

ABSTRACT
The phytohormones gibberellin (GA) and jasmonate (JA) regulate various aspects of plant development, growth and defense. Previous studies showed that both DELLA repressors in GA pathway and JA-ZIM domain (JAZ) proteins in JA pathway interact with and repress the WD-repeat/bHLH/MYB transcriptional complex to inhibit trichome initiation, and GA and JA respectively induce DELLAs and JAZs degradation to synergistically enhance trichome formation. In this study, we showed that the DELLA protein RGA and JAZ1 competitively bind to ENHANCER OF GLABRA3 (EGL3), a bHLH component of the WD-repeat/bHLH/MYB complex. GA and JA differently affect the expression and protein stability of the components of the WD-repeat/bHLH/MYB complex, and EGL3 and GL3 repress the expression of JAZ genes as a feedback. The novel findings help to understand the mechanism of the WD-repeat/bHLH/MYB complex in GA/JA-regulated trichome formation.
KEYWORDS: Gibberellin, jasmonate, WD-repeat/bHLH/MYB complex
The plant hormone gibberellin (GA) plays essential roles in root growth,1 seed germination,2 leaf expansion and senescence,3 stem elongation,4 flower development,5 trichome initiation,6 and plant resistance.7 The GA receptors GA-INSENSITIVE DWARF1a/b/c perceive GA signals and recruit DELLA repressors RGA, GAI, RGL1, RGL2, and RGL3 for ubiquitination and subsequent degradation, and release various DELLA-targeted proteins to control plant development and resistance.8,9
The phytohormone jasmonate (JA) induces the interactions of the JA receptor Coronatine Insensitive 1 (COI1) and JA-ZIM domain (JAZ) proteins, subsequent ubiquitination and degradation of JAZ proteins, which releases the diverse JAZ-inhibited transcriptional factors to regulate plant development, metabolism and defense, such as stamen development,10 trichome initiation,11 anthocyanin accumulation,12 root growth,13 leaf senescence14 and defense against herbivores and pathogens.15
The WD-repeat/bHLH/MYB transcriptional complex16,17 that is responsible for trichome initiation consists of the WD protein TRANS-PARENT TESTA GLABRA1 (TTG1), a bHLH transcription factor GLABRA3 (GL3) or ENHANCER OF GLABRA3 (EGL3), and the R2R3-MYB transcription factor GLABRA1 (GL1), and is the direct target of the DELLA repressors and JAZ repressors.12,18 Both JAZs and DELLAs interact with and repress the transcriptional activity of the bHLH and MYB components.12,18 GA and JA respectively promote DELLAs and JAZs degradation, and synergistically enhance trichome initiation by activating the WD-repeat/bHLH/MYB complex.12,18
Previous study exhibited that the DELLA protein RGA and JAZ1 physically associate with and synergistically repress the transcriptional activity of GL1 and EGL3.12,18 In this study we investigated whether RGA and JAZ1 competitively or concurrently bind to the components of the WD-repeat/bHLH/MYB complex by a pull-down assay. The purified EGL3 protein, fused with the maltose binding protein (MBP), was incubated with GST or GST-JAZ1. The results showed that MBP-EGL3 retained GST-JAZ1 but not GST (Fig. 1). When MBP-EGL3 was incubated with GST-JAZ1 and the increasing concentrations of myc-RGA, the ability of MBP-EGL3 in retaining GST-JAZ1 was attenuated by myc-RGA (Fig. 1), demonstrating that RGA competitively binds to EGL3.
Figure 1.
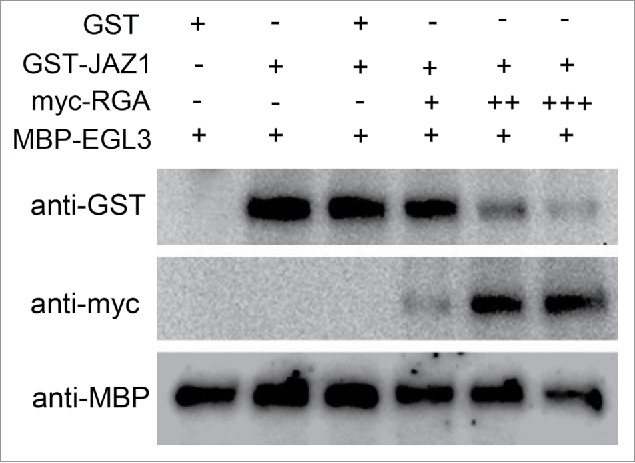
RGA and JAZ1 Competitively Bind to EGL3. In vitro pull-down assay showed that the interaction between MBP-EGL3 and GST-JAZ1 is attenuated by myc-RGA in a dosage-dependent manner. GST and GST-JAZ1 proteins were purified from E. coli using GST resin beads, and MBP-EGL3 protein was purified from E. coli using amylose resin beads. Total protein was extracted from 5 g of myc-RGA transgenic plants. Immobilized EGL3 was incubated with GST, GST-JAZ1 or myc-RGA as shown. The immunoprecipitated proteins were separated on SDS-PAGE and detected by anti-GST or anti-myc antibodies. The plus sign indicates the amount of myc-RGA. The total input of MBP-EGL3 was detected by anti-MBP antibody.
The expression of EGL3 could not be induced by JA, while GL3 and GL1 exhibit JA-induced expression pattern.12 Our quantitative real-time PCR analysis showed that GA treatment clearly enhanced the expression of GL1 and GL3 in wild-type Arabidopsis plant, but could not affect EGL3 and TTG1 expression (Fig. 2A). The expression of GL1 and GL3 was inhibited in the GA-deficient mutant ga1-3 (wild-type for RGA, GAI, RGL1, RGL2 and RGL3), and was recovered in Q3 (ga1-3 gai-t6 rgl1-1 rga-t2, wild-type for RGL2 and RGL3), penta (ga1-3 gai-t6 rga-t2 rgl1-1 rgl2-1, wild-type for RGL3), but not in Q1 (ga1-3 gai-t6 rgl1-1 rgl2-1, wild-type for RGA and RGL3) (Fig. 2B), suggesting that RGA inhibits the expression of GL3 and GL1. On the other hand, the expression of EGL3 and TTG1 was not affected in these GA-deficient mutants (Fig. 2B). Taken together, our results demonstrated that GA promotes the expression of GL1 and GL3, but not EGL3 and TTG1.
Figure 2.
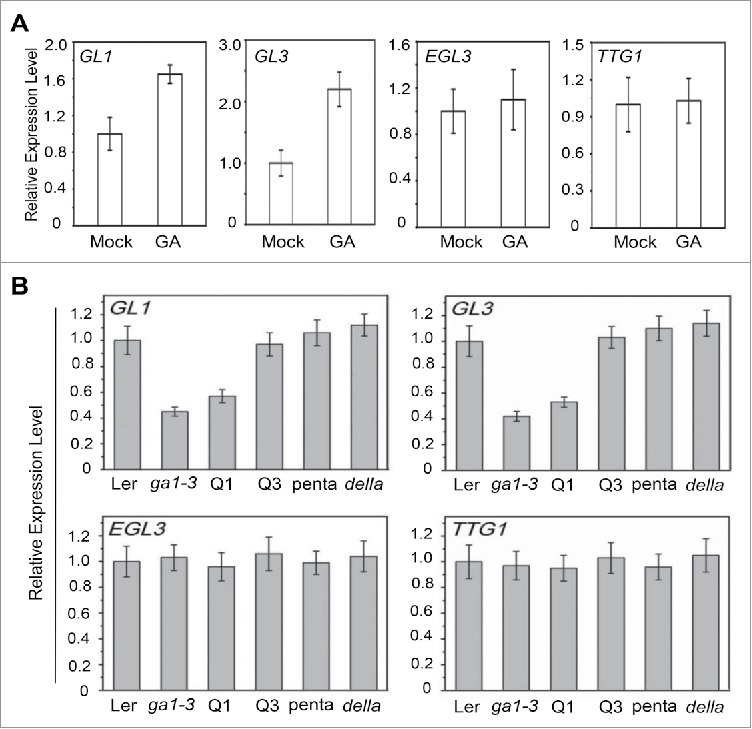
Regulation of the Expression of WD-repeat/bHLH/MYB Complex by Gibberellin. (A) Real-time PCR analysis for GL1, GL3, EGL3 and TTG1 in Col-0 wild-type treated without (Mock) or with Gibberellin (GA). ACTIN8 was used as the internal control. Error bars represent SE. (B) Real-time PCR analysis for GL1, GL3, EGL3 and TTG1 in the Landsberg erecta (Ler) wild-type, ga1-3, Q1 (ga1-3 gai-t6 rgl1-1 rgl2-1, wild-type for RGA and RGL3), Q3 (ga1-3 gai-t6 rgl1-1 rga-t2, wild-type for RGL2 and RGL3), penta (ga1-3 gai-t6 rga-t2 rgl1-1 rgl2-1, wild-type for RGL3) and della (gai-t6 rga-t2 rgl1-1 rgl2-1 rgl3-1) plants grown on MS medium. ACTIN8 was used as the internal control. Error bars represent SE.
Having shown that GA and JA induce the expression of GL1 and GL3 (Fig. 2),12 we next explored whether GA and JA affect the protein stability of GL1, GL3 and EGL3. As shown in Fig. 3A, JA displayed no effect on the GL1 degradation, while GA delayed the degradation of GL1. We observed that the degradation of myc-fused GL3 or EGL3, transiently expressed in N. benthamiana, was attenuated by the 26S proteasome inhibitor MG132 (Fig. 3B, 3C),19 suggesting that both GL3 and EGL3 are degraded through the 26S proteasome pathway. However, the degradation of GL3 and EGL3 was not enhanced or delayed by GA or JA (Fig. 3B, 3C), indicating that GA and JA don't affect their protein stability.
Figure 3.
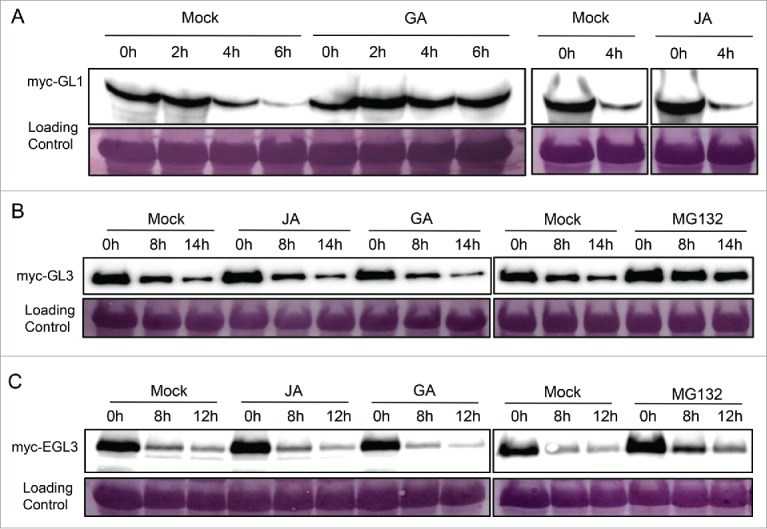
Effects of Jasmonate and Gibberellin on the Protein Stability of the WD-repeat/bHLH/MYB Complex. The Agrobacterium strain containing myc-fused GL1 (myc-GL1), GL3 (myc-GL3), or EGL3 (myc-EGL3) was infiltrated into N. benthamiana leaves. After 50 h, total protein was extracted from N. benthamiana leaves, incubated without (Mock) or with jasmonate (JA), gibberellin (GA), or MG132 for the indicated time (hours), and myc-GL1 (A), myc-GL3 (B), or myc-EGL3 (C) was detected by immunoblot using anti-myc antibody. The PVDF membrane was stained with Memstain to serve as loading control.
Previous studies showed that JAZ-targeted transcription factors may activate JAZ expression to serve as a negative feedback regulation.20 We further examined the JA-regulated expression of JAZ genes, including JAZ1, JAZ2, JAZ5, JAZ6, JAZ7, JAZ8, JAZ9 and JAZ10, in wild-type and the gl3 egl3 double mutant. The results showed that, compared with that in wild-type, gl3 egl3 exhibited enhanced JA-induced expression of JAZ genes (Fig. 4), suggesting that GL3 and EGL3 negatively regulate the expression of JAZs to form a positive feedback loop.
Figure 4.
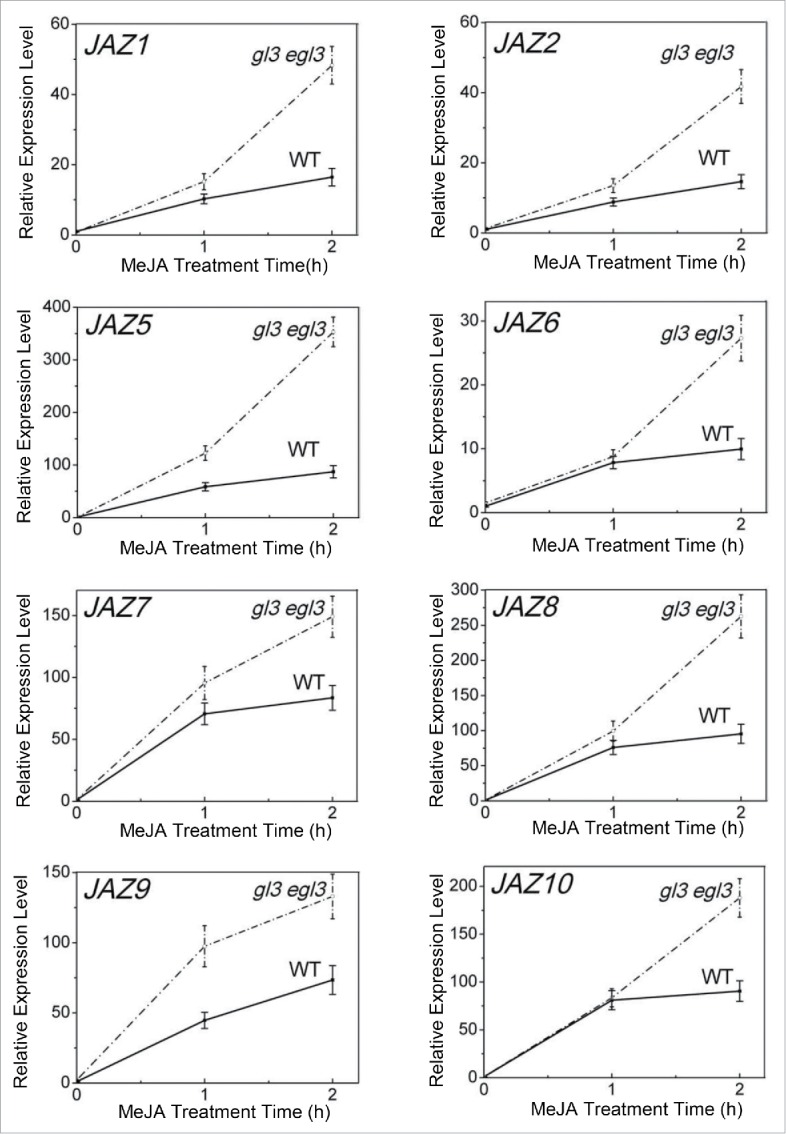
Expression of JAZ Genes in wide-type and the gl3 egl3 Double Mutant. Real-time PCR analysis for JAZ1, JAZ2, JAZ5, JAZ6, JAZ7, JAZ8, JAZ9 and JAZ10 in the 2 week-old seedlings of wild type (WT) and gl3 egl3 treated with 100 µM Methyl-jasmonate for 0, 1 or 2 hours. ACTIN8 was used as the internal control. Error bars represent SE.
GA and JA antagonistically regulate plant growth and defense,21,22 while they synergistically control trichome initiation, stamen development, and sesquiterpene biosynthesis.18,23,24 Our results provide novel mechanistic details of GA/JA-regulated trichome initiation: DELLAs and JAZs competitively bind to EGL3 for repression; GA and JA promote the expression of GL1 and GL3, but not EGL3, and only GA can stabilize the protein level of GL1; moreover, GL3 and EGL3 inhibit JAZ expression to form a positive feedback loop, which will be helpful for understanding the molecular synergism between GA and JA in plant development and metabolism.
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed.
Funding
This work was financially supported by the National Natural Science Foundation of China (Grant 31400254).
References
- 1.Fu X, Harberd NP. Auxin promotes Arabidopsis root growth by modulating gibberellin response. Nature 2003; 421:740-3; PMID:12610625; http://dx.doi.org/ 10.1038/nature01387 [DOI] [PubMed] [Google Scholar]
- 2.Piskurewicz U, Jikumaru Y, Kinoshita N, Nambara E, Kamiya Y, Lopez-Molina L. The gibberellic acid signaling repressor RGL2 inhibits Arabidopsis seed germination by stimulating abscisic acid synthesis and ABI5 activity. Plant Cell 2008; 20:2729-45; PMID:18941053; http://dx.doi.org/ 10.1105/tpc.108.061515 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Achard P, Gusti A, Cheminant S, Alioua M, Dhondt S, Coppens F, Beemster GT, Genschik P. Gibberellin signaling controls cell proliferation rate in Arabidopsis. Curr Biol 2009; 19:1188-93; PMID:19576768; http://dx.doi.org/ 10.1016/j.cub.2009.05.059 [DOI] [PubMed] [Google Scholar]
- 4.Kato J. Effect of Gibberellin on Elongation, Water Uptake, and Respiration of Pea-Stem Sections. Science 1956; 123:1132; PMID:17793433; http://dx.doi.org/ 10.1126/science.123.3208.1132 [DOI] [PubMed] [Google Scholar]
- 5.Cheng H, Qin L, Lee S, Fu X, Richards DE, Cao D, Luo D, Harberd NP, Peng J. Gibberellin regulates Arabidopsis floral development via suppression of DELLA protein function. Development 2004; 131:1055-64; PMID:14973286; http://dx.doi.org/ 10.1242/dev.00992 [DOI] [PubMed] [Google Scholar]
- 6.Gan Y, Yu H, Peng J, Broun P. Genetic and molecular regulation by DELLA proteins of trichome development in Arabidopsis. Plant Physiol 2007; 145:1031-42; PMID:17704233; http://dx.doi.org/ 10.1104/pp.107.104794 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Navarro L, Bari R, Achard P, Lison P, Nemri A, Harberd NP, Jones JD. DELLAs control plant immune responses by modulating the balance of jasmonic acid and salicylic acid signaling. Curr Biol 2008; 18:650-5; PMID:18450451; http://dx.doi.org/ 10.1016/j.cub.2008.03.060 [DOI] [PubMed] [Google Scholar]
- 8.Silverstone AL, Jung HS, Dill A, Kawaide H, Kamiya Y, Sun TP. Repressing a repressor: gibberellin-induced rapid reduction of the RGA protein in Arabidopsis. Plant Cell 2001; 13:1555-66; PMID:11449051; http://dx.doi.org/ 10.1105/tpc.13.7.1555 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Claeys H, De Bodt S, Inze D. Gibberellins and DELLAs: central nodes in growth regulatory networks. Trends Plant Sci 2013; 19(4):231-9; PMID:24182663; http://dx.doi.org/ 10.1016/j.tplants.2013.10.001 [DOI] [PubMed] [Google Scholar]
- 10.Song S, Qi T, Huang H, Ren Q, Wu D, Chang C, Peng W, Liu Y, Peng J, Xie D. The Jasmonate-ZIM Domain Proteins Interact with the R2R3-MYB Transcription Factors MYB21 and MYB24 to Affect Jasmonate-Regulated Stamen Development in Arabidopsis. Plant Cell 2011; 23:1000-13; PMID:21447791; http://dx.doi.org/ 10.1105/tpc.111.083089 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Yoshida Y, Sano R, Wada T, Takabayashi J, Okada K. Jasmonic acid control of GLABRA3 links inducible defense and trichome patterning in Arabidopsis. Development 2009; 136:1039-48; PMID:19234066; http://dx.doi.org/ 10.1242/dev.030585 [DOI] [PubMed] [Google Scholar]
- 12.Qi T, Song S, Ren Q, Wu D, Huang H, Chen Y, Fan M, Peng W, Ren C, Xie D. The Jasmonate-ZIM-Domain Proteins Interact with the WD-Repeat/bHLH/MYB Complexes to Regulate Jasmonate-Mediated Anthocyanin Accumulation and Trichome Initiation in Arabidopsis thaliana. Plant Cell 2011; 23:1795-814; PMID:21551388; http://dx.doi.org/ 10.1105/tpc.111.083261 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Chen Q, Sun J, Zhai Q, Zhou W, Qi L, Xu L, Wang B, Chen R, Jiang H, Qi J, et al.. The basic helix-loop-helix transcription factor MYC2 directly represses PLETHORA expression during jasmonate-mediated modulation of the root stem cell niche in Arabidopsis. Plant Cell 2011; 23:3335-52; PMID:21954460; http://dx.doi.org/ 10.1105/tpc.111.089870 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Shan X, Wang J, Chua L, Jiang D, Peng W, Xie D. The role of Arabidopsis Rubisco activase in jasmonate-induced leaf senescence. Plant physiology 2011; 155:751-64; PMID:21173027; http://dx.doi.org/ 10.1104/pp.110.166595 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Browse J. Jasmonate passes muster: a receptor and targets for the defense hormone. Annu Rev Plant Biol 2009; 60:183-205; PMID:19025383; http://dx.doi.org/ 10.1146/annurev.arplant.043008.092007 [DOI] [PubMed] [Google Scholar]
- 16.Zhang F, Gonzalez A, Zhao M, Payne CT, Lloyd A. A network of redundant bHLH proteins functions in all TTG1-dependent pathways of Arabidopsis. Development 2003; 130:4859-69; PMID:12917293; http://dx.doi.org/ 10.1242/dev.00681 [DOI] [PubMed] [Google Scholar]
- 17.Zhao M, Morohashi K, Hatlestad G, Grotewold E, Lloyd A. The TTG1-bHLH-MYB complex controls trichome cell fate and patterning through direct targeting of regulatory loci. Development 2008; 135:1991-9; PMID:18434419; http://dx.doi.org/ 10.1242/dev.016873 [DOI] [PubMed] [Google Scholar]
- 18.Qi T, Huang H, Wu D, Yan J, Qi Y, Song S, Xie D. Arabidopsis DELLA and JAZ Proteins Bind the WD-Repeat/bHLH/MYB Complex to Modulate Gibberellin and Jasmonate Signaling Synergy. Plant Cell 2014; 26:1118-33; PMID:24659329; http://dx.doi.org/ 10.1105/tpc.113.121731 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Patra B, Pattanaik S, Yuan L. Ubiquitin protein ligase 3 mediates the proteasomal degradation of GLABROUS 3 and ENHANCER OF GLABROUS 3, regulators of trichome development and flavonoid biosynthesis in Arabidopsis. Plant J 2013; 74:435-47; PMID:23373825; http://dx.doi.org/ 10.1111/tpj.12132 [DOI] [PubMed] [Google Scholar]
- 20.Chini A, Fonseca S, Fernandez G, Adie B, Chico JM, Lorenzo O, García-Casado G, López-Vidriero I, Lozano FM, Ponce MR, et al.. The JAZ family of repressors is the missing link in jasmonate signalling. Nature 2007; 448:666-71; PMID:17637675; http://dx.doi.org/ 10.1038/nature06006 [DOI] [PubMed] [Google Scholar]
- 21.Yang DL, Yao J, Mei CS, Tong XH, Zeng LJ, Li Q, Xiao LT, Sun TP, Li J, Deng XW, et al.. Plant hormone jasmonate prioritizes defense over growth by interfering with gibberellin signaling cascade. Proc Natl Acad Sci U S A 2012; 109:E1192-200; PMID:22529386; http://dx.doi.org/ 10.1073/pnas.1201616109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Hou X, Ding L, Yu H. Crosstalk between GA and JA signaling mediates plant growth and defense. Plant Cell Rep 2013; 32:1067-74; PMID:23525761; http://dx.doi.org/ 10.1007/s00299-013-1423-4 [DOI] [PubMed] [Google Scholar]
- 23.Cheng H, Song S, Xiao L, Soo HM, Cheng Z, Xie D, Peng J. Gibberellin acts through jasmonate to control the expression of MYB21, MYB24, and MYB57 to promote stamen filament growth in Arabidopsis. PLoS genetics 2009; 5:e1000440; PMID:19325888; http://dx.doi.org/ 10.1371/journal.pgen.1000440 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Hong GJ, Xue XY, Mao YB, Wang LJ, Chen XY. Arabidopsis MYC2 interacts with DELLA proteins in regulating sesquiterpene synthase gene expression. Plant Cell 2012; 24:2635-48; PMID:22669881; http://dx.doi.org/ 10.1105/tpc.112.098749 [DOI] [PMC free article] [PubMed] [Google Scholar]