

Abstract
Background
Corynebacterium pseudotuberculosis is the etiologic agent of caseous lymphadenitis (CL), a chronic disease that affects goats and sheep. CL is characterized by the formation of granulomas in lymph nodes and other organs, such as the lungs and liver. Current knowledge of CL pathogenesis indicates that the induction of humoral and cellular immune responses are fundamental to disease control. The aim of this study was to evaluate the humoral and cellular immune responses in BALB/c mice inoculated with a C. pseudotuberculosis strain isolated in the state of Bahia, Brazil.
Results
The lymphocyte proliferation and in vitro production of IFN-γ, IL-4, IL-10, IL-12 and nitric oxide by spleen cells stimulated with secreted and somatic antigens from the studied strain were evaluated. IgG subclasses were also analyzed. Results showed a significant increase of Th1-profile cytokines after 60 days post-inoculation, as well as an important humoral response, represented by high levels of IgG2a and IgG1 against C. pseudotuberculosis.
Conclusion
The T1 strain of C. pseudotuberculosis was shown to induce humoral and cellular immune responses in BALB/c mice, but, even at a dosage of 1x107 CFU, no signs of the disease were observed.
Keywords: Corynebacterium pseudotuberculosis, Cytokines, BALB/c, IgG isotypes
Background
Caseous lymphadenitis (CL) is a chronic disease caused by Corynebacterium pseudotuberculosis that mainly affects small ruminants. Despite the economic [1, 2] and zoonotic [3] relevance of CL, a satisfactory vaccine model has not been developed [4, 5].
C. pseudotuberculosis is a facultative intracellular pathogen that can persist inside macrophages and stimulate the formation of granulomas [6, 7]. This species is distributed worldwide, but has the most severe economic impacts in Oceania, Africa and South America [8].
The pathogenesis of CL in mice was demonstrated by monitoring the progression of lesions in the skin and viscera of infected animals [9]. Moreover, the physiology, pathogenicity and virulence mechanisms of C. pseudotuberculosis strains have been elucidated using genomics [8, 10], transcriptomics and proteomics methodologies [11, 12].
The immune response against C. pseudotuberculosis has a well-known humoral component and involves a complex cellular mechanism against secreted and somatic bacterial antigens [13–15].
The cytokines Tumor necrosis factor-α (TNF-α) and interferon-γ (IFN-γ) are important to mount an immune response in mice as well as sheep, whether naturally infected or inoculated with C. pseudotuberculosis [16–19]. It is known that, with respect to Mycobacterium tuberculosis, a microorganism largely phylogenetically similar to C. pseudotuberculosis, these cytokines play a major role in susceptibility and regulation of associated lesions in mice [20].
El-Enbaawy et al. (2005) [21] demonstrated that antigens obtained from a C. pseudotuberculosis strain isolated from a naturally infected sheep, specifically a toxoid associated with bacterin, induce the production of IFN-γ, as well as elicit a humoral immune response in BALB/c mice. The present study employed a naturally attenuated strain of C. pseudotuberculosis, denominated T1, isolated from a granuloma taken from a goat in a rural region of the state of Bahia, located in northeastern Brazil. Studies previously conducted with this strain show that it grows quickly in BHI broth medium, when compared to other strains, but is incapable of inducing disease in goats [22–24].
The present study characterized the immune response in BALB/c mice, considering five animals per group, against antigens derived from the T1 strain of C. pseudotuberculosis. This murine model was chosen because of impaired IFN-γ production in response to antigens derived from M. tuberculosis, which is very closely related to C. pseudotuberculosis [25]. The proliferation of spleen cells was investigated, as well as the production of cytokines, nitric oxide (NO) and serum IgG subclasses to expand the understanding of humoral and cellular immune responses against this strain, which may represent an ideal vaccine candidate against this disease.
Results
To determine the optimal inoculation dosage, four different infection dosages (5x105, 1x106, 5x106 and 1x107 CFU) of the T1 C. pseudotuberculosis strain were tested in BALB/c mice. ELISA results showed higher IgG levels in mice infected with the two higher dosages in comparison to the two lower levels tested (P < 0.001) (Fig. 1a). No significant differences in IgG levels were seen between the groups inoculated with 5x106 and 1x107 CFU, nor in the groups inoculated with 5x105 and 1x106 CFU. At 120 days post-infection, none of the animals presented any evidence of lesions characteristic of the disease under clinical examination or necropsy. Because the 1x107 CFU dosage was not observed to induce lesions, this experimental protocol was used to evaluate the production of IgG subclasses and cytokines.
Fig. 1.
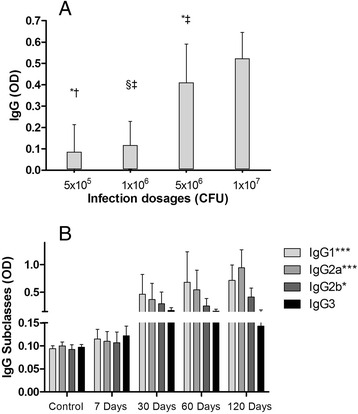
Serum IgG immune response in mice inoculated with T1 C. pseudotuberculosis strain, as evaluated by ELISA. Graph represents means of Optical Density (OD) values found for each group (n = 5 animals for group). Results are representative of the mean values obtained from two experiments. a. BALB/c mice were inoculated with increasing dosages: 5x105, 1x106, 5x106 and 1x107 CFU. Blood was collected 120 days after inoculation. Data were analyzed by ANOVA and Tukey post-hoc tests; *, †, ‡ and § indicate pairs with statistically significant differences. b. IgG subclass (IgG1, IgG2a, IgG2b and IgG3) production throughout the course of the experiment: control (before infection), 7, 30, 60, and 120 days after infection. Mice were inoculated with 107 CFU of T1 strain of C. pseudotuberculosis. Data were analyzed by ANOVA. *P < 0.05; ***P < 0.001
Analysis of the humoral immune response against T1 C. pseudotuberculosis revealed that IgG2a production gradually increased over time, being the predominant IgG subclass at 120 days after infection (P < 0.001). A significant increase in IgG1 levels (P < 0.001) was also observed, and a discrete, yet still statistically significant, increase of IgG2b (P < 0.05) was seen. No statistically significant differences in IgG3 levels were detected over the course of experimentation. Control group results are representative of the mean OD readings obtained from five animals before infection (Fig. 1b).
With respect to spleen cell response to antigenic stimuli, a significant lymphoproliferative response, expressed as SI, was observed after stimulation with secreted antigen (Se) (p < 0.05) at 60 days post-infection in comparison to 7 and 30 days (Fig. 2). Stimulation with Se provoked a significant difference in SI in comparison to So at 60 days after inoculation.
Fig. 2.
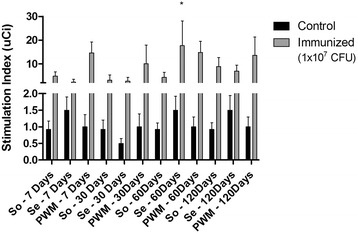
Proliferation of murine spleen cells stimulated with somatic (So) and secreted (Se) antigens. Results express the stimulation index (μCi) calculated from two independent experiments using splenocytes retrieved from five non-infected (control) and five infected (inoculated) animals from each group. Data were analyzed by ANOVA and Tukey post-hoc tests; *P < 0.05
In vitro production of interleukin-12 (IL-12) by spleen cells after stimulation with So or Se antigens is shown in Fig. 3a. Cell stimulated with both antigens had higher IL-12 concentrations at 60 and 120 days post-infection in comparison to controls (p < 0.05).
Fig. 3.
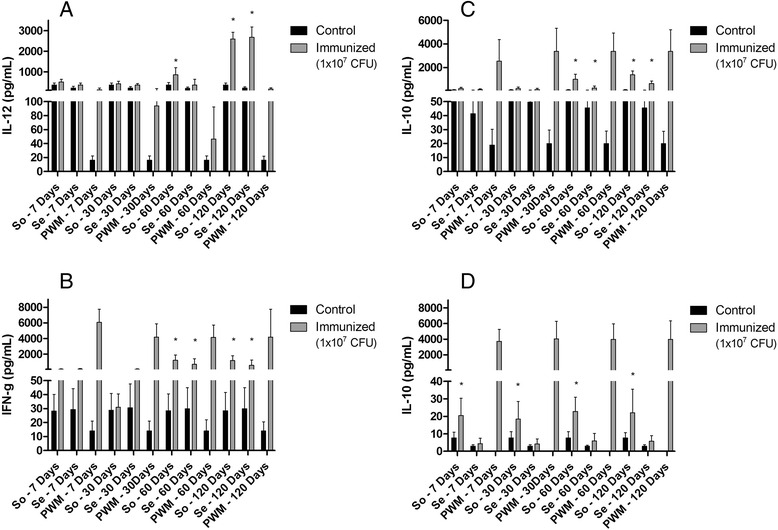
In vitro cytokine production by murine spleen cells stimulated with somatic (So) and secreted (Se) antigens. a. Interleukin-12 (IL-12). b. Interferon-γ (IFN-γ). c. Interleukin-10 (IL-10). d. Interleukin-4 (IL-4). Results are presented as ρg/mL, and represent the means of two independent experiments using spleen cells retrieved from five non-infected (control) and five infected (inoculated) animals from each group. Data were analyzed by ANOVA and Tukey post-hoc tests; *P < 0.05
No differences were seen in IFN-γ concentration in the antigen-stimulated culture supernatants in comparison to controls at seven and 30 days post-infection, but there significant increases were observed at 60 days (p < 0.05) and 120 days (p < 0.05) post infection. So also induced a higher and statistical significant INF-γ production, when compared to Se stimulation (Fig. 3b) at both of these times points.
With respect to in vitro interleukin-10 (IL-10) production, a significant statistical difference was observed in So-stimulated cells at 60 days post-inoculation (p < 0,05) in comparison to the previous infection times, and also in comparison to cells stimulated with Se at this same time point. A similar situation was observed at 120 days after inoculation (Fig. 3c). In addition, cells stimulated with So induced higher levels of IL-10 than Se throughout the experiment.
Interleukin-4 (IL-4) concentrations were very low at all experiment times evaluated with respect to both antigens. However, IL-4 production by cells stimulated with So was observed to significantly increase throughout the course of investigation (p < 0.05), but decreased at the 120 day time point (Fig. 3d).
Nitric oxide (NO) production measured in the supernatant of cell cultures stimulated So and Se is illustrated in Fig. 4. A significant increase (p < 0.05) in NO levels was seen only at 120 days post-infection in comparison to controls.
Fig. 4.
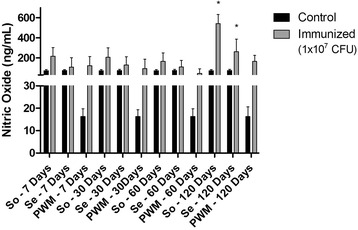
In vitro production of Nitric Oxide (NO) by murine spleen cells stimulated with somatic (So) and secreted (Se) antigens. Results are presented as ng/mL, and represent the means of two independent experiments using spleen cells retrieved from five non-infected (control) and five infected (inoculated) animals from each group. Data were analyzed by ANOVA and Tukey post-hoc tests; *P < 0.05
NO production by cells stimulated with So was also observed to be higher in comparison to Se, with statistical significance (p < 0.05) at 120 days after inoculation.
Discussion
The present study found that experimental inoculations of the attenuated T1 strain of C. pseudotuberculosis at a dosage of 107 CFU did not result in lesions in BALB/c mice, even though these animals have demonstrated susceptibility to intracellular pathogens [26, 27]. Nevertheless, a previous study has shown that a wild-type strain of C. pseudotuberculosis was able to induce lesions at a dosage of 102 [15].
In addition, cell cultures stimulated with T1 strain antigens were found to induce a proliferation of spleen cells, with secreted antigens (Se) demonstrating greater effectiveness than somatic antigens (So) two months after inoculation. A previous study showed that Se was able to enhance lymphocyte proliferation in PBMCs of an experimentally infected goat [23], which is consistent with our results. Se was found to induce a more intense proliferation than So due to the presence of phospholipase D, an exotoxin secreted by C. pseudotuberculosis at the beginning of infection to cleave the host cell membrane [28], which may cause a preeminent proliferation of B lymphocytes and elicit antibody production. Notably, lymphoproliferation in a murine model after stimulus with C. pseudotuberculosis antigens has not been described in the literature to date.
Experimental inoculation with T1 was observed to elicit high titers of IgG antibodies. The main IgG subclasses produced throughout the course of infection were IgG1 and IgG2a. As C. pseudotuberculosis is an intracellular pathogen that produces phospholipase D, an exotoxin with highly immunogenic properties [29, 30], the production of specific immunoglobulins is crucial to neutralize phospholipase D.
In addition, the cellular immune response is another way of reducing the dissemination of the pathogen, which can survive and multiply inside macrophages [7, 14, 18]. Accordingly, we found a significant production of IFN-γ by spleen cells after stimulation with So or Se C. pseudotuberculosis antigens two months after inoculation. This situation was sustained until the end of the experiment (120 days). Elevated IL-4 production was not detected, yet, in cells stimulated with So, the production of this cytokine was four times higher in comparison to those stimulated with Se and controls. This phenomenon may possibly have occurred because So has a larger amount of structural proteins and lipid antigens than Se [31]. Relatedly, Lan et al. (1999) [19] found a pronounced increase in IFN-γ production starting in the third week post-inoculation in splenic cell cultures of ICR-JCL mice inoculated with ATCC 1940 strain and stimulated with formalin-killed bacterial cells, which was sustained until the eighth week. In the same experiment, no significant production of IL-4 was observed.
IL-10 and IL-12 production by spleen cells stimulated by Se or So antigens increased post-inoculation time and was sustained at all time points evaluated. So was found to induce higher levels of IL-10 than Se, probably due the structural components of So, such as cytoplasm and membrane lipoproteins [31].
IL-10 may control IFN-γ synthesis during infection, thereby avoiding Th1 over-reactivity [32]. On the other hand, IL-12 can also trigger mechanisms related to cell proliferation and IFN-γ production [33]. Some studies have showed that IFN-γ, IL-10 and IL-12 are required to control persistent infections caused by intracellular parasites [34–36]. IL-12 is a cytokine crucial to Th1 shift, which is required to prevent the dissemination of pathogens within the host in order to control infection by facultative intracellular bacteria, such as C. pseudotuberculosis [14]. Accordingly, we found increased IL-12 production after 60 days, probably resulting from an immune response to reduce bacterial proliferation. Higher levels of IL-12 were detected at 120 days for both So and Se antigens, probably due to the persistence of infection. It is possible that, after this time, these levels would decrease as a result of IL-10 production.
NO production was also evaluated, due to its effectiveness in regulating the growth of intracellular pathogens [37]. Proteomic analysis has identified NO-responsive extracellular proteins of C. pseudotuberculosis and it also demonstrated the participation of the extracytoplasmic function sigma factor σE in composition of C. pseudotuberculosis exoproteome [38]. In the present studt, while NO production by spleen cells stimulated with So and Se antigens was higher at 120 days post-infection, So resulted in higher proliferations than Se, in accordance to what was observed in IFN-γ and IL-10 production.
Conclusion
The attenuated T1 strain of C. pseudotuberculosis was found to induce both humoral and cellular immune responses in an experimental model of susceptible BALB/c mice. A 107 CFU dosage did not result in any lesions in the mice evaluated. As the present study has demonstrated that, in addition to the production of antibodies, an efficient cellular response is important to the control of CL, the T1 strain can be considered as a promising option for potential vaccine candidates.
Methods
Bacterial strain
The T1 strain of C. pseudotuberculosis was isolated from granulomas obtained from goats raised in the municipality of Juazeiro, located in the state of Bahia in northeastern Brazil. Isolates were stored in the Department of Microbiology collection center at the Health Sciences Institute of the Federal University of Bahia (ICS - UFBA).
The identification of the T1 strain was confirmed by several microbiological methods: Gram staining, colony morphology, synergistic hemolytic (CAMP) reactions with Rhodococcus equi, urease and catalase production, as well as glucose and maltose fermentation. A commercial kit was used to aid in identification (API Coryne - BioMérieux, Merci l’Etoile, France). Since the T1 strain demonstrated a less severe pattern of hemolysis during synergistic hemolysis testing in comparison to other wild strains, other authors have suggested its use as a vaccinal strain [39].
The T1 strain was cultivated in Brain/Heart Infusion (BHI) broth and incubated for 72 h at 37 °C. The bacterial suspension was washed in Phosphate Buffer Saline (PBS) and centrifuged for 30 min at 3,000 g at 4 ° C.
Somatic antigen (So)
The bacterial pellet was homogenized in PBS (pH 7.4) and sonicated at 60 Hz under 4 °C for five cycles lasting 60 s each (Branson Sonifier 450, Branson, Dunbury, CT, USA). The sample was centrifuged for 30 min at 10,000 g and, after collection, the supernatant was stored at -20 °C in aliquots until use. Protein concentration was determined by Lowry’s modified method using a Bio-Rad Protein Assay (Bio-Rad, Hercules, CA, USA).
Secreted antigen (Se)
Se was obtained from the culture supernatant by saturation with 30 % ammonium sulfate (HCl) pH 4.0 and n-butanol under slow agitation at room temperature. The sample was homogenized, left undisturbed for 60 min, and then centrifuged for 10 min at 1,350 g under 4 ° C. The resulting interphase was dissolved in 20 mM of Tris buffer pH 7.4 (500 μL of buffer to 5 mL of culture supernatant), followed by dialysis in 50 mM Phosphate buffer pH 7.4 for 48 h. The sample was concentrated by ultra-filtration with a 10 kDa membrane (Millipore, Billerica, MA, USA). Protein concentration was determined by Lowry’s modified method using a Bio-Rad Protein Assay (Bio-Rad, Hercules, CA, USA).
Inoculation protocol and experimental design
Prior to experimental inoculation, an optimal inoculation dose experiment was performed to obtain maximum antibody production. Eight-week-old male and female BALB/c mice, provided by the Experimental Animal Facility at the Gonçalo Moniz Research Center, Oswaldo Cruz Foundation, Salvador, Bahia-Brazil, were used to establish the inoculation protocol. The optimal dose was determined using five groups of five mice. Four groups received an intraperitoneal inoculation of 5x105,106, 5x106 and 107 colony forming units (CFU) of C. pseudotuberculosis T1 strain diluted in sterile PBS at a final volume of 1 mL. The control group received 1 mL of sterile PBS by intraperitoneal inoculation. Blood was collected from the tail vein and the animals used for dosage experimentation were euthanized in a CO2 chamber. ELISA was performed 120 days after inoculation to evaluate humoral immune response by identifying the highest levels of IgG and its subclasses.
After determining the optimal inoculation dosage, male and female BALB/c mice received intraperitoneal inoculations with 107 CFU/mL of T1 strain in 1 mL of sterile PBS, while the control group was inoculated with 1 mL of sterile PBS. After blood sampling from the tail vein, five animals from each group were euthanized in a CO2 chamber at 7, 30, 60 and 120 days after receiving inoculation. The animals’ spleens were removed for splenocyte isolation in order to perform in vitro lymphocyte proliferation and cytokine production assays. Blood was also collected for immunoglobulin analysis.
Indirect ELISA for analysis of IgG and its isotypes
ELISA plates (Costar, Corning Life Sciences, Lowell, MA, USA) were coated with So (1 μg in 100 μL of 50 mM Carbonate-bicarbonate buffer pH 9.6, in each well), incubated overnight at 4 oC and washed twice in 0.05 % PBS Tween-20 (PBS-T). Plates were then blocked with 200 μL of 5 % skim milk in 0.05 % PBS-T and incubated for 2 h at 37 oC. Next, the plates were washed once with PBS-T and 50 μL of diluted serum (1:50 in 1 % skim milk/PBS-T) was added to each well. Plates were then incubated for 1 h at 37 °C and washed five times with PBS-T. Next, wells were filled with 50 μL/well of HRP conjugated rabbit Ig antimouse IgG (Sigma-Aldrich, St Louis, MO, USA) at a dilution of 1:10,000 in 1 % skim milk/PBS-T to assess total IgG. To evaluate IgG1, IgG2a, IgG2b and IgG3, wells were filled with 50 μL/well of HRP conjugated rabbit Ig antimouse IgG1, IgG2a, IgG2b or IgG3 (Zymed, San Francisco, CA, USA), respectively, each diluted at 1:8.000 in 1 % skim milk/PBS-T. All plates were then incubated for 45 min at 37 oC. Each plate was washed five times in PBS-T and 50 μL/well of Citrate Phosphate Buffer pH 5.1, ortho-phenyl-diamine (Sigma, St Louis, MO, USA) and 30 % H2O2] were added and left for 15 min at room temperature in a dark chamber. Reactions were stopped with 25 μL/well of 4 N H2SO4 and Optical Density (OD) was measured at 490 nm using an ELISA Plate Reader (BIORAD, Hercules, CA, USA).
Lymphocyte proliferation assay
The spleen of each mouse was removed, washed three times with Hanks’ solution, and then placed in a petri dish containing 5 mL of RPMI 1640 medium (Gibco Laboratories, North Andover, MA, USA) supplemented with penicillin and streptomycin. The spleens were then macerated and the cellular suspension was transferred to a conical tube containing 5 mL of the same medium, followed by centrifugation at 400 g for 3 min. Pellets were resuspended in 0.17 M of NH4Cl for 5 min at 4 °C in order to lyse erythrocytes. Cells were washed 3 times with RPMI and then resuspended in RPMI enriched with 10 % bovine fetal serum.
Cell viability was determined by a Trypan Blue exclusion assay. 106 cells/mL were cultivated in 96-well microculture plates in RPMI-1640 medium supplemented with L-glutamine, penicillin/streptomycin, gentamicin and 10 % fetal calf serum. Cells were stimulated by So or Se C. pseudotuberculosis antigens (40 μg/mL), and pokeweed mitogen (2.5 μg/mL) was used as a positive control, while medium alone was used as a negative control. All plates were incubated for 120 h at 37o C under 5 % CO2. 1 μBq/well (10 μL) of fresh [3H] thymidine (GE Healthcare, Bucks, UK) was added 18 h prior to the end of the incubation time using a beta counter system (iMatic Canberra, Meriden, USA). After 120 h, plates were frozen at -20 oC and β-radiation was measured as described by Paule et al. (2004) [23]. Results are expressed in terms of a Stimulation Index (SI), calculated by dividing the β-radiation found for each stimulated sample by the radiation measured from its respective negative control.
ELISA for cytokines quantification
Cytokine analysis was performed in a culture supernatant obtained from cells (106/mL) cultivated in the same medium used for lymphoproliferation assay. Spleen cells were stimulated with So and Se C. pseudotuberculosis antigens (40 μg/mL), pokeweed mitogen (2,5 μg/mL) as a positive control, and the medium alone (negative control). The plates were incubated for 120 h at 37o C in 5 % CO2 [40]. The supernatant was collected, centrifuged, and kept at -20o C until use.
Cytokine profile analysis was performed using commercial kits for IFN-γ and IL-10 (R&D Systems, Minneapolis, MN, USA), and IL-12 and IL-4 (Pharmigen, San Jose, CA, USA) according to the manufacturer instructions. Results are expressed in ρg/mL.
Nitric oxide (NO) production assay
The presence of NO in the supernatant of spleen cells cultures that were incubated for 120 h was measured by nitrite assay, based on Griess reaction [41]. Briefly, supernatant (50 μL) was mixed with 50 μL of Griess reagent (1 % sulfanilamide and 0.1 % N-(1-naphthyl)ethylenediamine, in 5 % phosphoric acid) and incubated for 10 min at room temperature. Absorbances were measured at 492 ηm using an ELISA microplate reader (BioRad, Hercules, CA, USA). The standard curve of NO2− was prepared by diluting nitrite stock solution (1 M NaNO2 diluted in Milli-Q water) in spleen cell culture media. Results are expressed in ηg/mL.
Statistical analysis
For determination of statistical significance between experimental groups at an individual time-point, a analysis of variance (ANOVA) was performed using SPSS 12.0. (IBM Statistics, Chicago, EUA). A p value of <0.05 was considered significant.
Acknowledgements
Authors are sincerely grateful to the technical staff of the LABIMUNO/ICS for their assistance.
Funding
This study was supported by Laboratory of Immunology of Health Sciences Institute of Federal University of Bahia (Laboratório de Imunologia do Instituto de Ciências da Saúde da Universidade Federal da Bahia - LABIMUNO/ICS/UFBA) and by the Scientific and Technological Development Fund (Fundo de Desenvolvimento Científico e Tecnológico - FUNDECI) of Banco do Nordeste do Brasil S. A. (BNB).
Availability of data and materials
All the data supporting these findings is contained within the manuscript.
Authors’ contributions
MCS, APS and EJM carried out the immunoassays. HSPC and LFMC carried out the microbiological experiments. BJAP and SCT participated in the design of the study and performed the statistical analysis. EKNSL and ILON helped to draft the manuscript. VLCV and RJMN conceived of the study and participated in its design and coordination. All authors read and approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Consent for publication
Not applicable.
Ethics approval and consent to participate
This study is in agreement with the Ethical Principles for Animal Research established by Brazilian Society of Laboratory Animal Science (SBCAL), and was approved by the Institutional Review Board for Animal Research of the Institute of Health Sciences (CEUA - ICS) at the Federal University of Bahia (UFBA), Salvador-Brazil.
Abbreviations
- CL
Caseous lymphadenitis
- NO
Nitric oxide
- OD
Optical density
- Se
Secreted antigen
- So
Somatic antigen.
Contributor Information
Vera Lúcia Costa Vale, Phone: (+55) 71 9961 3187, Email: vvale@uneb.br.
Marcos da Costa Silva, Email: macosil@gmail.com.
Andréia Pacheco de Souza, Email: andreia.pachecosouza@yahoo.com.br.
Soraya Castro Trindade, Email: soraya.castrotrindade@gmail.com.
Lília Ferreira de Moura-Costa, Email: lmouracosta@gmail.com.
Ellen Karla Nobre dos Santos-Lima, Email: ellenobre@hotmail.com.
Ivana Lucia de Oliveira Nascimento, Email: ivana@nob-ba.com.br.
Hugo Saba Pereira Cardoso, Email: hugosaba@gmail.com.
Edson de Jesus Marques, Email: edjmarques@ig.com.br.
Bruno Jean Adrien Paule, Email: brunojeanadrien@yahoo.com.br.
Roberto José Meyer Nascimento, Email: rmeyer@ufba.br.
References
- 1.Unanian MM, Feliciano Silva AE, Pant KP. Abscesses and caseous lymphadenitis in goats in tropical semi-arid north-east Brazil. Trop Anim Health Prod. 1985;17:57–62. doi: 10.1007/BF02356137. [DOI] [PubMed] [Google Scholar]
- 2.Guimarães AS, Carmo FB, Heinemann MB, Portela RW, Meyer R, Lage AP, et al. High sero-prevalence of caseous lymphadenitis identified in slaughterhouse samples as a consequence of deficiencies in sheep farm management in the state of Minas Gerais, Brazil. BMC Vet Res. 2011;7:68. doi: 10.1186/1746-6148-7-68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Bastos BL, Dias Portela RW, Dorella FA, Ribeiro D, Seyffert N, Castro TLP, et al. Corynebacterium pseudotuberculosis: immunological responses in animal models and zoonotic potential. J Clin Cell Immunol. 2012;S4:005. [Google Scholar]
- 4.Costa MP, McCulloch JA, Almeida SS, Dorella FA, Fonseca CT, Oliveira DM, et al. Molecular characterization of the Corynebacterium pseudotuberculosis hsp60-hsp10 operon, and evaluation of the immune response and protective efficacy induced by hsp60 DNA vaccination in mice. BMC Res Notes. 2011;4:243. doi: 10.1186/1756-0500-4-243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Bastos BL, Loureiro D, Raynal JT, Guedes MT, Vale VL, Moura-Costa LF, et al. Association between haptoglobin and IgM levels and the clinical progression of caseous lymphadenitis in sheep. BMC Vet Res. 2013;9:254. doi: 10.1186/1746-6148-9-254. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Batey RG. Pathogenesis of caseous lymphadenitis in sheep and goats. Aust Vet J. 1986;63:269–72. doi: 10.1111/j.1751-0813.1986.tb08064.x. [DOI] [PubMed] [Google Scholar]
- 7.McKean S, Davies J, Moore R. Identification of macrophage induced genes of Corynebacterium pseudotuberculosis by differential fluorescence induction. Microbes Infect. 2005;7:1352–63. doi: 10.1016/j.micinf.2005.05.002. [DOI] [PubMed] [Google Scholar]
- 8.Ruiz JC, D’Afonseca V, Silva A, Ali A, Pinto AC, Santos AR, et al. Evidence for reductive genome evolution and lateral acquisition of virulence functions in two Corynebacterium pseudotuberculosisstrains. PLoS ONE. 2011;6:e18551. doi: 10.1371/journal.pone.0018551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Batey RG. Aspects of pathogenesis in a mouse model of infection by Corynebacterium pseudotuberculosis. Aust J Exp Biol Med Sci. 1986;64:237–49. doi: 10.1038/icb.1986.26. [DOI] [PubMed] [Google Scholar]
- 10.Soares SC, Silva A, Trost E, Blom J, Ramos R, Carneiro A, et al. The pan-genome of the animal pathogen Corynebacterium pseudotuberculosis reveals differences in genome plasticity between the biovar ovis and equi strains. PLoS ONE. 2013;8:e53818. doi: 10.1371/journal.pone.0053818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Pacheco LGC, Slade SE, Seyffert N, Santos AR, Castro TLP, Silva WM, et al. A combined approach for comparative exoproteome analysis of Corynebacterium pseudotuberculosis. BMC Microbiol. 2011;11:12. doi: 10.1186/1471-2180-11-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Dorella FA, Gala-Garcia A, Pinto AC, Sarrouh B, Antunes CA, Ribeiro D, et al. Progression of “OMICS” methodologies for understanding the pathogenicity of Corynebacterium pseudotuberculosis: the Brazilian experience. Comput Struct Biotechnol J. 2013;6:e201303013. doi: 10.5936/csbj.201303013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Cameron CM, Engelbrecht MM. Mechanism of immunity to Corynebacterium pseudotuberculosis (Buchanan, 1911) in mice using inactivated vaccine. Onderstepoort J Vet Res. 1971;38:73–82. [PubMed] [Google Scholar]
- 14.Pépin M, Seow HF, Corner L, Rothel JS, Hodgson AL, Wood PR. Cytokine gene expression in sheep following experimental infection with various strains of Corynebacterium pseudotuberculosis differing in virulence. Vet Res. 1997;28:149–63. [PubMed] [Google Scholar]
- 15.de Souza AP, Vale VLC, Silva MC, Araújo IBO, Trindade SC, Moura-Costa LF, et al. MAPK involvement in cytokine production in response to Corynebacterium pseudotuberculosis infection. BMC Microbiol. 2014;14:230. doi: 10.1186/s12866-014-0230-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Ellis JA, Lairmore MD, Otoole D, Campos M. Differential induction of Tumor Necrosis Factor Alpha in ovine pulmonary alveolar macrophages following infection with Corynebacterium pseudotuberculosis, Pasteurella haemolytica, or Lentiviruses. Infect Immun. 1991;59:3254–60. doi: 10.1128/iai.59.9.3254-3260.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Ellis JA, Russell HI, Du CW. Effect of selected cytokines on the replication of Corynebacteriurn pseudotuberculosis and ovine lentiviruses in pulmonary macrophages. Vet Immunol Immunopathol. 1994;40:31–47. doi: 10.1016/0165-2427(94)90013-2. [DOI] [PubMed] [Google Scholar]
- 18.Lan DT, Taniguhi S, Makino S, Shirahata N, Nakane A. Role of endogenous Tumor Necrosis Factor Alpha and Gamma Interferon in resistance to Corynebacterium pseudotuberculosis infection in mice. Microbiol Immunol. 1998;42:863–70. doi: 10.1111/j.1348-0421.1998.tb02362.x. [DOI] [PubMed] [Google Scholar]
- 19.Lan DT, Makino SI, Shirahata T, Yamada M, Nakane A. Tumor necrosis factor alpha and gama interferon are required for the development of protective immunity to secondary Corynebacterium pseudotuberculosis infection in mice. J Vet Med Sci. 1999;11:1203–8. doi: 10.1292/jvms.61.1203. [DOI] [PubMed] [Google Scholar]
- 20.Flynn JL, Chan J, Triebold KJ, Dalton DK, Stewart TA, Bloom BR. An essential role for Interferon gamma in resistance to Mycobacterium tuberculosis infection. J Exp Med. 1993;178:2249–54. doi: 10.1084/jem.178.6.2249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.El-Enbaawy MI, Saad MM, Selim SA. Humoral and cellular immune responses of a murine model against Corynebacterium pseudotuberculosis antigens. Egypt J Immunol. 2005;12:13–9. [PubMed] [Google Scholar]
- 22.Paule BJ, Azevedo V, Regis LF, Carminati R, Bahia CR, Vale VL, et al. Experimental Corynebacterium pseudotuberculosis primary infection in goats: kinetics of IgG and interferon-gamma production, IgG avidity and antigen recognition by Western blotting. Vet Immunol Immunopathol. 2003;96:129–39. doi: 10.1016/S0165-2427(03)00146-6. [DOI] [PubMed] [Google Scholar]
- 23.Paule BJ, Meyer R, Moura-Costa LF, Bahia RC, Carminati R, Regis LF, et al. Three-phase partitioning as an efficient method for extraction / concentration of immunoreactive excreted-secreted proteins of Corynebacterium pseudotuberculosis. Protein Expr Purif. 2004;34:311–16. doi: 10.1016/j.pep.2003.12.003. [DOI] [PubMed] [Google Scholar]
- 24.Meyer R, Regis L, Vale V, Paule B, Carminati R, Bahia R, et al. In vitro IFN-gamma production by goat blood cells after stimulation with somatic and secreted Corynebacterium pseudotuberculosis antigens. Vet Immunol Immunopathol. 2005;107:249–54. doi: 10.1016/j.vetimm.2005.05.002. [DOI] [PubMed] [Google Scholar]
- 25.Huygen K, Palfliet K. Strain variation in interferon gamma production of BCG-sensitized mice challenged with PPD II. Importance of one major autosomal locus and additional sexual influences. Cell Immunol. 1984;85:75–81. doi: 10.1016/0008-8749(84)90279-X. [DOI] [PubMed] [Google Scholar]
- 26.Oswald IP, Afroun S, Bray D, Petit JF, Lemaire G. Low response of BALB/c macrophages to priming and activating signals. J Leukoc Biol. 1992;52:315–22. doi: 10.1002/jlb.52.3.315. [DOI] [PubMed] [Google Scholar]
- 27.Tumitan AR, Monnazzi LG, Ghiraldi FR, Cilli EM. Machado de Medeiros BM. Pattern of macrophage activation in yersinia-resistant and yersinia-susceptible strains of mice. Microbiol Immunol. 2007;51:1021–8. doi: 10.1111/j.1348-0421.2007.tb03986.x. [DOI] [PubMed] [Google Scholar]
- 28.Muckle CA, Menzies PI, Li Y, Hwang YT, van Wesenbeeck M. Analysis of the immunodominant antigens of Corynebacterium pseudotuberculosis. Vet Microbiol. 1992;30:47–58. doi: 10.1016/0378-1135(92)90093-9. [DOI] [PubMed] [Google Scholar]
- 29.Eggleton DG, Haynes JA, Middleton HD, Cox JC. Immunisation against ovine caseous lymphadenitis: correlation between Corynebacterium pseudotuberculosis toxoid content and protective efficacy in combined clostridial-corynebacterial vaccines. Aust Vet J. 1991;68:322–5. doi: 10.1111/j.1751-0813.1991.tb03088.x. [DOI] [PubMed] [Google Scholar]
- 30.Hodgson AL, Tachedjian M, Corner LA, Radford AJ. Protection of sheep against caseous lymphadenitis by use of a single oral dose of live recombinant Corynebacterium pseudotuberculosis. Infect Immun. 1994;62:5275–80. doi: 10.1128/iai.62.12.5275-5280.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Paule BJA, Azevedo V, Moura-Costa LF, Freire SM, Regis LF, Vale VLC, et al. SDS-PAGE and Western blot analysis of somatic and extracellular antigens of Corynebacterium pseudotuberculosis. Rev Ciênc Méd Biol. 2004;3:44–52. [Google Scholar]
- 32.Pestka S, Krause CD, Sarkar D, Walter MR, Shi Y, Fisher PB. Interleukin-10 and related cytokines and receptors. Annu Rev Immunol. 2004;22:929–79. doi: 10.1146/annurev.immunol.22.012703.104622. [DOI] [PubMed] [Google Scholar]
- 33.Trinchieri G, Scott P. Interleukin-12: a proinflammatory cytokine with immunoregulatory functions. Res Immunol. 1995;146:423–31. doi: 10.1016/0923-2494(96)83011-2. [DOI] [PubMed] [Google Scholar]
- 34.Pohl-Koppe A, Balashov KE, Steere AC, Logigian EL, Hafler DA. Identification of a T cell subset capable of both IFN-γ and IL-10 secretion in patients with chronic Borrelia burgdorferi infection. J Immunol. 1998;160:1804–10. [PubMed] [Google Scholar]
- 35.Trinchieri G. Regulatory role of T cells producing both interferon gamma and interleukin 10 in persistent infection. J Exp Med. 2001;194:F53–7. doi: 10.1084/jem.194.10.f53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Fortune SM, Solache A, Jaeger A, Hill PJ, Belisle JT, Bloom BR, et al. Mycobacterium tuberculosis inhibits macrophage responses to IFN-γ through myeloid differentiation factor 88-dependent and -independent mechanisms. J Immunol. 2004;172:6272–80. doi: 10.4049/jimmunol.172.10.6272. [DOI] [PubMed] [Google Scholar]
- 37.Green SJ, Nacy CA, Meltzer MS. Cytokine-induced synthesis of nitrogen oxides in macrophages: a protective host response to Leishmania and other intracellular pathogens. J Leukoc Biol. 1991;50:93–103. doi: 10.1002/jlb.50.1.93. [DOI] [PubMed] [Google Scholar]
- 38.Pacheco LGC, Castro TLP, Carvalho RD, Moraes PM, Dorella FA, Carvalho NB, et al. A role for sigma factor σE in Corynebacterium pseudotuberculosis resistance to nitric oxide / peroxide stress. Front Microbiol. 2012;3:126. doi: 10.3389/fmicb.2012.00126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Moura-Costa LF, Bahia RC, Carminati R, Vale VL, Paule BJ, Portela RW, et al. Evaluation of the humoral and cellular immune response to different antigens of Corynebacterium pseudotuberculosis in Canindé goats and their potential protection against caseous lymphadenitis. Vet Immunol Immunopathol. 2008;126:131–41. doi: 10.1016/j.vetimm.2008.06.013. [DOI] [PubMed] [Google Scholar]
- 40.Salas-Téllez E, Núñez del Arco A, Tenorio V, Díaz-Aparicio E, de la Garza M, Suárez-Güemes F. Subcellular fractions of Brucella ovis distinctively induce the production of interleukin-2, interleukin-4, and interferon-gamma in mice. Can J Vet Res. 2005;69:53–7. [PMC free article] [PubMed] [Google Scholar]
- 41.Green LC, Wagner DA, Glogowski J, Skipper PL, Wishnok JS, Tannenbaun SR. Analysis of nitrite and (15N) nitrate in biological fluids. Anal Biochem. 1982;126:131–8. doi: 10.1016/0003-2697(82)90118-X. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All the data supporting these findings is contained within the manuscript.