

Abstract
Background
In patients with asthma, the fungus Aspergillus fumigatus can cause allergic bronchopulmonary aspergillosis (ABPA). Familial ABPA is reported, and some genetic factors have been associated with the disease, however, these are small studies (n ≤ 38) and do not explain all cases of ABPA.
Methods
We analysed SNPs in 95 ABPA patients, comparing frequencies to 152 atopic asthmatic and 279 healthy controls. Twenty two genes were selected from literature, and 195 tagging SNPs were analysed for genetic association with ABPA using logistic regression corrected for multiple testing. We also analysed monocyte-derived macrophage gene expression before and during co-culture with A. fumigatus.
Results
Seventeen ABPA-associated SNPs (ABPA v Atopic asthma) were identified. Three remained significant after correction for multiple testing; IL13 rs20541, IL4R rs3024656, TLR3 rs1879026. We also identified minor differences in macrophage gene expression responses in the ABPA group compared to the control groups.
Conclusions
Multiple SNPs are now associated with ABPA. Some are novel associations. These associations implicate cytokine pathways and receptors in the aberrant response to A. fumigatus and susceptibility to ABPA, providing insights into the pathogenesis of ABPA and/or its complications. We hope these results will lead to increased understanding and improved treatment and diagnostics for ABPA.
Electronic supplementary material
The online version of this article (doi:10.1186/s13223-016-0152-y) contains supplementary material, which is available to authorized users.
Keywords: ABPA, Asthma, Genetic susceptibility, TLR3, IL4R, IL13
Background
The fungus Aspergillus fumigatus is ubiquitous and humans inhale several hundred conidia each day [1]. Most overtly immunocompetent individuals clear A. fumigatus conidia without infection or sequelae, however, some asthmatic patients develop allergic bronchopulmonary aspergillosis (ABPA) following inhalation and airway colonisation [1, 2]. The exact numbers affected is unclear, but if systemically sought, ABPA is identified in 1–8 % of asthmatics seen in hospital referral clinics [3, 4]. Presentation usually involves poorly controlled asthma, wheezing, expectoration of brown mucus plugs, and ‘pneumonia’. Elevated total blood IgE levels and IgE reactivity to A. fumigatus is observed in patients, and A. fumigatus is often isolated from sputum [1]. Often central bronchiectasis and mucoid impaction of bronchi with distal atelectasis occurs. Untreated ABPA can result in pulmonary fibrosis and eventually respiratory failure [1].
The human immune response to A. fumigatus involves many cell types, including macrophages and neutrophils. These phagocytose and kill the fungus and produce chemotactic and proinflammatory cytokines to continue and orchestrate the immune response. As macrophages are present in the airways they may be the first innate immune cell to contact inhaled fungi.
In ABPA patients, an allergic Th2 response develops on exposure to A. fumigatus. Stimulation of PBMCs with Aspergillus results in production of Th2 cytokines IL5 and IL13, and ABPA patients show increased Aspergillus-induced IL5 and IL13, and decreased IFNγ production, compared to healthy controls [5]. IgE production, eosinophil recruitment, and production of an abnormal host inflammatory response in the bronchi and bronchioles of the lungs is observed [6]. This is followed by excessive mucin production, eosinophil infiltration of the bronchial mucin, and development of the features of ABPA [1]. Why some asthmatic individuals develop ABPA while others are unaffected by exposure to A. fumigatus remains unclear, despite studies. In unsensitised mice, chronic intranasal administration of mould spores or extracts can lead to allergic lung inflammation, hyper-reactivity and lung remodelling [7], but the effect in humans is unclear. Cases of ABPA within families suggest a common genetic basis with low penetrance [8, 9]. In one case series from India, 5 % of cases were found to be familial in nature [10]. The structural gene CFTR has been previously associated with ABPA [11], and small genetic association studies (involving ≤38 patients) have identified associations between ABPA and SNPs in the immune genes IL4R, IL10, TLR9, SFTPA2 and HLA (HLA-DR) [12–16], however, these results do not explain all cases of ABPA, and the immune mechanisms that underlie ABPA remain unclear.
To advance our understanding of immune responses and genetic susceptibility to ABPA, we conducted a much larger genetic association study, involving almost 100 ABPA patients and using atopic asthmatic patients as controls, as well as a second control of healthy subjects. Candidate genes included known A. fumigatus recognition receptors (dectin-1, TLR2, TLR4 and TLR9 [17–20]) and other possible recognition receptors (TLR1, TLR3 and TLR6 [21, 22]), downstream response genes, many of which are upregulated in response to A. fumigatus (IL1α, IL1β, IL1RN, IL1RAP, IL6, IL10, IL17A, TNF-α, CCL2, TGFB1, PTX3) [20, 23], mannose binding lectin (MBL) and plasminogen, both of which bind A. fumigatus [23, 24], other Th1 (TNF-α, IL15) and Th2 (IL4, IL5, IL13, CCL17) cytokines [25, 26] and signal transducer and activator of transcription-3 (STAT3), which mediates IL-10 anti-inflammatory functions in macrophages and neutrophils [27]. Other immune genes not previously investigated in Aspergillus infection, such as DENN/MADD domain containing 1B (DENND1B), a negative regulator of the TNF-α receptor, and adenosine A2a receptor (ADORA2A) were also included [28]. In addition, we analysed expression of some of these immune genes in monocyte-derived macrophages (MDMs) at baseline and during co-culture with live A. fumigatus, comparing expression in cells derived from patients to those from control individuals.
Methods
Subjects
ABPA subjects, asthmatic controls and healthy controls were defined as described in Table 1. Both atopic (but fungally non-atopic) asthmatic and non-atopic asthmatic controls were recruited. ABPA subjects complicated by CCPA were excluded. Only Caucasian subjects were used. ABPA patients were recruited from the National Aspergillosis Centre [University Hospital of South Manchester (UHSM), UK] tertiary referral clinic from March 2006 to August 2010. Previous recruited healthy and asthmatic subjects were used as controls [29, 30]. The Local Research Ethics Committee approved the study and all participants gave informed consent. The healthy control subjects have been used as controls for previous studies into genetic susceptibility to CCPA [31, 32].
Table 1.
Diagnostic criteria for recruited subjects
| Disease | Diagnostic criteria |
|---|---|
| ABPA |
All the following are required
Total serum IgE 1000 IU/ml (at any time) Either positive SPT for Aspergillus or Aspergillus specific IgE Current or historical evidence of eosinophilia Further indicators Almost all patients have asthma (or cystic fibrosis, n = 3) and over 50 % central bronchiectasis on CT, but these were not required for inclusion Either recurrent obstruction (mucoid impaction) or episodes coughing up plugs of thick mucus (containing hyphae and eosinophils) Positive Aspergillus precipitins or raised Aspergillus IgG titre |
| Atopic (non-fungally atopic) asthmatic |
All the following are required
Physician diagnosed asthma No diagnosis of aspergillosis Negative SPT (at 3 mm cut-off) and/or IgE (<0.4) to all fungi tested, including Alternaria alternata, Candida albicans, Cladosporium herbarum, Penicillium notatum, Trichophyton rubrum, A. fumigatus Positive SPT (at 3 mm cut-off) and/or IgE (<0.4) to any allergen non-fungal allergen tested (e.g. mite, cat, dog and grasses) Note Only SPT or IgE need be completed, but if both are done and one is positive this is classed as a positive result |
| Non-atopic asthmatic |
All the following are required
Physician diagnosed asthma No diagnosis of aspergillosis Negative SPT (at 3 mm cut-off) and/or IgE (<0.4) to all allergens tested, including mite, cat, dog and grasses Negative SPT (at 3 mm cut-off) and/or IgE (<0.4) to all fungi tested, including Alternaria alternata, Candida albicans, Cladosporium herbarum, Penicillium notatum, Trichophyton rubrum, A. fumigatus Note: Only SPT or IgE need be completed, but if both are done and one is positive this is classed as a positive result |
| Healthy control |
All the following are required
No diagnosis of asthma No diagnosis of aspergillosis Negative SPT (at 3 mm cut-off) and/or IgE (<0.4) to all allergens tested, including mite, cat, dog and grasses Negative SPT (at 3 mm cut-off) and/or IgE (<0.4) to all fungi tested, including Alternaria alternata, Candida albicans, Cladosporium herbarum, Penicillium notatum, Trichophyton rubrum, A. fumigatus |
DNA and PBMC extraction from blood
For DNA, blood was collected in EDTA-treated blood collection tubes (Becton Dickinson; BD, Oxford, UK). This was centrifuged to separate the plasma and cellular sections and then DNA was then extracted from the cellular section using a phenol chloroform extraction method. Both the plasma and DNA were stored at −80 °C. For the previously recruited subjects, DNA had been collected previously [30].
For PBMC extraction, blood (≤80 ml) was collected in sodium heparin treated blood collection tubes (BD) then layered onto Ficoll-paque Plus (GE Lifesciences, Buckingham, UK) in a 1:1 ratio and centrifuged (23 °C, 0.5 h, 480g). The PBMC layer was transferred into growth media (RPMI 1640 with l-glutamine and NaHCO3, with 10 % heat inactivated FBS, 100 units/ml penicillin, and 0.1 mg/ml of streptomycin [Sigma-Aldrich Company Ltd, Dorset, UK]) and centrifuged to pellet the PBMCs (24 °C, 10 min, 390g). These PBMCs were frozen in freezing media [Heat inactivated FBS (Life Technologies Ltd, Paisley, UK) with 5 % DMSO Hybri-max (Sigma-Aldrich)] at a concentration of ~1 × 107 cells/ml. Frozen PBMCs were transferred to liquid nitrogen for long term storage. Recovery and viability after freezing were assessed by trypan blue staining and found to be high (normally >90 %).
Gene and SNP selection, genotyping, quality control and data analysis
Twenty two biologically plausible and previously associated candidate genes with immune functions were selected from the literature (Additional file 1: Table S1). These included genes involved in immune recognition and response, especially those involved in recognition and response to fungus specifically. It did not include the CFTR gene, which has structural functions.
A total of 253 haplotype tagging SNPs within these genes of interest were selected for genotyping, using the Genome Variation Server (GVS, http://gvs.gs.washington.edu/GVS/) (Additional file 1: Table S1). These were usually selected to encompass the entire gene, plus 2500 bp up- and 1500 bp down-stream.
Genotyping was completed on 237 SNPs using the Sequenom® MassArray® iPLEX™ Gold system. SNPs with Hardy–Weinberg Equilibrium p < 0.0001 or call rates <90 % were excluded from the analysis. After this, subjects with call rates <90 % were excluded from the analysis. Genotyping was completed in two rounds. In the first round, genotyping was completed successfully in 95 ABPA, 279 healthy and 152 atopic asthmatic subjects; an additional 14 atopic asthmatic subjects were genotyped successfully in the second round. Results were analysed using SNP and Variation Suite (SVS; version 7.4.3, Golden Helix). Redundant SNPs (r2 > 0.80) were excluded from analysis after evaluation of the LD within our population, as were SNPs that were monomorphic within our population. Statistical analysis was completed using Stata (Statacorp). Logistic regression was used to determine association for the remaining 195 SNPs using dominant and recessive models. Atopic asthmatics were used as controls for this analysis. Correction for multiple testing was completed using the Benjamini–Hochberg correction for False Discovery Rate [33]. A p value of p < 0.05 was considered significant. The p values were calculated using the R software [34]. The widely used Benjamini–Hochberg correction for False Discovery Rate is less stringent than the Bonferroni correction in which p values are multiplied by the number of comparisons. The Benjamini–Hochberg correction for FDR is shown to have higher power compared to other procedures including the Bonferroni method for multiple testing [35]. For SNPs associated with ABPA in the comparison to asthmatic subjects, a genetic association test was completed to identify the p value for the comparison to healthy subjects.
Macrophage-A. fumigatus co-culture
Ten each of ABPA, non-atopic asthmatic and healthy subjects were selected for the macrophage-A. fumigatus co-culture experiment. MDMs were generated (approx. 2 × 105/well), live A. fumigatus conidia added (4 × 105/well) and RNA extracted as described previously [31]. Previous work shows that during the incubation period, proportions of the different fungal morphologies change, and that the time points of 0–3, 6 and 9 h are representative of exposure to conidia, germ tubes and hyphae respectively [31]. Experiments were repeated in triplicate.
Measuring expression by MDMs
The human innate and adaptive immune responses RT2 profiler PCR array (SABiosciences) was used to measure expression of various genes related to those we genotyped in pooled RNA samples (1 μg) from each disease group (n = 10 subjects) using HRPT1, RPL13A and GAPDH as housekeeping normaliser genes. Arrays were repeated in triplicate. The investigated genes included TLRs (TLR1, 2, 3, 4, 9, 10) and related receptors (TREM1) and intracellular signalling genes (IRAK2, TRAF6, MYD88). For the majority of the arrays, the three housekeeping genes HRPT1, RPL13A and GAPDH were used as the normalizer genes, however, HPRT1 was found to be variable in the healthy 9 h samples and so only RPL13A and GAPDH were used for the normalization of this experiment. Results were available for only two replicates of the asthmatic 6 h time point due to technical difficulties. The data was analysed using the manufacturer’s online data analysis tool (http://pcrdataanalysis.sabiosciences.com/pcr/arrayanalysis.php) to calculate fold changes relative to a calibrator.
Statistical analysis
Statistical analysis was completed in Stata, SPSS (Version 16; SPSS Inc.) and GraphPad Prism (Version 5.02; GraphPad Software Inc.). Ages and % males were compared between the groups using Mann-Whitney tests as the data was not normally distributed. Expression data was analysed using T-tests and repeated measures one-way ANOVA, using data from the triplicate arrays. Standard deviation was calculated for these. Data for TLR1 was highly variable between replicates for the healthy group and this was excluded from analysis. As the one-way ANOVA required three replicates, the 6 h time point was excluded from this analysis of the asthma samples.
Results
Characteristics of study participants
The characteristics of the subjects recruited for genotyping are shown in Table 2. All are Caucasian. Those subjects with ABPA tended to be older than other groups (p < 0.0001), and had poorer lung function (% predicted FEV1, % predicted FVC or FEV1/FVC ratio, all p < 0.0001) (Table 2). Of the 97 ABPA, 280 healthy, 167 atopic asthmatic subjects recruited, 95 ABPA, 279 healthy and 166 atopic asthmatic subjects were successfully genotyped.
Table 2.
Characteristics of patients and controls recruited
| Characteristic | ABPA | Atopic asthma | Healthy |
|---|---|---|---|
| n | 97 | 167 | 280 |
| Age (year) (median, IQR) | 61.7 year (54.0–69.5) | 50.1 year (43.8–64.9) | 47.0 year (44.2–50.6) |
| % Male | 53.6 % (52/97) | 41.3 % | 40.4 % (113/280) |
| Asthma (%) | 88.7 % (86/97) | 100 % (167/167) | 0 % (0/280) |
| Lung function tests (n) | 67 | 165 | 255 |
| FEV1 % predicted (median, IQR) | 64.0 (51.0–78.0) | 96.0 (78.4–108.2) | 107.5 (99.6–117.2) |
| FVC % predicted (median, IQR) | 96.0 (87.5–106.5) | 108.9 (99.0–118.5) | 114.7 (105.6–123.6) |
| FEV1/FVC ratio (median, IQR) | 56.0 (47.9–64.0) | 72.0 (63.0–79.0) | 79.0 (75.0–83.0) |
| Markers of ABPA | |||
| Bronciectasis (%) | 67.0 % (65/97) | N/A | N/A |
| Total serum IgE (kU/L) (median, IQR) (n) | 2250 (1100–4425) (96) | N/A | N/A |
| Aspergillus specific IgE (kUA/L) (median, IQR) (n) | 9.7 (3.2–36.0) (62) | N/A | N/A |
IQR interquartile range
The characteristics of the subjects selected for the gene expression study are shown in Table 3. The ABPA group were older than the healthy group (media age 59.5 years vs. 38.0 years, p = 0.0007), but not significantly different to the non-atopic asthmatic group (Table 3).
Table 3.
Characteristics of patients and controls used for gene expression experiment
| Characteristic | ABPA | Non-atopic asthma | Healthy |
|---|---|---|---|
| n | 10 | 10 | 10 |
| Age (year) (median, IQR) | 59.5 year (55.7–66.1) | 46.0 year (37.4–57.3) | 38.0 year (31.2–51.1) |
| % Male | 40.0 % (4/10) | 20 % (2/10) | 40 % (4/10) |
| Asthma (%) | 70 % (7/10) | 100 % (10/10) | 0 % (0/10) |
| Lung function tests (n) | 5 | 9 | N/A |
| FEV1 % predicted (median, IQR) | 75.0 (71.0–80.0) | 85.0 (58.0–105.0) | N/A |
| FVC % predicted (median, IQR) | 103.0 (102.0–106.0) | 113.0 (89.0–116.0) | N/A |
| FEV1/FVC ratio (median, IQR) | 60.8 (55.0–63.4) | 78.0 (46.0–81.0) | N/A |
| Markers of ABPA | N/A | ||
| Bronciectasis (%) | 80 % (8/10) | 50 % (5/10) | N/A |
| Total serum IgE (kU/L) (median, IQR) (n) | 4100 (2650–7075) (10) | 71.9 (63.4–110.0) (10) | N/A |
| Aspergillus specific IgE (kUA/L) (median, IQR) (n) | 34.0 (2.9–60.3) (6) | N/A | N/A |
The recruitment criteria specified that the asthma control patients had a negative specific IgE or skin test to aspergillus. Therefore Aspergillus specific IgE level was not measured on every patient but was negative when measured
IQR interquartile range
Multiple SNPs are associated with ABPA
All the p values (uncorrected and FDR corrected) for all the models tested can be seen in Additional file 1: Table S2. Of the 195 SNPs analysed (Additional file 1: Table S1), 17 SNPs in nine genes were found to be associated with ABPA (p < 0.05, Table 4). Three of these survived correction for multiple testing (Table 4; Fig. 1). Compared to atopic asthmatics, ABPA patients were more likely to be carriers of the rare A allele of the exonic missense mutation in IL13 (rs20541). They were also more likely to carry the common G allele and common GG genotype of the intronic SNPs in IL4R rs3024656 and TLR3 rs1879026, respectively. Further analysis found that four of the SNPs associated with ABPA on the ABPA vs. Atopic asthma model were also associated on the ABPA vs. Healthy model (Additional file 1: Table S3). These were Dectin-1 rs7959451, IL13 rs1800925, IL4R rs3024656 and IL4R rs1029489.
Table 4.
SNPs associated with ABPA
| Gene | SNP | Alleles (M/m) | MAF (%) | Model for association | Genotype count | Odds ratio (95 % CI) | p value* | BH FDR p value* | Location | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Genotype | ABPA | Atopic asthma | Healthya | |||||||||
| ADORA2A | rs2236624 | C/T | 26 | CC + CT vs. TT | TT CC + CT |
11 84 |
7 145 |
18 261 |
0.37 (0.14–0.99) |
0.047 | 0.130 | Intronic |
| DECTIN1 | rs11053624 | T/C | 9 | CC + TC vs. TT | TT CC + CT |
72 23 |
132 20 |
233 46 |
2.11 (1.08–4.10) |
0.028 | 0.086 | 5′ near gene |
| rs7959451 | C/T | 14 | TT + CT vs. CC | CC TT + CT |
62 33 |
120 32 |
212 67 |
2.00 (1.12–3.55) |
0.018 | 0.061 | 3′ UTR | |
| IL13 | rs20541 | G/A | 20 | AA + GA vs. GG | GG AA + GA |
51 44 |
116 48 |
167 105 |
2.08 (1.23–3.53) |
0.006 | 0.025 | Exonic (R > Q): missense |
| rs1800925 | C/T | 18 | TT + TC vs. CC | CC TT + TC |
53 42 |
115 49 |
186 87 |
1.86 (1.10–3.14) |
0.021 | 0.067 | 5′ near gene | |
| IL17A | rs3819024 | A/G | 36 | GG + GA vs. AA | AA GG + GA |
33 62 |
74 78 |
115 164 |
1.78 (1.05–3.02) |
0.032 | 0.097 | 5′ near gene |
| IL4R | rs3024656 | G/A | 33 | GG + GA vs. AA | AA GG + GA |
3 92 |
22 141 |
33 239 |
4.78 (1.39–16.44) |
0.013 | 0.045 | Intronic |
| rs1029489 | G/A | 39 | AA + GA vs. GG | GG AA + GA |
24 71 |
64 100 |
113 160 |
2.00 (1.14–3.52) |
0.016 | 0.054 | 3′ near gene | |
| rs6498012 | G/C | 38 | GG + GC vs. CC | CC GG + GC |
20 75 |
19 145 |
35 237 |
0.49 (0.25–0.98) |
0.043 | 0.122 | Intronic | |
| MBL2 | rs2099903 | C/A | 25 | CC + CA vs. AA | AA CC + CA |
11 84 |
6 146 |
21 258 |
0.31 (0.11–0.88) |
0.027 | 0.086 | 3′ UTR |
| PLAT | rs8178880 | A/G | 4 | GG + AG vs. AA | AA GG + AG |
91 3 |
135 17 |
256 23 |
0.26 (0.07–0.92) |
0.036 | 0.108 | Intronic |
| PLG | rs4252053 | A/G | 15 | GG + AG vs. AA | AA GG + AG |
64 31 |
122 30 |
194 85 |
1.97 (1.10–3.54) |
0.023 | 0.075 | 5′ near gene |
| TLR3 | rs1879026 | G/T | 17 | TT + GT vs. GG | GG TT + GT |
75 20 |
94 57 |
197 82 |
0.44 (0.24–0.80) |
0.007 | 0.026 | Intronic |
| rs10025405 | A/G | 43 | GG + GA vs. AA | AA GG + GA |
26 69 |
62 90 |
95 184 |
1.83 (1.05–3.18) |
0.033 | 0.100 | Intergenic (3′ of TLR3) | |
| rs5743303 | A/T | 19 | TT + AT vs. AA | AA TT + AT |
56 39 |
112 40 |
184 95 |
1.95 (1.13–3.36) |
0.016 | 0.56 | 5′ near gene | |
| rs5743305 | T/A | 37 | AA + TA vs. TT | TT AA + TA |
48 47 |
54 98 |
117 162 |
0.54 (0.32–0.91) | 0.020 | 0.67 | 5′ near gene | |
| rs7668666 | C/A | 26 | AA + CA vs. CC | CC AA + CA |
49 46 |
99 53 |
164 115 |
1.75 (1.04–2.96) | 0.035 | 0.105 | Intronic | |
Risk allele shown in bold. SNPs in bold remain significant after Benjamini–Hochberg adjustment for false discovery rate (FDR). Benjamini–Hochberg adjusted p value shown (BH FDR p value)
CI confidence interval; M/m Major allele/Minor allele
* p value calculated for the model indicated using logistic regression in Stata (ABPA v Atopic asthma)
aGenotype frequencies in our healthy population are shown for the readers interest but are not used for analysis
Fig. 1.
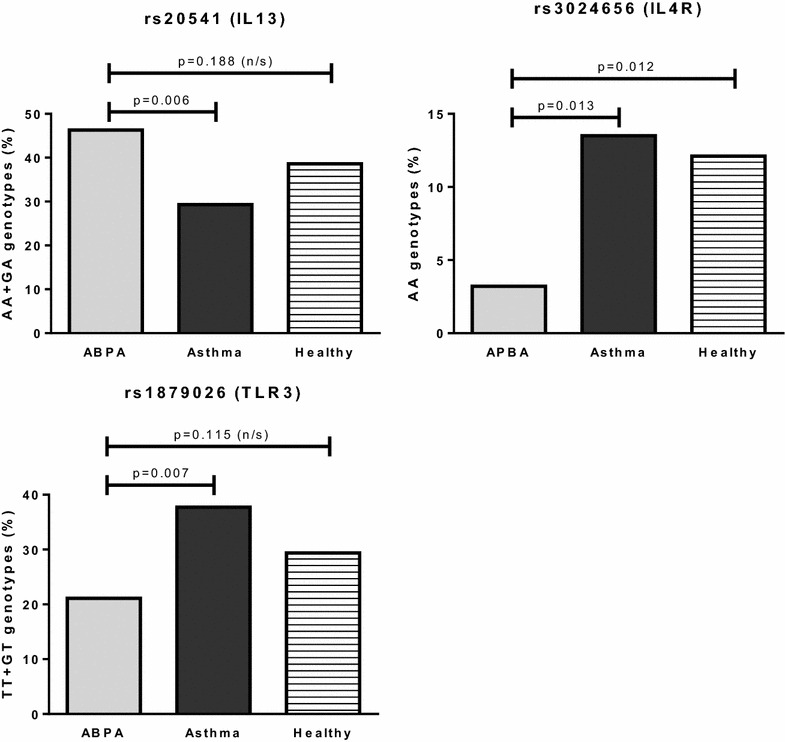
Genotype frequencies of SNPs associated with ABPA after correction for multiple testing
Expression of TLR genes varies little over time after stimulation with A. fumigatus
Expression of TLR2, TLR4, TLR6, and, to a lesser extent, TLR3, is lower in the ABPA and non-atopic asthmatic groups than in the healthy group at 6 h, and higher at 9 h (Fig. 2a–d). For TLR2 and TLR4 expression at 9 h is also significantly higher in asthma than in ABPA. The differences compared to the healthy group are a result of increased 6 h and reduced 9 h expression in the healthy subjects, which are not observed in the asthma or ABPA groups (Fig. 2e–h). The downstream signalling genes, TRAF6, IRAK2 and to a lesser extent MYD88 follow a similar expression pattern, although expression of other downstream signalling molecules does not (Additional file 1: Fig. S1). Expression of the activating receptor, triggering receptor expressed on myeloid cells 1 (TREM1) is significantly reduced in the ABPA group at baseline, compared to both the asthmatic and healthy groups, and remains lower at all time points (Fig. 3), which may suggest a reduced amplification of the TLR response in the ABPA group. Expression of further TLRs, including TLR9 and TLR10 is shown in the (Additional file 1: Fig. S2).
Fig. 2.
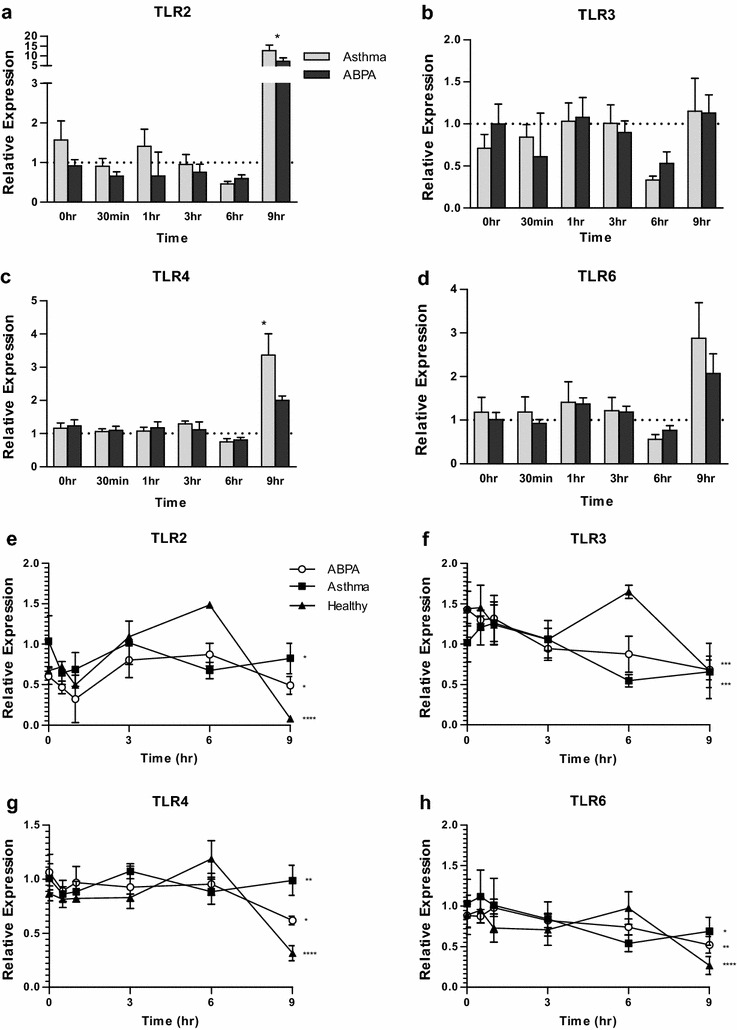
Expression of TLRs by MDMs from ABPA, asthmatic and healthy subjects (n = 10, pooled). a–d show expression in the ABPA (black bars) and asthma groups (grey bars) relative to the healthy group (dotted line) at each time point. Stars indicate significant differences between ABPA and asthma, calculated by t-test. e–h show expression in ABPA (open circles), asthmatic (closed squares) and healthy (closed triangles) subjects over time, relative to asthma 0 h. Stars indicate significant changes over time, calculated by repeated measures 1-way ANOVA. *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001. Bars indicate standard deviation of the three replicates
Fig. 3.
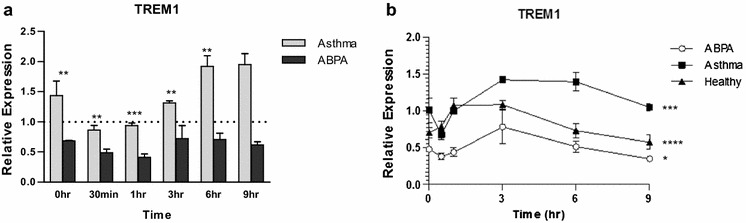
Expression of TREM1 by MDMs from ABPA, asthmatic and healthy subjects (n = 10, pooled). a shows expression in the ABPA (black bars) and asthma groups (grey bars) relative to the healthy group (dotted line) at each time point. Stars indicate significant differences between ABPA and asthma, calculated by t-test. b shows expression in ABPA (open circles), asthmatic (closed squares) and healthy (closed triangles) subjects over time, relative to asthma 0 h. Stars indicate significant changes over time, calculated by repeated measures 1-way ANOVA. *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001. Bars indicate standard deviation of the three replicates
Discussion
In the largest genetic association study of ABPA conducted to date, we have identified associations with SNPs in three immune genes, IL13 (rs20541), IL4R (rs3024656) and TLR3 (rs1879026), which remained significant after correction for multiple testing. Of these genes, only IL4R has previously been associated with ABPA; IL13 and TLR3 are novel candidate genes, which help to increase our understanding of the aberrant immune response occurring in ABPA patients. These associations now require replication in other populations to confirm their importance.
The association of SNPs in IL13 and IL4R with ABPA is interesting. IL13 and IL4 are Th2 cytokines and ABPA is known to involve a Th2 response [5]. PBMCs from ABPA patients show increased Aspergillus-induced IL-5 and IL-13 production, and decreased IFNγ production, compared to healthy controls [5]. Five SNPs in IL13 and IL4R were associated with ABPA before correction for multiple testing, and one in each remained significant after correction. One IL4R SNP has been previously associated with ABPA (rs1805010) [12]. This SNP has also been suggested to affect sensitivity to IL4 stimulation [12]. Unfortunately, this SNP failed the genotyping QC in the current study, however, a different SNP in IL4R (rs3024656) was found to be associated with ABPA. RS1805010 is a missense SNP in located in the 5′UTR of the IL4R gene (position 16:27344882), while rs3024656 is intronic (position 16:27358288). There is a very low LD between them (r2 = 0.16).
IL13 and IL4 act via the IL4R receptor and are involved in the allergic response to antigens such as those produced by A. fumigatus [5]. It has been shown that polymorphisms in IL4R and IL13 can have synergistic effects [36]. IL4 is a key cytokine required for differentiation of Th2 cells [26]. Uncontrolled Th2 are thought to be detrimental for both survival and disease progression in aspergillosis, and the identification of these SNPs supports this theory [17, 20, 37, 38].
The IL4R SNP (rs3024656) has been previously associated with only invasive squamous cell cervical cancer [39] and is not in high LD (r2 > 0.80) with any other SNPs, however, the IL13 SNP (rs20541) has been previously associated with many diseases and phenotypes, including asthma, atopy and serum IgE level [40–42]. In addition, rs20541 is in high LD with other SNPs (rs849, rs848, rs1295685, rs1295686) that have also been extensively associated with disease, including with asthma and serum IgE level [41]. As discussed, ABPA is an allergic disease often found in asthmatics, however, our use of atopic asthmatic controls should control for any association with asthma or non-fungal atopy in our population and suggests that this SNP is associated with ABPA or fungal atopy specifically. RS20541 is an exonic missense mutation in IL13, which results in an codon change from CGG to CAG, and an arginine (R) to glutamine (Q) substitution at amino acid position 110. Compared to atopic asthmatics, ABPA patients were more likely to be carriers of the rare A allele, coding for glutamine. This Q110 variant of IL13 appears to be less susceptible to clearance and more stable in plasma than the R110 variant, which may cause increased circulating levels in vivo [43]. This is supported by reports demonstrating significantly higher serum IL13 levels in human carriers of the Q110 homozygous genotype, compared to carriers of the R110 homozygous genotype [43]. As mentioned, Aspergillus-induced IL-13 is increased in PBMCs from ABPA patients compared to those from healthy controls [5]. The RS20541 A allele and the glutamine variant is also associated with elevated IgE levels [44]. IgE levels are also raised in ABPA [6]. In addition to rs20541, another SNP in IL13 (rs1800925) was found to be associated with ABPA, although this association did not survive correction for multiple testing. Like rs20541, IL13 rs1800925 has been extensively associated with atopic disease, including asthma, psoriatic arthritis and eczema [40, 42, 45, 46], as well as with allergy and IgE levels [42, 47]. It has also recently been associated with COPD [48]. The T allele of this SNP was associated with ABPA in the current study. This allele has been demonstrated to increase IL13 promoter activity in primary human and murine CD4 + Th2 cells by creation of a Yin-Yang binding site, which overlaps a STAT motif [49]. STAT6-mediated repression of IL13 transcription is reduced, allowing increased promoter activity and increased IL13 expression [49]. In addition, mitogen-activated PBMCs from humans homozygous for the T allele have been shown to secrete significantly higher levels of IL13 compared with those from CC and CT individuals [49]. The TT genotype is also associated with increased cord blood IgE [47]. The A allele (glutamine) of rs20541 and the T allele of rs1800925 may increase susceptibility to ABPA by increasing IL13 expression and increasing the allergic Th2 response, including IgE production, which is detrimental in aspergillosis. Other SNPs in IL13 and in other Th2 cytokines and receptors such as IL4 and IL4R could increase susceptibility to ABPA in a similar way.
Toll like receptors (TLRs) are known to be important in the recognition of A. fumigatus and as such it is likely that mutations in these receptors will influence susceptibility to aspergillosis. Traditionally, the most studied are TLR2, TLR4 and TLR9, however, recent work suggests that other TLRs such as TLR1, TLR3 and TLR6 may also be important [17–22]. Two SNPs in TLR1 (rs5743611, rs4833095) have been previously associated with IA, and one SNP in TLR9 (rs5743836) has been previously associated with ABPA [13, 22]. The study that identified the TLR9 SNP was small, involving only 22 ABPA patients and 88 controls [13]. None of these SNPs was found to be associated with ABPA in the current study; however, we did identify five SNPs at various locations within TLR3 as associated with ABPA. Only one of these (rs1879026) survived correction for multiple testing. This is an intronic SNP that has not been associated with disease in the past and is not in high LD (r2 > 0.80) with any other SNPs. TLR3 has only recently been implicated as a recognition receptor for A. fumigatus, with studies identifying a role in epithelial cell mediated protection against this fungus [19, 50], and the discovery of this association with ABPA is helpful in supporting this role.
We also analysed the expression of many of the TLR genes known to be expressed on human macrophages, in MDMs from healthy subjects, non-atopic asthmatics and ABPA subjects, in response to stimulation with A. fumigatus. As has been described by previous groups, we only observed small changes in gene expression over time after stimulation with A. fumigatus [19, 21, 23], however, 9 h TLR2 and TLR4 expression was significantly lower in the ABPA group compared to the asthmatic group. This could represent a differential response to the hyphal form of the fungus (predominant at the 9 h time point), or a differential time-dependant response to 9 h of culture. However, as the fold changes are low, it may also be that these differences and patterns are not biologically relevant. In addition, it may be that downstream signalling and the response to this is more important in the response to fungus than any change in TLR expression. This may account for the small changes in expression observed in the current study as well as in previous studies [19, 21, 23]. Alternatively, as the expression of some downstream signalling molecules follows a similar pattern to expression of the TLRs, there may indeed be a deficient response in the ABPA group.
The activating receptor TREM1 is found on neutrophils and monocytes and acts to amplifies inflammation induced by stimulation of TLRs, including TLR2 and TLR4 [51, 52]. Expression of TREM1 is reduced in the ABPA group, which may indicate a reduced ability to respond to TLR stimulation, including stimulation by fungi such as A. fumigatus. As TLR2 and TLR4 are key in the response to A. fumigatus, it could be that TREM1 plays a role in the response to A. fumigatus via interactions with these. We recently reported increased TREM1 expression by MDMs from CCPA subjects compared to those from healthy controls, and suggested that this might be an attempt to combat reduced baseline expression of TLRs by the CCPA group [32]. This does not appear to be the case in ABPA; in this situation, baseline TLR expression is not reduced, and increased TREM1 expression is not observed. Although both ABPA and CCPA occur in overtly immunocompetent subjects, they are very different diseases; CCPA is not considered an allergic disease and is not related to asthma. It is therefore not surprising that increased TREM1 expression is observed in MDMs from CCPA and reduced TREM1 expression is observed in ABPA. It is, however, interesting and suggests that this previously unstudied molecule may be important in susceptibility to fungal diseases and may benefit from further study.
We do not have information about the specific treatments patients were receiving at the time that blood was collected, however, because a small proportion of patients with ABPA in our clinic are prescribed corticosteroid therapy, either continuously or intermittently, it may be that some of those tested are currently taking this medication. The same is true for the asthmatic population. Systemic corticosteroid treatment can influence cell proliferation and RNA expression [53], however, as this effect of steroids on cells is temporary, we would expect any effect to be lost after the freezing, thawing and 15 days of culture that these cells undergo prior to use in our experiments.
While there remains uncertainty about the relevance of the expression profiles observed for the TLR genes, the association of the SNP in TLR3 with ABPA supports a role for this receptor in recognition of A. fumigatus and in response to this and TLR3 may affect susceptibility to ABPA, with SNPs that reduce the fungal recognition leaving people more susceptible to this disease.
We acknowledge that the alveolar macrophage could be considered as the ideal cell type in which to study interactions between host and pathogen as the initial interactions in the host are likely to occur within the lungs. However, if we had recruited only subjects that were fit for bronchoscopy we would have skewed the study population in favour of patients with mild ABPA. Collection of blood is a much less invasive procedure and is possible for subjects with all severities of ABPA; the use of MDMs therefore prevents population skewing.
We opted to use pooled samples for our gene expression work as although this prevents analysis of inter-patient variability, it allows for analysis of a greater number of genes (due to limited availability of patient cells). As has been discussed in detail previously [31], mean values for cases and controls are likely to be broadly similar to the results of pooled samples, however, we acknowledge the possibility that our pooled value could be skewed by a minority of individuals, Unfortunately, pooling also prevented us from associating gene expression with individual genotypes and therefore from identifying functional consequences of the SNPs. In addition, we felt that in the smaller gene expression study, the use of atopic asthmatic controls such as those used in the genetic association study, could present a risk as results could be skewed due to association with atopy to another non-fungal allergen (if two or three of the small control group were atopic to the same allergen), and so opted for non-atopic controls. In the genetic study, the larger control group with lots of different allergic profiles meant this was less likely to be a problem. We understand this is primarily a hypothesis generating study and further work will be required.
In addition, we acknowledge that the controls within our study are older than the cases, and this may mean that some controls may go on to develop ABPA later in life. This would reduce (rather than increase) our power to find associations with ABPA, and as such does not invalidate our findings. As a hypothesis generating study, and we appreciate that future work is required to support our results. This work will include replication of the genetic associations, functional work and experiments to confirm that gene expression differences translate to differences in protein levels. Despite this, we believe our results are important in suggesting novel genes and pathways that are associated with ABPA susceptibility and may be involved in the pathogenesis of ABPA. We hope these results are useful in providing novel directions for future research.
Conclusions
The genetic association study presented here is the most extensive study into genetic susceptibility to ABPA to date, involving three times as many patients as most previous studies and investigating many more SNPs. We have identified SNPs in candidate genes including TLR3, IL4R and IL13 that are associated with ABPA, and which help to support the involvement of these previously theorised receptors and pathways in the immune response to A. fumigatus and in susceptibility to ABPA. We hope that this increased understanding will in future lead to developments in the treatment and diagnosis of ABPA.
Authors’ contributions
AS, PB and DWD contributed to study conception and obtained funding. AS, PB, DWD and NO contributed to study design. NS contributed to acquisition of data. NO, AS, PB and DWD analysed and interpreted data. NS drafted the report, and all authors contributed to revision of the report and approved the version submitted. All authors read and approved the final manuscript.
Acknowledgements
We would like to thank the control subjects and the patients for their participation, Danielle Belgrave at the University of Manchester who provided statistical advice, and staff at the Centre for Integrated Genomics Research (CIGMR), including Dr Jenny Hankinson, who completed the genotyping.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
The dataset supporting the conclusions of this article is included within the article and its additional files.
Authors’ information
AS and DWD are clinicians working at the University Hospital of South Manchester; AS is a specialist in asthma and allergy at the North West Lung Centre, while DWD heads the National Aspergillosis Centre and is a specialist in aspergillosis. Both have strong research interests and are Honorary Professors of the University of Manchester. PB is a senior lecturer at the University of Manchester while NO is a research scientist, both studying Aspergillus and aspergillosis. All of the authors are interested in the host response to Aspergillus and in genetic susceptibility to ABPA and other forms of aspergillosis. NO was previously known as N Smith.
Consent for publication
Not applicable. This manuscript does not contain any individual person’s data.
Ethics approval and consent to participate
The Local Research Ethics Committee approved the study and all participants gave informed consent. Healthy and Asthmatic subjects were collected under REC references 05/Q1406/70, 06/Q1403/110 and 09/H1003/95, while ABPA subjects were collected under REC references 07/Q1403/70 and 08/H1003/45.
Funding
This work was supported by the European Union’s Seventh Framework Programme [FP7/2007-2013] under grant agreement n° HEALTH-2010-260338 (ALLFUN). Funding was also provided by the National Aspergillosis Centre, Medical Research Council, Fungal Infection Trust, and JP Moulton Charitable Foundation and the North West Lung Centre Charity. This report is independent research supported by the National Institute for Health Research Clinical Research Facility at University Hospital of South Manchester NHS Foundation Trust. The views expressed in this publication are those of the author(s) and not necessarily those of the NHS, the National Institute for Health Research or the Department of Health. The funders had no role in study design, data collection and interpretations, or the decision to submit the work for publication.
Abbreviations
- ABPA
allergic bronchopulmonary aspergillosis
- SNP
single nucleotide polymorphism
- IL
interleukin
- TLR
toll like receptor
- IgE
immunoglobulin E
- Th
t helper
- MDM
monocyte derived macrophage
- UHSM
University Hospital of South Manchester
- PBMCs
peripheral blood mononuclear cells
- PCR
polymerase chain reaction
- FDR
false discovery rate
Additional file
10.1186/s13223-016-0152-y Supplementary tables and figures.
Contributor Information
Nicola L. D. Overton, Email: nicola.overton@manchester.ac.uk
David W. Denning, Email: david.denning@manchester.ac.uk
Paul Bowyer, Email: paul.bowyer@manchester.ac.uk.
Angela Simpson, Phone: +44 161 291 5871, Email: angela.simpson@manchester.ac.uk.
References
- 1.Latge JP. Aspergillus fumigatus and aspergillosis. Clin Microbiol Rev. 1999;12:310–350. doi: 10.1128/cmr.12.2.310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Stevens DA, Moss RB, Kurup VP, Knutsen AP, Greenberger P, Judson MA, Denning DW, Crameri R, Brody AS, Light M, et al. Allergic bronchopulmonary aspergillosis in cystic fibrosis–state of the art: cystic Fibrosis Foundation Consensus Conference. Clin Infect Dis. 2003;37(Suppl 3):S225–S264. doi: 10.1086/376525. [DOI] [PubMed] [Google Scholar]
- 3.Agarwal R, Aggarwal AN, Gupta D, Jindal SK. Aspergillus hypersensitivity and allergic bronchopulmonary aspergillosis in patients with bronchial asthma: systematic review and meta-analysis. Int J Tuberc Lung Dis. 2009;13:936–944. [PubMed] [Google Scholar]
- 4.Denning DW, Pleuvry A, Cole DC. Global burden of allergic bronchopulmonary aspergillosis with asthma and its complication chronic pulmonary aspergillosis in adults. Med Mycol. 2013;51:361–370. doi: 10.3109/13693786.2012.738312. [DOI] [PubMed] [Google Scholar]
- 5.Becker KL, Gresnigt MS, Smeekens SP, Jacobs CW, Magis-Escurra C, Jaeger M, Wang X, Lubbers R, Oosting M, Joosten LA, et al. Pattern recognition pathways leading to a Th2 cytokine bias in ABPA patients. Clin Exp Allergy. 2015;45:423–437. doi: 10.1111/cea.12354. [DOI] [PubMed] [Google Scholar]
- 6.Soubani AO, Chandrasekar PH. The clinical spectrum of pulmonary aspergillosis. Chest. 2002;121:1988–1999. doi: 10.1378/chest.121.6.1988. [DOI] [PubMed] [Google Scholar]
- 7.Denis O, van den Brule S, Heymans J, Havaux X, Rochard C, Huaux F, Huygen K. Chronic intranasal administration of mould spores or extracts to unsensitized mice leads to lung allergic inflammation, hyper-reactivity and remodelling. Immunology. 2007;122:268–278. doi: 10.1111/j.1365-2567.2007.02636.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Halwig JM, Kurup VP, Greenberger PA, Patterson R. A familial occurrence of allergic bronchopulmonary aspergillosis: a probable environmental source. J Allergy Clin Immunol. 1985;76:55–59. doi: 10.1016/0091-6749(85)90804-8. [DOI] [PubMed] [Google Scholar]
- 9.Graves TS, Fink JN, Patterson R, Kurup VP, Scanlon GT. A familial occurrence of allergic bronchopulmonary aspergillosis. Ann Intern Med. 1979;91:378–382. doi: 10.7326/0003-4819-91-3-378. [DOI] [PubMed] [Google Scholar]
- 10.Shah A, Kala J, Sahay S, Panjabi C. Frequency of familial occurrence in 164 patients with allergic bronchopulmonary aspergillosis. Ann Allergy Asthma Immunol. 2008;101:363–369. doi: 10.1016/S1081-1206(10)60311-0. [DOI] [PubMed] [Google Scholar]
- 11.Agarwal R, Khan A, Aggarwal AN, Gupta D. Link between CFTR mutations and ABPA: a systematic review and meta-analysis. Mycoses. 2012;55:357–365. doi: 10.1111/j.1439-0507.2011.02130.x. [DOI] [PubMed] [Google Scholar]
- 12.Knutsen AP, Kariuki B, Consolino JD, Warrier MR. IL-4 alpha chain receptor (IL-4Ralpha) polymorphisms in allergic bronchopulmonary sspergillosis. Clin Mol Allergy. 2006;4:3–9. doi: 10.1186/1476-7961-4-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Carvalho A, Pasqualotto AC, Pitzurra L, Romani L, Denning DW, Rodrigues F. Polymorphisms in toll-like receptor genes and susceptibility to pulmonary aspergillosis. J Infect Dis. 2008;197:618–621. doi: 10.1086/526500. [DOI] [PubMed] [Google Scholar]
- 14.Vaid M, Kaur S, Sambatakou H, Madan T, Denning DW, Sarma PU. Distinct alleles of mannose-binding lectin (MBL) and surfactant proteins A (SP-A) in patients with chronic cavitary pulmonary aspergillosis and allergic bronchopulmonary aspergillosis. Clin Chem Lab Med. 2007;45:183–186. doi: 10.1515/CCLM.2007.033. [DOI] [PubMed] [Google Scholar]
- 15.Brouard J, Knauer N, Boelle PY, Corvol H, Henrion-Caude A, Flamant C, Bremont F, Delaisi B, Duhamel JF, Marguet C, et al. Influence of interleukin-10 on Aspergillus fumigatus infection in patients with cystic fibrosis. J Infect Dis. 2005;191:1988–1991. doi: 10.1086/429964. [DOI] [PubMed] [Google Scholar]
- 16.Chauhan B, Hutcheson PS, Slavin RG, Bellone CJ. MHC restriction in allergic bronchopulmonary aspergillosis. Front Biosci. 2003;8:s140–s148. doi: 10.2741/971. [DOI] [PubMed] [Google Scholar]
- 17.Steele C, Rapaka RR, Metz A, Pop SM, Williams DL, Gordon S, Kolls JK, Brown GD. The beta-glucan receptor dectin-1 recognizes specific morphologies of Aspergillus fumigatus. PLoS Pathog. 2005;1:e42. doi: 10.1371/journal.ppat.0010042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Meier A, Kirschning CJ, Nikolaus T, Wagner H, Heesemann J, Ebel F. Toll-like receptor (TLR) 2 and TLR4 are essential for Aspergillus-induced activation of murine macrophages. Cell Microbiol. 2003;5:561–570. doi: 10.1046/j.1462-5822.2003.00301.x. [DOI] [PubMed] [Google Scholar]
- 19.Bellocchio S, Moretti S, Perruccio K, Fallarino F, Bozza S, Montagnoli C, Mosci P, Lipford GB, Pitzurra L, Romani L. TLRs govern neutrophil activity in aspergillosis. J Immunol. 2004;173:7406–7415. doi: 10.4049/jimmunol.173.12.7406. [DOI] [PubMed] [Google Scholar]
- 20.Gersuk GM, Underhill DM, Zhu L, Marr KA. Dectin-1 and TLRs permit macrophages to distinguish between different Aspergillus fumigatus cellular states. J Immunol. 2006;176:3717–3724. doi: 10.4049/jimmunol.176.6.3717. [DOI] [PubMed] [Google Scholar]
- 21.Shin SH, Lee YH. Airborne fungi induce nasal polyp epithelial cell activation and Toll-like receptor expression. Int Arch Allergy Immunol. 2010;153:46–52. doi: 10.1159/000301578. [DOI] [PubMed] [Google Scholar]
- 22.Kesh S, Mensah NY, Peterlongo P, Jaffe D, Hsu K, VDB M, O’Reilly R, Pamer E, Satagopan J, Papanicolaou GA. TLR1 and TLR6 polymorphisms are associated with susceptibility to invasive aspergillosis after allogeneic stem cell transplantation. Ann NY Acad Sci. 2005;1062:95–103. doi: 10.1196/annals.1358.012. [DOI] [PubMed] [Google Scholar]
- 23.Loeffler J, Haddad Z, Bonin M, Romeike N, Mezger M, Schumacher U, Kapp M, Gebhardt F, Grigoleit GU, Stevanovic S, et al. Interaction analyses of human monocytes co-cultured with different forms of Aspergillus fumigatus. J Med Microbiol. 2009;58:49–58. doi: 10.1099/jmm.0.003293-0. [DOI] [PubMed] [Google Scholar]
- 24.Zaas AK, Liao G, Chien JW, Weinberg C, Shore D, Giles SS, Marr KA, Usuka J, Burch LH, Perera L, et al. Plasminogen alleles influence susceptibility to invasive aspergillosis. PLoS Genet. 2008;4:e1000101. doi: 10.1371/journal.pgen.1000101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Katakura T, Miyazaki M, Kobayashi M, Herndon DN, Suzuki F. CCL17 and IL-10 as effectors that enable alternatively activated macrophages to inhibit the generation of classically activated macrophages. J Immunol. 2004;172:1407–1413. doi: 10.4049/jimmunol.172.3.1407. [DOI] [PubMed] [Google Scholar]
- 26.Furuzawa-Carballeda J, Vargas-Rojas MI, Cabral AR. Autoimmune inflammation from the Th17 perspective. Autoimmun Rev. 2007;6:169–175. doi: 10.1016/j.autrev.2006.10.002. [DOI] [PubMed] [Google Scholar]
- 27.Kobayashi M, Kweon MN, Kuwata H, Schreiber RD, Kiyono H, Takeda K, Akira S. Toll-like receptor-dependent production of IL-12p40 causes chronic enterocolitis in myeloid cell-specific Stat3-deficient mice. J Clin Invest. 2003;111:1297–1308. doi: 10.1172/JCI17085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Sleiman PM, Flory J, Imielinski M, Bradfield JP, Annaiah K, Willis-Owen SA, Wang K, Rafaels NM, Michel S, Bonnelykke K, et al. Variants of DENND1B associated with asthma in children. N Engl J Med. 2010;362:36–44. doi: 10.1056/NEJMoa0901867. [DOI] [PubMed] [Google Scholar]
- 29.Langley SJ, Goldthorpe S, Craven M, Morris J, Woodcock A, Custovic A. Exposure and sensitization to indoor allergens: association with lung function, bronchial reactivity, and exhaled nitric oxide measures in asthma. J Allergy Clin Immunol. 2003;112:362–368. doi: 10.1067/mai.2003.1654. [DOI] [PubMed] [Google Scholar]
- 30.Marinho SF: Genetic and environmental factors in asthma and allergic phenotypes in adults. Ph.D. thesis submitted to The University of Manchester 2010.
- 31.Smith NL, Hankinson J, Simpson A, Bowyer P, Denning DW. A prominent role for the IL1 pathway and IL15 in susceptibility to chronic cavitary pulmonary aspergillosis. Clin Microbiol Infect. 2014;20:O480–O488. doi: 10.1111/1469-0691.12473. [DOI] [PubMed] [Google Scholar]
- 32.Smith NL, Hankinson J, Simpson A, Denning DW, Bowyer P. Reduced expression of TLR3, TLR10 and TREM1 by human macrophages in CCPA, and novel associations of VEGFA, DENND1B and PLAT. Clin Microbiol Infect. 2014;20:O960–O968. doi: 10.1111/1469-0691.12643. [DOI] [PubMed] [Google Scholar]
- 33.Benjamini Y, Hochberg Y. Controlling the False Discovery Rate: a Practical and Powerful Approach to Multiple Testing. J R Statist Soc B. 1995;57:289–300. [Google Scholar]
- 34.RDevelopmentCoreTeam R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria; 2012.
- 35.Lix LM, Sajobi T. Testing multiple outcomes in repeated measures designs. Psychol Methods. 2010;15:268–280. doi: 10.1037/a0017737. [DOI] [PubMed] [Google Scholar]
- 36.Chen W, Ericksen MB, Levin LS, Khurana Hershey GK. Functional effect of the R110Q IL13 genetic variant alone and in combination with IL4RA genetic variants. J Allergy Clin Immunol. 2004;114:553–560. doi: 10.1016/j.jaci.2004.04.044. [DOI] [PubMed] [Google Scholar]
- 37.Murdock BJ, Shreiner AB, McDonald RA, Osterholzer JJ, White ES, Toews GB, Huffnagle GB. Coevolution of TH1, TH2, and TH17 responses during repeated pulmonary exposure to Aspergillus fumigatus conidia. Infect Immun. 2011;79:125–135. doi: 10.1128/IAI.00508-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Hohl TM, Van Epps HL, Rivera A, Morgan LA, Chen PL, Feldmesser M, Pamer EG. Aspergillus fumigatus triggers inflammatory responses by stage-specific beta-glucan display. PLoS Pathog. 2005;1:e30. doi: 10.1371/journal.ppat.0010030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Johnson LG, Schwartz SM, Malkki M, Du Q, Petersdorf EW, Galloway DA, Madeleine MM. Risk of cervical cancer associated with allergies and polymorphisms in genes in the chromosome 5 cytokine cluster. Cancer Epidemiol Biomarkers Prev. 2011;20:199–207. doi: 10.1158/1055-9965.EPI-10-0779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Cui L, Jia J, Ma CF, Li SY, Wang YP, Guo XM, Li Q, Yu HB, Liu WH, Gao LB. IL-13 polymorphisms contribute to the risk of asthma: a meta-analysis. Clin Biochem. 2012;45:285–288. doi: 10.1016/j.clinbiochem.2011.12.012. [DOI] [PubMed] [Google Scholar]
- 41.Bottema RW, Nolte IM, Howard TD, Koppelman GH, Dubois AE, de Meer G, Kerkhof M, Bleecker ER, Meyers DA, Postma DS. Interleukin 13 and interleukin 4 receptor-alpha polymorphisms in rhinitis and asthma. Int Arch Allergy Immunol. 2010;153:259–267. doi: 10.1159/000314366. [DOI] [PubMed] [Google Scholar]
- 42.Black S, Teixeira AS, Loh AX, Vinall L, Holloway JW, Hardy R, Swallow DM. Contribution of functional variation in the IL13 gene to allergy, hay fever and asthma in the NSHD longitudinal 1946 birth cohort. Allergy. 2009;64:1172–1178. doi: 10.1111/j.1398-9995.2009.01988.x. [DOI] [PubMed] [Google Scholar]
- 43.Arima K, Umeshita-Suyama R, Sakata Y, Akaiwa M, Mao XQ, Enomoto T, Dake Y, Shimazu S, Yamashita T, Sugawara N, et al. Upregulation of IL-13 concentration in vivo by the IL13 variant associated with bronchial asthma. J Allergy Clin Immunol. 2002;109:980–987. doi: 10.1067/mai.2002.124656. [DOI] [PubMed] [Google Scholar]
- 44.Graves PE, Kabesch M, Halonen M, Holberg CJ, Baldini M, Fritzsch C, Weiland SK, Erickson RP, von Mutius E, Martinez FD. A cluster of seven tightly linked polymorphisms in the IL-13 gene is associated with total serum IgE levels in three populations of white children. J Allergy Clin Immunol. 2000;105:506–513. doi: 10.1067/mai.2000.104940. [DOI] [PubMed] [Google Scholar]
- 45.Eder L, Chandran V, Pellett F, Pollock R, Shanmugarajah S, Rosen CF, Rahman P, Gladman DD. IL13 gene polymorphism is a marker for psoriatic arthritis among psoriasis patients. Ann Rheum Dis. 2011;70:1594–1598. doi: 10.1136/ard.2010.147421. [DOI] [PubMed] [Google Scholar]
- 46.Miyake Y, Kiyohara C, Koyanagi M, Fujimoto T, Shirasawa S, Tanaka K, Sasaki S, Hirota Y. Case-control study of eczema associated with IL13 genetic polymorphisms in Japanese children. Int Arch Allergy Immunol. 2011;154:328–335. doi: 10.1159/000321825. [DOI] [PubMed] [Google Scholar]
- 47.Hong X, Tsai HJ, Liu X, Arguelles L, Kumar R, Wang G, Kuptsova-Clarkson N, Pearson C, Ortiz K, Bonzagni A, et al. Does genetic regulation of IgE begin in utero? Evidence from T(H)1/T(H)2 gene polymorphisms and cord blood total IgE. J Allergy Clin Immunol. 2010;126:1059–1067. doi: 10.1016/j.jaci.2010.08.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Arja C, Ravuri RR, Pulamaghatta VN, Surapaneni KM, Raya P, Adimoolam C, Kanala KR. Genetic determinants of chronic obstructive pulmonary disease in South Indian male smokers. PLoS One. 2014;9:e89957. doi: 10.1371/journal.pone.0089957. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Cameron L, Webster RB, Strempel JM, Kiesler P, Kabesch M, Ramachandran H, Yu L, Stern DA, Graves PE, Lohman IC, et al. Th2 cell-selective enhancement of human IL13 transcription by IL13-1112C > T, a polymorphism associated with allergic inflammation. J Immunol. 2006;177:8633–8642. doi: 10.4049/jimmunol.177.12.8633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.de Luca A, Bozza S, Zelante T, Zagarella S, D’Angelo C, Perruccio K, Vacca C, Carvalho A, Cunha C, Aversa F, Romani L. Non-hematopoietic cells contribute to protective tolerance to Aspergillus fumigatus via a TRIF pathway converging on IDO. Cell Mol Immunol. 2010;7:459–470. doi: 10.1038/cmi.2010.43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Bouchon A, Dietrich J, Colonna M. Cutting edge: inflammatory responses can be triggered by TREM-1, a novel receptor expressed on neutrophils and monocytes. J Immunol. 2000;164:4991–4995. doi: 10.4049/jimmunol.164.10.4991. [DOI] [PubMed] [Google Scholar]
- 52.Zheng H, Heiderscheidt CA, Joo M, Gao X, Knezevic N, Mehta D, Sadikot RT. MYD88-dependent and -independent activation of TREM-1 via specific TLR ligands. Eur J Immunol. 2010;40:162–171. doi: 10.1002/eji.200839156. [DOI] [PubMed] [Google Scholar]
- 53.El-Gedaily A, Schoedon G, Schneemann M, Schaffner A. Constitutive and regulated expression of platelet basic protein in human monocytes. J Leukoc Biol. 2004;75:495–503. doi: 10.1189/jlb.0603288. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The dataset supporting the conclusions of this article is included within the article and its additional files.