Blood, 112(1), pp. 90–99. 10.1182/blood-2007-12-127001 [DOI] [PubMed] [Google Scholar]
- [13]. Leng, L. , Kashiwagi, H. , Ren, X. D. , and Shattil, S. J. , 1998, “ RhoA and the Function of Platelet Integrin alphaIIbbeta3,” Blood, 91(11), pp. 4206–4215.http://www.bloodjournal.org/content/91/11/4206 [PubMed] [Google Scholar]
- [14]. Calaminus, S. D. J. , Auger, J. M. , McCarty, O. J. T. , Wakelam, M. J. O. , Machesky, L. M. , and Watson, S. P. , 2007, “ Myosinlla Contractility is Required for Maintenance of Platelet Structure During Spreading on Collagen and Contributes to Thrombus Stability,” J. Thromb. Haemostasis, 5(10), pp. 2136–2145. 10.1111/j.1538-7836.2007.02696.x [DOI] [PubMed] [Google Scholar]
- [15]. Cohen, I. , and De Vries, A. , 1973, “ Platelet Contractile Regulation in an Isometric System,” Nature, 246(5427), pp. 36–37. 10.1038/246036a0 [DOI] [PubMed] [Google Scholar]
- [16]. Carr, M. E., Jr. , and Zekert, S. L. , 1991, “ Measurement of Platelet-Mediated Force Development During Plasma Clot Formation,” Am. J. Med. Sci., 302(1), pp. 13–18. 10.1097/00000441-199107000-00004 [DOI] [PubMed] [Google Scholar]
- [17]. Jen, C. J. , and McIntire, L. V. , 1982, “ The Structural Properties and Contractile Force of a Clot,” Cell Motil., 2(5), pp. 445–455. 10.1002/cm.970020504 [DOI] [PubMed] [Google Scholar]
- [18]. Feghhi, S. , and Sniadecki, N. J. , 2011, “ Mechanobiology of Platelets: Techniques to Study the Role of Fluid Flow and Platelet Retraction Forces at the Micro- and Nano-Scale,” Int. J. Mol. Sci., 12(12), pp. 9009–9030. 10.3390/ijms12129009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [19]. Lam, W. A. , Chaudhuri, O. , Crow, A. , Webster, K. D. , Li, T. D. , Kita, A. , Huang, J. , and Fletcher, D. A. , 2011, “ Mechanics and Contraction Dynamics of Single Platelets and Implications for Clot Stiffening,” Nat. Mater., 10(1), pp. 61–66. 10.1038/nmat2903 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [20]. Schwarz Henriques, S. , Sandmann, R. , Strate, A. , and Koster, S. , 2012, “ Force Field Evolution During Human Blood Platelet Activation,” J. Cell Sci., 125(Pt. 16), pp. 3914–3920. 10.1242/jcs.108126 [DOI] [PubMed] [Google Scholar]
- [21]. Liang, X. M. , Han, S. J. , Reems, J. A. , Gao, D. , and Sniadecki, N. J. , 2010, “ Platelet Retraction Force Measurements Using Flexible Post Force Sensors,” Lab Chip, 10(8), pp. 991–998. 10.1039/b918719g [DOI] [PMC free article] [PubMed] [Google Scholar]
- [22]. Ishizaki, T. , Uehata, M. , Tamechika, I. , Keel, J. , Nonomura, K. , Maekawa, M. , and Narumiya, S. , 2000, “ Pharmacological Properties of Y-27632: A Specific Inhibitor of Rho-Associated Kinases,” Mol. Pharmacol., 57(5), pp. 976–983.http://molpharm.aspetjournals.org/content/57/5/976 [PubMed] [Google Scholar]
- [23]. Itoh, K. , Hara, T. , Yamada, F. , and Shibata, N. , 1992, “ Diphosphorylation of Platelet Myosin Ex Vivo in the Initial Phase of Activation by Thrombin,” Biochim. Biophys. Acta, 1136(1), pp. 52–56. 10.1016/0167-4889(92)90084-O [DOI] [PubMed] [Google Scholar]
- [24]. Kovacs, M. , Toth, J. , Hetenyi, C. , Malnasi-Csizmadia, A. , and Sellers, J. R. , 2004, “ Mechanism of Blebbistatin Inhibition of Myosin II,” J. Biol. Chem., 279(34), pp. 35557–35563. 10.1074/jbc.M405319200 [DOI] [PubMed] [Google Scholar]
- [25]. Tan, J. L. , Tien, J. , Pirone, D. M. , Gray, D. S. , Bhadriraju, K. , and Chen, C. S. , 2003, “ Cells Lying on a Bed of Microneedles: An Approach to Isolate Mechanical Force,” Proc. Natl. Acad. Sci. U.S.A., 100(4), pp. 1484–1489. 10.1073/pnas.0235407100 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [26]. Sniadecki, N. J. , and Chen, C. S. , 2007, “ Microfabricated Silicone Elastomeric Post Arrays for Measuring Traction Forces of Adherent Cells,” Methods Cell Biol., 83, pp. 313–328. 10.1016/S0091-679X(07)83013-5 [DOI] [PubMed] [Google Scholar]
- [27]. Burridge, K. , and Wennerberg, K. , 2004, “ Rho and Rac Take Center Stage,” Cell, 116(2), pp. 167–179. 10.1016/S0092-8674(04)00003-0 [DOI] [PubMed] [Google Scholar]
- [28]. Zhang, M. , and Rao, P. V. , 2005, “ Blebbistatin: A Novel Inhibitor of Myosin II ATPase Activity, Increases Aqueous Humor Outflow Facility in Perfused Enucleated Porcine Eyes,” Invest. Ophthalmol. Visual Sci., 46(11), pp. 4130–4138. 10.1167/iovs.05-0164 [DOI] [PubMed] [Google Scholar]