

Abstract
Previous studies have shown that metformin, an AMP-activated protein kinase activator widely prescribed for type 2 diabetes, is especially beneficial in cases of diabetic retinopathy (DR) with undetermined mechanisms. Here, we used a streptozotocin-induced diabetes model in mice to study the effects of metformin on the development of DR. We found that 10 weeks after STZ treatment, DR was induced in STZ-treated mice, regardless treatment of metformin. However, metformin alleviated the DR, seemingly through attenuating the retina neovascularization. The total vascular endothelial cell growth factor A (VEGF-A) in eyes was not altered by metformin, but the phosphorylation of the VEGF receptor 2 (VEGFR2) was decreased, which inhibited VEGF signaling. Further analysis showed that metformin may induce VEGF-A mRNA splicing to VEGF120 isoform to reduce its activation of the VEGFR2. These findings are critical for generating novel medicine for DR treatment.
Keywords: Diabetic retinopathy (DR), vascular endothelial cell growth factor A (VEGF-A), VEGF receptor 2 (VEGFR2), splicing, VEGF120, VEGF164
Introduction
Diabetes is a chronic disease that affect a great number of people worldwide. Among all diabetes patients, more than 90% are type 2 diabetes, which is treated by metformin as the first-line therapy [1]. Metformin is particularly more protective in the management of overweight or obese type 2 diabetes patients at high risk of developing cardiovascular complications, compared to sulphonylureas or insulin, and have protective effects against development of diabetic retinopathy (DR) [2].
Metformin is also found to reduce plasminogen activator inhibitor 1 (PAI-1), resulting in increases in fibrinolytic activity. Moreover, metformin is also found to inhibit inflammatory mediated angiogenesis in a murine model and in clinic, seemingly through a PAI-1-mediated increases in thrombospondin 1, an antiangiogenic adipokine that is highly expressed in insulin-resistant individuals with obesity [1]. In the literature, there are conflicting reports on the effects of metformin, as suggested as anti-angiogenic or pro-angiogenic, largely resulting from the difference in time course at analysis and experimental model [3]. Nevertheless, the exact molecular mechanisms are still unclear.
Vascular endothelial growth factor A (VEGF-A) plays a crucial role during organ development, regeneration and many pathological situations. Alternative VEGF mRNA splicing results in 3 protein isoforms (VEGF120, VEGF164 and VEGF188 in mice), of which VEGF164 has been shown to have the highest bioavailability and biological potency [4-6]. VEGF120, however, is freely diffusible due to the lack of heparan-sulfate proteoglycan binding domains and thus being low potent in activating VEGF Receptor 2 (VEGFR2), which is the main tyrosine kinase receptor for VEGF-A that is expressed mainly by endothelial cells to mediate most of the biological effects of VEGF-A, including blood vessel growth and branching, endothelial cell survival and vessel permeability [7,8].
Here, we used a streptozotocin-induced diabetes model in mice to study the effects of metformin on the development of DR [9-12]. We found that 10 weeks after STZ treatment, DR was induced in STZ-treated mice, regardless treatment of metformin. However, metformin reduced the DR severity, seemingly through attenuating the retina neovascularization. The total vascular endothelial cell growth factor A (VEGF-A) in eyes was not altered by metformin, but the phosphorylation of the VEGF receptor 2 (VEGFR2) was decreased, which inhibited VEGF signaling. Further analysis showed that metformin may induce VEGF-A mRNA splicing to VEGF120 isoform to reduce its activation of the VEGFR2. These findings are critical for generating novel medicine for DR treatment.
Materials and methods
Mouse manipulation
All mouse experiments were approved by the Animal Research and Care Committee at the Ningbo Eye Hospital. Male C57BL/6 mice were purchased from the Jackson Lab (Bar Harbor, Maine, USA), and were used in the experiments at 10 weeks of age. Fasting blood glucose monitoring were performed at 9am in the morning. The beta cell toxin streptozotocin (STZ, Sigma-Aldrich, St Louis, MO, USA) was injected intraperitoneally at 90 mg/kg body weight, which allowed development of sustained high blood glucose and long survival of the animals. Two week after STZ treatment, mice in the metformin group were administered an intraperitoneal injection of metformin (120 mg/kg body weight) every day for 10 weeks. Mice in the saline-treated group mice were administered an intraperitoneal injection of an equivalent volume of normal saline at same frequency. The mice were analyzed at 8 or 10 weeks after metformin (or control), or 10 or 12 weeks after STZ.
Fluorescein angiography and analysis of retina vessel density
At the time of sacrifice, mice were anesthetized by 2% isoflurane inhalation, and fluorescein angiography images were obtained after intraperitoneal injection of 0.15 ml of 2% fluorescein sodium (Alcon Laboratories, Inc., Fort Worth, TX, USA). Mouse eyeballs were harvested and immersed in 4% paraformaldehyde at room temperature for 2 hour. Afterwards, sclera, uveal tissue, cornea, and lens were removed from the retina. The isolated retina was washed twice with phosphate buffered saline for 5 minutes. Retinal vessels were stained by storing the retina in tomato lectin (Molecular Probes, Carlsbad, CA, USA) at 4°C for 2 days. Stained retinal tissues were flat-mounted on glass slides with coverslips.
Retinopathy grading
Avascular areas were evaluated using Image J (NIH, Bethesda, MA, USA) by two experienced investigators blinded to group-identifying information. Retinopathy grade was assessed by two experienced investigators using the scoring system proposed described before [13]. This scoring system incorporates assessments of blood vessel growth, blood vessel tufts, extra-retinal neovascularization, central vasoconstriction, retinal hemorrhage, and degree of vessel tortuosity.
Enzyme-linked immunosorbent assay (ELISA) for VEGF-A
Whole eyes were homogenized in PBS and sonicated. After centrifugation, supernatants containing most soluble proteins were collected. VEGF-A levels in the supernatant were determined using an ELISA kit (RayBiotech, Inc., Norcross GA, USA). Absorbance values were obtained at 450-570 nm using an Emax spectrophotometer (Molecular Devices, Sunnyvale, CA, USA).
RNA isolation and RT-qPCR
Total RNA was extracted from mouse eyes with Trizol (Invitrogen, Carlsbad, California, USA). RNA was quantified with Nanodrop1000 (Thermo Scientific, Rockford, IL, USA) according to the manufacturer’s instructions, followed by cDNA synthesis (Qiagen, Valencia, CA, USA). Quantitative PCR primers were shown in the figure. RT-qPCR Reactions were performed in triplicates with QuantiTect SYBR Green PCR Kit (Qiagen). Specificity of the amplified products was determined by melting peak analysis. Quantification was performed with the 2-ΔΔCt method. Values of genes were normalized against tubulin, which proved to be stable across the samples.
Immunohistochemistry
All samples are fixed in 4% paraformaldehyde at room temperature for 2 hour, then cryo-protected in 30% sucrose overnight before further approaches. DAB staining was performed with a DAB chromogen system (Dako, Carpinteria, CA, USA). Primary antibodies for immunostaining are: guinea pig polyclonal anti-insulin (Dako) and rat polyclonal anti-CD31 (Becton-Dickinson Biosciences, San Jose, CA, USA). No antigen retrieval is necessary for these antigens. Secondary antibodies for indirect fluorescent staining are Cy2-conjugated antibodies generated from donkey (Jackson Immuno Research Labs, West Grove, PA, USA).
Western blot
Protein was extracted using RIPA buffer (Sigma-Aldrich) for Western Blot. The supernatants were collected after centrifugation at 12000×g at 4°C for 20 min. Protein concentration was determined using BCA protein assay, and whole lysates were mixed with 4×SDS loading buffer at a ratio of 1:3. Samples were heated at 100°C for 5 min and were separated on SDS-polyacrylamide gels. The separated proteins were then transferred to a PVDF membrane. The membrane blots were first probed with a primary antibody. After incubation with horseradish peroxidase-conjugated second antibody, autoradiograms were prepared using the enhanced chemiluminescent system to visualize the protein antigen. The signals were recorded using X-ray film. Primary antibodies for Western Blot are anti-phosphorylated VEGFR2 (pVEGFR2), anti-VEGFR2 (pIGF1R) and anti-α-tubulin (all purchased from Cell Signaling, San Jose, CA, USA). Secondary antibody is HRP-conjugated anti-rabbit (Jackson ImmunoResearch Labs). Images shown in the figure were representative from 5 repeats. Densitometry of Western blots was quantified with ImageJ software (NIH).
Statistical analysis
All statistical analyses were carried out using GraphPad prism 6.0 (GraphPad Software Inc. La Jolla, CA, USA). All values are depicted as mean ± SD and are considered significant if p<0.05. All data were statistically analyzed using one-way ANOVA with a Bonferroni correction, followed by Fisher’s Exact Test for comparison of two groups.
Results
Experimental model
In order to study the effects of metformin on DR, we used STZ to induce diabetes in mice. The beta cell toxin STZ was injected intraperitoneally at 90 mg/kg body weight, which allowed development of sustained high blood glucose and long survival of the animals. Two week after STZ treatment, mice in the metformin group (STZ+Met) were administered an intraperitoneal injection of metformin (120 mg/kg body weight) every day for 10 weeks. Mice in the saline-treated group mice (STZ) were administered an intraperitoneal injection of an equivalent volume of normal saline at same frequency. The mice were analyzed at 8 weeks after metformin treatment for Retinopathy score, and at 10 weeks after metformin treatment for retina vessel density and mechanisms (Figure 1A).
Figure 1.
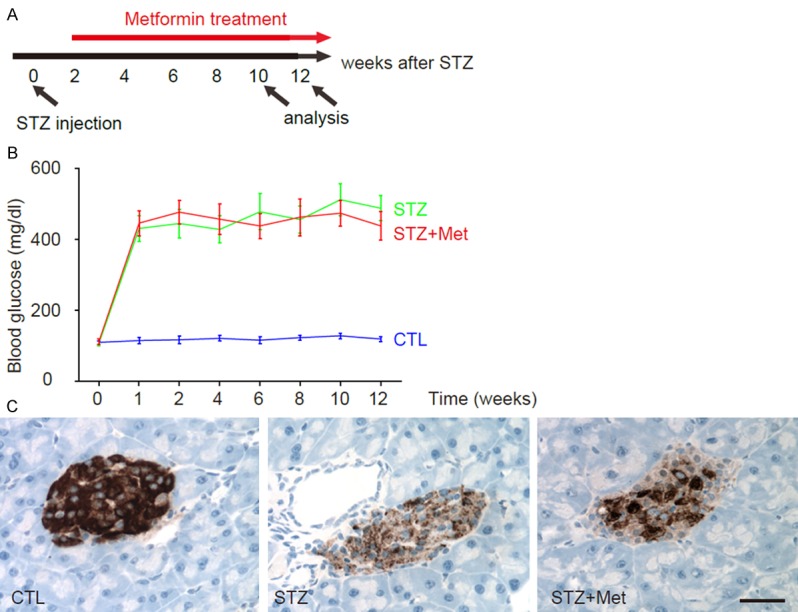
Experimental model and Induction of diabetes in mice by STZ administration. A: Experimental model. The beta cell toxin STZ was injected. Two week after STZ treatment, mice in the metformin group (STZ+Met) were administered an intraperitoneal injection of metformin every day for 10 weeks. Mice in the saline-treated group mice (STZ) were administered an intraperitoneal injection of an equivalent volume of normal saline at same frequency. The mice that did not received STZ and metformin were used as negative controls (STZ). The mice were analyzed at 8 weeks after metformin treatment for Retinopathy score, and at 10 weeks after metformin treatment for retina vessel density and mechanisms. B: Fasting blood glucose. C: The analysis of pancreas sections at 12 weeks after STZ showed significant loss of islet beta cells in both groups, based on insulin immunostaining. *p<0.05. N=10. Scale bar is 20 µm.
Induction of diabetes in mice by STZ administration
We found that ever since 1 week after STZ injection, the mice developed sustained hyperglycemia, in both STZ and STZ+Met groups. There are no differences in the levels of high blood glucose between two groups (Figure 1B). The analysis of pancreas sections at 12 weeks after STZ showed significant loss of islet beta cells in both groups, based on insulin immunostaining (Figure 1C). Thus, STZ induces diabetes in mice and metformin does not revert hyperglycemia.
Induction of DR in mice by STZ administration
Then, we examined whether STZ may induce DR in these diabetic mice. We found that 10 weeks after STZ treatment, DR was induced in STZ-treated mice, regardless treatment of metformin, based on retinopathy scoring (Figure 2A). However, metformin seemed to alleviate the severity of DR (Figure 2A). Interestingly, metformin treatment did not alter the avascular area in eyes at this point, shown by quantification (Figure 2B), and by representative images (Figure 2C), suggesting that metformin may not affect retinopathy at the initial ischemic period. Thus, we hypothesize that metformin may affect the neovascularization in the development of DR.
Figure 2.
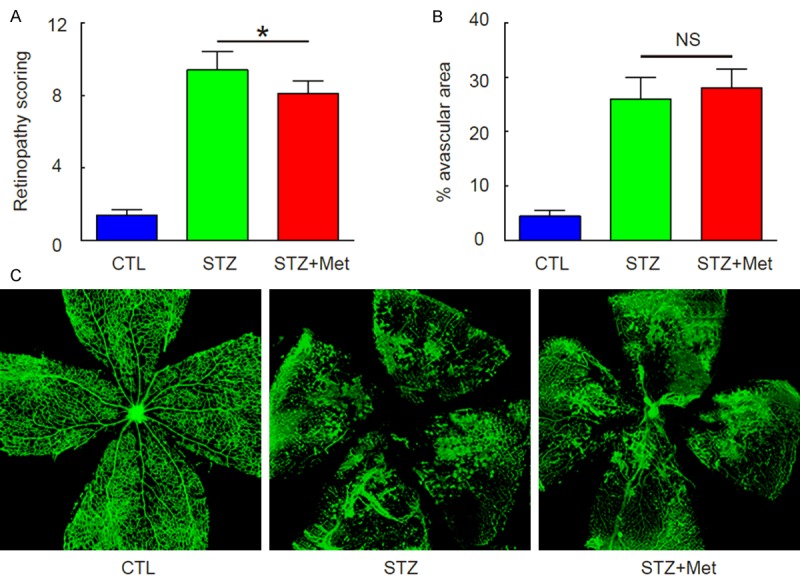
Induction of DR in mice by STZ administration. (A) Retinopathy scoring at 10 weeks after STZ. (B, C) Avascular area in eyes at 10 weeks after STZ, by quantification (B), and by representative images (C). *p<0.05. NS: non-significant. N=10.
Metformin attenuates neovascularization in the development of DR without affecting total VEGF-A protein
Next, we examined the mice at 12 weeks after STZ, when the neovascularization occurs. We found that metformin significantly attenuated vessel density at this time point, shown by representative images (Figure 3A), and by quantification (Figure 3B). Since VEGF-A is the well-known trigger of neovascularization in the development of DR, we analyzed VEGF-A protein levels by ELISA in mouse eyes. Surprisingly, although we confirmed significant increases in VEGF-A protein in eyes by STZ, we did not detect an effect of metformin on total VEGF-A levels (Figure 3C). Hence, Metformin attenuates neovascularization in the development of DR without affecting total VEGF-A protein.
Figure 3.
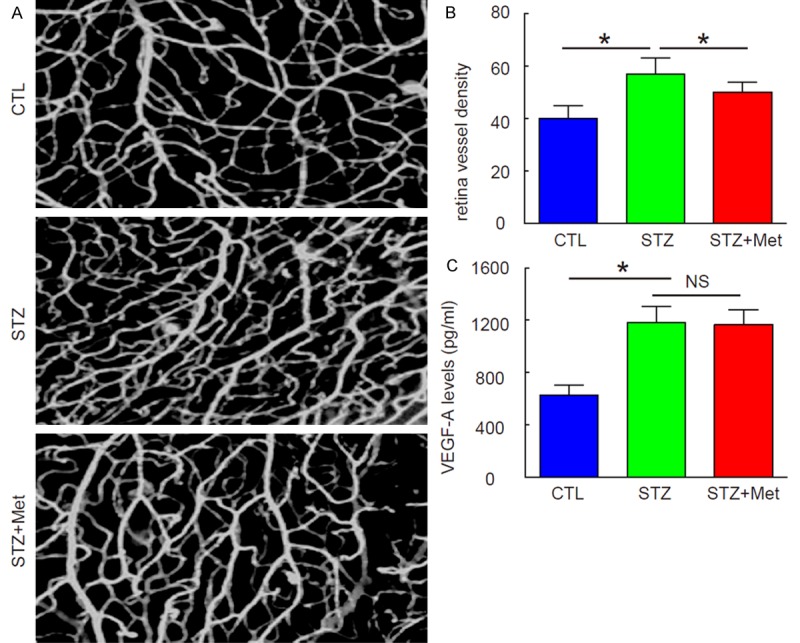
Metformin attenuates neovascularization in the development of DR without affecting total VEGF-A protein. (A, B) Attenuated vessel density was analyzed at 12 weeks after STZ, shown by representative images (A), and by quantification (B). (C) ELISA for total VEGF-A protein in mouse eyes. *p<0.05. NS: non-significant. N=10.
Metformin attenuates activation of VEGF signaling in the development of DR
Next, we examined whether Metformin may affect phosphorylation of VEGFR2, which represents the activation of VEGF signaling. We found that neither STZ nor metformin altered the total levels of VEGFR2 in mouse eyes, shown by representative Western blot images (Figure 4A), and by quantification (Figure 4B). However, STZ significantly increased the levels of VEGFR2 phosphorylation, which was attenuated by metformin, shown by representative Western blot images (Figure 4A), and by quantification (Figure 4C). Thus, Metformin attenuates activation of VEGF signaling in the development of DR.
Figure 4.
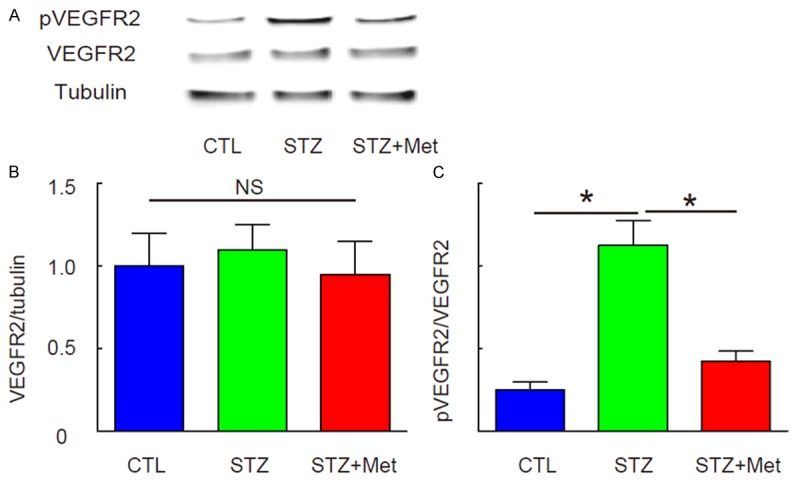
Metformin attenuates activation of VEGF signaling in the development of DR. The phosphorylation of VEGFR2 (pVEGFR2), which represents the activation of VEGF signaling, was analyzed by Western blot. A: Representative Western blot images. B: Quantification for total VEGFR2. C: Quantification for pVEGF2. *p<0.05. NS: non-significant. N=10.
Metformin alters VEGFR164 vs VEGF120 splicing to reduce VEGFR2 activation
VEGF signaling was altered by metformin while the total protein of VEGF-A, and the ligand of the VEGF signaling was unaltered. Therefore, we examined the isoforms of VEGF-A, since they are differently potent in activation of the receptor signaling. Isoform-specific RT-qPCR primers were thus designed (Figure 5A). We did not find VEGF188 isoform in mouse eyes. However, STZ significantly increased the levels of VEGF164, which was attenuated by metformin (Figure 5B). Thus, Metformin attenuates activation of VEGF signaling through alteration of VEGF-A mRNA splicing to increase the ratio of VEGFR120 vs VEGF164.
Figure 5.
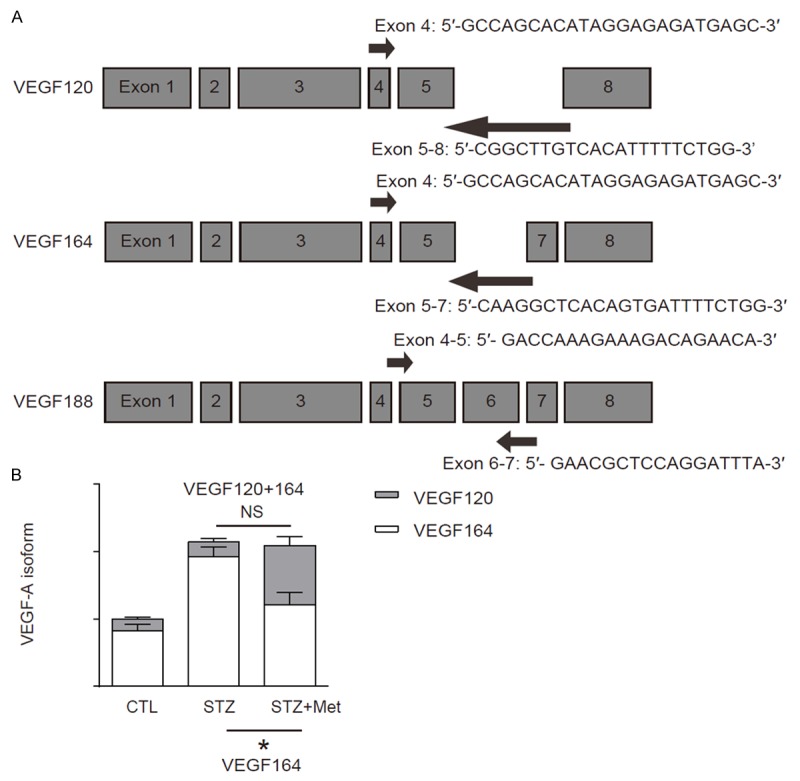
Metformin alters VEGFR164 vs VEGF120 splicing to reduce VEGFR2 activation. A: Isoform-specific RT-qPCR primers for VEGF-A. B: RT-qPCR for VEGF120 and VEGF164. *p<0.05. NS: non-significant. N=10.
Discussion
Metformin improves blood glucose control by different mechanisms than those of sulphonylureas or insulin, and its cardio-protective effects are independent of glycemic control. Metformin seems to lower fasting plasma insulin concentrations and enhance insulin sensitivity, to increase glucose uptake by peripheral tissues and decreasing hepatic glucose output [14-16]. Given the evidence of the anti-inflammatory and antiangiogenic activity of metformin, treatment with metformin may activate protective mechanisms against DR, because both inflammation-mediated pathways and angiogenesis are thought to underlie the progression of DR. However, the exact mechanisms have yet to be fully determined.
Here, we used a mouse model of diabetes to study the effects of metformin on DR. First, we confirmed sustained diabetes and development of DR in this model. The dose of STZ in the current study was well adjusted to induce a sustained but relatively modest hypoglycemia, which allows the mice to survive the relative long period for developing DR. we indeed found a modest protective effects of metformin against DR, seemingly at the second stage of DR when neovascularization occurs after ischemia period. These data are consistent with literature showing that metformin may activate protective mechanisms against angiogenesis during the progression of DR.
VEGF signaling is a key player in regulation of DR-associated neovascularization. The activation of this signaling pathway initiates from binding of the ligand (VEGF-A) to the receptor (VEGFR2) [17]. We thus examined the VEGF-A and VEGFR2 phosphorylation by STZ and metformin. Although VEGF-A levels were not affected by metformin, metformin indeed increased phosphorylation of VEGFR2. This paradoxical data inspired us to investigate the mechanisms underlying the altered activation of VEGFR2 without changes in total VEGF-A.
By differential mRNA splicing, the murine VEGF-A gene can give rise to three protein isoforms, VEGF120, VEGF164 and VEGF188. VEGF188 binds to heparin to be associated with the cell surface and the extracellular matrix, while VEGF120 is freely diffusible. The major biological isoform is VEGF164 that has intermediate properties between VEGF120 and VEGF188 [4-6]. We found that although metformin did not alter total VEGF-A, but it altered the ratio of VEGF120 and VEGF164 in the total VEGF-A in mouse eyes after STZ treatment. The VEGF120 levels were significantly increased and the potential of total VEGF-A was thus decreased, which explained the attenuated activation of VEGFR2, or VEGF signaling.
To summarize, our findings are critical for generating novel medicine through modulating VEGF mRNA splicing for DR treatment.
Acknowledgements
This study is supported by the Natural Science Foundation of Ningbo City (2016A610014).
Disclosure of conflict of interest
None.
References
- 1.Alusik S, Paluch Z. Metformin: the past, presence, and future. Minerva Med. 2015;106:233–238. [PubMed] [Google Scholar]
- 2.Effect of intensive blood-glucose control with metformin on complications in overweight patients with type 2 diabetes (UKPDS 34). UK Prospective Diabetes Study (UKPDS) Group. Lancet. 1998;352:854–865. [PubMed] [Google Scholar]
- 3.Dallaglio K, Bruno A, Cantelmo AR, Esposito AI, Ruggiero L, Orecchioni S, Calleri A, Bertolini F, Pfeffer U, Noonan DM, Albini A. Paradoxic effects of metformin on endothelial cells and angiogenesis. Carcinogenesis. 2014;35:1055–1066. doi: 10.1093/carcin/bgu001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Ferrara N, Gerber HP, LeCouter J. The biology of VEGF and its receptors. Nat Med. 2003;9:669–676. doi: 10.1038/nm0603-669. [DOI] [PubMed] [Google Scholar]
- 5.Stalmans I, Ng YS, Rohan R, Fruttiger M, Bouche A, Yuce A, Fujisawa H, Hermans B, Shani M, Jansen S, Hicklin D, Anderson DJ, Gardiner T, Hammes HP, Moons L, Dewerchin M, Collen D, Carmeliet P, D’Amore PA. Arteriolar and venular patterning in retinas of mice selectively expressing VEGF isoforms. J Clin Invest. 2002;109:327–336. doi: 10.1172/JCI14362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Dor Y, Djonov V, Keshet E. Making vascular networks in the adult: branching morphogenesis without a roadmap. Trends Cell Biol. 2003;13:131–136. doi: 10.1016/s0962-8924(03)00022-9. [DOI] [PubMed] [Google Scholar]
- 7.Xiao X, Prasadan K, Guo P, El-Gohary Y, Fischbach S, Wiersch J, Gaffar I, Shiota C, Gittes GK. Pancreatic duct cells as a source of VEGF in mice. Diabetologia. 2014;57:991–1000. doi: 10.1007/s00125-014-3179-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Xiao X, Guo P, Chen Z, El-Gohary Y, Wiersch J, Gaffar I, Prasadan K, Shiota C, Gittes GK. Hypoglycemia reduces vascular endothelial growth factor a production by pancreatic Beta cells as a regulator of Beta cell mass. J Biol Chem. 2013;288:8636–8646. doi: 10.1074/jbc.M112.422949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Bhatt LK, Addepalli V. Attenuation of diabetic retinopathy by enhanced inhibition of MMP-2 and MMP-9 using aspirin and minocycline in streptozotocin-diabetic rats. Am J Transl Res. 2010;2:181–189. [PMC free article] [PubMed] [Google Scholar]
- 10.Gong CY, Lu B, Hu QW, Ji LL. Streptozotocin induced diabetic retinopathy in rat and the expression of vascular endothelial growth factor and its receptor. Int J Ophthalmol. 2013;6:573–577. doi: 10.3980/j.issn.2222-3959.2013.05.03. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Si YF, Wang J, Guan J, Zhou L, Sheng Y, Zhao J. Treatment with hydrogen sulfide alleviates streptozotocin-induced diabetic retinopathy in rats. Br J Pharmacol. 2013;169:619–631. doi: 10.1111/bph.12163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Papachristodoulou D, Heath H, Kang SS. The development of retinopathy in sucrose-fed and streptozotocin-diabetic rats. Diabetologia. 1976;12:367–374. doi: 10.1007/BF00420981. [DOI] [PubMed] [Google Scholar]
- 13.Higgins RD, Yu K, Sanders RJ, Nandgaonkar BN, Rotschild T, Rifkin DB. Diltiazem reduces retinal neovascularization in a mouse model of oxygen induced retinopathy. Curr Eye Res. 1999;18:20–27. doi: 10.1076/ceyr.18.1.20.5390. [DOI] [PubMed] [Google Scholar]
- 14.Yang Z, Chen X, Chen Y, Zhao Q. PGC-1 mediates the regulation of metformin in muscle irisin expression and function. Am J Transl Res. 2015;7:1850–1859. [PMC free article] [PubMed] [Google Scholar]
- 15.Chen WP, Wang YD, Ma Y, Zhang ZY, Hu LY, Lin JL, Lin BQ. Danhong Huayu Koufuye combined with metformin attenuated diabetic retinopathy in Zucker diabetic fatty rats. Int J Ophthalmol. 2015;8:1094–1100. doi: 10.3980/j.issn.2222-3959.2015.06.03. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Joe SG, Yoon YH, Choi JA, Koh JY. Anti-angiogenic effect of metformin in mouse oxygen-induced retinopathy is mediated by reducing levels of the vascular endothelial growth factor receptor Flk-1. PLoS One. 2015;10:e0119708. doi: 10.1371/journal.pone.0119708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Cao Y. Angiogenesis and vascular functions in modulation of obesity, adipose metabolism, and insulin sensitivity. Cell Metab. 2013;18:478–489. doi: 10.1016/j.cmet.2013.08.008. [DOI] [PubMed] [Google Scholar]