

ABSTRACT
Aging has been associated with a decline in relational memory, which is critically supported by the hippocampus. By adapting the transitivity paradigm (Bunsey and Eichenbaum (1996) Nature 379:255‐257), which traditionally has been used in nonhuman animal research, this work examined the extent to which aging is accompanied by deficits in relational learning and flexible expression of relational information. Older adults' performance was additionally contrasted with that of amnesic case DA to understand the critical contributions of the medial temporal lobe, and specifically, the hippocampus, which endures structural and functional changes in healthy aging. Participants were required to select the correct choice item (B versus Y) based on the presented sample item (e.g., A). Pairwise relations must be learned (A‐>B, B‐>C, C‐>D) so that ultimately, the correct relations can be inferred when presented with a novel probe item (A‐>C?Z?). Participants completed four conditions of transitivity that varied in terms of the degree to which the stimuli and the relations among them were known pre‐experimentally. Younger adults, older adults, and DA performed similarly when the condition employed all pre‐experimentally known, semantic, relations. Older adults and DA were less accurate than younger adults when all to‐be‐learned relations were arbitrary. However, accuracy improved for older adults when they could use pre‐experimentally known pairwise relations to express understanding of arbitrary relations as indexed through inference judgments. DA could not learn arbitrary relations nor use existing knowledge to support novel inferences. These results suggest that while aging has often been associated with an emerging decline in hippocampal function, prior knowledge can be used to support novel inferences. However, in case DA, significant damage to the hippocampus likely impaired his ability to learn novel relations, while additional damage to ventromedial prefrontal and anterior temporal regions may have resulted in an inability to use prior knowledge to flexibly express indirect relational knowledge. © 2015 The Authors Hippocampus Published by Wiley Periodicals, Inc.
Keywords: memory, aging, amnesia, transitivity, semantic knowledge, hippocampus
INTRODUCTION
An important facet of memory is the ability to flexibly express stored knowledge. In particular, the organization of memory as a set of overlapping representations allows for relations among items to be accessed and used to make inferential judgments in novel situations (Bunsey and Eichenbaum, 1996). The act of making inferences—that is, making relations indirectly, rather than through direct learning—is a central component of problem solving and creativity and guides behavior, including social interactions (Koscik and Tranel, 2012; Holyoak, 2012). However, the establishment of new relational memories is typically disrupted in healthy aging (Naveh‐Benjamin, 2000; Bastin and Van der Linden, 2006), which ultimately has negative consequences for inferential behavior (Ryan et al., 2009). Age‐related deficits in relational memory have been attributed to a decline in hippocampal function: older adults show structural and/or synaptic changes in the hippocampus (Driscoll et al., 2003; Morrison and Baxter, 2012; Gordon et al., 2013), altered functional recruitment of the hippocampus as revealed through neuroimaging (Grady et al., 2003; Daselaar et al., 2006; Tsukiura et al., 2011), and impaired performance relative to younger adults on tasks on which human and/or non‐human animals with hippocampal lesions typically show deficits (Castel and Craik, 2003; Healy et al., 2005; Ryan et al., 2007; Old and Naveh‐Benjamin, 2008; Ryan et al., 2009; Ostreicher et al., 2010).
Nonetheless, the use of prior knowledge has been shown to mitigate the traditionally observed relational memory deficit in aging, elevating performance to levels commensurate with those of younger adults (Castel, 2005; Ostreicher et al., 2010). Existing schemas of prior knowledge provide an organizational structure to support new learning (Bartlett, 1932) and in particular, allow for the rapid integration of hippocampal‐dependent learning to become assimilated into a cortical network (Tse et al., 2007, 2011), that critically includes the functions of the anterior cingulate cortex and vmPFC (Wang and Morris, 2010; Wang et al., 2012). Yet, it remains to be determined whether older adults can make use of prior knowledge to support the flexible expression of novel inferences. To date, the paradigms that have examined the influence of prior knowledge on the acquisition of relational memory have primarily focused on the testing of direct, previously studied, relations and have seldom focused on the testing of novel inferences.
For instance, the transverse patterning (TP) task requires relations to be learned among three stimuli (A,B,C) where each stimulus wins in the context of one of the other items and loses in the context of the other item. All possible relations (A wins over B, B wins over C, C wins over A) are learned and tested, and the testing of inferences is therefore not possible (e.g., Ostreicher et al., 2010). When task designs have allowed for inferential performance to be examined, inferential judgments could conceivably have been made based on associative strength of individual items rather than the relations among them, such as in the transitive inference (TI) task in which participants learn a series of pairwise relations (A wins over B, B wins over C, C wins over D, D wins over E) and infer a hierarchy of stimuli (A‐B‐C‐D‐E) in which one stimulus always wins (A) and one stimulus always loses (E) (e.g., Moses et al., 2010a,2010b). Additionally, while findings from neuropsychological and neuroimaging studies suggest that age‐related deficits on relational inferential tasks may be due to declining hippocampal function (Heckers et al., 2004; Smith and Squire, 2005; Greene et al., 2006; Zalesak and Heckers, 2009), performance of older adults has not been directly compared with that of amnesic individuals with hippocampal damage. To examine the extent to which prior knowledge can be used to flexibly support inferential judgments of novel pairings, younger and older adults performed a transitivity task adapted from Bunsey and Eichenbaum (1996) for use with humans. During training of the transitivity task, a sample stimulus (e.g., A) and two choice stimuli (e.g., B, X) are provided, and participants must select the choice stimulus that “belongs” with a given sample stimulus (e.g., if “A,” choose “B”). Ultimately, the participant learns the relations among pairs of stimuli that comprise distinct sets (e.g., A‐>B, B‐>C, C‐>D; W‐>X, X‐>Y, Y‐>Z) such that an inference can be made when presented with a novel pairing (e.g., if “A” choose “C,” not “Y”). The present adaptation of the transitivity task included multiple conditions that varied in terms of whether the items and/or the relations among them were known before the experimental session through accumulated semantic knowledge. Critically, one condition required the expression of novel inferences, but the expression of such inferences could be done within the context of previously known relations (i.e., prior semantic knowledge), whereas another condition required the expression of novel inferences that were based on the learning of arbitrary relations.
Comparative data on this transitivity paradigm were also obtained from the amnesic case DA. As outlined in Rosenbaum et al. (2008), D.A. became amnesic after contracting herpes simplex encephalitis in 1993. Detailed volumetric analysis of high‐resolution MRI revealed severe reductions in D.A.'s medial temporal lobes, including the hippocampus bilaterally. Given the link between successful transitivity performance based on learning of arbitrary relations and the integrity of the hippocampal system (Bunsey and Eichenbaum, 1996), direct comparison of DAs performance to that of older vs. younger adults would illuminate the role of the hippocampus in any observed age‐related differences in the learning of arbitrary relations and subsequent inference performance.
DA also has damage to the right ventromedial prefrontal cortex (vmPFC; right ventral frontal cortex: z = −1.86 compared with the distribution of control data, see Rosenbaum et al., 2008), as well as the right anterior‐lateral temporal lobe (z = −11.15), regions that are not typically affected to the same extent as the hippocampus in healthy aging (Scahill et al., 2003). The vmPFC has been shown to be critical for making inference judgments, as individuals with lesions to this region are significantly impaired on tasks such as TI, despite showing intact learning and retention for the premise pairs (Koscik and Tranel, 2012). However, computational modeling suggests that the expression of inferential knowledge may be achieved through the continual strengthening of the memory traces for the premise pairs, as achieved through recurrence in the hippocampal system (Kumaran and McClelland, 2012). Thus comparing performance between DA and older adults on conditions that require flexible inference behavior as predicated on pre‐experimentally known (semantic) versus arbitrary relations might reveal the extent to which aging is associated with differential integrity of the vmPFC versus the hippocampus, and/or reveal the extent to which the hippocampus is required for all inferential behavior.
Participants were presented with four conditions of transitivity that varied in the extent to which the stimuli and the relations among them were known before the experiment. It was expected that DA and older adults would show similar performance to younger adults on conditions and problems for which known items with semantic relations were used, but show impaired performance relative to younger adults on the condition for which all relations were arbitrary and had to be learned within the confines of the experiment (Moses et al., 2008; Ostreicher et al., 2010; Ryan et al., 2013). Previous work has suggested that the hippocampus is engaged in the formation (Moscovitch et al., 2006; Konkel et al., 2008) and retrieval (Ryan et al., 2010; Hoscheidt et al., 2010) of all types of relational memory, including semantic memories, and that the hippocampus is critical for the rapid linking of new information to existing schemas (Tse et al., 2007, 2011), Therefore, it was expected that only the older adults, and not DA, would benefit from the use of semantic, pairwise, relations to make novel, flexible inferences and show performance similar to that of younger adults.
Additionally, the sample and the choice stimuli were provided either simultaneously during training and/or test or separated by a delay. Given the particular role for the hippocampus in bridging information across time (Wallenstein et al., 1998; Bangasser et al., 2006), this manipulation allowed us to investigate the extent to which relational memory performance could be altered through simultaneous versus delayed presentations as well as through the use of prior knowledge. The present work then allowed us to examine the extent to which deficits in relational memory and inferential judgments, either due to age‐related changes or due to a case of amnesia, can be mitigated through the use of previously acquired knowledge and through altered training procedures.
METHODS
Participants
Thirty‐six healthy younger adults (age range: 18–28; mean age = 23.1, SE = 0.5) and 36 older adults (age range: 57–84; mean age = 68.5, SE = 1.3) with no known pathology were recruited from the volunteer participant pool at the Rotman Research Institute at Baycrest and participated in exchange for monetary compensation. All participants gave informed written consent. The rights of the participants were protected and the guidelines of the Toronto Academic Health Science Council were followed.
Three versions of the experiment were manipulated between‐subjects which varied the presentation timing procedures of the sample and choice stimuli during training and test phases (see Procedure, below). Each experimental version contained the same four within‐subjects conditions. Participant age was not significantly different across experimental versions (F < 1) and there was no interaction between experiment version and participant group (younger, older) on participant age (F < 1).
Younger adults (M = 16.3, SE = 0.3) and older adults (M = 16.7, SE = 0.6) had similar years of education that did not vary across experiment version (F < 1). However, there was a significant experiment version by group interaction (F (2,72) = 4.58; P < 0.05). Follow‐up univariate ANOVAs revealed that the younger adults showed a significant difference in education across the experiment versions (F (2,33) = 3.40; P < 0.05); participants in the onset + delay version had the most years of education (M = 17.2, SE = 0.5) and those in the onset condition had the least (M = 15.4, SE = 0.6). Education did not significantly vary across experimental versions for older adults (F (2,33) = 2.08; P > 0.10). As previously found (Ryan et al., 2007), older adults (M = 28.1, SE = 1.5) outperformed the younger adults (14.9, SE = 1.4) on the Extended Range Vocabulary Test (F (1,72) = 37.09, P < 0.001). The main effect of experiment version and the interaction with age group were each nonsignificant (Fs < 1).
Eleven additional older adults were tested in the current study, but were excluded for the following reasons. Four older adults from the Onset condition (age range: 83–86) were replaced in order to match age across experimental conditions. One participant was excluded as they were tested in the wrong experimental condition. An additional six older adults (age range: 63–82, mean age = 68) were tested in the study, but were excluded for having failed the training phase in the novel objects/arbitrary relations conditions, and thus did not complete the test phase in this condition. Importantly, while these older adults had low accuracy in the novel objects/arbitrary relations and known objects/arbitrary relations conditions (mean accuracy = 0.52 and 0.68, respectively), these same older adults were highly accurate in the semantic and pairwise relations conditions (mean accuracy = 0.96 and 0.91, respectively). The seven older adults who were excluded for reasons other than age matching did not differ from the included older adults in terms of age (P = 0.63), education (P = 0.87), or ERVT scores (P = 0.33).
Data were also obtained from amnesic case DA (56 years old at initial testing) who was tested over nine sessions across 2 years. As outlined in Rosenbaum et al. (2008), DA is a right‐handed man with 17 years of education who contracted herpes encephalitis in 1993, resulting in bilateral MTL damage. DA's right MTL damage includes the hippocampus and parahippocampal gyrus along its entire extent (perirhinal, entorhinal, and parahippocampal cortices), and the right anterior and posterior temporal lobe (see Fig. 1). The left hippocampus and entorhinal cortex are less than 1/3 volume of that of age‐matched controls, and there is nearly complete loss of his left perirhinal and parahippocampal cortices. DA's cognitive function is generally intact other than extensive anterograde amnesia and a temporally graded retrograde amnesia extending approximately 30 years for autobiographical events (Rosenbaum et al., 2008). Critically for the work here, DA shows preserved access to, and use of, information stored within semantic memory (Westmacott and Moscovitch, 2002), including the ability to successfully retrieve and process previously acquired relations, as demonstrated in a transverse patterning task (Ryan et al., 2013). DA was exposed to each experimental version across the nine sessions (two sessions of the “delay” version, five sessions of the “onset‐delay” version, and two sessions of the “onset” version; see details below); however, performance did not vary across the experimental versions, and data from all nine sessions were combined.
Figure 1.
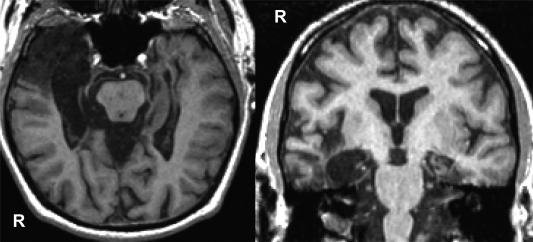
T1‐weighted images in axial (left) and coronal (right) views showing lesions to bilateral medial temporal lobe and right anterior temporal cortex in DA.
Stimuli
Transitivity performance for all participants was assessed across four conditions that varied on the extent to which the items and the relations among them were known before the experiment: (1) known objects/semantic relations; (2) known objects/pairwise relations; (3) known objects/arbitrary relations; (4) novel objects/arbitrary relations. For the younger and the older adults, the presentation of conditions was counterbalanced across participants such that each condition was presented equally often as the first condition. However, DA always performed the experimental conditions in the following order: known objects/semantic relations; known objects/pairwise relations; known objects/arbitrary relations; novel objects/arbitrary relations. This was done to ensure that DA had maximum understanding of the task requirements. The stimuli for each condition are shown in Figure 2; eight unique colored stimuli were used for each condition, which were grouped into two sets of four (A‐B‐C‐D; W‐X‐Y‐Z, wherein each letter represents a unique item). The novel stimuli were created in Corel Draw v.12 and were among the set used in our prior work (Ryan et al., 2007). The known stimuli were common, nameable objects (e.g., hammer) selected from the Hemera 3.01 database.
Figure 2.
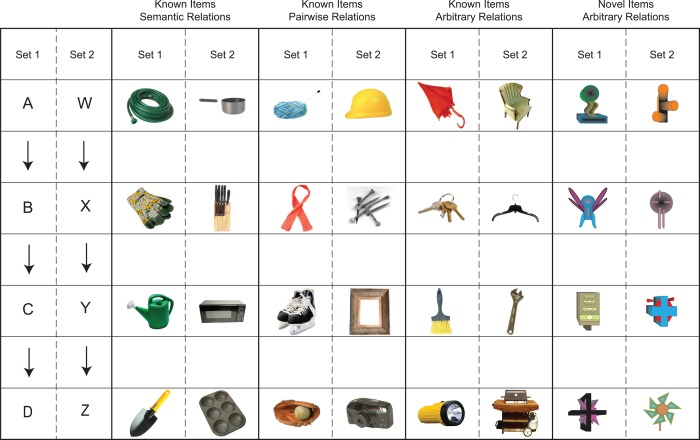
Pictures used for the ABCD and WXYZ stimulus sets across the four experimental conditions (Known Items/Semantic Relations, Known Items/Pairwise Relations; Known Items/Arbitrary Relations; Novel Items/Arbitrary Relations). [Color figure can be viewed in the online issue, which is available at wileyonlinelibrary.com.]
The stimuli for the known objects/semantic relations included a group of four kitchen instruments and a group of four gardening tools. The stimuli for the known objects/pairwise relations consisted of items for which there are meaningful relationships within each consecutive pair (A‐B, B‐C, C‐D). For instance, as in Figure 2, a ball of yarn can be used to make a scarf (A‐B), and a scarf is worn with ice skates to go skating (B‐C), and ice skates and baseball gloves are sports equipment (C‐D). However, the relations among nonconsecutive items (A‐C, B‐D, A‐D) are arbitrary; that is, there is not an obvious prior relationship between a ball of yarn and ice skates, between a scarf and a baseball glove, nor between a ball of yarn and a baseball glove.
Training Procedure
Participants were presented with three objects on a computer screen—a sample object (e.g., A) and two choice objects (e.g., B versus X)—and were required to select the choice object that made a correct pairing with the sample object (e.g., B) through trial and error. Three training blocks were given followed by a no‐feedback test block (Fig. 3). During the first training block, each problem pair for one stimulus set (ABCD) was presented consecutively, and the sequence was then repeated four times (4 × {A→B vs. X, B→C vs. Y, C→D vs. Z}). The same procedure was followed for the second training block for the alternate stimulus set (WXYZ; 4 × {W→B vs. X, X→C vs. Y, Y→D vs. Z}). The third training block presented the problem pairs for each stimulus set from six times each, in random order (6 × {A→B vs. X, B→C vs. Y, C→D vs. Z, W→B vs. X, X→C vs. Y, Y→D vs. Z}). This methodology is similar to our previous transverse patterning and transitive inference paradigms (e.g., Moses et al., 2006; Ryan et al., 2013). Presentations of the choice stimuli were counterbalanced for right/left position across trials. Stimuli were presented using E‐prime 1.1 on a 19" monitor set at 1,024 × 768 pixel resolution and 24‐bit high color. Participants were asked to select the choice stimulus that “best fit” with the sample stimulus. Responses were made using the “p” or “q” keys on a standard computer keyboard to select the choice object on the right and left side of the screen, respectively.
Figure 3.

Top: depiction of the training and test sequences in the transitivity task. Trials were self‐paced and feedback was provided during the training phase only. The sample image was presented at the top of the screen and the two choice stimuli were presented along the bottom of the screen. The participants were required to select one of the two choice stimuli that was best associated with the sample stimulus. Test trials could depict a previously studied relation, or participants were required to make an inference across pairs of previously studied relations (novel probe pair). Novel probe pairs include pairs in which the sample was separated from the choice items by one‐away, two‐away or a mixture of one and two intervening items and their respective relations (mixed). Bottom: Depiction of the three experimental conditions (Onset, Onset + Delay, Delay). In the Onset condition, the training and test phases were identical. In this condition, the sample stimulus was presented alone for 2 s, after which the two choice stimuli appeared along with the sample stimulus. All three stimuli remained on the screen until a response as made. In the Onset + Delay condition, the training phase was identical to that of the Onset condition, while the test phase was identical to that of the Delay condition (described below). In the Delay condition, the training and test phases were identical. In this condition, the sample stimulus was presented alone for 2 s, followed by a blank screen was presented for 3 s, after which the two choice stimuli appeared on the screen alone until a response was made. [Color figure can be viewed in the online issue, which is available at wileyonlinelibrary.com.]
A criterion of 70% accuracy for each training block was required to advance to the subsequent training block, otherwise the block was repeated until criterion was reached. Testing within a condition was terminated if a training block was repeated six times without achieving criterion. Each trial was self‐initiated and terminated upon response. Feedback was provided in all of the training blocks. A correct response was rewarded with the appearance of a happy cartoon face and the caption “Good Job!,” and an incorrect response was followed by the appearance of an angry cartoon face and the caption “Wrong!.”
Test Procedure
In the test block, previously studied pairs (as noted above) and novel probe pairs were presented six times each in random order. Novel probe pairs include pairs in which the sample was separated from the choice items by one (one‐away: A→C vs. Y, B→D vs. Z, X→C vs. Y, Y→D vs. Z), two (2‐away: A→D vs. Z, X→ D vs. Z), or a mixture of one and two intervening items and their respective relations (mixed: A→C vs. Z, A→D vs. Y, X→C vs. Z, X→D vs. Y). Feedback was not provided during the test block.
Experimental Conditions
The above conditions and trials were used in three separate experimental versions that varied in the timing and presentation of the sample and choice items: delay, onset, onset‐delay (see Fig. 3). In the delay version, the sample item was presented for 2 s centered at the top half of the screen, followed by a 3‐s delay during which a blank screen was shown, and finally the two choice objects were shown on the bottom half of the screen until a response was given. This procedure was employed throughout the training and the test blocks.
During the training and test blocks for the onset version, the sample item was presented for 2 s centered on the top half of the screen, and then the two choice objects were added to the bottom half of the screen. The sample and choice items all remained on the screen until the participant responded.
For the onset‐delay version, training blocks followed the training procedures for the onset experiment, whereas the test block followed the procedures of the delay version of the experiment, as described above.
Issues of Awareness
Post‐experimental questionnaires were administered to each participant following each condition to assess awareness of the relationships and perceived learning strategies (c.f., Moses et al., 2006, 2010a,b). As part of the questionnaires, participants were shown the eight objects for the current condition and were asked to answer the following question: “Based on your understanding of the relationships among these objects, arrange the following objects in the most appropriate way. Use the spaces provided to place the objects in groups of four.” Individuals were considered aware of the relations if they grouped together the objects into the two groups correctly without any errors; otherwise, they were considered unaware.
The proportion of individuals classified as aware are listed in Table 1 as a function of age group and stimulus condition. Given the unequal proportions of individuals who were aware of the relationships across conditions and group, the discussion of awareness is limited here to a description of the patterns of awareness across groups and conditions. Overall, most of the younger and older adults were aware of the relationships in the semantic and pairwise relations conditions. In contrast, while more than half of the younger adults were aware in the two conditions that employed arbitrary relations, fewer older adults were aware in these two conditions, with the smallest proportion in the novel objects/arbitrary relations condition.
Table 1.
Proportion of Individuals Classified as “Aware” as a Function of Age Group and Stimulus Condition
| Known objects/known relations | Known objects/pairwise relations | Known objects/novel relations | Novel objects/novel relations | |
|---|---|---|---|---|
| Young Adults | 0.97 | 0.89 | 0.78 | 0.72 |
| Older Adults | 1 | 0.89 | 0.50 | 0.22 |
The effect of awareness on accuracy was examined by adding awareness as a between‐subject factor in separate analyses for each stimulus condition. Overall, aware participants had higher accuracy than unaware participants. Although awareness did interact with age group and trial type in the test phase for the known objects/pairwise relations and known objects/arbitrary relations conditions, it is difficult to ascertain the importance of these interactions with such a limited number of unaware participants in some conditions, and for such reasons, issues of awareness are not considered further here.
Analysis
A repeated‐measures ANOVA was conducted for the measure of accuracy on the training blocks. Between‐subjects factors included: age group (older, younger), experiment version (delay, onset, onset + delay), and the condition completed first in the experiment (known objects/semantic relations, known objects/pairwise relations, known objects/arbitrary relations, novel objects/arbitrary relations). Within‐subjects factors included condition (known objects/semantic relations, known objects/pairwise relations, known objects/arbitrary relations, novel objects/arbitrary relations). A separate, identical analysis was conducted on the number of trials required to complete the training phase. A repeated‐measures ANOVA was conducted for the measure of accuracy on the no‐feedback test block. The between‐subjects factors included were identical to the analysis on the training blocks. The within‐subjects factors included condition and trial type (studied pairs, one‐away, two‐away, mixed pairs). All interactions involving age group were broken down using independent samples t‐tests assuming unequal variance, and are reported with the nominal degrees of freedom and with P values that have been adjusted for multiple comparisons.
RESULTS
Although the years of education for younger adults differed significantly across experiment versions, education level was not a significant covariate (F = 1.02; P > 0.30), and education did not significantly interact with the within‐subject factors of condition (F (3,69) < 1), trial type (F (3,69) = 1.51; P > 0.20) or the interaction of condition by trial type (F (9,207) < 1). Therefore, education was not considered further in the present results. Below, training and test results for the younger and older adults are presented first, followed by the results for amnesic case DA.
Training Blocks—Accuracy and Number of Training Trials Required
Effects of experiment version
The main effect of experiment version was not significant in either analysis of the training blocks (accuracy: F (2,48) = 1.27, P = 0.29, number of trials: F < 1); however, the interaction between experiment version and stimulus version was significant in the analysis of accuracy (F (6,144) = 3.42, P = 0.01, ε = 0.72). The range in performance across stimulus conditions was largest in the onset condition (M = 0.79–0.98), followed by the delay condition (M = 0.82–0.97), and smallest in the onset + delay condition (M = 0.85–0.99). The interaction between experiment version and stimulus version was not significant in the analysis of number of training trials required (F (6,144) = 1.89, P = 0.09) No other interactions with experiment version were significant for either accuracy or number of trials.
Effects of condition order
The main effect of the condition completed first in the experiment was marginally significant, (accuracy: F (3,48) = 2.68, P = 0.06, number of trials: F (3,48) = 2.56, P = 0.07); participants who completed the novel objects/arbitrary relations condition first had lower accuracy (M = 0.85, SE = 0.02) and required more trials (M = 80, SE = 5.7) than the participants who completed the three other conditions (accuracy: M = 0.90, SE = 0.02, number of trials: range = 68–74, for other three conditions).
Effects of known versus novel relations
The main effect of stimulus condition was significant for accuracy (F (3,144) = 109.58, P < 0.001) and for the number of training trials (F (3,144) = 19.92, P < 0.001). As expected, accuracy was higher, and participants did not require additional training blocks, for the conditions in which the items and the relations among them were pre‐experimentally known. Accuracy was highest (M = 0.98, SE = 0.00) and the number of training trials was at the minimum (M = 60 trials, SE = 0) for the known objects/semantic relations condition, followed by the known objects/pairwise relations condition (accuracy: M = 0.97, SE = 0.00; trials: M = 60, SE = 0.5)), known objects/arbitrary relations condition (accuracy; M = 0.87; SE = 0.01; trials: M = 78, SE = 5.1), and the novel objects/arbitrary relations condition (accuracy: M = 0.82, SE = 0.02; trials: M = 92 trials, SE = 6.1). In sum, accuracy decreased, and the number of training trials increased, when the conditions required that the objects and the relations among them be learned within the confines of the experiment.
Importantly, the interaction between stimulus condition and the condition completed first was also significant for accuracy (F (9,144) = 6.83, P < 0.001) and for the number of training trials required (F (9,144) = 3.29, P = 0.001). The order of the conditions had the most dramatic effect on accuracy for the novel objects/arbitrary relations condition. When the novel objects/arbitrary relations condition was presented first, accuracy was low (M = 0.73; SE = 0.03) and the number of training trials was high (M = 121 trials; SE = 16.3), but if this condition followed any of the other conditions, accuracy was considerably higher (range = 0.83–0.88) and fewer trials were required during training (range = 77–89 trials). Similarly, for the known objects/arbitrary relations condition, accuracy was the lowest (M = 0.83, SE = 0.02) and the number of training trials required was highest (M = 97 trials, SE = 15.9), when this condition was presented first. Accuracy was higher (range = 0.87–0.90), and fewer training trials were required (range = 67–77 trials), on the known objects/arbitrary relations condition when it followed any of the other conditions. By contrast, when at least some of the relations were known before the experiment, accuracy (known objects/semantic relations: M = 0.97, SE = 0.01; known objects/pairwise relations: M = 0.96, SE = 0.01) was high, and the minimum number of trials were required for training (known objects/semantic relations: M = 60 trials, SE = 0; known objects/pairwise relations: M = 60 trials, SE = 0) regardless of whether it was presented first or followed another condition (known objects/semantic relations: accuracy range = 0.98–0.99; known objects/pairwise relations: accuracy range = 0.96–0.98; training trials M = 60 for all). Therefore, consistent with previous findings (Castel, 2005; Moses et al., 2010a,b; Ostreicher et al., 2010), when arbitrary relations had to be learned participants benefited from experience with a condition that used a relational schema.
Effects of age
In general, younger adults were more accurate (M = 0.93; SE = 0.01) and required fewer training trials (M = 64 trials, SE = 1.0) compared with the older adults (accuracy: M = 0.84, SE = 0.01; training trials required: M = 81 trials; SE = 4.0), as revealed by significant effects of age group on both measures (accuracy: F (1,48) = 27.44; P < 0.001; training trials required: F (1,48) = 24.18; P < 0.001; Fig. 4). The interaction of age group with stimulus condition was significant for accuracy (F (3,144) = 25.96, P < 0.001; Fig. 4, top panel) and for the number of training trials required (F (3,144) = 10.39, P < 0.001). Older adults were significantly less accurate (older adults: M = 0.75, SE = 0.02; younger adults: M = 0.89, SE = 0.02; t (70) = 5.47, P < 0.001) and required significantly more training trials (older adults: M = 114, SE = 10.5; younger adults: M = 69, SE = 3.1; t (70) = 4.15, P < 0.001) than younger adults on the novel objects/arbitrary relations condition . Older adults were also significantly less accurate on the known objects/arbitrary relations condition (older adults: M = 0.83, SE = 0.02; younger adults: M = 0.92, SE = 0.01; t (70) = 4.34, P < 0.001), and required numerically more training trials (older adults: M = 90, SE = 9.7; younger adults: M = 66, SE = 1.9). As expected, the older adults' performance was similar to that of the younger adults for the condition in which all the relations were pre‐experimentally known (for known objects/semantic relations; accuracy older adults: M = 0.98, SE = 0.00; younger adults: M = 0.97, SE = 0.00; t (70) = −1.77, P > 0.05; training trials required: M = 60 trials, SE = 0, for both groups). Critically, older adults also performed like younger adults for the pairwise relation condition on accuracy (older adults: M = 0.97, SE = 0.00; younger adults: M = 0.97, SE = 0.01; t (70) = 0.91, P > 0.05) and number of training trials required (older adults: M = 60, SE = 0.0; younger adults: M = 61, SE = 1.0; t (70) = −1.0, P > 0.05).
Figure 4.
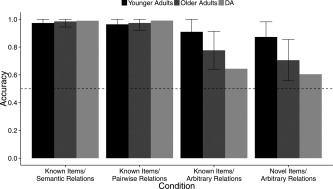
Accuracy in the training phase for younger adults, older adults and DA on the four conditions of transitivity that vary in the extent to which the items and the relations among them were known before the experiment. Younger adults showed high accuracy across all conditions. DAs performance was highly accurate only for the Known Items/Semantic Relations and the Known Items/Pairwise Relations conditions; his performance was low on the conditions that required learning of arbitrary. Older adults showed impairments relative to younger adults when arbitrary relations had to be learned. However, like DA, older adults showed performance comparable to that of younger adults on all trial types when arbitrary relations had to be acquired, but were presented in the context of known, pairwise relations (Known Items/Pairwise Relations). Dotted line here in all figures represents chance performance (accuracy = 0.50). Error bars here and in all figures represent the 95% confidence interval of the mean.
No‐Feedback Test Block
Effects of experiment version
The main effect of experiment version was not significant (F (2,48) = 2.71; P = 0.08). The main effect of first condition completed was also not significant (F (3,48) = 1.04; P > 0.05), as were all possible between‐subject interactions of experiment version, age group, and first condition completed.
Effects of known versus novel relations
As in the training blocks, the main effect of stimulus condition was significant (F (3,144) = 50.01, P < 0.001) in the no‐feedback test block; as expected, accuracy was higher for the conditions that used known items and semantic relations, and decreased when novel objects and arbitrary relations were leaned within the confines of the experiment (known objects/semantic relations: M = 0.97, SE = 0.01; known objects/pairwise relations: M = 0.90, SE = 0.01; known objects/arbitrary relations: M = 0.80; SE = 0.02; novel objects/arbitrary relations: M = 0.77, SE = 0.02).
As in the training blocks, the interaction of stimulus condition by first condition completed was significant in the no‐feedback test block (F (9,144) = 4.67, P < 0.001). The order of the conditions had the most dramatic effect on accuracy for the novel objects/arbitrary relations condition. When this condition was presented first, accuracy was low (M = 0.62; SE = 0.04), but if it followed any of the other conditions, accuracy was considerably higher (range = 0.80–0.83). Similarly, for the known objects/arbitrary relations condition, accuracy was the lowest when this condition was presented first (M = 0.73, SE = 0.05), but was higher when it followed any of the other conditions (range = 0.80–0.85). By contrast, when at least some of the relations were pre‐experimentally known, performance was high regardless of whether it was presented first (known objects/semantic relations: M = 0.97, SE = 0.01; known objects/pairwise relations: M = 0.88, SE = 0.03) or followed another condition (known objects/semantic relations: range = 0.96–0.98; known objects/pairwise relations: range = 0.87–0.92).
Trial type: Effects of relational distance
Increasing relational distance between the sample and choice stimuli resulted in decreasing accuracy, as revealed by a significant main effect of trial type (F (3,144) = 34.04, P < 0.001). Participants had the highest accuracy for the previously studied pairs (M = 0.90, SE = 0.01), and the lowest accuracy for two‐away pairs (M = 0.82; SE = 0.02).
The interaction of stimulus condition with trial type was significant (F (9,432) = 7.63; P < 0.001). The general pattern of highest accuracy for studied pairs and lowest accuracy for the two‐away pairs was found for each of the stimulus conditions; however, the range of accuracies across the trial types was exaggerated for the known objects/pairwise relations condition (range = 0.83–0.96). This finding may not be surprising as the studied pairs represented relations known before the experiment, whereas the other trial types (one‐away, two‐away, mixed) presented relations that had to be acquired during the confines of the experiment. The trial types in each of the other conditions presented relations of the same order: either all were pre‐experimentally known or all had to be acquired within the experimental session.
The interaction of condition, trial type, and experiment version was significant (F (18,432) = 1.82; P < 0.05). Again, performance tended to be most accurate for the previously studied pairs and least accurate for the two‐away pairs across stimulus conditions and experiment versions. Also, as noted above, the range of accuracies across trial types was most exaggerated for the known objects/pairwise relations condition in each experimental version and the range of accuracies smaller in the delay version of the task (range: 0.89–0.96) relative to the onset + delay version (range: 0.83–0.97) and the onset version of the task (range: 0.76–0.95). A similar change in the range of accuracies across trial types and the experimental versions was also observed in the known objects/novel relations condition, where the range of accuracies was smallest in the delay version (range: 0.86–0.91), followed by the onset + delay version (range: 0.76–0.84), and the onset version (range: 0.70–0.80).
Effects of age
All the younger and older adults achieved criterion during training and proceeded to the test blocks. In general, younger adults (M = 0.90; SE = 0.02) were more accurate than the older adults (M = 0.82, SE = 0.02), as revealed by a significant effect of age group (F (1,48) = 12.87; P = 0.001). The interaction of age group with stimulus condition was significant (F (1,144) = 12.69, P < 0.001; Fig. 5). Older adults were significantly less accurate than younger adults on arbitrary relations conditions (with known objects t (70) = 2.67, P < 0.05; with novel objects t (70) = 4.47, P < 0.001). As expected, the older adults' performance was similar to that of the younger adults for known objects/semantic relations condition (t (70) = −1.07, P > 0.05). Critically, older adults also performed like younger adults for the condition in which the pairwise relations were known before the experiment (t (70) = 1.03, P > 0.05).
Figure 5.

Accuracy in the test phase for younger adults, older adults, and DA on the four conditions of transitivity that vary in the extent to which the items and the relations among them were known before the experiment. Younger adults showed high accuracy across all conditions. DAs performance was highly accurate only for the Known Items/Semantic Relations condition; his performance was low on all other conditions which required arbitrary relations to be acquired within the experimental session. Older adults showed impairments relative to younger adults when arbitrary relations had to be learned. However, older adults showed performance comparable to that of younger adults on all trial types when arbitrary relations had to be acquired, but were presented in the context of known, pairwise relations (Known Items/Pairwise Relations).
This latter effect is not due merely to the inclusion of the previously studied trials. When accuracy for the younger and older adults was compared on the known objects/pairwise relations condition, omitting the studied trials from the analysis for which there was pre‐existing knowledge, there was still no significant difference between younger and older adults (t (70) = 1.13, P > 0.05). By contrast, without the inclusion of the studied trials, a significant difference remained between the older and younger adults on accuracy for the known objects/arbitrary relations condition (t (70) = 3.4, P < 0.01). Thus, successful performance on the novel probe trials in both conditions required that novel inferences be expressed between known objects, however, older adults only performed like younger adults when those novel inferences were expressed with respect to known (pairwise) relations. When novel inferences had to be expressed in the absence of an existing relational schema, accuracy for the older adults was significantly worse than that of younger adults.
Across conditions, performance for the older adults was less accurate than younger adults across trial types, with the largest difference in accuracy for the two‐away pairs, resulting in a significant interaction of trial type with age group (F (3,144) = 8.75, P < 0.001). Older adults were significantly less accurate than the younger adults for every type of trial except for the studied trials (studied: t (70) = 2.07, P > 0.05; one‐away: t (70) = 3.39, P < 0.01; two‐away: t (70) = 3.61, P < 0.01; mixed: t (30) = 3.98, P < 0.001).
The interaction of stimulus condition, trial type and age group was significant (F (9,432) = 2.45; P < 0.05; see Fig. 6). As noted above, typically, accuracy on the studied pairs was the highest and accuracy for the two‐away pairs was the lowest; this occurred for the older and younger adults regardless of stimulus condition. However, the range of accuracy across trial types was the greatest for both older adults (range: 0.79–0.96) and younger adults (range: 0.86–0.96) for the pairwise relations condition. The range of accuracies across trial types for any of the other stimulus conditions was fairly restricted for the younger adults (novel objects/arbitrary relations: 0.83–0.88; known objects/arbitrary relations: 0.85–0.87; known objects/semantic relations: 0.95–0.97), whereas for the older adults, the range of accuracies across trials types was restricted only for the known objects/semantic relations condition (range: 0.97–0.99; novel objects/arbitrary relations: 0.60–0.73; known objects/arbitrary relations: 0.67–0.83).
Figure 6.

Accuracy in the test phase for younger adults, older adults, and DA on the two conditions of transitivity involving known items where the inference trials contained arbitrary relations (left: Known Items/Pairwise Relations; right: Known Items/Arbitrary Relations) for each trial type (studied, one‐away, two‐away, mixed). Older adults are impaired relative to the younger adults on all trial types when arbitrary relations had to be acquired within the experimental session (right), except for when those relations are presented in the context of known pairwise relations (Studied pairs; Known Items/Pairwise Relations; left). In contrast, DA's performance is impaired in both conditions on all trial types involving arbitrary relations—he did not benefit from premise pairs supported by semantic knowledge.
Amnesic case DA
DA completed a total of nine sessions of testing. However, data were lost for the known objects/semantic relations condition for the first session, resulting in eight sessions of data for this condition. DA failed to achieve criterion (the experimental condition terminated following six repeated presentations of the training blocks in which accuracy was below 70%) for the known objects/arbitrary relations condition in three separate sessions; thus, the reported test block accuracy is for six sessions of data. Due to time constraints, DA was only able to complete the novel objects/arbitrary relations in six of the nine sessions as this condition was always given last. He failed to achieve criterion in three of those six sessions; therefore, the reported test block accuracy is for three sessions of data for the novel objects/arbitrary relations condition. Failure to reach criterion for both of the arbitrary relations conditions (with known or novel objects) occurred in only one session. Interestingly, the instances in which DA failed to achieve criterion occurred when the sample and choice stimuli were presented simultaneously during training rather than over a delay; however, test performance did not differ across experimental versions and therefore, test accuracy data for each condition were combined across experimental versions. DA's accuracy for each experimental condition in the training phase is plotted in Figure 4 and in the test phase is plotted in Figure 5. In each figure his performance is plotted alongside performance for the younger and older adults, and was contrasted against the 95% confidence interval (CI) of older adults as done in our previous work (Moses et al., 2008; Ryan et al., 2013). In the training phase, DA's performance was highly accurate for the known objects/semantic relations condition and the known objects/pairwise relations condition, and his performance was within the within the 95% CI for both younger and older adults for both conditions. DA's performance was well outside the 95% CI of both younger and older adults on the arbitrary relations conditions (known objects, novel objects). DA also required more trials during training in both conditions with arbitrary relations (see Fig. 4). In the test phase, DA's performance was highly accurate only for the known objects/semantic relations condition, and was within the within the 95% CI for both younger and older adults. DA's performance was well outside the 95% CI of both younger and older adults on the known objects/pairwise relations, and both conditions with arbitrary relations (known objects, novel objects).
DA's performance was also examined within each stimulus condition, for each type of trial. DA's performance was highly accurate, and within the 95% CI for younger and older adults, on all trial types of the known items/semantic relations condition. Within the known objects/pairwise relations condition, DA demonstrated intact performance (within the 95% CI of both the younger and older adults) on the studied pairs, for which he had pre‐existing knowledge, but he could not express novel inferences with respect to these known pairwise relations, as demonstrated by his poor performance on every other pair (Fig. 6). Likewise, DA performed outside of the 95% CI of younger and older adults for all trial types for the two arbitrary relations conditions (known objects; novel objects).
It should be noted that although DA exhibited poor performance in the conditions for which arbitrary relations were to be learned, following the experimental sessions, he was nonetheless able to appropriately name each of the known objects, and create names for the novel objects. Further, when presented with all of the items in a given experimental condition and asked to create two groups of items, he was able to create plausible stories which connected the objects (e.g., “the keys would get lost in the cushion of the chair”), although the groupings did not consistently match those provided within the experimental context.
DISCUSSION
By testing younger adults, older adults, and amnesic case DA on a variant of the transitivity task, originally developed for use with non‐human animals (Bunsey and Eichenbaum, 1996), we examined the extent to which existing knowledge could support the flexible expression of novel inference judgments. Findings from the current study demonstrate that while the processing of semantic relations is intact in aging, the acquisition of arbitrary relations, and by consequence, inferential judgments, is compromised (Ryan et al., 2009). Further, the age‐related deficit in the acquisition of arbitrary relations as observed here, was similar in nature to the impairment observed in amnesia, although not as severe. These age‐related impairments in inference were shown in a task in which performance could likely not be attributed to the use of a strategy that considers differences in the associative strength of individual items. Importantly, successful inference performance can occur by flexibly expressing knowledge among known relations (Castel, 2005; McGillivray and Castel, 2010; Moses et al., 2010a,b; Ostreicher et al., 2010). However, DA, who has both hippocampal and neocortical damage, could not successfully acquire arbitrary relations, and could not flexibly express previously stored relational, semantic, knowledge during inference judgments, suggesting that the flexible expression of relational knowledge critically requires contributions from hippocampal‐neocortical interactions (Tse et al., 2007, 2011).
In the training phase, younger adults, older adults, and DA performed similarly in the two conditions where prior knowledge could support performance (known objects/semantic relations and known objects/pairwise relations). In contrast, older adults and DA had lower accuracy and required more training trials to reach criteria when learning arbitrary relations among known or novel objects, with DA showing larger impairments than the older adults. Therefore, relative to younger adults, both older adults and DA were impaired when arbitrary relations had to be learned, but had intact performance in conditions where the expression of relations could be referenced to semantic information.
Accuracy for the younger adults in the test phase was high across all conditions, regardless of whether relations had to be learned among known or novel objects. Older adults were impaired relative to younger adults when arbitrary relations had to be learned (known items/arbitrary relations, novel items/arbitrary relations). While accuracy for the older adults on the novel items/arbitrary relations condition was better than that of DA, their performance nonetheless points to a deficit similar to that of DA, namely that older adults have significant difficulties in the learning and expression of arbitrary relations, particularly among novel objects. These age‐related deficits in relational memory are consistent with findings from prior studies (Naveh‐Benjamin, 2000; Ryan et al., 2007; Old and Naveh‐Benjamin, 2008; Ostreicher et al., 2010; Moses et al., 2010a,b) and demonstrate that impaired relational memory impacts inference performance (Ryan et al., 2009). These age‐related deficits on the transitivity task also mirror the transitivity performance observed in the senescence‐accelerated mouse model (SAMP8) (Ohta et al., 2002). Interestingly, age‐related differences in accuracy did not show a significant interaction with the timing procedures at training/test. This suggests that the critical element for performance on the transitivity task is not necessarily the bridging of a temporal gap, but rather the formation of the relations themselves and the ability to select the appropriate relation in the presence of a distracter.
Older adults could flexibly express relational knowledge when asked to make novel inference judgments (known items/pairwise relations; McGillivray and Castel, 2010; Moses et al., 2010a,b; Ostreicher et al., 2010; Crespo‐Garcia et al., 2012). Importantly, high performance by the older adults on the pairwise relations condition was not due solely to accuracy on the studied pairs for which participants had pre‐existing knowledge. Rather, older adults, like younger adults, were able to organize pre‐existing relational knowledge accordingly, resulting in accurate inference performance on the one‐away, two‐away, and mixed trials. This is critical because accurate performance on these problem sets required the expression of arbitrary relations, just as in the novel objects/arbitrary relations condition. In fact, when performance on the one‐away, two‐away, and mixed trials for the pairwise condition was contrasted with that on the arbitrary relations condition, it became clear that the use of pre‐existing knowledge could support novel inferences in older adults as accuracy was higher on all these problem pairs in the pairwise condition compared with the arbitrary relations condition.
Like the older adults, DA was significantly impaired whenever arbitrary relations had to be acquired for inference to occur. These findings suggest that the learning of relations, and consequently, inference performance, critically requires contributions from the hippocampus. Although DA's damage extends beyond the hippocampus into the cortices of the medial temporal lobe, when the present findings are considered in conjunction with previous work with nonhuman animals with lesions restricted to the hippocampus (Bunsey and Eichenbaum, 1996), the specific role of the hippocampus in the learning of new relations to support transitivity performance, in human and non‐human animals alike, becomes highlighted.
In contrast to the older adults, DA was unable to express flexible knowledge of relations that were based on prior knowledge. Flexible expression of relations in the pairwise condition may require storage of new relational knowledge representing the indirect relations (e.g., Tse et al., 2007); any residual hippocampal function in DA may not have been sufficient to support this function (Kumaran and McClelland, 2012). DA's damage to vmPFC and anterior‐lateral temporal neocortex likely contributed to his impaired expression of indirect relations on the pairwise relations condition. Findings from neuroimaging provide support for the role of the prefrontal cortex in relational processing (Wendelken and Bunge, 2010). Specifically, the vmPFC may integrate shared content among distinct events held in memory, ultimately supporting inference (Zeithamova et al., 2012). Further non‐human animal research from Tse et al. (2011) demonstrates the critical and equal role of the rat mPFC and hippocampus in the assimilation of new relations into previously stored knowledge (Tse et al., 2011), although the homolog to the rat mPFC is under debate (Kesner, 2000; Farovik et al.; 2008; for review see Uylings et al., 2003).
Although DA has damage to the hippocampus and to the vmPFC, he was able to express his pre‐existing knowledge regarding the relations among known stimuli. Successful relational processing in DA is consistent with our previous reports of intact performance by amnesic cases (including DA) on versions of the transverse patterning task that used relations learned by the amnesics before their neurological insult (Moses et al., 2008; Ryan et al., 2013). Such findings argue that the hippocampus is not necessary for the expression of relational knowledge, but that the hippocampus contributes to the initial binding of relations (Ryan et al., 2009).
It should be noted for some problems, particularly in the pairwise relations condition, DA exhibited performance that was considerably below chance levels. DA's choices were queried following the experimental sessions, and he would often group items into sets based on his prior experiences, such as pairing the wrench with the BBQ because he has used a wrench to fix his BBQ, or pairing the keys with the chair as his keys would often fall out of his pockets and get lost in the chair cushions. However, some of his groupings, such as the keys with the chair, would not have been possible to make during the experiment itself given the design (e.g., parallel stimulus items, B and X). However, although DA's strategic use of prior knowledge to support test performance and his stated groupings were typically consistent across experimental sessions, his post‐experiment explicit responses did not always map onto his task responses. While some older adults may have used a prior knowledge strategy as well to counteract declining relational memory, there was a subset of older adults (n=14) who scored perfectly on problems such as the two‐away trial of the known items/pairwise relations condition, suggesting that DA particularly relied on his prior knowledge of known relations to support performance due to his inability to learn arbitrary relations. Additional analyses (data not included) were conducted to determine if there was an effect of ordinal strength on the mixed trials within each condition. There was no evidence to indicate that ordinal position in the hierarchy biased performance on the mixed trials for DA, the older adults or the younger adults, suggesting that memory “strength” was not generally used as a strategy to support performance, in contrast to the use of prior knowledge.
The order of the conditions may have particularly biased DA to rely on his prior knowledge. DA was always given the known items/semantic relations condition first, similar to our prior work with transverse patterning (Ryan et al., 2013), in an effort to ensure he understood the task demands. The order of the conditions had a significant influence on performance for the younger and the older adults; performance on the arbitrary relations conditions was more accurate if participants were first presented with the semantic relations condition, suggesting that participants were likely applying the relevant schemas across stimulus conditions to aid performance. We have also observed this influence of prior knowledge on subsequent learning of arbitrary relations in our prior work with older adults on the transverse patterning task (Ostreicher et al., 2010). However, whereas older adults were able to use prior knowledge and schemas to boost learning and inference of novel relations within and across conditions, DA did not show such benefits despite always receiving the conditions employing prior knowledge first during the experimental session.
Intact use of semantic relations in DA nonetheless contrasts with findings regarding individuals with vmPFC lesions who were selectively impaired on inference itself despite intact learning of novel premise statements (Koscik and Tranel, 2012). However, unlike the current study, inferences were to be made on recently acquired premise relationships rather than on relations acquired before the individuals' neurological insult. Thus, humans with vmPFC lesions could be impaired on inference for arbitrary (hippocampal‐dependent) relations, but show intact performance for already consolidated (hippocampal‐independent) relations. Previously acquired knowledge may be more likely to be accessed and represented through ventrolateral PFC along with the anterior temporal lobe than through the vmPFC. In fact, our prior magnetoencephalography work (Moses et al., 2009) has shown a shift in the relative contributions from the hippocampal versus anterior‐lateral temporal and ventrolateral frontal systems that were engaged to support transverse patterning when the relations to be processed were either acquired recently (i.e., more hippocampus, less anterior‐lateral temporal and frontal) or long before the experimental session (i.e., less hippocampus, more anterior temporal and frontal; see also Hanlon et al., 2005; Doeller et al., 2005). Whereas the acquisition of arbitrary relations requires the hippocampus, the encoding of information that is consistent with prior knowledge, as well as the retrieval and processing of previously stored relations may be facilitated by anterior temporal‐ventral frontal interactions (Van Kesteren et al., 2013).
The flexible expression of arbitrary, indirect, relations as based on prior knowledge may occur through continual hippocampal‐neocortical functional interactions, which then further serve to update the knowledge schema and stored memory representations (Wang and Morris, 2010; McKenzie and Eichenbaum, 2011). In DA, damage to the vmPFC, anterior temporal lobe and/or the hippocampus impaired knowledge of the novel, indirect, relations that were based on sets of previously known relations. It remains for future work to determine if amnesic cases who have some residual function in the hippocampus and an intact frontal cortex and anterior temporal lobe could successfully perform novel inference judgments that were predicated on semantic relations. Recent findings indicate that amnesic cases who have damage restricted to the medial temporal lobe, with sparing of the anterior and lateral temporal lobes, show benefits for acquiring novel information that is congruent versus incongruent with existing knowledge schemas (Kan et al., 2009), and such learning is likely mediated by the medial prefrontal system (Van Kesteren et al., 2010, 2012).
Interestingly, DA fails here on the transitivity paradigm when arbitrary relations must be acquired and/or expressed, but he has demonstrated uncharacteristically intact performance for an amnesic person on the transverse patterning task which similarly requires arbitrary relations to be acquired (Ryan et al., 2013). We have argued that DA's intact performance on transverse patterning was likely due to the use of a unitization strategy in which he generates a scenario involving the physical integration of single objects. When DA did not attempt such integration, transverse patterning performance was impaired. In the present investigation, it was initially expected that DA would demonstrate relative success on versions of the experiment in which the sample and choice stimuli were presented simultaneously rather than across a delay, as simultaneous presentation of the stimuli may have allowed the opportunity for unitization to occur. However, inspection of the data revealed that DA's performance did not vary across experimental versions; under no conditions was DA successfully able to acquire the arbitrary relations needed to support transitivity. Ultimately, unitization may have been difficult to employ in the current task, as during all problems, the correct choice was presented alongside the incorrect choice. Thus, any integration would have required DA to evaluate and exclude one of the presented objects before creating a unitized image, which may have been difficult and/or required additional processing time that would have impacted the online maintenance of the relations.
Taken together, the current findings reveal that aging is associated with a decline in establishing arbitrary relations, likely as a result of declining hippocampal function, that would otherwise support the expression of novel inferences (Ryan et al., 2009). However, existing relational knowledge can be used flexibly by older adults to make novel inference judgments. By contrast, amnesic case DA could not make use of prior knowledge to support novel inferences, likely due to damage to vmPFC and anterior temporal cortex as well as the hippocampus. This work, when considered in conjunction with findings from non‐human animal research (Tse et al., 2007, 2011), highlights the importance of ventral frontal‐temporal‐hippocampal interactions in the ongoing learning and flexible expression of relations.
Acknowledgments
The authors acknowledge DA and his family for his continued involvement in research. The authors also thank Arber Kacollja for his assistance in data collection and Rosanna Olsen for her helpful comments on an earlier draft of this article.
REFERENCES
- Bangasser DA, Waxler DE, Santollo J, Shors TJ. 2006. Trace conditioning and the hippocampus: The importance of contiguity. J Neurosci 26:8702–8706. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bartlett, F.C . (1932). Remembering: A Study in Experimental and Social Psychology. Cambridge, UK: Cambridge University Press. [Google Scholar]
- Bastin C, Van der Linden M. 2006. The effects of aging on recognition of different types of associations. Exp Aging Res 32:61–77. [DOI] [PubMed] [Google Scholar]
- Bunsey M, Eichenbaum H. 1996. Conservation of hippocampal memory function in rats and humans. Nature 379:255–257. [DOI] [PubMed] [Google Scholar]
- Castel AD. 2005. Memory for grocery prices in younger and older adults: The role of schematic support. Psychol Aging 20:718–21. [DOI] [PubMed] [Google Scholar]
- Castel AD, Craik FIM. 2003. The effects of aging and divided attention on memory for item and associative information. Psychol Aging 18:873–885. [DOI] [PubMed] [Google Scholar]
- Crespo‐Garcia, M , Cantero. JL , M Atienza. (2012). Effects of semantic relatedness on age‐related associative deficits: the role of theta oscillations. Neuroimage, 61(4), 1235–1248. [DOI] [PubMed] [Google Scholar]
- Daselaar SM, Fleck MS, Dobbins IG, Madden DJ, Cabeza R. 2006. Effects of healthy aging on hippocampal and rhinal memory functions: an event‐related fMRI study. Cereb Cortex 16:1771–1782. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doeller CF, Opitz B, Krick CM, Mecklinger A, Reith W. 2005. Prefrontal hippocampal dynamics involved in learning regularities across episodes. Cereb Cortex 15:1123–1133. [DOI] [PubMed] [Google Scholar]
- Driscoll I, Hamilton DA, Petropoulos H, Yeo RA, Brooks WM, Baumgartner RN, Sutherland RJ. 2003. The aging hippocampus: cognitive, biochemical and structural findings. Cereb Cortex 13:1344–1351. [DOI] [PubMed] [Google Scholar]
- Farovik A, Dupont LM, Arce M, Eichenbaum H. 2008. Medial prefrontal cortex supports recollection, but not familiarity, in the rat. J Neurosci 28:13428–13434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gordon BA, Blazey T, Benzinger TL, Head D. 2013. Effects of aging and Alzheimer's disease along the longitudinal axis of the hippocampus. J Alzheimer's Dis 37:41–50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grady CL, McIntosh AR, Craik FIM. 2003. Age‐related differences in the functional connectivity of the hippocampus during memory encoding. Hippocampus 13:572–586. [DOI] [PubMed] [Google Scholar]
- Greene AJ, Gross WL, Elsinger CL, Rao SM. 2006. An FMRI analysis of the human hippocampus: Inference, context, and task awareness. J Cognit Neurosci 18:1156–1173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hanlon FM, Weisend MP, Yeo RA, Huang M, Lee RR, Thoma RJ, Moses SN, Paulson KM, Petropoulos H, Miller GA, Cañive JM. 2005. A specific test of hippocampal deficit in schizophrenia. Behav Neurosci 119:863–875. [DOI] [PubMed] [Google Scholar]
- Healy MR, Light LL, Chung C. 2005. Dual‐process models of associative recognition in young and older adults: Evidence from receiver operating characteristics. J Exp Psychol 31:768–788. [DOI] [PubMed] [Google Scholar]
- Heckers S, Zalesak M, Weiss AP, Ditman T, Titone D. 2004. Hippocampal activation during transitive inference in humans. Hippocampus 14:153–162. [DOI] [PubMed] [Google Scholar]
- Holyoak KJ. (2012). Analogy and relational reasoning In: The Oxford Handbook of Thinking and Reasoning. New York, NY: Oxford University Press; pp 234–259. [Google Scholar]
- Hoscheidt SM, Nadel L, Payne J, Ryan L. 2010. Hippocampal activation during retrieval of spatial context from episodic and semantic memory. Behav Brain Res 212:121–132. [DOI] [PubMed] [Google Scholar]
- Kan IP, Alexander MP, Verfaellie M. 2009. Contribution of prior semantic knowledge to new episodic learning in amnesia. J Cognit Neurosci 938–944. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kesner RP. 2000. Subregional analysis of mnemonic functions of the prefrontal cortex in the rat. Psychobiology 28:219–228. [Google Scholar]
- Konkel A, Warren DE, Duff MC, Tranel DN, Cohen NJ. 2008. Hippocampal amnesia impairs all manner of relational memory. Front Hum Neurosci, 2:15. doi: 10.3389/neuro.09.015.2008. eCollection 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koscik TR, Tranel D. 2012. The human ventromedial prefrontal cortex is critical for transitive inference. J Cognit Neurosci 24:1191–1204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kumaran D, McClelland JL. 2012. Generalization through the recurrent interaction of episodic memories: A model of the hippocampal system. Psychol Rev 119:573–616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McGillivray S, Castel AD. 2010. Memory for age‐face associations in younger and older adults: The role of generation and schematic support. Psychol Aging 25:822–832. [DOI] [PubMed] [Google Scholar]
- McKenzie S, Eichenbaum H. 2011. Consolidation and reconsolidation: Two lives of memories? Neuron 71:224–233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morrison JH, Baxter MG. 2012. The ageing cortical synapse: Hallmarks and implications for cognitive decline. Nat Rev Neurosci 13:240–250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moscovitch M, Nadel L, Winocur G, Gilboa A, Rosenbaum RS. 2006. The cognitive neuroscience of remote episodic, semantic and spatial memory. Curr Opin Neurobiol 16:179–190. [DOI] [PubMed] [Google Scholar]
- Moses SN, Villate C, Ryan JD. 2006. An investigation of learning strategy supporting transitive inference performance in humans. Neuropsychologia 44:1370–1387. [DOI] [PubMed] [Google Scholar]
- Moses SN, Ostreicher M, Rosenbaum RS, Ryan JD. 2008. Successful transverse patterning in amnesia using semantic knowledge. Hippocampus 18:121–124. [DOI] [PubMed] [Google Scholar]
- Moses SN, Ryan JD, Bardouille T, Kovacevic N, Hanlon FM, McIntosh AR. 2009. Semantic information alters neural activation during transverse patterning performance. Neuroimage 46:863–873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moses SN, Brown T, Ryan JD, McIntosh AR. 2010a. Neural system interactions underlying human transitive inference. Hippocampus 8:894–901. [DOI] [PubMed] [Google Scholar]
- Moses SN, Ostreicher M, Ryan JD. 2010b. Relational strategy bias improves transitive inference performance. Psychol Res 74:207–218. [DOI] [PubMed] [Google Scholar]
- Naveh‐Benjamin M. 2000. Adult age differences in memory performance: Tests of an associative deficit hypothesis. J Exp Psychol 26:1170–1187. [DOI] [PubMed] [Google Scholar]
- Ohta A, Akiguchi I, Seriu N, Ohnishi K, Yagi H, Higuchi K, Hosokawa M. 2002. Deterioration in learning and memory of inferential tasks for evaluation of transitivity and symmetry in aged SAMP8 mice. Hippocampus 12:803–810. [DOI] [PubMed] [Google Scholar]
- Old SR, Naveh‐Benjamin M. 2008. Differential effects of age on item and associative measures of memory: A meta‐analysis. Psychol Aging 23:104–118. [DOI] [PubMed] [Google Scholar]
- Ostreicher M, Moses SN, Rosenbaum RS, Ryan JD. 2010. Prior experience supports new learning of relations in aging. J Gerontol 65:32–41. [DOI] [PubMed] [Google Scholar]
- Rosenbaum RS, Moscovitch M, Foster JK, Schnyer DM, Gao F, Kovacevic N, Verfaellie M, Black SE, Levine B. 2008. Patterns of autobiographical memory loss in medial‐temporal lobe amnesic patients. J Cognit Neurosci 20:1490–1506. [DOI] [PubMed] [Google Scholar]
- Ryan JD, Leung GL, Turk‐Browne NB, Hasher L. 2007. Assessment of age‐related changes in inhibition and relational memory binding using eye movement monitoring. Psychol Aging 22:239–250. [DOI] [PubMed] [Google Scholar]
- Ryan JD, Moses SN, Villate C. 2009. Impaired relational organization of propositions, but intact transitive inference, in aging: implications for understanding underlying neural integrity. Neuropsychologia 47:338–353. [DOI] [PubMed] [Google Scholar]
- Ryan L, Lin CY, Ketcham K, Nadel L. 2010. The role of medial temporal lobe in retrieving spatial and nonspatial relations from episodic and semantic memory. Hippocampus 20:11–18. [DOI] [PubMed] [Google Scholar]
- Ryan JD, Moses SN, Barense M, Rosenbaum RS. 2013. Intact learning of new relations in amnesia as achieved through unitization. J Neurosci 33:9601–9613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scahill RI, Frost C, Jenkins R, Whitwell JL, Rossor MN, Fox NC. 2003. A longitudinal study of brain volume changes in normal aging using a serial registered magnetic resonance imaging. Arch Neurol 60:989–994. [DOI] [PubMed] [Google Scholar]
- Smith C, Squire LR. 2005. Declarative memory, awareness, and transitive inference. J Neurosci 25:10138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tse D, Langston RF, Kakeyama M, Bethus I, Spooner PA, Wood ER, Witter MP, Morris RG. 2007. Schemas and memory consolidation. Science 316:76–82. [DOI] [PubMed] [Google Scholar]
- Tse D, Takeuchi T, Kakeyama M, Kajii Y, Okuno H, Tohyama C, Bito H, Morris RG. 2011. Schema‐dependent gene activation and memory encoding in neocortex. Science 333:891–895. [DOI] [PubMed] [Google Scholar]
- Tsukiura T, Sekiguchi A, Yomogida Y, Nakagawa Shigemune Y, Kambara T, Kawashima R. 2011. Effects of aging on hippocampal and anterior temporal activations during successful retrieval of memory for face‐name associations. J Cognit Neurosci 23:200–213. [DOI] [PubMed] [Google Scholar]
- Uylings HBM, Groenewegen HJ, Kolb B. 2003. Do rats have a prefrontal cortex? Behav Brain Res 146:3–17. [DOI] [PubMed] [Google Scholar]
- Van Kesteren MT, Rijokema M, Ruiter DJ, Fernandez G. 2010. Retrieval of associative information congruent with prior knowledge is related to increased medial prefrontal activity and connectivity. Journal of Neuroscience, 30(47), 15888–15894. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Kesteren MTR, Ruiter DJ, Fernandez G, Henson RN. 2012. How schema and novelty augment memory formation. Trends Neurosci 35:211–219. [DOI] [PubMed] [Google Scholar]
- Van Kesteren MTR, Beul SF, Takashima A, Henson RN, Ruiter DJ, Fernandez G. 2013. Differential roles for medial prefrontal and medial temporal cortices in schema‐dependent encoding: From congruent to incongruent. Neuropsychologia 51:2352–2359. [DOI] [PubMed] [Google Scholar]
- Wallenstein GV, Eichenbaum H, Hasslemo ME. 1998. The hippocampus as an associator of discontiguous events. Trends Neurosci 21:317–323. [DOI] [PubMed] [Google Scholar]
- Wang S‐H, Morris RGM. 2010. Hippocampal‐neocortical interactions in memory formation, consolidation and reconsolidation. Annu Rev Psychol 61:49–79. [DOI] [PubMed] [Google Scholar]
- Wang S‐H, Tse D, Morris RGM. 2012. Anterior cingulate cortex in schema assimilation and expression. Learn Mem 19:315–318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wendelken C, Bunge SA. 2010. Transitive inference: distinct contributions of rostrolateral prefrontal cortex and the hippocampus. J Cognit Neurosci 22:837–847. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Westmacott R, Moscovitch M. 2002. Temporally‐graded retrograde memory loss for famous names and vocabulary terms in amnesia and semantic dementia: Further evidence for opposite gradients using implicit memory tests. Cognit Neuropsychol 19:135–163. [DOI] [PubMed] [Google Scholar]
- Zalesak M, Heckers S. 2009. The role of the hippocampus in transitive inference. Psychiatry Res 172:24–30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zeithamova D, Dominick AL, Preston AR. 2012. Hippocampal and ventral medial prefrontal activation during retrieval‐mediated learning supports novel inference. Neuron 75:168–179. [DOI] [PMC free article] [PubMed] [Google Scholar]