

Abstract
The lipid bilayer of the plasma membrane is thought to be compartmentalized by the presence of lipid-protein microdomains. In eukaryotic cells, microdomains composed of sterols and sphingolipids packed in a liquid-ordered state, commonly known as lipid rafts, are believed to exist. While less studied in bacterial cells, reports on the presence of sterol or protein-mediated microdomains in bacterial cell membranes are also appearing with increasing frequency. Recent efforts have been focused on addressing the biophysical and biochemical properties of lipid rafts. However, most studies have been focused on synthetic membranes, mammalian cells, and/or model, non-pathogenic microorganisms. Much less is known about microdomains in the plasma membrane of pathogenic microorganisms. This review attempts to provide an overview of the current state of knowledge of lipid rafts in pathogenic fungi and the developing field of microdomains in pathogenic bacteria. The current literature on the structure and function and of microdomains is reviewed and the potential role of microdomains in growth, pathogenesis, and drug resistance of pathogens are discussed. Better insight into the structure and function of membrane microdomains in pathogenic microorganisms might lead to a better understanding of the process of pathogenesis and development of raft-mediated approaches for new methods of therapy.
Keywords: plasma membrane, lipid rafts, microdomain, sterol, pathogen
1. INTRODUCTION
Through the 1970s, the dominant view of the physical state of the cell membrane was that the membrane is a ‘fluid-mosaic’, in which the proteins are randomly distributed (Singer & Nicolson, 1972). In the late 1960s and early 1970s, in vitro studies with lipid monolayers and bilayers showed that long-chain, saturated lipids pack favorably with sterols to form a compact state (Chapman, Owens, Phillips, & Walker, 1969; Lee, 1977; Papahadjopoulos, Jacobson, Nir, & Isac, 1973). These observations gradually raised questions about the physical state of the plasma membrane leading to suggestions that natural membranes could have a heterogenous lipid bilayer (Karnovsky, Kleinfeld, Hoover, & Klausner, 1982; Linden & Fox, 1975; Thompson & Tillack, 1985). Since then, evidence has accumulated indicating that ordered sphingolipid/sterol-rich domains (i.e. lipid rafts) that co-exist with disordered fluid phases exist in cell membranes, and play an important role in cellular functions and lipid-protein and protein-protein interactions by selective inclusion or exclusion of proteins in specific domains (Ahmed, Brown, & London, 1997; Brown & London, 1997; Brown & Rose, 1992; Fastenberg, Shogomori, Xu, Brown, & London, 2003; Field, Holowka, & Baird, 1999; Harder, Scheiffele, Verkade, & Simons, 1998; Levental, Grzybek, & Simons, 2010; Lin & London, 2013; Schroeder, London, & Brown, 1994; Simons & Ikonen, 1997; Simons & Van Meer, 1988; Skibbens, Roth, & Matlin, 1989).
Recent years have seen an increasing interest in the studies of lipid rafts in the function of different cells. Several recent reviews have discussed the importance of lipid rafts in various cell functions such as signal transduction, endocytosis, ion homeostasis and cell growth among others (Hicks, Nalivaeva, & Turner, 2012; Mollinedo, 2012; Yaqoob & Shaikh, 2010). However, the majority of studies have been focused on mammalian cells or model microorganisms such as Saccharomyces cerevisiae (Bagnat, Keränen, Shevchenko, Shevchenko, & Simons, 2000; Bagnat & Simons, 2002a). With lipid rafts being suggested to play a role in many important cellular functions, an understanding of their functions in pathogenic microorganisms could help better define their role in cell function and pathogenicity and potentially pave the way for new therapeutic strategies. To this aim, the current chapter focuses on the role membrane microdomains in pathogenic bacteria and fungi, and provides an overview of microdomain composition, function, and role in pathogenicity.
2. WHAT ARE MEMBRANE DOMAINS MADE OF?
Lipid phase segregation is the central element to the formation of lipid microdomains. Thus, many pioneering studies focused on lipid phase in an attempt to understand the physical state of lipid domains in the plasma membrane. Early studies with synthetic bilayers of phospholipids using spin-labels and freeze fracture microscopy revealed the presence of lateral phase separations in the bilayer with areas of reduced fluidity in contact with more fluid areas (Grant, Wu, & McConnell, 1974; Kleemann, Grant, & McConnell, 1974). The areas of reduced fluidity were named the solid phase (Lβ), in which the lipid acyl chains are tightly packed and there is a low rate of lateral diffusion. The name solid ordered (So) is also sometimes used for this state. In contrast, the more fluid areas were named the liquid crystalline (Lα) phase (Shimshick & McConnell, 1973; Wu & McConnell, 1975). This state is now more commonly called the liquid disordered (Ld) state. The phase behavior of lipids is temperature dependent and the Lβ phase transition into Ld phase when the temperature is increased. The temperature at which this transition occurs is known as “gel to liquid transition temperature (Tm)” and depends on the structure of the lipid. The dependence of Tm on the acyl chains of lipids is well characterized. Lipids with longer saturated fatty acyl chains (e.g. most sphingolipids) have high Tm, whereas lipids with fatty acids having cis double bonds (e.g. most cellular phospholipids) have low Tm.
A number of studies using lipid monolayers and bilayers showed that the presence of cholesterol can significantly affect the phase behavior of lipids (Chapman et al., 1969; Ladbrooke, Williams, & Chapman, 1968; Recktenwald & McConnell, 1981; Vist & Davis, 1990). Notably, high concentrations of cholesterol leads to the emergence of a lipid phase that is characterized by the tight packing of the Lβ phase and the same time the relatively high lateral diffusion close to that Ld phase, leading to the name “liquid-ordered (Lo)” phase (Hjort Ipsen, Karlström, Mourtisen, Wennerström, & Zuckermann, 1987). The Lo phase was observed in lipid bilayers which contained cholesterol and high Tm phospholipids or sphingolipids, and was suggested to be due to the favorable packing of cholesterol with these lipids (Sankaram & Thompson, 1990), an observation that was confirmed by numerous later studies (Ahmed et al., 1997; Almeida, Vaz, & Thompson, 1993; Schroeder, Ahmed, Zhu, London, & Brown, 1998).
The discovery of the cholesterol-mediated Lo phase led to numerous studies focused on the phase behavior of ternary lipid mixtures as a simplistic model of eukaryotic cell membrane (a sterol, a high Tm and a low Tm lipid) (Ahmed et al., 1997; de Almeida, Fedorov, & Prieto, 2003; Schroeder et al., 1994; Silvius, del Giudice, & Lafleur, 1996; Veatch & Keller, 2003; Yuan, Furlong, Burgos, & Johnston, 2002). Most such studies have used a three-component phase diagram to represent the phase behavior of ternary lipid mixtures. A general three-component phase diagram is shown in Figure 1. In this diagram, each vertex represents a pure lipid component and each side represents a binary mixture. Points inside the triangle refer to ternary mixtures, in which the concentration of the high Tm lipid and cholesterol increases from left to the right and bottom to top, respectively.
Figure 1.
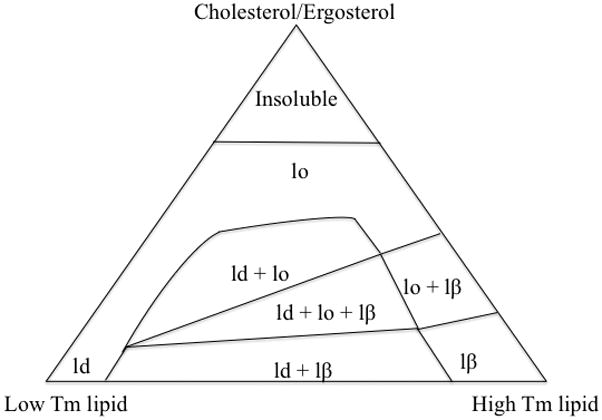
A hypothetical three phase diagram showing the interactions between a high Tm, a low Tm and a sterol. Ld: liquid-disordered phase, lo: liquid-ordered phase (lipid raft), lβ: solid-ordered phase (gel).
In discussing three-phase diagrams it should be borne in mind that apart from lipid composition, phases in such diagrams are highly dependent on temperature (de Almeida et al., 2003). The diagram in Figure 1 presents a situation in which the temperature is lower than the phase transition temperature of the high Tm lipid and higher than the transition temperature of the low Tm lipid. Such conditions are likely true for most mammalian cells at physiological temperature. Sphingomyelin a saturated sphingolipid abundant in mammalian cells has a transition temperature of 35–45 °C (Calhoun & Shipley, 1979), whereas unsaturated phospholipids abundant in mammalian cells have Tm values of less than 0 °C (Koynova & Caffrey, 1998). The first and most important conclusion from a three-phase diagram is that multiple lipid phases can exist at the same time in a mixture of lipids (Figure 1). In conditions of interest for biological membranes where high Tm lipids and cholesterol are lower than 50% of the lipid content (Lange, Swaisgood, Ramos, & Steck, 1989), these coexisting phases include the Ld phase in the presence of Lo or So phases. This observation indicates that ordered and disordered phases can coexist in biologically relevant lipid ratios. In such conditions, an increase in cholesterol concentrations translates to an increase in lipid order and often favors the presence of more ordered phases. Note that the cholesterol solubility limit in binary mixtures with phospholipids has been reported to be between 51% to 66% with higher concentrations leading to crystallization (Huang, Buboltz, & Feigenson, 1999). Such conditions are usually physiologically irrelevant and are shown as “insoluble” in Figure 1.
Evidence from phase diagrams suggest that ordered lipid phases in biological membranes are likely rich in sterols and high Tm lipids as each of these compounds promote ordered phase formation. The outer leaflet of the plasma membrane of mammalian cells is rich in sphingolipids and cholesterol so these membranes are likely to have ordered domains in their membranes. This hypothesis was first supported in mammalian cell membranes using an interesting characteristic of ordered lipid phases: insolubility in cold, non-ionic detergents (explained in detail in Section 3.2.1). Since the Lo lipid phases do not solubilize in detergents, isolation of detergent insoluble fractions from mammalian cell membranes was considered as the first evidence for the presence of lipid rafts in mammalian cells (Brown & Rose, 1992; Schroeder et al., 1994). Since the first observation of detergent resistant membranes numerous studies have followed reporting the presence of lipid rafts in mammalian cells membranes, and although the details remain controversial it is now widely accepted that lipid rafts exist in the plasma membrane of mammalian cells and play a role in various cell functions (Babina, Donatello, Nabi, & Hopkins, 2011; Brown, 2006; George & Wu, 2012; Grassme et al., 2003; Simons & Toomre, 2000; Suzuki, 2012; Triantafilou, Miyake, Golenbock, & Triantafilou, 2002).
In contrast to mammalian cells, membrane microdomains are highly understudied in bacteria and fungi. This is partly due to the smaller cell size of these microorganisms, differences in lipid compositions compared to mammalian cells and diversity in morphology and growth conditions. The remainder of this chapter focuses on membrane microdomains in pathogenic fungi and bacteria with a view to provide insight on methods of study, microdomain composition, and potential roles in pathogenesis. It should be noted that studies on non-pathogenic microorganisms, most notably the yeast S. cerevisiae (Bagnat, Chang, & Simons, 2001; Bagnat et al., 2000; Bagnat & Simons, 2002b; Klose et al., 2010), have been invaluable in understanding the role of rafts in cellular functions. However, such studies are not the focus of this chapter and the interested reader is referred to the pertinent literature for more information on those topics.
3. MEMBRANE DOMAINS IN PATHOGENIC FUNGI
3.1. Raft-forming Lipids in Fungi
Due to their different lipid composition, the sterols and saturated lipids that can form the Lo phase in a fungal plasma membrane are different than those in mammalian cells. The primary sterol in fungal cells is ergosterol, not cholesterol. Also, the sphingolipids in fungi are generally glycosphingolipids (i.e. sphingolipids connected to sugar molecules), not sphingomyelin. Glycosphingolipids in fungi include glucosylceramide (GlcCer), inositolphosphorylceramide (IPC), mannosyl-inositolphosphoylceramide (MIPC), and mannosyl-diinositolphosphorylceramide (MIP2C) (Bagnat et al., 2000; Rittershaus et al., 2006) (inositol is not a sugar, but does have a carbohydrate-like structure). These sphingolipids differ significantly from the glycosphingolipids of mammalian cells.
Sterols and saturated sphingolipids in fungi are better raft formers compared to their mammalian counterparts based on in vitro studies. Using fluorescence quenching experiments, Xu and colleagues (Xu et al., 2001) showed that mixtures of ergosterol with various saturated lipids form more stable lipid domains compared to mixtures of cholesterol with the same lipids. This was likely due to the presence of an additional double bond in the B ring, which led to better sterol-saturated lipid packing. Glycosphingolipids are also better ordered domain formers than non-glycosylated sphingolipids, i.e. they form gel states that melt at higher temperatures than sphingomyelins. This is likely due to their long acyl chains and the structure of their headgroups, which allows for extensive hydrogen bonding due to the presence of many hydroxyl groups (Redfern & Gericke, 2004). IPC has been shown to have higher Tm compared to acyl-chain matched sphingomyelin consistent with a role of the headgroup structure in promoting raft formation (Björkbom et al., 2010). In addition, IPC isolated from the yeast S. cerevisiae, has been shown to form ordered domains in mixtures with ergosterol and phosphatidylinositol (Klose et al., 2010). The strong raft-promoting properties of ergosterol (Xu et al., 2001) and fungal glycosphingolipids does not necessarily mean that all fungal cells contain lipid rafts, as the degree of unsaturation in unsaturated phospholipids and sterol-sphingolipid-unsaturated lipid ratios play a significant role in raft-formation, and is different for various cell types. However, the in vitro observations reveal that fungal cells possess the compositional requirements for the formation of lipid rafts.
3.2. Methods to Study Membrane Domains in Fungi
Except for the model yeast S. cerevisiae, a limited number of methods have been used to study membrane domains in pathogenic fungi. Isolation of detergent resistant membranes, staining and extraction/substitution of sterols are the main methods used in fungal cells, which are explained in more detail below. Several other methods can potentially be used for domain studies in pathogenic fungi; however, this section is focused on methods that have been used to detect membrane domains in pathogenic fungi and other methods that might be applicable are not discussed.
3.2.1. Isolation of Detergent Resistant Membranes
The most widely used method for characterization of lipid rafts in cells is the isolation of detergent resistant membranes. Due to their tight packing, lipids in the gel (Dennis, 1974) and liquid-ordered phase (Schroeder et al., 1994) do not solubilize in ice-cold non-ionic detergents such as Triton X-100. Exposure to TX-100 followed by density centrifugation will result in floatation of insoluble lipids known as detergent resistant membranes (DRMs) (Brown & Rose, 1992; London & Brown, 2000). DRMs have been generally used as a surrogate for lipid rafts and their lipid and protein composition has been characterized to provide information on raft composition. A number of proteins such as plasma membrane-ATPase1 (Pma1) and glycosylphosphatidylinositol-anchored plasma membrane protein, Gas1p, have been shown to be concentrated in the DRMs of S. cerevisiae (Bagnat et al. 2002) and have been typically used as a marker for DRMs in other fungi.
3.2.2. Staining and Light Microscopy
The polyene antibiotic filipin has been the molecule of choice for sterol visualization in fungi. Filipin is a sterol-binding molecule, which has a structure similar to amphotericin B (Loura, Castanho, Fedorov, & Prieto, 2001). Filipin is fluorescent and allows for visualization of sterols using fluorescence microscopy. Since sterols are a crucial component of lipid rafts, filipin fluorescence can be used to detect sterol rich areas in the cell, which might correspond to areas in which lipid rafts are present. A problem in working with filipin is that it is very sensitive to photobleaching. Also, filipin can disrupt membrane architecture at high concentrations or long incubation times and care should be taken in using filipin for imaging (Smejkal & Hoff, 1994; Takeshita, Higashitsuji, Konzack, & Fischer, 2008).
Sterol rich domains observed in cells using light microscopy should not necessarily be equated with lipid raft domains. Sterol rich regions can be as large as a few microns, whereas rafts are believed to be most frequently nano-sized (London, 2005; Pike, 2009); thus, SRDs could likely represent of a region containing a high concentration of lipid rafts. However, given the metabolic activity of cells, rather than the presence of raft domains at equilibrium, sterol rich regions also could represent membrane regions to which a high level of sterols have been delivered, and not linked to the presence of distinct sphingolipid enriched liquid ordered domains.
It should be noted that in addition to filipin, by staining non-raft regions, the dye FAST diI (with affinity towards Ld phases) has also been used for detection of large of lipid rafts in S. cerevisiae (Toulmay & Prinz, 2013). However, to the best of our knowledge, its use has been limited to non-pathogenic fungi.
3.2.3. Sterol Depletion/Substitution
The use of chemical compounds to deplete sterols has been a widely used method to study lipid rafts. Since lipid rafts in eukaryotes are made of sterols and sphingolipids, depletion of sterols should often lead to raft disruption allowing for studies of their role in cellular processes. The cyclic oligosaccharide, methyl beta cyclodextrin (MβCD), has been widely used to extract cholesterol from mammalian and in some cases fungal cells (Siafakas, Wright, Sorrell, & Djordjevic, 2006; Zidovetzki & Levitan, 2007). MβCD is water soluble, but has a hydrophobic core, which can efficiently bind sterols. Cyclodextrins are sterol donor-acceptor molecules and have also been used to extract one sterol and replace it with another (LaRocca et al., 2013; Tagliari et al., 2012). Sterol substitution experiments have the added benefit that they allow for studies of sterol structure without drastically changing the sterol mass balance in the cell, thus reducing the undesired effects that might be due to mechanisms other than raft-disruption. Cyclodextrins have been used to extract ergosterol from fungal cells and also to perform sterol substitution studies (Siafakas et al., 2006; Tagliari et al., 2012). However, their use has been limited and there has been a report suggesting that cyclodextrins are not effective in ergosterol extraction from S. cerevisiae (Bagnat et al., 2000). This might be due to the presence of cell wall that hinders the interaction of MβCD with the plasma membrane.
3.2.4. Inhibiting Sterol/sphingolipid Biosynthesis
In addition to MβCD, inhibitors of sterol and sphingolipid synthesis have also been used to study raft function in the cells. Azoles are a class of drugs that block the 14-α-demethylation of lanosterol in the ergosterol biosynthesis pathway (Ghannoum & Rice, 1999) and have been used to study the role of sterols in raft formation in fungi. Compounds that inhibit sphingolipid biosynthesis have also been used to disrupt fungal lipid rafts as a method to study their function in cells. Primary among such compounds are myriocin, which inhibits serine palmitoyltransferase at the first committed step of sphingolipid biosynthesis (Miyake, Kozutsumi, Nakamura, Fujita, & Kawasaki, 1995) and aureobasidin A, which inhibits IPC synthase and thus the synthesis of glycosphingolipids (Takesako et al., 1993).
3.3. Observation of Membrane Domains in Pathogenic Fungi
Many of the concepts later used in studies of pathogenic fungi were first discovered in the non-pathogenic yeast S. cerevisiae. The first observation of lipid rafts in fungal cells was reported in S. cerevisiae (Bagnat et al., 2000). In this work, detergent resistant membranes were successfully isolated from S. cerevisiae and analyzed for their lipids and proteins. Glycosphingolipids and ergosterol were reported as the main raft-forming lipids, while the glycophosphatidylinositol (GPI)-anchored protein Gas1p and the membrane bound proton-pump protein Pma1 were found to be two of the most abundant raft proteins. The findings of this study were important as these lipids and proteins have been generally used to identify lipid rafts in other fungi. There is a growing body of research further analyzing the role and composition of lipid rafts in S. cerevisiae (Aresta-Branco et al., 2011; Klose et al., 2010; Toulmay & Prinz, 2013). However, in this section we have focused on reports regarding lipid rafts in pathogenic fungi, namely C. albicans, C. neoformans, A. nidulans and H. capsulatum.
Candida albicans
C. albicans is a dimorphic opportunistic pathogen capable of growing both by budding (yeast) and hyphal (mold) formation. C. albicans is generally benign, but can cause mucosal or even blood stream infections under certain conditions, and immunocompromised individuals are particularly susceptible. C. albicans is also able to form biofilms on medical implants, causing infections which might require the removal of the implant (Ramage, Martínez, & Lopez-Ribot, 2006).
The first evidence for the existence of lipid rafts in C. albicans was generated by Martin and Konopka (Martin & Konopka, 2004), who assessed the role of sterols in morphogenesis by filipin staining C. albicans cells during budding and hyphal growth. It was shown that sterols are homogenously distributed in the plasma membrane during budding growth, but are significantly concentrated at hyphal tips during hyphal growth. Detergent resistant membranes were isolated from budding cells and hyphae and showed similar protein composition indicating that differences in staining are due to localization of sterol-rich domains (SRDs) at hyphal tips. Disrupting sphingolipid biosynthesis by myriocin and sterol biosynthesis by ketoconazole led to defects in hyphal formation demonstrating the importance of sphingolipids and sterol-rich domains for hyphal growth of C. albicans.
Lipid microdomains in C. albicans seem to play a role in another change of morphology: changing from planktonic form to biofilms. A study by Lattif and colleagues (Lattif et al., 2011) showed that filipin stained C. albicans biofilms much more intensely than planktonic forms suggesting the presence of more lipid rafts. This proposal is supported by the presence of significantly larger amounts of glycosphingolipids in biofilms than in planktonic forms. The ability of C. albicans to form biofilms was significantly reduced after treatment with myriocin and aureobacidin A suggesting that sphingolipids play an important role in biofilm formation, likely by regulating the amount of lipid rafts.
The protein composition of DRMs in C. albicans has also been the focus of various studies. Proteome analysis has revealed the presence of 29 proteins in the DRM of C. albicans (Insenser, Nombela, Molero, & Gil, 2006). Interestingly, many of the identified proteins were those known to be located in the mitochondria, the Golgi apparatus or the endoplasmic reticulum suggesting that lipid rafts may also be present in intracellular membranes as has been suggested by others (Mollinedo et al., 2011). The presence of lipid rafts in intracellular membranes of Candida spp. is also supported by another study reporting a number of mitochondrial proteins as part of the proteome of DRMs in C. guilliermondii (An, Chen, Li, Qin, & Tian, 2014). It should be noted that the possibility that detergent insolubility of mitochondria arises from properties other than the presence of Lo domains has not been ruled out.
Proteomic studies performed on the DRMs isolated from the budding and hyphal C. albicans cells revealed that plasma membrane proteome differs based on the morphology (Alvarez & Konopka, 2007). Several notable proteins have been identified in C. albicans DRM. A novel protein, Ngt1, was discovered and shown to be a transporter of N-acetylglucosamine, a known inducer of hyphae in C. albicans (Simonetti, Strippoli, & Cassone, 1974). The presence of Ngt1 in DRMs further supports the importance of lipid rafts in regulating changes in morphology. Other important proteins suggested to be part of the DRM proteome in C. albicans include CaCdr1P, an efflux pump protein and a drug transporter (Pasrija, Panwar, & Prasad, 2008), and Rta2p, which acts to transfer long chain sphingoid bases from the cytofacial to the exofacial side of the plasma membrane (Wang et al., 2012). Both of these proteins were highly concentrated in the DRM fractions, and localized at the plasma membrane of C. albicans. Detergent resistance and plasma membrane localization of these proteins was abolished in response to mutations or drugs that affect ergosterol and sphingolipid biosynthesis, suggesting that raft-association is required for the proper localization of these proteins (Pasrija et al., 2008; Wang et al., 2012).
C. neoformans
The budding yeast Cryptococcus neoformans is a facultative intracellular pathogen and the leading cause of fungal meningitis. C. neoformans is able to grow at both acidic and neutral conditions allowing it to survive and proliferate inside immune cells and in the bloodstream. Studies of lipid rafts in C. neoformans have been focused on virulence factors. Virulence factors have been defined as “mechanisms that allow the fungus to efficiently divide and survive inside the host,…resist host immune attack, and…damage the host” (Coelho, Bocca, & Casadevall, 2014). Similar to C. albicans, the first observation of membrane domains in C. neoformans was provided by filipin staining, revealing the presence of sterol-rich domains in the bud tips of C. neoformans (Nichols, Fraser, & Heitman, 2004). The first isolation of detergent resistant membranes in C. neoformans was performed by Siafakas and colleagues (Siafakas et al., 2006). This analysis revealed the presence of PE, PC, PS, ergosterol and ceramide monohexosides (such as GlcCer) as the main lipids of C. neoformans DRMs. Interestingly, IPC, MIPC and MIP2C were not reported as part of the DRM lipidome in C. neoformans. This might be due to the techniques used in the study (these lipids have been generally visualized using radiolabelling (Rittershaus et al., 2006) or lower amounts of these lipids in C. neoformans compared to other fungi. Two virulence factors of C. neoformans were found in the DRM fractions: Phospholipase B1 (which acts to degrade the phospholipids of the host cell (Chrisman, Albuquerque, Guimaraes, Nieves, & Casadevall, 2011)) and Cu/Zn superoxide dismutase (which protects the fungus against oxidative stress (Cox et al., 2003)).
Studies have also focused on the role of lipid rafts in the ability of C. neoformans to survive at different levels of ambient pH. A C. neoformans mutant that lacks the gene sphingolipid C9 methyltransferase (Smt1) produces GlcCer that is not methylated. This strain loses its ability to grow at neutral pH and shows attenuated virulence (Singh et al., 2011). This mutant strain showed increased membrane fluidity as evidenced by a slight reduction in the fluorescent anisotropy of live cells. In addition, atomic force microscopy analysis of reconstituted lipid extracts showed a reduction in the height of the ordered lipid phases suggesting a less ordered packing of the vertical fatty acyl chains. The role of GlcCer methylation in membrane fluidity is unclear; however, it is possible that methylation is required for efficient packing of GlcCer with sterols and other sphingolipids (Del Poeta, Nimrichter, Rodrigues, & Luberto, 2014). Another possibility is that a mutation in the sphingolipid pathway might result in complex changes in membrane composition by affecting other pathways and the synthesis of sterols and/or unsaturated phospholipids.
The connection between membrane fluidity and growth remains unclear. A recent study has shown that similar to S. cerevisiae and C. albicans the protein Pma1 is concentrated in DRMs of C. neoformans and plays an important role in fungal growth and survival in acidic conditions (Farnoud, Mor, Singh, & Del Poeta, 2014). Further studies on characterization of lipid raft proteins might help elucidate the role of lipid rafts in growth at pH 7.4.
Aspergillus nidulans
Aspergillus is a fungus commonly found as mold in the environment. Aspergillus spp. are able to cause disease in immunocompromised patients with inhalation of conidia (spores) being the common route of infection. Aspergillus infections are generally limited to the lungs, but can disseminate to the bloodstream in severely immunocompromised patients. Most Aspergillus infections are caused by A. fumigatus; however, other strains, such as A. nidulans are also known to cause infections (Segal et al., 1998). Compared to C. albicans and C. neoformans, much less is known about the membrane microdomains in Aspergillus. However, in similarity to those fungi, filipin staining has revealed the presence of SRDs at the hyphal tips and the sites of septation in A. nidulans (Pearson, Xu, Sharpless, & Harris, 2004; Takeshita et al., 2008). SRDs localize the cell end marker protein, Tea1, and disruption of SRDs displaces this protein from the hyphal tips (Takeshita et al., 2008). Interestingly, the interactions between SRDs and proteins appear to be reciprocal. Flotillin is a lipid raft marker in mammalian cells (Dermine et al., 2001) and there is a homologue in Aspergillus. A. nidulans mutants lacking this protein showed disrupted SRDs and reduced growth rate (Takeshita, Diallinas, & Fischer, 2012). However, the flotillin did not localize in the SRDs, suggesting an indirect relationship between its localization and SRD formation.
Histoplasma capsulatum
H. capsulatum is a temperature-dependent dimorphic fungus, which is found as mold in the environment but grows as yeast in physiological temperatures. Inhalation of conidia can lead to severe lung infections in immunocompromised patients. To our knowledge, only one study has focused on characterization of membrane microdomains in H. capsulatum (Tagliari et al., 2012). In this study, DRM were isolated from H. capsulatum revealing the presence of ergosterol, PE, PC, PI and glycosphingolipids GlcCer, IPC, MIPC and MIP2C as detergent insoluble lipids. Extraction of sterols using MβCD resulted in the displacement of Pma1 and several other proteins from the detergent resistant fraction. It should be noted that sterol extraction can act not only by abolishing ordered domains, but also by simply reducing the resistance of ordered domains to solubilization to detergent (London, 2005; Xu & London, 2000). However, in this study residual DRM containing sphingolipids were detected after MβCD treatment. Interestingly, sterol depletion also significantly reduced the infectivity of alveolar macrophages by H. capsulatum. Both protein displacement and reduced infectivity persisted even after substituting ergosterol with cholesterol indicating the importance of sterol chemical structure in proper protein localization and infectivity.
3.4. The Role Membrane Domains in Fungal Pathogenesis
A summary of studies of lipid rafts in pathogenic fungi is provided in Table 1. As shown in the Table, the literature on lipid rafts in pathogenic fungi remains fairly small and is divided between various microorganisms. Based on the limited literature, it is difficult to pinpoint one universal role for lipid rafts in all pathogenic fungi. However, one area in which lipid rafts appear to facilitate fungal infections is by regulating changes in fungal morphology. SRDs were observed in hyphal tips in C. albicans (Martin & Konopka, 2004) and A. nidulans (Pearson et al., 2004; Takeshita et al., 2008) suggesting that lipid rafts regulate hyphal growth. In addition, the involvement of lipid rafts in biofilm formation (Lattif et al., 2011) suggests that rafts might play a role in all forms of changes in morphology. Hyphal growth and biofilm formation are important determinants of C. albicans pathogenicity. Hyphae have been suggested to promote tissue penetration and facilitate invasive growth (Sudbery, Gow, & Berman, 2004). In fact, C. albicans mutants that are locked in the yeast form are avirulent in a mouse model (Lo et al., 1997). In addition, biofilms are significantly more resistant to antifungal drugs than planktonic cells (Ramage, VandeWalle, Bachmann, Wickes, & López-Ribot, 2002) and are able to colonize implant devices causing significant health problems.
Table 1.
Published studies of lipid rafts in pathogenic fungi.
| Microorganism | Study | Methods | Proposed Raft Function |
|---|---|---|---|
| C. albicans | Martin & Konopka (2004) | Filipin staining, DRM isolation, Blocking sterol/sphingolipid synthesis | Hyphal growth |
| Alvarez & Konopka (2007) | DRM isolation, Proteomics | Hyphal growth | |
| Lattif et al. (2011) | Filipin staining, Blocking sphingolipid synthesis | Biofilm formation | |
| Inserner, Nombela, Molero & Gil (2006) | DRM isolation, Proteomics | Not Applicable, proteomic study | |
| Wang et al. (2012) | Filipin staining, DRM isolation, Blocking sterol/sphingolipid synthesis, Proteomics | Protein sorting | |
| C. neoformans | Nichols, Fraser & Heitman (2004) | Filipin staining | Budding growth |
| Siafakas, Wright, Sorrell & Djordjevic (2006) | DRM isolation, Sterol extraction | Protein sorting | |
| Singh et al. (2012) | Fluorescent anisotropy, Atomic force microscopy | Membrane integrity, Growth at neutral pH | |
| Farnoud, Mor, Singh & Del Poeta (2014) | DRM isolation | Protein sorting, Growth at acidic pH | |
| A. nidulans | Pearson, Xu, Sharpless & Harris (2004) | Filipin staining | Hyphal growth, Protein sorting |
| Takeshita, Higashitsuji, Konzack & Fischer (2008) | Filipin staining | Hyphal growth, Protein sorting | |
| Takeshita, Diallinas & Fischer (2012) | Filipin staining, DRM isolation | Hyphal growth, Protein sorting | |
| H. capsulatum | Tagliari et al. (2012) | DRM isolation, Sterol substitution, Lipidomics | Infectivity |
Lipid rafts may also contribute to fungal pathogenesis by regulating drug resistance. CaCdr1p is an efflux pump protein involved in multi-drug resistance in C. albicans (Shukla, Saini, Jha, Ambudkar, & Prasad, 2003). When expressed in S. cerevisiae, this protein was associated with detergent resistant membranes and disruption of ergosterol or sphingolipid biosynthesis pathway displaced this protein from detergent resistant fractions and inhibited the ability of the fungus to resist methotrexate and fluconazole (Pasrija et al., 2008). It has also been suggested that lipid rafts might also be involved in drug resistance of filamentous fungi by regulating membrane fluidity. The polymeric compound chitosan has been shown to have fungicidal effect against the filamentous fungi by permeabilizing the fungal plasma membrane (Palma-Guerrero et al., 2009). Interestingly, a study of chitosan-resistant and chitosan-sensitive filamentous fungi revealed that a N. crassa desaturase mutant with greatly reduced levels of polyunsaturated fatty acids, and thus increased membrane rigidity, was chitosan resistant (Palma-Guerrero et al., 2010). These observations suggest that a rigid plasma membrane, a high fraction of, or all of which, is in the Lo state, might act as a barrier to membrane-permeabilizing drugs.
Lipid rafts also play a role in pathogenicity by concentrating virulence factors. Phosphalipase B1 (Plb1) and Cu/Zn SOD, two important virulence factors of C. neoformans, were reported to be concentrated in detergent resistant membranes (Siafakas et al., 2006). Plb1 acts to degrade the phospholipids of the host immune cells, possibly resulting in the escape of phagocyotosed C. neoformans from the phagolysosomal membrane, reach the cytosol, and acquire nutrients (Chrisman et al., 2011). Plb1 mutants in C. neoformans show less nonlytic exocytosis (escape of cryptococcal cells from the macrophages after ingestion, with survival of both fungal and mammalian cells (Nicola, Robertson, Albuquerque, da Silveira Derengowski, & Casadevall, 2011)), likely due to reduced ability to survive the hostile environment of host phagolysosomes (Chayakulkeeree et al., 2011). Cu/Zn superoxide dismutase protects the fungus against oxidative stress inside the phagolysosomes by reducing super-oxide anions to hydrogen peroxide (Cox et al., 2003). Lipid rafts may also play a role in pathogenicity of C. neoformans by concentrating the cryptococcal Pma1 protein. As noted above, Pma1 is a proton pump, and it maintains the intracellular pH of fungal cells in the physiological range (Holyoak et al., 1996). Pma1 is a raft-localized protein and its association with rafts is required for surface delivery and protein stability at the plasma membrane (Bagnat et al., 2001; Gong & Chang, 2001). C. neoformans mutants in which DRM-associated Pma1 was not functioning properly showed attenuated virulence, likely due to their inability to survive the acidic environment of the phagolysosome (Farnoud et al., 2014; Shea, Kechichian, Luberto, & Del Poeta, 2006). Finally, reduced phagocytosis of ergosterol-depleted H. capsulatum cells by macrophages suggests the fungal lipid rafts may also play a role in fungal infection (Tagliari et al. 2012). It is possible that changes in membrane fluidity upon ergosterol depletion trigger the secretion of proteins with anti-phagocytic function. The secretion of such proteins have been reported in C. neoformans (Luberto et al., 2003), but their dependence on membrane fluidity has not been studied.
It is likely that rafts play other roles in pathogenic fungi, or that they play different roles at different times. For example, it is likely that fungal raft proteins may have multiple conformations or varying protein-protein interactions, and thus different functions in different host environments. In addition, the principles of raft organization have not been fully explored, and may involve more than just lipid-lipid interaction. Some specialized fungal membrane domains (e.g. eisosomes) may be organized by plasma membrane proteins (Douglas & Konopka, 2014). Thus, more studies and better characterization of raft lipids and proteins are needed to elucidate all raft functions in pathogenic fungi.
4. MEMBRANE DOMAINS IN PATHOGENIC BACTERIA
4.1. Domain-forming Lipids in Bacteria
Bacterial microdomains are significantly different from those in eukaryotes. Bacterial membranes are enriched in phospholipids, lipopolysaccharides and various lipoproteins (Huijbregts, de Kroon, & de Kruijff, 2000). However, most bacteria do not have sphingolipids (Parsons & Rock, 2013) and only a few bacteria contain sterols, with sterol containing bacteria including Helicobacter, Mycoplasma, Ehrlichia, Anaplasma, Brachyspira and Borrelia. (Haque, Hirai, Yokota, & Oguma, 1995; Hirai et al., 1995; Lin & Rikihisa, 2003; Livermore, Bey, & Johnson, 1978; Smith, 1971; Trott, Alt, Zuerner, Wannemuehler, & Stanton, 2001). Due to the important role of sterols in promoting lipid rafts in eukaryotic cells, much of the studies on lipid rafts in bacteria has been focused on sterol-containing species, and in particular Borrelia burgdorferi. However, the presence of microdomains has been shown even in sterol-lacking bacteria, as will be described below. It is worth mentioning that some non-pathogenic bacteria synthesize hopanoids, which have a chemical structure similar to that of cholesterol (Rohmer, Bouvier, & Ourisson, 1979) and studies have already started addressing the question whether these bacteria contain membrane microdomains (Sáenz, Sezgin, Schwille, & Simons, 2012). Such studies will not be discussed in the following section, which is focused on pathogenic bacteria. In addition, the methods to study lipid rafts in bacteria are similar to those in fungal cells and thus are not discussed in detail.
4.2. Observation of Membrane Domains in Pathogenic Bacteria
Borrelia burgdorferi
B. burgdorferi is an extracellular tick-borne spirochete that causes Lyme disease. The most prominent manifestation of Lyme disease is a skin rash with central clearing at the site of inoculation known as eritema migrans. Other organs and tissues including the heart, joints and the nervous system can be affected due to the ability of bacterium to disseminate through the blood stream (Benach et al., 1983). The membrane of B. burgdorferi contains phosphatidycholine, phospatidylglycerol and lipoproteins (Belisle, Brandt, Radolf, & Norgard, 1994; Brooks, Vuppala, Jett, & Akins, 2006; Jones, Bourell, Norgard, & Radolf, 1995; Kenedy, Lenhart, & Akins, 2012; Radolf et al., 1995). In addition it has free cholesterol and two cholesterol glycolipids, acylated cholesteryl galactoside (ACGal) and cholesteryl galactoside (CGal) as well as a non-cholesterol glycolipid, monogalactosyl diacylglycerol (MGalD) (Ben-Menachem, Kubler-Kielb, Coxon, Yergey, & Schneerson, 2003; Schroder et al., 2003; Stubs et al., 2009; Stubs et al., 2011; Wheeler et al., 1993). Interestingly, Borrelia does not have the biosynthetic ability to synthesize cholesterol and has to acquire it from the mammalian host. B. burgdorferi recruits the cholesterol needed to form the glycolipids from its host cells. The mechanisms by which B. burgdorferi interacts with the host cell has not been elucidated yet but it has been hypothesized that such interaction could be mediated through lipid raft-lipid raft interactions (Crowley et al., 2013). In any case, in this recruitment process there is an exchange of cholesterol, antigens and lipids between the host cell and the spirochete (Crowley et al., 2013). In addition to recruiting cholesterol through direct contact with the cell, B. burgdorferi has the ability to recruit cholesterol from host derived vesicles (Crowley et al., 2013). In both cases, the acquisition of cholesterol in B. burgdorferi is time dependent (Crowley et al., 2013) and cholesterol is glycosylated in the spirochete by undetermined bacterial enzymes (Östberg, Berg, Comstedt, Wieslander, & Bergström, 2007) before it is subsequently exported to the membrane.
The presence of sterols in B. burgdorferi has led to questions whether Borrelia contains lipid rafts similar to those observed in eukaryotic cells. To test this hypothesis, LaRocca and colleagues (LaRocca et al., 2010) performed ultrastructure analysis on B. burgdorferi. Previous studies established the cross-reaction between Borrelia glycolipids and host gangliosides by showing that B. burgdorferi elicits an antibody response in infected rats that targeted the host gangliosides (Garcia-Monco, Seidman, & Benach, 1995). The cross-reaction between monosialotetrahexosyl ganglioside (GM1) and Borrelia cholesterol glycolipids allowed for the use of an asialo-GM1 antibody to study the localization of cholesterol glycolipids with TEM. Using a particle-conjugated asialo-GM1 antibody and another particle-conjugated CB2 antibody, which binds to the outer surface protein B (OspB), it was shown that the surface protein and cholesteryl glycolipids form microdomains in B. burgdorferi. Moreover, the size and the number of these clusters were temperature-dependent and were present in both culture and mouse derived spirochetes (LaRocca et al., 2010). Detergent resistant membranes similar to those of eukaryotic cells could be isolated from the bacterium and revealed the presence of cholesteryl glycolipids and proteins, in the floating (detergent insoluble) fractions further corroborating the presence of lipid rafts containing specific proteins. [It should be noted that cholesteryl glycolipids are also found in the DRM fraction from plant cells (Lefebvre et al., 2007; Simon-Plas, Perraki, Bayer, Gerbeau-Pissot, & Mongrand, 2011)]. Treatment with MβCD, which led to extraction of sterols and sterol-glycolipids, resulted in the relocalization of proteins in the detergent soluble fractions (LaRocca et al., 2010). Also, both lipoproteins detected in the insoluble fraction were shown to co-localize with Borrelia glycolipids in the membrane of the spirochete by TEM.
Definitive proof for the existence of lipid rafts in B. burgdorferi was found when treatment with MβCD was followed by supplementing the bacterium with a set of different sterols (LaRocca et al., 2013). In vitro experiments with synthetic lipid bilayers have demonstrated that the ability of sterols to promote raft-formation is dependent on their chemical structure (Bakht & London, 2006; Wang, Megha, & London, 2004; Xu et al., 2001; Xu & London, 2000). Substituting cholesterol with strongly raft-promoting sterols (e.g. ergosterol) or intermediate raft-promoting sterols (e.g. lanosterol) did not affect the presence of sterol glycolipids domains in the ultrastructure of B. burgdorferi as shown by TEM and Förster Resonance Electron Transfer (FRET) in live cells. However, raft-disrupting sterols (e.g. coprostanol) led to disruption of these structures. Interestingly, upon prolonged incubation, the bacterium lost its membrane integrity after sterol depletion or in the presence of raft-disrupting sterols as evidenced by the visualization of exposed periplasmic flagella, that are not visible in normal spirochetes, and the release of cytosolic proteins into the external environment (LaRocca et al., 2013). These observations indicated that sterol-mediated membrane microdomains exist in Borrelia and regulate its membrane integrity.
Although lipid rafts contain a subset of specific proteins little is known about the contribution of proteins to the stability of the microdomains. A recent study showed that outer surface proteins play a role in membrane domain stability in B. burgdorferi although their role is not as crucial as that of sterols. A double-mutant lacking raft-associated proteins, OspA and OspB, showed significantly reduced domain stability as detected by FRET. Membrane domain stability was recovered in the double mutant when it was complemented with OspA, but not when it was complemented with another surface lipoprotein, OspC, that is not associated with the microdomains, suggesting that only those proteins that localize into the microdomain regulate membrane domain stability (Toledo et al., 2014). The evidence that bacterial surface proteins play an important role in the stability of microdomains is of significance not only for Borrelia but also for bacteria that lack sterols. Although only a few such bacteria have been studied for their ability to form microdomains (see below), the possibility of domain promotion by surface proteins suggests that many more bacteria might contain membrane microdomains, which should be a focus of future studies.
Bacillus subtilis
B. subtilis is a rod-shaped gram-positive bacterium commonly found in soil. B. subtilis is generally used in probiotics; however, infections in immune compromised individuals have been reported both with B. subtilis and with other strains (Nelms, Larson, & Barnes-Josiah, 1997; Oggioni, Pozzi, Valensin, Galieni, & Bigazzi, 1998). B. subtilis cells lack cholesterol; however evidence is accumulating that membrane proteins play a role in microdomain formation in their membranes. Flotillin is a protein frequently reported in the DRM of eukaryotic cells (Langhorst, Reuter, & Stuermer, 2005) and is commonly used as a raft-marker protein (Macdonald & Pike, 2005). B. subtilis contains two flotillin-like proteins floT and floA (Mielich-Süss, Schneider, & Lopez, 2013). To examine the role of flotillin in microdomain formation in B. subtilis, Donovan and Bramkamp (Donovan & Bramkamp, 2009) used a genetically modified strain producing fluorescently tagged floT. Observation of this strain under fluorescence microscopy revealed the presence of several flotillin domains across the membranes. These domains were dynamic and changed in their size and location with time. FloT was only found in the membrane and floT enriched DRMs were isolated from B. subtilis providing further evidence for the involvement of floT in the formation of ordered membrane domains.
Analogously to lipid rafts in budding yeast, membrane microdomains in B. subtilis appear to play a role in morphogenetic processes, such as sporulation, biofilm formation, and cell division. A genetically modified B. subtilis strain lacking floT showed defective sporulation (Donovan & Bramkamp, 2009). In addition, floA and floT were observed in the septum of dividing cells indicating that they interact with septum-localized proteins (Yepes et al., 2012). Overexpression of flotillins promoted biofilm formation and increased cell division leading to a reduction in cell size, and loss of the rod shape (Mielich-Süss et al., 2013).
Even in sterol-lacking bacteria, sterol-like molecules are required for organization of microdomains. The yisP gene in B. subtilis shows similarity with squalene synthases in fungi. Since squalene is a molecule involved in ergosterol biosynthesis in fungi (Ghannoum et al., 1994), this finding implied that sterol-like molecules might be present in B. subtilis (López & Kolter, 2010). Incubation of bacteria with zaragozic acid, a molecule known to inhibit squalene synthases, inhibited biofilm formation by B. subtilis, changed the protein profile of DRMs and abrogated flotillin microdomains suggesting that flotillin microdomains depend on a sterol-like molecule for stability, and are analogous to eukaryotic lipid rafts (López & Kolter, 2010). These observations were confirmed by Laurdan spectroscopy experiments (Bach & Bramkamp, 2013). The emission spectrum of the fluorescent dye Laurdan is dependent on the polarity of the surrounding environment and shows a difference in disordered vs. ordered lipid phases due to the presence of less water molecules in ordered phases. Staining wild-type B. subtilis cells with Laurdan revealed the presence of dispersed areas of high lipid order due to the presence of membrane microdomains. However, in B. subtilis ΔyisP a coalescence of these ordered areas into a single larger domain was observed indicating a change in membrane heterogeneity. Similar phenomena were observed in mutants lacking flotillin-like proteins suggesting that these proteins and sterol-like molecules are both involved in maintaining domain integrity (Bach & Bramkamp, 2013).
Staphylococcus aureus
S. aureus is a gram-positive bacterium generally found in hair and noses of humans and animals. S. aureus commonly causes skin infections; however, it can also infect the bloodstream and various organs. Infections with S. aureus can be hard to treat due to the emergence of methicillin-resistant S. aureus (MRSA) (Enright et al., 2002). A sequence homolog of flotillin exists in S. aureus and the possibility for the presence of membrane microdomains containing flotillin-like proteins was examined by fusion of a fluorescent tag to this protein (López & Kolter, 2010). The fused fluorescent protein was localized to a single domain in the bacterial membrane suggesting the presence of flotillin-induced microdomains. The localization of this protein at the membrane was abolished by zaragozic acid, suggesting the importance of sterol like molecules in domain formation.
Isolation of DRMs from S. aureus followed by protein sequencing showed the presence of 18 proteins in the DRMs including elastin-binding protein EbpS, which is involved in biofilm formation (López & Kolter, 2010). Treating the bacterium with clomazone, statin, and zaragozic acid (inhibitors of various steps of sterol biosynthesis) abrogated biofilm formation, and in case of clomazone and statin led to cell death, suggesting that sterol-like molecules play a role in biofilm formation and survival of S. aureus. Interestingly, S. aureus, contains a lipid, staphyloxanthin, that is structurally similar to cholesteryl glycolipids. A polyisoprene replaces cholesterol (which itself is derived biosynthetically from isoprene units). Whether this lipid participates in raft formation is not clear.
Based on the current evidence from the literature, microdomains differ significantly in fungal cells, sterol-containing, and sterol-lacking bacteria (Figure 2). While in fungal cells, sphingolipids and cholesterol pack to form microdomains (Figure 2-a), in sterol-containing bacteria either cholesterol glycolipids may play the role of both sterols and sphingolipids (Figure 2-b) or other lipids substitute for sphingolipids. In these microorganisms, membrane proteins stabilize microdomains but are not necessary for microdomain formation. Finally, in sterol-lacking bacteria, microdomain formation appears to be mediated by the packing of flotillin-like proteins and yet unknown sterol-like lipids (Figure 2-c). Although membrane microdomains have been identified only in Borrelia burgdorferi, Bacillus subtilis, and S. aureus, it seems likely that sterol or protein-mediated membrane microdomains will soon be discovered in other bacteria.
Figure 2.
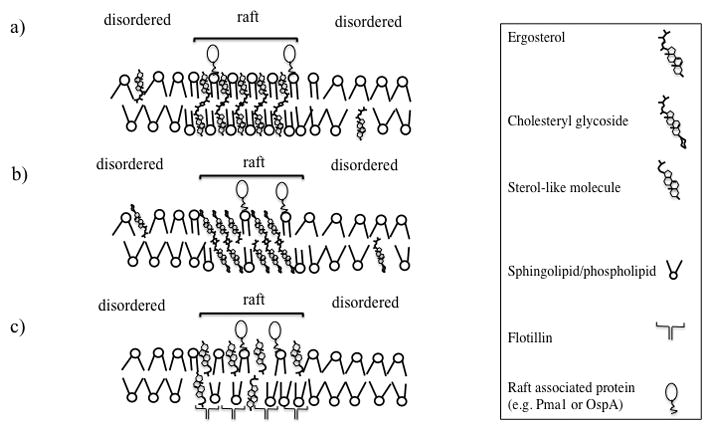
A hypothetical schematic of lipid rafts in (a) fungal, (b) sterol-containing and (c) sterol-lacking bacteria: (a) in fungal cells, ergosterol and glycosphingolipids pack to form membrane microdomains, (b) in sterol-containing bacteria, cholesteryl glycolipids play the role of both sterols and sphingolipids, and (c)in sterol- lacking bacterial flotillin and sterol-like molecules mediate raft formation. Certain membrane lipids and proteins are not included for simplicity.
4.3. Role of Membrane Domains in Bacterial Pathogenesis
Prokaryotic membrane microdomains appear to play a similar role to eukaryotic microdomains with regard to cell division and morphogenesis. Similarly to sterol-rich domains in fungi, flotillin-like proteins are concentrated in the septa in dividing B. subtilis cells. In both cases, the microdomains likely act as a hub for concentrating proteins required for septum formation. Microdomains also play a significant role in biofilm formation in bacteria, and B. subtilis mutants deficient in flotillin-like proteins (and thus heterogeneous microdomains) do not form biofilms. The inability of bacteria to form biofilms upon microdomain disruptions is strikingly similar to the inability of fungi to form biofilms upon lipid raft disruption (Lattif et al., 2011). This phenomenon is likely caused by disruption of microdomain-mediated, protein-protein interaction. FtsH is a protease essential for the differentiation of biofilm matrix producers. This protein is shown to directly interact with flotillin-like proteins and does not localize to DRM fractions in the absence of flotillin-like proteins suggesting a link between protein-protein interaction, microdomains, and biofilm formation (Yepes et al., 2012). This is of biomedical importance because biofilms have superior resistance to antibiotics and host immune response, and so significantly contribute to the persistence of bacterial infections.
Protein function and secretion machinery also depend on the function of membrane microdomains. Analysis of the protein profile of detergent resistant fractions in B. subtilis revealed that flotillin-like proteins interact with proteins that have a role in diverse cell functions, such as cell wall and energy metabolism and protein secretion, and mutants deficient in flotillin-like proteins also show reduced protein secretion (Bach & Bramkamp, 2013). Extracellular secretion is an important part of bacterial pathogenesis and allows bacteria to release toxins and virulence factors to the host (Lee & Schneewind, 2001). For example S. aureus produces extracellular toxins able to form pores in eukaryotic plasma membrane (Dinges, Orwin, & Schlievert, 2000). Extracellular release is also utilized by B. burgdorferi, in which raft components such as choleteryl glycolipids are released into the medium via outer membrane vesicles (Crowley et al., 2013). If delivered to the host cell, cholesteryl glycolipids might not be recognized by the host immune response due to their structural similarity to host lipids, or cause the host cells to become the target of immune response due to the presence of lipid antigens (Crowley et al., 2013).
Lipid rafts might also play the role of an environmental sensor for the pathogen. It has been shown that the size of microdomains in B. burgdorferi decreases with increasing the temperature (LaRocca et al., 2010). Borrelia is exposed to a wide range of temperatures as it has to survive both inside ticks and in mammalian hosts. Temperature–dependent changes in microdomain size and membrane fluidity might act as a sensor of the extracellular environment, so that, for example, a reduction in microdomain size can signal the entrance to the mammalian host. Changes in microdomains could also affect the raft-localized lipoproteins with further implications in signal transduction.
From a therapeutic point of view, bacterial lipid rafts may also be a drug target. The bactericidal CB2 antibody against Borrelia depends on sterols for its efficacy and sterol depletion significantly reduces its bactericidal activity (LaRocca et al., 2010). Also, zaragozic acid, which inhibits the activity of squalene synthase, has been shown to prevent biofilm synthesis in B. subtilis and S. aureus. In fact, inhibitors of squalene synthase have been shown to inhibit the virulence of S. aureus (Liu et al., 2008). Thus, drugs that affect membrane structure via altering lipid raft formation might be an effective therapy against bacterial infections, and in particular resistant biofilms.
5. UNEXPLORED AREAS AND FUTURE DIRECTIONS
In the past two decades, there has been increasing interest in the presence of membrane microdomains in living cells and the literature in this field is rapidly expanding. The fundamental knowledge gained on lipid-lipid interactions has been significantly improved by in vitro studies, while the presence of membrane microdomains in living cells has been demonstrated by a number of in vivo studies. Despite this progress, there are currently many outstanding questions and areas that need to be further explored to provide an understanding on the role of membrane domain in pathogenesis. The number of studies performed in pathogenic microorganisms is significantly less than those performed in model microorganisms such as S. cerevisiae, and pathogenic bacteria in particular are still highly understudied. While many outstanding questions remain regarding the role of membrane domains in pathogenic microorgansisms, we believe the following are some of the major unexplored areas that need to be addressed.
Do all bacteria have membrane domains?
While the presence of lipid rafts in eukaryotic cells is now widely accepted, it is unclear whether all bacteria include microdomains. Of the several bacterial species that contain sterols only Borrelia has been studied in detail and it is not clear if microdomains exist in other sterol-containing pathogenic bacteria. Also it is unclear whether sterol-containing molecules mediate domain formation in sterol-lacking bacteria. It has been suggested that sterol-like molecules mediate membrane domain in B. subtilis (Bach & Bramkamp, 2013; López & Kolter, 2010), while other sterol-like molecules such as hopanoids have been suggested to induce raft-formation in bacteria (Sáenz et al., 2012). The answers to these questions can have important implications for therapeutic strategies. For example, if sterol-like molecules contribute to membrane integrity in bacterial pathogens, then inhibitors of their synthesis might be very potent drugs against a variety of infections, as noted above (Liu et al., 2008).
More methods are needed to study microdomains
The paucity of appropriate methods has been a road-block in studies of lipid rafts. This is perhaps epitomized by the fact that the conventional method of DRM extraction has many limitations (London, 2005; London & Brown, 2000). Biophysical methods such as FRET, fluorescent anisotropy, and Laurdan spectroscopy have provided a significant amount of information about lipid rafts in live bacterial cells; however, such methods have been rarely used in fungal cells. These methods depend on effective delivery of fluorescent probes into the cell membrane, a requirement that is hampered by the presence of cell wall and in some cases a capsule in fungal cells. Nanotechnology-based application such as the use antibody/probe-conjugated particles or the use of small fluorescent particles may be able overcome this problem. In addition, the use of new microscopy techniques and lipidomics can significantly improve the current state of knowledge on molecular diffusion in rafts and the molecular species of raft-forming molecules, respectively.
How do lipid rafts change during the course of growth and pathogenesis?
Although existing studies have provided information on the proteins associated with lipid rafts, the changes in the protein profile of lipid rafts during various stages of pathogenesis is still unknown. For example, it has been proposed that different proteins are expressed in B. burgdorferi during various stages of infection (Tilly, Bestor, Dulebohn, & Rosa, 2009; Tilly, Bestor, & Rosa, 2013). However, it is unclear whether these proteins perform similar functions in maintaining lipid rafts and/or if these proteins are released to the extracellular environment. It also not known if lipid rafts differ in commensal and non-commensal microorganisms. It is possible that the amount of lipid rafts increases during infections, presenting a platform for the concentration of virulence factors. Studies comparing pathogenic and non-pathogenic strains, especially in microorganisms that switch from commensalism to pathogenicity might help investigate this possibility. A related issue is that the role of rafts in various stages of bacterial or fungal growth remains unclear. B. subtilis cells show few microdomains during exponential growth, but the number of microdomains increases as the bacterium reaches the stationary phase. A similar appearance of vacuolar microdomains at various stages of growth has been noted in S. cerevisiae (Toulmay & Prinz, 2013). Finally, the role of microdomains in maintaining cell viability during various stages of growth and inside the host requires further studies. Clearly, the structure and biological function of lipid rafts in pathogenic microorganisms is a field that is only in its infancy, and is likely to be a major area of future research.
Acknowledgments
This work was supported by NIH grants GM 099082 (to E.L.), RO1 AI47837 (to J.B.K.), and AI56168, AI71142, and AI100631 (to M.D.P.). M.D.P. is a Burroughs Wellcome Investigator in Infectious Diseases.
References
- Ahmed SN, Brown DA, London E. On the origin of sphingolipid/cholesterol-rich detergent-insoluble cell membranes: physiological concentrations of cholesterol and sphingolipid induce formation of a detergent-insoluble, liquid-ordered lipid phase in model membranes. Biochemistry. 1997;36:10944–10953. doi: 10.1021/bi971167g. [DOI] [PubMed] [Google Scholar]
- Almeida P, Vaz W, Thompson T. Percolation and diffusion in three-component lipid bilayers: effect of cholesterol on an equimolar mixture of two phosphatidylcholines. Biophysical Journal. 1993;64:399–412. doi: 10.1016/S0006-3495(93)81381-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alvarez FJ, Konopka JB. Identification of an N-acetylglucosamine transporter that mediates hyphal induction in Candida albicans. Molecular Biology of the Cell. 2007;18:965–975. doi: 10.1091/mbc.E06-10-0931. [DOI] [PMC free article] [PubMed] [Google Scholar]
- An B, Chen Y, Li B, Qin G, Tian S. Ca 2+–CaM regulating viability of Candida guilliermondii under oxidative stress by acting on detergent resistant membrane proteins. Journal of Proteomics. 2014;109:38–49. doi: 10.1016/j.jprot.2014.06.022. [DOI] [PubMed] [Google Scholar]
- Aresta-Branco F, Cordeiro AM, Marinho HS, Cyrne L, Antunes F, de Almeida RF. Gel domains in the plasma membrane of Saccharomyces cerevisiae Highly ordered, ergosterol-free, and sphingolipid-enriched lipid rafts. Journal of Biological Chemistry. 2011;286:5043–5054. doi: 10.1074/jbc.M110.154435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Babina IS, Donatello S, Nabi IR, Hopkins AM. Lipid rafts as master regulators of breast cancer cell function 2011 [Google Scholar]
- Bach JN, Bramkamp M. Flotillins functionally organize the bacterial membrane. Molecular Microbiology. 2013;88:1205–1217. doi: 10.1111/mmi.12252. [DOI] [PubMed] [Google Scholar]
- Bagnat M, Chang A, Simons K. Plasma membrane proton ATPase Pma1p requires raft association for surface delivery in yeast. Molecular Biology of the Cell. 2001;12:4129–4138. doi: 10.1091/mbc.12.12.4129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bagnat M, Keränen S, Shevchenko A, Shevchenko A, Simons K. Lipid rafts function in biosynthetic delivery of proteins to the cell surface in yeast. Proceedings of the National Academy of Sciences. 2000;97:3254–3259. doi: 10.1073/pnas.060034697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bagnat M, Simons K. Cell surface polarization during yeast mating. Proceedings of the National Academy of Sciences. 2002a;99:14183–14188. doi: 10.1073/pnas.172517799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bagnat M, Simons K. Lipid rafts in protein sorting and cell polarity in budding yeast Saccharomyces cerevisiae. Biological Chemistry. 2002b;383:1475–1480. doi: 10.1515/BC.2002.169. [DOI] [PubMed] [Google Scholar]
- Bakht O, London E. Cholesterol precursors stabilize ordinary and ceramide-rich ordered lipid domains (lipid rafts) to different degrees Implications for the Bloch hypothesis and sterol biosynthesis disorders. Journal of Biological Chemistry. 2006;281:21903–21913. doi: 10.1074/jbc.M600395200. [DOI] [PubMed] [Google Scholar]
- Belisle JT, Brandt ME, Radolf JD, Norgard MV. Fatty acids of Treponema pallidum and Borrelia burgdorferi lipoproteins. Journal of Bacteriology. 1994;176:2151–2157. doi: 10.1128/jb.176.8.2151-2157.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ben-Menachem G, Kubler-Kielb J, Coxon B, Yergey A, Schneerson R. A newly discovered cholesteryl galactoside from Borrelia burgdorferi. Proceedings of the National Academy of Sciences. 2003;100:7913–7918. doi: 10.1073/pnas.1232451100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benach JL, Bosler EM, Hanrahan JP, Coleman JL, Habicht GS, Bast TF, … Burgdorfer W. Spirochetes isolated from the blood of two patients with Lyme disease. New England Journal of Medicine. 1983;308:740–742. doi: 10.1056/NEJM198303313081302. [DOI] [PubMed] [Google Scholar]
- Björkbom A, Ohvo-Rekilä H, Kankaanpää P, Nyholm TK, Westerlund B, Slotte JP. Characterization of membrane properties of inositol phosphorylceramide. Biochimica et Biophysica Acta (BBA)-Biomembranes. 2010;1798:453–460. doi: 10.1016/j.bbamem.2009.11.003. [DOI] [PubMed] [Google Scholar]
- Brooks CS, Vuppala SR, Jett AM, Akins DR. Identification of Borrelia burgdorferi outer surface proteins. Infection and Immunity. 2006;74:296–304. doi: 10.1128/IAI.74.1.296-304.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown DA. Lipid rafts, detergent-resistant membranes, and raft targeting signals. Physiology. 2006;21:430–439. doi: 10.1152/physiol.00032.2006. [DOI] [PubMed] [Google Scholar]
- Brown DA, London E. Structure of detergent-resistant membrane domains: does phase separation occur in biological membranes? Biochemical and Biophysical Research Communications. 1997;240:1–7. doi: 10.1006/bbrc.1997.7575. [DOI] [PubMed] [Google Scholar]
- Brown DA, Rose JK. Sorting of GPI-anchored proteins to glycolipid-enriched membrane subdomains during transport to the apical cell surface. Cell. 1992;68:533–544. doi: 10.1016/0092-8674(92)90189-j. [DOI] [PubMed] [Google Scholar]
- Calhoun WI, Shipley GG. Sphingomyelin-lecithin bilayers and their interaction with cholesterol. Biochemistry. 1979;18:1717–1722. doi: 10.1021/bi00576a013. [DOI] [PubMed] [Google Scholar]
- Chapman D, Owens N, Phillips M, Walker D. Mixed monolayers of phospholipids and cholesterol. Biochimica et Biophysica Acta (BBA)-Biomembranes. 1969;183:458–465. doi: 10.1016/0005-2736(69)90160-6. [DOI] [PubMed] [Google Scholar]
- Chayakulkeeree M, Johnston SA, Oei JB, Lev S, Williamson PR, Wilson CF, … Meyer W. SEC14 is a specific requirement for secretion of phospholipase B1 and pathogenicity of Cryptococcus neoformans. Molecular Microbiology. 2011;80:1088–1101. doi: 10.1111/j.1365-2958.2011.07632.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chrisman CJ, Albuquerque P, Guimaraes AJ, Nieves E, Casadevall A. Phospholipids trigger Cryptococcus neoformans capsular enlargement during interactions with amoebae and macrophages. PLoS pathogens. 2011;7:e1002047. doi: 10.1371/journal.ppat.1002047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coelho C, Bocca AL, Casadevall A. The tools for virulence of Cryptococcus neoformans. Advances in Applied Microbiology. 2014;87:1–41. doi: 10.1016/B978-0-12-800261-2.00001-3. [DOI] [PubMed] [Google Scholar]
- Cox GM, Harrison TS, McDade HC, Taborda CP, Heinrich G, Casadevall A, Perfect JR. Superoxide dismutase influences the virulence of Cryptococcus neoformans by affecting growth within macrophages. Infection and immunity. 2003;71:173–180. doi: 10.1128/IAI.71.1.173-180.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crowley JT, Toledo AM, LaRocca TJ, Coleman JL, London E, Benach JL. Lipid exchange between Borrelia burgdorferi and host cells. PLoS pathogens. 2013;9:e1003109. doi: 10.1371/journal.ppat.1003109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Almeida RF, Fedorov A, Prieto M. Sphingomyelin/phosphatidylcholine/cholesterol phase diagram: boundaries and composition of lipid rafts. Biophysical Journal. 2003;85:2406–2416. doi: 10.1016/s0006-3495(03)74664-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Del Poeta M, Nimrichter L, Rodrigues ML, Luberto C. Synthesis and biological properties of fungal glucosylceramide. PLoS Pathogens. 2014;10:e1003832. doi: 10.1371/journal.ppat.1003832. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dennis EA. Formation and characterization of mixed micelles of the nonionic surfactant Triton X-100 with egg, dipalmitoyl, and dimyristoyl phosphatidylcholines. Archives of Biochemistry and Biophysics. 1974;165:764–773. doi: 10.1016/0003-9861(74)90305-1. [DOI] [PubMed] [Google Scholar]
- Dermine JF, Duclos S, Garin J, St-Louis F, Rea S, Parton RG, Desjardins M. Flotillin-1-enriched lipid raft domains accumulate on maturing phagosomes. Journal of Biological Chemistry. 2001;276:18507–18512. doi: 10.1074/jbc.M101113200. [DOI] [PubMed] [Google Scholar]
- Dinges MM, Orwin PM, Schlievert PM. Exotoxins of Staphylococcus aureus. Clinical Microbiology Reviews. 2000;13:16–34. doi: 10.1128/cmr.13.1.16-34.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Donovan C, Bramkamp M. Characterization and subcellular localization of a bacterial flotillin homologue. Microbiology. 2009;155:1786–1799. doi: 10.1099/mic.0.025312-0. [DOI] [PubMed] [Google Scholar]
- Douglas LM, Konopka JB. Fungal membrane organization: the eisosome concept. Annual Review of Microbiology. 2014;68:377–393. doi: 10.1146/annurev-micro-091313-103507. [DOI] [PubMed] [Google Scholar]
- Enright MC, Robinson DA, Randle G, Feil EJ, Grundmann H, Spratt BG. The evolutionary history of methicillin-resistant Staphylococcus aureus (MRSA) Proceedings of the National Academy of Sciences. 2002;99:7687–7692. doi: 10.1073/pnas.122108599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Farnoud AM, Mor V, Singh A, Del Poeta M. Inositol phosphosphingolipid phospholipase C1 regulates plasma membrane ATPase (Pma1) stability in Cryptococcus neoformans. FEBS Letters. 2014;588:3932–3938. doi: 10.1016/j.febslet.2014.09.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fastenberg ME, Shogomori H, Xu X, Brown DA, London E. Exclusion of a transmembrane-type peptide from ordered-lipid domains (rafts) detected by fluorescence quenching: extension of quenching analysis to account for the effects of domain size and domain boundaries. Biochemistry. 2003;42:12376–12390. doi: 10.1021/bi034718d. [DOI] [PubMed] [Google Scholar]
- Field KA, Holowka D, Baird B. Structural aspects of the association of FcεRI with detergent-resistant membranes. Journal of Biological Chemistry. 1999;274:1753–1758. doi: 10.1074/jbc.274.3.1753. [DOI] [PubMed] [Google Scholar]
- Garcia-Monco JC, Seidman RJ, Benach JL. Experimental immunization with Borrelia burgdorferi induces development of antibodies to gangliosides. Infection and Immunity. 1995;63:4130–4137. doi: 10.1128/iai.63.10.4130-4137.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- George KS, Wu S. Lipid raft: a floating island of death or survival. Toxicology and Applied Pharmacology. 2012;259:311–319. doi: 10.1016/j.taap.2012.01.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghannoum MA, Rice LB. Antifungal agents: mode of action, mechanisms of resistance, and correlation of these mechanisms with bacterial resistance. Clinical Microbiology Reviews. 1999;12:501–517. doi: 10.1128/cmr.12.4.501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghannoum MA, Spellberg BJ, Ibrahim AS, Ritchie JA, Currie B, Spitzer ED, … Casadevall A. Sterol composition of Cryptococcus neoformans in the presence and absence of fluconazole. Antimicrobial Agents and Chemotherapy. 1994;38:2029–2033. doi: 10.1128/aac.38.9.2029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gong X, Chang A. A mutant plasma membrane ATPase, Pma1-10, is defective in stability at the yeast cell surface. Proceedings of the National Academy of Sciences. 2001;98:9104–9109. doi: 10.1073/pnas.161282998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grant CW, Wu SHW, McConnell HM. Lateral phase separations in binary lipid mixtures: correlation between spin label and freeze-fracture electron microscopic studies. Biochimica et Biophysica Acta (BBA)-Biomembranes. 1974;363:151–158. doi: 10.1016/0005-2736(74)90055-8. [DOI] [PubMed] [Google Scholar]
- Grassme H, Jendrossek V, Riehle A, Von Kürthy G, Berger J, Schwarz H, … Gulbins E. Host defense against Pseudomonas aeruginosa requires ceramide-rich membrane rafts. Nature Medicine. 2003;9:322–330. doi: 10.1038/nm823. [DOI] [PubMed] [Google Scholar]
- Haque M, Hirai Y, Yokota K, Oguma K. Steryl glycosides: a characteristic feature of the Helicobacter spp.? Journal of Bacteriology. 1995;177:5334–5337. doi: 10.1128/jb.177.18.5334-5337.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harder T, Scheiffele P, Verkade P, Simons K. Lipid domain structure of the plasma membrane revealed by patching of membrane components. The Journal of Cell Biology. 1998;141:929–942. doi: 10.1083/jcb.141.4.929. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hicks DA, Nalivaeva NN, Turner AJ. Lipid rafts and Alzheimer’s disease: protein-lipid interactions and perturbation of signaling. Frontiers in Physiology. 2012;3 doi: 10.3389/fphys.2012.00189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hirai Y, Haque M, Yoshida T, Yokota K, Yasuda T, Oguma K. Unique cholesteryl glucosides in Helicobacter pylori: composition and structural analysis. Journal of Bacteriology. 1995;177:5327–5333. doi: 10.1128/jb.177.18.5327-5333.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hjort Ipsen J, Karlström G, Mourtisen O, Wennerström H, Zuckermann M. Phase equilibria in the phosphatidylcholine-cholesterol system. Biochimica et Biophysica Acta (BBA)-Biomembranes. 1987;905:162–172. doi: 10.1016/0005-2736(87)90020-4. [DOI] [PubMed] [Google Scholar]
- Holyoak C, Stratford M, McMullin Z, Cole M, Crimmins K, Brown A, Coote P. Activity of the plasma membrane H (+)-ATPase and optimal glycolytic flux are required for rapid adaptation and growth of Saccharomyces cerevisiae in the presence of the weak-acid preservative sorbic acid. Applied and Environmental Microbiology. 1996;62:3158–3164. doi: 10.1128/aem.62.9.3158-3164.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang J, Buboltz JT, Feigenson GW. Maximum solubility of cholesterol in phosphatidylcholine and phosphatidylethanolamine bilayers. Biochimica et Biophysica Acta (BBA)-Biomembranes. 1999;1417:89–100. doi: 10.1016/s0005-2736(98)00260-0. [DOI] [PubMed] [Google Scholar]
- Huijbregts RP, de Kroon AI, de Kruijff B. Topology and transport of membrane lipids in bacteria. Biochimica et Biophysica Acta (BBA)-Reviews on Biomembranes. 2000;1469:43–61. doi: 10.1016/s0304-4157(99)00014-3. [DOI] [PubMed] [Google Scholar]
- Insenser M, Nombela C, Molero G, Gil C. Proteomic analysis of detergent-resistant membranes from Candida albicans. Proteomics. 2006;6:S74–S81. doi: 10.1002/pmic.200500465. [DOI] [PubMed] [Google Scholar]
- Jones JD, Bourell KW, Norgard MV, Radolf JD. Membrane topology of Borrelia burgdorferi and Treponema pallidum lipoproteins. Infection and Immunity. 1995;63:2424–2434. doi: 10.1128/iai.63.7.2424-2434.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karnovsky MJ, Kleinfeld AM, Hoover RL, Klausner RD. The concept of lipid domains in membranes. The Journal of Cell Biology. 1982;94:1–6. doi: 10.1083/jcb.94.1.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kenedy MR, Lenhart TR, Akins DR. The role of Borrelia burgdorferi outer surface proteins. FEMS Immunology & Medical Microbiology. 2012;66:1–19. doi: 10.1111/j.1574-695X.2012.00980.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kleemann W, Grant C, McConnell H. Lipid phase separations and protein distribution in membranes. Journal of Supramolecular Structure. 1974;2:609–616. doi: 10.1002/jss.400020508. [DOI] [PubMed] [Google Scholar]
- Klose C, Ejsing CS, García-Sáez AJ, Kaiser HJ, Sampaio JL, Surma MA, … Simons K. Yeast lipids can phase-separate into micrometer-scale membrane domains. Journal of Biological Chemistry. 2010;285:30224–30232. doi: 10.1074/jbc.M110.123554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koynova R, Caffrey M. Phases and phase transitions of the phosphatidylcholines. Biochimica et Biophysica Acta (BBA)-Reviews on Biomembranes. 1998;1376:91–145. doi: 10.1016/s0304-4157(98)00006-9. [DOI] [PubMed] [Google Scholar]
- Ladbrooke BD, Williams RM, Chapman D. Studies on lecithin-cholesterol-water interactions by differential scanning calorimetry and X-ray diffraction. Biochimica et Biophysica Acta-Biomembranes. 1968;150:333–340. doi: 10.1016/0005-2736(68)90132-6. [DOI] [PubMed] [Google Scholar]
- Lange Y, Swaisgood M, Ramos B, Steck T. Plasma membranes contain half the phospholipid and 90% of the cholesterol and sphingomyelin in cultured human fibroblasts. Journal of Biological Chemistry. 1989;264:3786–3793. [PubMed] [Google Scholar]
- Langhorst MF, Reuter A, Stuermer C. Scaffolding microdomains and beyond: the function of reggie/flotillin proteins. Cellular and Molecular Life Sciences CMLS. 2005;62:2228–2240. doi: 10.1007/s00018-005-5166-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- LaRocca TJ, Crowley JT, Cusack BJ, Pathak P, Benach J, London E, … Benach JL. Cholesterol lipids of Borrelia burgdorferi form lipid rafts and are required for the bactericidal activity of a complement-independent antibody. Cell Host & Microbe. 2010;8:331–342. doi: 10.1016/j.chom.2010.09.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- LaRocca TJ, Pathak P, Chiantia S, Toledo A, Silvius JR, Benach JL, London E. Proving lipid rafts exist: membrane domains in the prokaryote Borrelia burgdorferi have the same properties as eukaryotic lipid rafts. PLoS Pathogens. 2013;9:e1003353. doi: 10.1371/journal.ppat.1003353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lattif AA, Mukherjee PK, Chandra J, Roth MR, Welti R, Rouabhia M, Ghannoum MA. Lipidomics of Candida albicans biofilms reveals phase-dependent production of phospholipid molecular classes and role for lipid rafts in biofilm formation. Microbiology. 2011;157:3232–3242. doi: 10.1099/mic.0.051086-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee A. Lipid phase transitions and phase diagrams II. Mixtures involving lipids. Biochimica et Biophysica Acta (BBA)-Reviews on Biomembranes. 1977;472:285–344. doi: 10.1016/0304-4157(77)90001-6. [DOI] [PubMed] [Google Scholar]
- Lee VT, Schneewind O. Protein secretion and the pathogenesis of bacterial infections. Genes & Development. 2001;15:1725–1752. doi: 10.1101/gad.896801. [DOI] [PubMed] [Google Scholar]
- Lefebvre B, Furt F, Hartmann MA, Michaelson LV, Carde JP, Sargueil-Boiron F, … Bessoule JJ. Characterization of lipid rafts from Medicago truncatula root plasma membranes: a proteomic study reveals the presence of a raft-associated redox system. Plant Physiology. 2007;144:402–418. doi: 10.1104/pp.106.094102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levental I, Grzybek M, Simons K. Greasing their way: lipid modifications determine protein association with membrane rafts. Biochemistry. 2010;49:6305–6316. doi: 10.1021/bi100882y. [DOI] [PubMed] [Google Scholar]
- Lin M, Rikihisa Y. Ehrlichia chaffeensis and Anaplasma phagocytophilum lack genes for lipid A biosynthesis and incorporate cholesterol for their survival. Infection and Immunity. 2003;71:5324–5331. doi: 10.1128/IAI.71.9.5324-5331.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin Q, London E. Transmembrane protein (Perfringolysin O) association with ordered membrane domains (rafts) depends upon the raft-associating properties of protein-bound sterol. Biophysical Journal. 2013;105:2733–2742. doi: 10.1016/j.bpj.2013.11.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Linden CD, Fox CF. Membrane physical state and function. Accounts of Chemical Research. 1975;8:321–327. [Google Scholar]
- Liu C, Liu GY, Song Y, Yin F, Hensler ME, Jeng W, … Oldfield E. A cholesterol biosynthesis inhibitor blocks Staphylococcus aureus virulence. Science. 2008;319:1391–1394. doi: 10.1126/science.1153018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Livermore B, Bey R, Johnson R. Lipid metabolism of Borrelia hermsi. Infection and Immunity. 1978;20:215–220. doi: 10.1128/iai.20.1.215-220.1978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lo H, Köhler JR, DiDomenico B, Loebenberg D, Cacciapuoti A, Fink GR. Nonfilamentous C. albicans mutants are avirulent. Cell. 1997;90:939–949. doi: 10.1016/s0092-8674(00)80358-x. [DOI] [PubMed] [Google Scholar]
- London E. How principles of domain formation in model membranes may explain ambiguities concerning lipid raft formation in cells. Biochimica et Biophysica Acta (BBA)-Molecular Cell Research. 2005;1746:203–220. doi: 10.1016/j.bbamcr.2005.09.002. [DOI] [PubMed] [Google Scholar]
- London E, Brown DA. Insolubility of lipids in triton X-100: physical origin and relationship to sphingolipid/cholesterol membrane domains (rafts) Biochimica et Biophysica Acta (BBA)-Biomembranes. 2000;1508:182–195. doi: 10.1016/s0304-4157(00)00007-1. [DOI] [PubMed] [Google Scholar]
- López D, Kolter R. Functional microdomains in bacterial membranes. Genes & Development. 2010;24:1893–1902. doi: 10.1101/gad.1945010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Loura L, Castanho MA, Fedorov A, Prieto M. A photophysical study of the polyene antibiotic filipin: Self-aggregation and filipin–ergosterol interaction. Biochimica et Biophysica Acta (BBA)-Biomembranes. 2001;1510:125–135. doi: 10.1016/s0005-2736(00)00341-2. [DOI] [PubMed] [Google Scholar]
- Luberto C, Martinez-Mariño B, Taraskiewicz D, Bolaños B, Chitano P, Toffaletti DL, … Balish E. Identification of App1 as a regulator of phagocytosis and virulence of Cryptococcus neoformans. Journal of Clinical Investigation. 2003;112:1080–1094. doi: 10.1172/JCI18309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Macdonald JL, Pike LJ. A simplified method for the preparation of detergent-free lipid rafts. Journal of Lipid Research. 2005;46:1061–1067. doi: 10.1194/jlr.D400041-JLR200. [DOI] [PubMed] [Google Scholar]
- Martin SW, Konopka JB. Lipid raft polarization contributes to hyphal growth in Candida albicans. Eukaryotic Cell. 2004;3:675–684. doi: 10.1128/EC.3.3.675-684.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mielich-Süss B, Schneider J, Lopez D. Overproduction of flotillin influences cell differentiation and shape in Bacillus subtilis. mBio. 2013;4:e00719–00713. doi: 10.1128/mBio.00719-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miyake Y, Kozutsumi Y, Nakamura S, Fujita T, Kawasaki T. Serine palmitoyltransferase is the primary target of a sphingosine-like immunosuppressant, ISP-1/myriocin. Biochemical and Biophysical Research Communications. 1995;211:396–403. doi: 10.1006/bbrc.1995.1827. [DOI] [PubMed] [Google Scholar]
- Mollinedo F. Lipid raft involvement in yeast cell growth and death. Frontiers in Oncology. 2012;2 doi: 10.3389/fonc.2012.00140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mollinedo F, Fernández M, Hornillos V, Delgado J, Amat-Guerri F, Acuña AU, … Ceña V. Involvement of lipid rafts in the localization and dysfunction effect of the antitumor ether phospholipid edelfosine in mitochondria. Cell Death & Disease. 2011;2:e158. doi: 10.1038/cddis.2011.41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nelms PK, Larson O, Barnes-Josiah D. Time to B. cereus about hot chocolate. Public Health Reports. 1997;112:240. [PMC free article] [PubMed] [Google Scholar]
- Nichols CB, Fraser JA, Heitman J. PAK kinases Ste20 and Pak1 govern cell polarity at different stages of mating in Cryptococcus neoformans. Molecular Biology of the Cell. 2004;15:4476–4489. doi: 10.1091/mbc.E04-05-0370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nicola AM, Robertson EJ, Albuquerque P, da Silveira Derengowski L, Casadevall A. Nonlytic exocytosis of Cryptococcus neoformans from macrophages occurs in vivo and is influenced by phagosomal pH. mBio. 2011;2:e00167–00111. doi: 10.1128/mBio.00167-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oggioni MR, Pozzi G, Valensin PE, Galieni P, Bigazzi C. Recurrent septicemia in an immunocompromised patient due to probiotic strains of Bacillus subtilis. Journal of Clinical Microbiology. 1998;36:325–326. doi: 10.1128/jcm.36.1.325-326.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Östberg Y, Berg S, Comstedt P, Wieslander Å, Bergström S. Functional analysis of a lipid galactosyltransferase synthesizing the major envelope lipid in the Lyme disease spirochete Borrelia burgdorferi. FEMS Microbiology Letters. 2007;272:22–29. doi: 10.1111/j.1574-6968.2007.00728.x. [DOI] [PubMed] [Google Scholar]
- Palma-Guerrero J, Huang IC, Jansson HB, Salinas J, Lopez-Llorca L, Read N. Chitosan permeabilizes the plasma membrane and kills cells of Neurospora crassa in an energy dependent manner. Fungal Genetics and Biology. 2009;46:585–594. doi: 10.1016/j.fgb.2009.02.010. [DOI] [PubMed] [Google Scholar]
- Palma-Guerrero J, Lopez-Jimenez J, Pérez-Berná A, Huang I, Jansson H, Salinas J, … Lopez-Llorca L. Membrane fluidity determines sensitivity of filamentous fungi to chitosan. Molecular Microbiology. 2010;75:1021–1032. doi: 10.1111/j.1365-2958.2009.07039.x. [DOI] [PubMed] [Google Scholar]
- Papahadjopoulos D, Jacobson K, Nir S, Isac I. Phase transitions in phospholipid vesicles fluorescence polarization and permeability measurements concerning the effect of temperature and cholesterol. Biochimica et Biophysica Acta (BBA)-Biomembranes. 1973;311:330–348. doi: 10.1016/0005-2736(73)90314-3. [DOI] [PubMed] [Google Scholar]
- Parsons JB, Rock CO. Bacterial lipids: Metabolism and membrane homeostasis. Progress in Lipid Research. 2013;52:249–276. doi: 10.1016/j.plipres.2013.02.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pasrija R, Panwar SL, Prasad R. Multidrug transporters CaCdr1p and CaMdr1p of Candida albicans display different lipid specificities: both ergosterol and sphingolipids are essential for targeting of CaCdr1p to membrane rafts. Antimicrobial Agents and Chemotherapy. 2008;52:694–704. doi: 10.1128/AAC.00861-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pearson CL, Xu K, Sharpless KE, Harris SD. MesA, a novel fungal protein required for the stabilization of polarity axes in Aspergillus nidulans. Molecular Biology of the Cell. 2004;15:3658–3672. doi: 10.1091/mbc.E03-11-0803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pike LJ. The challenge of lipid rafts. Journal of Lipid Research. 2009;50:S323–S328. doi: 10.1194/jlr.R800040-JLR200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Radolf JD, Goldberg MS, Bourell K, Baker SI, Jones JD, Norgard MV. Characterization of outer membranes isolated from Borrelia burgdorferi, the Lyme disease spirochete. Infection and Immunity. 1995;63:2154–2163. doi: 10.1128/iai.63.6.2154-2163.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramage G, Martínez JP, Lopez-Ribot JL. Candida biofilms on implanted biomaterials: a clinically significant problem. FEMS Yeast Research. 2006;6:979–986. doi: 10.1111/j.1567-1364.2006.00117.x. [DOI] [PubMed] [Google Scholar]
- Ramage G, VandeWalle K, Bachmann SP, Wickes BL, López-Ribot JL. In vitro pharmacodynamic properties of three antifungal agents against preformed Candida albicans biofilms determined by time-kill studies. Antimicrobial Agents and Chemotherapy. 2002;46:3634–3636. doi: 10.1128/AAC.46.11.3634-3636.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Recktenwald DJ, McConnell HM. Phase equilibriums in binary mixtures of phosphatidylcholine and cholesterol. Biochemistry. 1981;20:4505–4510. doi: 10.1021/bi00518a042. [DOI] [PubMed] [Google Scholar]
- Redfern DA, Gericke A. Domain formation in phosphatidylinositol monophosphate/phosphatidylcholine mixed vesicles. Biophysical Journal. 2004;86:2980–2992. doi: 10.1016/S0006-3495(04)74348-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rittershaus PC, Kechichian TB, Allegood JC, Merrill AH, Hennig M, Luberto C, Del Poeta M. Glucosylceramide synthase is an essential regulator of pathogenicity of Cryptococcus neoformans. The Journal of Clinical Investigation. 2006;116:1651–1659. doi: 10.1172/JCI27890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rohmer M, Bouvier P, Ourisson G. Molecular evolution of biomembranes: structural equivalents and phylogenetic precursors of sterols. Proceedings of the National Academy of Sciences. 1979;76:847–851. doi: 10.1073/pnas.76.2.847. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sáenz JP, Sezgin E, Schwille P, Simons K. Functional convergence of hopanoids and sterols in membrane ordering. Proceedings of the National Academy of Sciences. 2012;109:14236–14240. doi: 10.1073/pnas.1212141109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sankaram MB, Thompson TE. Interaction of cholesterol with various glycerophospholipids and sphingomyelin. Biochemistry. 1990;29:10670–10675. doi: 10.1021/bi00499a014. [DOI] [PubMed] [Google Scholar]
- Schroder NW, Schombel U, Heine H, Gobel UB, Zahringer U, Schumann RR. Acylated cholesteryl galactoside as a novel immunogenic motif in Borrelia burgdorferi sensu stricto. J Biol Chem. 2003;278:33645–33653. doi: 10.1074/jbc.M305799200. [DOI] [PubMed] [Google Scholar]
- Schroeder R, London E, Brown D. Interactions between saturated acyl chains confer detergent resistance on lipids and glycosylphosphatidylinositol (GPI)-anchored proteins: GPI-anchored proteins in liposomes and cells show similar behavior. Proceedings of the National Academy of Sciences. 1994;91:12130–12134. doi: 10.1073/pnas.91.25.12130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schroeder RJ, Ahmed SN, Zhu Y, London E, Brown DA. Cholesterol and sphingolipid enhance the Triton X-100 insolubility of glycosylphosphatidylinositol-anchored proteins by promoting the formation of detergent-insoluble ordered membrane domains. Journal of Biological Chemistry. 1998;273:1150–1157. doi: 10.1074/jbc.273.2.1150. [DOI] [PubMed] [Google Scholar]
- Segal BH, DeCarlo ES, Kwon-Chung K, Malech HL, Gallin JI, Holland SM. Aspergillus nidulans infection in chronic granulomatous disease. Medicine. 1998;77:345–354. doi: 10.1097/00005792-199809000-00004. [DOI] [PubMed] [Google Scholar]
- Shea JM, Kechichian TB, Luberto C, Del Poeta M. The cryptococcal enzyme inositol phosphosphingolipid-phospholipase C confers resistance to the antifungal effects of macrophages and promotes fungal dissemination to the central nervous system. Infection and immunity. 2006;74:5977–5988. doi: 10.1128/IAI.00768-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shimshick EJ, McConnell HM. Lateral phase separation in phospholipid membranes. Biochemistry. 1973;12:2351–2360. doi: 10.1021/bi00736a026. [DOI] [PubMed] [Google Scholar]
- Shukla S, Saini P, Jha S, Ambudkar SV, Prasad R. Functional characterization of Candida albicans ABC transporter Cdr1p. Eukaryotic cell. 2003;2:1361–1375. doi: 10.1128/EC.2.6.1361-1375.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Siafakas AR, Wright LC, Sorrell TC, Djordjevic JT. Lipid rafts in Cryptococcus neoformans concentrate the virulence determinants phospholipase B1 and Cu/Zn superoxide dismutase. Eukaryotic cell. 2006;5:488–498. doi: 10.1128/EC.5.3.488-498.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Silvius JR, del Giudice D, Lafleur M. Cholesterol at different bilayer concentrations can promote or antagonize lateral segregation of phospholipids of differing acyl chain length. Biochemistry. 1996;35:15198–15208. doi: 10.1021/bi9615506. [DOI] [PubMed] [Google Scholar]
- Simon-Plas F, Perraki A, Bayer E, Gerbeau-Pissot P, Mongrand S. An update on plant membrane rafts. Current opinion in plant biology. 2011;14:642–649. doi: 10.1016/j.pbi.2011.08.003. [DOI] [PubMed] [Google Scholar]
- Simonetti N, Strippoli V, Cassone A. Yeast-mycelial conversion induced by N-acetyl-D-glucosamine in Candida albicans. Nature. 1974;250:344–346. doi: 10.1038/250344a0. [DOI] [PubMed] [Google Scholar]
- Simons K, Ikonen E. Functional rafts in cell membranes. Nature. 1997;387:569–572. doi: 10.1038/42408. [DOI] [PubMed] [Google Scholar]
- Simons K, Toomre D. Lipid rafts and signal transduction. Nature reviews Molecular cell biology. 2000;1:31–39. doi: 10.1038/35036052. [DOI] [PubMed] [Google Scholar]
- Simons K, Van Meer G. Lipid sorting in epithelial cells. Biochemistry. 1988;27:6197–6202. doi: 10.1021/bi00417a001. [DOI] [PubMed] [Google Scholar]
- Singer SJ, Nicolson GL. The fluid mosaic model of the structure of cell membranes. Science. 1972;175:720–731. doi: 10.1126/science.175.4023.720. [DOI] [PubMed] [Google Scholar]
- Singh A, Wang H, Silva LC, Na C, Prieto M, Futerman AH, … Del Poeta M. Methylation of glycosylated sphingolipid modulates membrane lipid topography and pathogenicity of Cryptococcus neoformans. Cellular Microbiology. 2011;14:500–516. doi: 10.1111/j.1462-5822.2011.01735.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Skibbens JE, Roth MG, Matlin KS. Differential extractability of influenza virus hemagglutinin during intracellular transport in polarized epithelial cells and nonpolar fibroblasts. The Journal of cell biology. 1989;108:821–832. doi: 10.1083/jcb.108.3.821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smejkal GB, Hoff HF. Filipin staining of lipoproteins in polyacrylamide gels: sensitivity and photobleaching of the fluorophore and its use in a double staining method. Electrophoresis. 1994;15:922–925. doi: 10.1002/elps.11501501133. [DOI] [PubMed] [Google Scholar]
- Smith PF. Biosynthesis of cholesteryl glucoside by Mycoplasma gallinarum. Journal of bacteriology. 1971;108:986–991. doi: 10.1128/jb.108.3.986-991.1971. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stubs G, Fingerle V, Wilske B, Gobel UB, Zahringer U, Schumann RR, Schroder NW. Acylated cholesteryl galactosides are specific antigens of borrelia causing lyme disease and frequently induce antibodies in late stages of disease. Journal of Biological Chemistry. 2009;284:13326–13334. doi: 10.1074/jbc.M809575200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stubs G, Fingerle V, Zahringer U, Schumann RR, Rademann J, Schroder NW. Acylated cholesteryl galactosides are ubiquitous glycolipid antigens among Borrelia burgdorferi sensu lato. FEMS Immunology and Medical Microbiology. 2011;63:140–143. doi: 10.1111/j.1574-695X.2011.00827.x. [DOI] [PubMed] [Google Scholar]
- Sudbery P, Gow N, Berman J. The distinct morphogenic states of Candida albicans. Trends in Microbiology. 2004;12:317–324. doi: 10.1016/j.tim.2004.05.008. [DOI] [PubMed] [Google Scholar]
- Suzuki KG. Lipid rafts generate digital-like signal transduction in cell plasma membranes. Biotechnology Journal. 2012;7:753–761. doi: 10.1002/biot.201100360. [DOI] [PubMed] [Google Scholar]
- Tagliari L, Toledo MS, Lacerda TG, Suzuki E, Straus AH, Takahashi HK. Membrane microdomain components of Histoplasma capsulatum yeast forms, and their role in alveolar macrophage infectivity. Biochimica et Biophysica Acta (BBA)-Biomembranes. 2012;1818:458–466. doi: 10.1016/j.bbamem.2011.12.008. [DOI] [PubMed] [Google Scholar]
- Takesako K, Kuroda H, Inoue T, Haruna F, Yoshikawa Y, Kato I, … Yamaguchi H. Biological properties of aureobasidin A, a cyclic depsipeptide antifungal antibiotic. The Journal of Antibiotics. 1993;46:1414–1420. doi: 10.7164/antibiotics.46.1414. [DOI] [PubMed] [Google Scholar]
- Takeshita N, Diallinas G, Fischer R. The role of flotillin FloA and stomatin StoA in the maintenance of apical sterol-rich membrane domains and polarity in the filamentous fungus Aspergillus nidulans. Molecular Microbiology. 2012;83:1136–1152. doi: 10.1111/j.1365-2958.2012.07996.x. [DOI] [PubMed] [Google Scholar]
- Takeshita N, Higashitsuji Y, Konzack S, Fischer R. Apical sterol-rich membranes are essential for localizing cell end markers that determine growth directionality in the filamentous fungus Aspergillus nidulans. Molecular Biology of the Cell. 2008;19:339–351. doi: 10.1091/mbc.E07-06-0523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thompson T, Tillack TW. Organization of glycosphingolipids in bilayers and plasma membranes of mammalian cells. Annual Review of Biophysics and Biophysical Chemistry. 1985;14:361–386. doi: 10.1146/annurev.bb.14.060185.002045. [DOI] [PubMed] [Google Scholar]
- Tilly K, Bestor A, Dulebohn DP, Rosa PA. OspC-independent infection and dissemination by host-adapted Borrelia burgdorferi. Infection and Immunity. 2009;77:2672–2682. doi: 10.1128/IAI.01193-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tilly K, Bestor A, Rosa PA. Lipoprotein succession in Borrelia burgdorferi: similar but distinct roles for OspC and VlsE at different stages of mammalian infection. Molecular Microbiology. 2013;89:216–227. doi: 10.1111/mmi.12271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Toledo A, Crowley JT, Coleman JL, LaRocca TJ, Chiantia S, London E, Benach JL. Selective association of outer surface lipoproteins with the lipid rafts of Borrelia burgdorferi. mBio. 2014;5:e00899–00814. doi: 10.1128/mBio.00899-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Toulmay A, Prinz WA. Direct imaging reveals stable, micrometer-scale lipid domains that segregate proteins in live cells. The Journal of Cell Biology. 2013;202:35–44. doi: 10.1083/jcb.201301039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Triantafilou M, Miyake K, Golenbock DT, Triantafilou K. Mediators of innate immune recognition of bacteria concentrate in lipid rafts and facilitate lipopolysaccharide-induced cell activation. Journal of Cell Science. 2002;115:2603–2611. doi: 10.1242/jcs.115.12.2603. [DOI] [PubMed] [Google Scholar]
- Trott DJ, Alt DP, Zuerner RL, Wannemuehler MJ, Stanton TB. The search for Brachyspira outer membrane proteins that interact with the host. Animal Health Research Reviews. 2001;2:19–30. [PubMed] [Google Scholar]
- Veatch SL, Keller SL. Separation of liquid phases in giant vesicles of ternary mixtures of phospholipids and cholesterol. Biophysical Journal. 2003;85:3074–3083. doi: 10.1016/S0006-3495(03)74726-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vist MR, Davis JH. Phase equilibria of cholesterol/dipalmitoylphosphatidylcholine mixtures: deuterium nuclear magnetic resonance and differential scanning calorimetry. Biochemistry. 1990;29:451. doi: 10.1021/bi00454a021. [DOI] [PubMed] [Google Scholar]
- Wang J, Megha &, London E. Relationship between sterol/steroid structure and participation in ordered lipid domains (lipid rafts): implications for lipid raft structure and function. Biochemistry. 2004;43:1010–1018. doi: 10.1021/bi035696y. [DOI] [PubMed] [Google Scholar]
- Wang L, Jia Y, Tang RJ, Xu Z, Cao YB, Jia XM, Jiang YY. Proteomic analysis of Rta2p-dependent raft-association of detergent-resistant membranes in Candida albicans. PLoS One. 2012;7:e37768. doi: 10.1371/journal.pone.0037768. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wheeler CM, Garcia Monco JC, Benach JL, Golightly MG, Habicht GS, Steere AC. Nonprotein antigens of Borrelia burgdorferi. Journal of Infectious Diseases. 1993;167:665–674. doi: 10.1093/infdis/167.3.665. [DOI] [PubMed] [Google Scholar]
- Wu SH, McConnell HM. Phase separations in phospholipid membranes. Biochemistry. 1975;14:847–854. doi: 10.1021/bi00675a032. [DOI] [PubMed] [Google Scholar]
- Xu X, Bittman R, Duportail G, Heissler D, Vilcheze C, London E. Effect of the structure of natural sterols and sphingolipids on the formation of ordered sphingolipid/sterol domains (rafts) Comparison of cholesterol to plant, fungal, and disease-associated sterols and comparison of sphingomyelin, cerebrosides, and ceramide. Journal of Biological Chemistry. 2001;276:33540–33546. doi: 10.1074/jbc.M104776200. [DOI] [PubMed] [Google Scholar]
- Xu X, London E. The effect of sterol structure on membrane lipid domains reveals how cholesterol can induce lipid domain formation. Biochemistry. 2000;39:843–849. doi: 10.1021/bi992543v. [DOI] [PubMed] [Google Scholar]
- Yaqoob P, Shaikh SR. The nutritional and clinical significance of lipid rafts. Current Opinion in Clinical Nutrition & Metabolic Care. 2010;13:156–166. doi: 10.1097/MCO.0b013e328335725b. [DOI] [PubMed] [Google Scholar]
- Yepes A, Schneider J, Mielich B, Koch G, García-Betancur JC, Ramamurthi KS, … López D. The biofilm formation defect of a Bacillus subtilis flotillin-defective mutant involves the protease FtsH. Molecular Microbiology. 2012;86:457–471. doi: 10.1111/j.1365-2958.2012.08205.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yuan C, Furlong J, Burgos P, Johnston LJ. The size of lipid rafts: an atomic force microscopy study of ganglioside GM1 domains in sphingomyelin/DOPC/cholesterol membranes. Biophysical Journal. 2002;82:2526–2535. doi: 10.1016/S0006-3495(02)75596-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zidovetzki R, Levitan I. Use of cyclodextrins to manipulate plasma membrane cholesterol content: evidence, misconceptions and control strategies. Biochimica et Biophysica Acta (BBA)-Biomembranes. 2007;1768:1311–1324. doi: 10.1016/j.bbamem.2007.03.026. [DOI] [PMC free article] [PubMed] [Google Scholar]