

Drug-resistant Staphylococcus species are a major concern in human and veterinary medicine. There is a need for new antibiotics that exhibit a selective effect in treating infections in companion and livestock animals and that would not be used to treat human bacterial infections. We have identified fosmidomycin as an antibiotic that selectively targets certain Staphylococcus species that are often encountered in skin infections in cats and dogs. These findings expand our understanding of Staphylococcus evolution and may have direct implications for treating staphylococcal infections in veterinary medicine.
KEYWORDS: Staphylococcus, companion animals, comparative genomics, fosmidomycin, isoprenoid biosynthesis, One Health
ABSTRACT
Staphylococcus species are a leading cause of skin and soft tissue infections in humans and animals, and the antibiotics used to treat these infections are often the same. Methicillin- and multidrug-resistant staphylococcal infections are becoming more common in human and veterinary medicine. From a “One Health” perspective, this overlap in antibiotic use and resistance raises concerns over the potential spread of antibiotic resistance genes. Whole-genome sequencing and comparative genomics analysis revealed that Staphylococcus species use divergent pathways to synthesize isoprenoids. Species frequently associated with skin and soft tissue infections in companion animals, including S. schleiferi and S. pseudintermedius, use the nonmevalonate pathway. In contrast, S. aureus, S. epidermidis, and S. lugdunensis use the mevalonate pathway. The antibiotic fosmidomycin, an inhibitor of the nonmevalonate pathway, was effective in killing canine clinical staphylococcal isolates but had no effect on the growth or survival of S. aureus and S. epidermidis. These data identify an essential metabolic pathway in Staphylococcus that differs among members of this genus and suggest that drugs such as fosmidomycin, which targets enzymes in the nonmevalonate pathway, may be an effective treatment for certain staphylococcal infections.
IMPORTANCE Drug-resistant Staphylococcus species are a major concern in human and veterinary medicine. There is a need for new antibiotics that exhibit a selective effect in treating infections in companion and livestock animals and that would not be used to treat human bacterial infections. We have identified fosmidomycin as an antibiotic that selectively targets certain Staphylococcus species that are often encountered in skin infections in cats and dogs. These findings expand our understanding of Staphylococcus evolution and may have direct implications for treating staphylococcal infections in veterinary medicine.
INTRODUCTION
Staphylococcus infections pose a serious health threat to humans, companion animals, and livestock (1–3). Staphylococcus aureus is a Gram-positive bacterium that can asymptomatically colonize human skin and the anterior nares, but it is also responsible for mild to severe skin and soft tissue infections and life-threatening endocarditis, pneumonia, and sepsis. The Centers for Disease Control and Prevention estimated that in 2014 there were 72,000 invasive methicillin-resistant S. aureus (MRSA) infections in the United States, which resulted in 9,000 deaths (4). Although the prevalence of MRSA carriage in companion animals is low (approximately 0 to 4%) (5, 6) and infections are rare (7), other Staphylococcus species are common commensals and pathogens in veterinary medicine (8). S. schleiferi and S. pseudintermedius are the leading causes of canine and feline skin and ear infections (9); S. hyicus causes high-morbidity skin infections in pigs (10) and osteomyelitis in birds (11), while S. aureus, S. agnetis, and S. chromogenes cause mastitis in cattle and are associated with reduced milk quality (5, 12, 13).
The emergence of drug-resistant Staphylococcus is a global problem (14–21). Drugs such as erythromycin and cephalexin are commonly used to treat infections in both humans and animals, leading to concern that as resistance to shared antibiotics becomes more widespread, zoonotic transmission of either drug-resistant Staphylococcus or horizontal transfer of resistance genes (22) may render these treatments ineffective for both humans and animals. Despite the major animal health burden posed by a range of Staphylococcus species, much of our knowledge of Staphylococcus biology stems from studies on only a handful species that are important causes of human disease. To address this knowledge gap, we have used whole-genome sequencing of S. schleiferi and comparative genomics to identify novel drug targets to treat staphylococcal infections in companion animals.
RESULTS
High-quality genome sequences for S. schleiferi obtained via long-read sequencing technology.
We previously reported the use of single-molecule, real-time sequencing to generate complete genome sequences of four canine clinical isolates of S. schleiferi (23). These isolates represented a spectrum of antibiotic resistance profiles, from susceptible to all tested antibiotics (isolate 2142-05), intermediate resistance (2317-03 and 5909-02), to multidrug resistant (1360-13) (see Table S1 in the supplemental material). The long reads produced by this approach improved the ability to assemble highly repetitive regions of the S. schleiferi genome. For example, rRNA operons represent some of the longest repetitive regions in most bacterial genomes (24), and therefore they pose the biggest challenge for assembly. In addition, clustered, regularly interspaced short palindromic repeat (CRISPR) loci, a type of bacterial adaptive immunity, also pose a challenge to assembly, as they are comprised of a variable number of short, repetitive sequence elements. Our sequencing generated numerous long reads that completely spanned both of these difficult-to-assemble regions of the bacterial genome (Fig. S1a and b). Taken together, these data show that high-quality finished genomes have been generated for this important veterinary pathogen.
Long sequence reads assisted in assembling long repetitive regions. (a) Long reads generated by single-molecule real-time sequencing span a 5-kb rRNA operon and nearby coding regions. Red, yellow, and blue bars represent rRNA genes, tRNA genes, and protein-coding sequences, respectively. Black and gray lines indicate sequences that map to strain 5909-02 in the forward and reverse direction, respectively (total coverage in this region is ~90×). (b) Alignment of reads in the clustered regularly interspaced short palindromic repeat (CRISPR) region. Blue bars and pink triangles represent protein-coding cas genes and 36-bp CRISPR repeat sequences, respectively. Black and gray lines indicate sequences that map across the entire CRISPR repeat-spacer region in strain 1360-13 in the forward and reverse direction, respectively (total coverage in this region is ~80×). The genomes are ordered from smallest number of repeat sequences (4; strain 2142-05) to largest (36; strain 1360-13). Download Figure S1, PDF file, 0.8 MB (886.9KB, pdf) .
Copyright © 2016 Misic et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Antimicrobial susceptibility and metadata for S. schleiferi strains (the S. schleiferi strain metadata are shown, including date of collection, body site, and antibiotic susceptibility profiles; the antibiotic susceptibilities were determined using the Microscan Walkaway 40 PC20 Gram-positive combo-panel [Dade Behring, Sacramento, CA, USA]). Download Table S1, DOCX file, 0.02 MB (17.3KB, docx) .
Copyright © 2016 Misic et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Comparative genomic analysis of staphylococcal species.
To better understand the population genetics of Staphylococcus and identify putative drug targets, we compared our S. schleiferi genomes with publicly available complete genome sequences for four other Staphylococcus species. These included S. pseudintermedius, a leading cause of companion animal infections, and S. epidermidis, S. lugdunensis, and S. aureus USA300, important causes of community-acquired infections in humans. Genomes were aligned to strain USA300 via BLAST v2.2.22 (25) and visualized with Brig v0.95 (26) (Fig. 1a). A visual inspection of the circular alignments revealed genomic regions that were present in the human-associated S. aureus strain USA300 but absent from canine-associated species (Fig. 1a, arrows). Similarly, other regions were absent from all species examined except for S. aureus USA300 (Fig. 1a, arrowheads). To more robustly identify genetic pathways differentially abundant among Staphylococcus species, we expanded our analysis to include two to four strains for each of the five species shown in Fig. 1a. In total, genomes from 14 strains were annotated by using Rapid Annotation through Subsystems Technology (RAST) (27), allowing gene membership for 382 subsystems to be compared (see Table S2 in the supplemental material). A total of 147 subsystems were conserved, with an equal number of genes present in each subsystem across all 14 strains examined. In contrast, genes in 235 subsystems were differentially abundant among two or more species.
FIG 1 .

Comparative genomic analysis of Staphylococcus genomes. (a) Circular alignment of whole-genome sequences for five Staphylococcus species. Working from the inner-most ring out: S. aureus subsp. aureus USA300 (solid black ring); GC content of S. aureus USA300; GC skew of S. aureus USA300; S. lugdunensis (purple); S. epidermidis (blue); S. pseudintermedius ED99 (green); S. schleiferi strain 1360-13 (red). Arrows indicate genomic regions missing from canine-associated Staphylococcus. Triangles point to regions missing from all strains examined but which were present in S. aureus USA300. (b) A hierarchical clustering heat map of differentially abundant subsystems among 14 selected Staphylococcus genomes from five species. (c) Five selected subsystems found in higher abundance in S. aureus, S. lugdunensis, and S. epidermidis than in S. pseudintermedius or S. schleiferi. (d) Five selected subsystems found in higher abundance in S. aureus than in all the other species examined. (e) Two selected subsystems found in higher abundance in canine-associated Staphylococcus species (S. schleiferi and S. pseudintermedius) than in human-associated species (S. aureus, S. epidermidis, and S. lugdunensis). In panels c, d, and e, only subsystems for which at least two genes were different between the compared groups are shown. The red text in panels c and e highlights isoprenoid biosynthesis pathways.
RAST-annotated subsystems of 14 Staphylococcus genomes (from a total of 382 genomes, 147 subsystems were conserved, with an equal number of genes present in each subsystem across all 14 strains examined; genes in 235 subsystems were differentially abundant among two or more species). Download Table S2, XLSX file, 0.03 MB (32.9KB, xlsx) .
Copyright © 2016 Misic et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
To identify relationships within the subsystems data, hierarchical clustering was performed to group bacterial strains based on similar subsystems profiles (Fig. 1b, top dendrogram), as well as to group subsystems (Fig. 1b, rows) that were enriched in canonically human-associated species (S. aureus, S. epidermidis, and S. lugdunensis) or canine-associated species (S. schleiferi and S. pseudintermedius). As expected, strains from the same species clustered most closely together (Fig. 1b, top dendrogram). The human- and canine-associated Staphylococcus species segregated, indicating that they differ in their functional gene content.
Differences in virulence factors and metabolic pathways dominate subsystems analysis.
At least three distinct subsystem clusters emerged from our analysis. Cluster 1 consisted of 15 subsystems enriched in all four S. aureus strains but absent from all other species. This subsystem cluster included adhesins and pore-forming cytolysins (Fig. 1d). Given the importance of virulence factors in Staphylococcus biology, we examined these subsystems in more detail. S. aureus isolates contained more than 20 adhesins, while S. epidermidis and S. lugdunensis isolates had between 6 and 8 adhesins and the S. schleiferi and S. pseudintermedius isolates contained 9 to 10 adhesins. Table S3 in the supplemental material includes a list of 23 adhesins and their presence or absence among 14 genomes. One adhesin, staphylocoagulase, activates prothrombin to coagulate blood. Although S. pseudintermedius and S. schleiferi subsp. coagulans (S. schleiferi strain 1360-13) can coagulate blood, our subsystems analysis showed that there were no staphylocoagulase genes (coa) present in the genomes (see Table S3). Consistent with previous studies of S. aureus, we found that S. aureus contains more virulence factors than other Staphylococcus species (28–30). Table S3 shows select adhesins and antimicrobial resistance factors found among the staphylococcal genomes. A surprising finding was that S. schleiferi 1360-13 is methicillin resistant (see Table S1 in the supplemental material), yet a mecA or mecC gene was not found in the genome nor on a plasmid (see Table S3). A recent report documented a case of an S. schleiferi human clinical isolate that was resistant to methicillin yet was penicillin binding protein 2 negative (31). The basis of methicillin resistance in these isolates is not yet understood.
Select adhesin and antimicrobial resistance genes found among 14 Staphylococcus genomes (the presence or absence of selected virulence genes in the respective genomes is indicated with a + or − symbol). Download Table S3, XLSX file, 0.02 MB (26.4KB, xlsx) .
Copyright © 2016 Misic et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Cluster 2 comprised 15 subsystems that were absent from canine-associated Staphylococcus isolates but which were present across all other species examined (Fig. 1c). Among these subsystems was the mevalonate pathway for isoprenoid biosynthesis. This finding was surprising, given that isoprenoids are an essential class of natural products and that staphylococci have been shown to use the mevalonate pathway for isoprenoid biosynthesis (32, 33). Further examination of the canine-associated Staphylococcus species (cluster 3, 13 subsystems) showed that these species use the nonmevalonate pathway, an alternative route to isoprenoid biosynthesis (Fig. 1e). Taken together, these data suggest that staphylococcal species use different routes to generate isoprenoids.
Fosmidomycin selectively kills bacteria associated with veterinary skin and ear infections.
Isoprenoid biosynthesis is a highly conserved and essential process in bacteria, eukaryotes, and plants (33, 34), and inhibitors have been used to target both the mevalonate and the nonmevalonate pathways. We hypothesized that if S. schleiferi and S. pseudintermedius used the nonmevalonate pathway to synthesize isoprenoids, they should be sensitive to the drug fosmidomycin, a phosphonic acid derivative that blocks the first committed step of the nonmevalonate pathway via inhibition of 1-deoxy-d-xylulose 5-phosphate reductoisomerase (Dxr) (35). To test this hypothesis, S. schleiferi and S. pseudintermedius (canine associated) and S. aureus and S. epidermidis (human associated) were grown on Mueller-Hinton agar plates supplemented with 50 µg/ml fosmidomycin. As expected, S. aureus and S. epidermidis, both of which are reported to use the mevalonate pathway (33), grew in the presence of fosmidomycin. In contrast, fosmidomycin completely restricted growth of S. schleiferi and S. pseudintermedius (Fig. 2a). To expand on this analysis, we used MIC and minimum bactericidal concentration (MBC) assays to quantify fosmidomycin activity against a panel of Staphylococcus strains from the five species evaluated in Fig. 1. While S. aureus, S. lugdunensis, and S. epidermidis grew normally even in the presence of 256 µg/ml of the drug, S. schleiferi and S. pseudintermedius were inhibited by fosmidomycin concentrations as low as 0.5 µg/ml and were killed by concentrations of 4 to 16 µg/ml (Fig. 2b). These results provide a biochemical validation of our comparative genomics data (Fig. 1) and suggest that this antibiotic is active against specific Staphylococcus species.
FIG 2 .
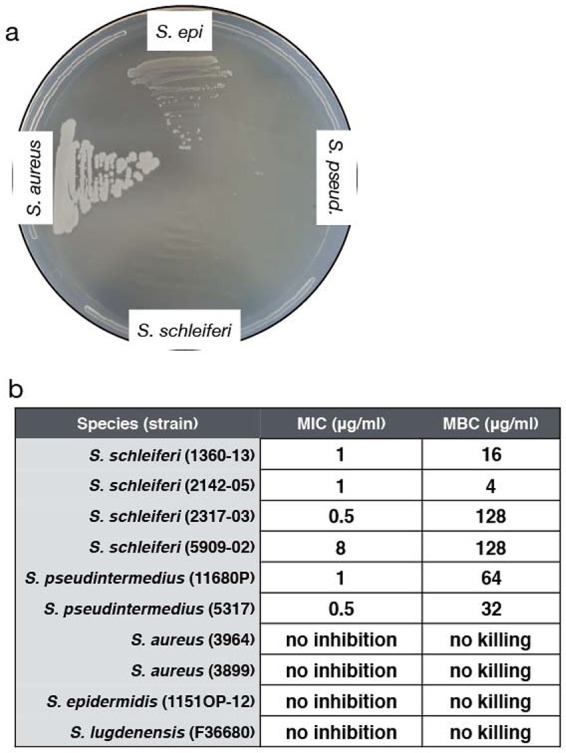
Fosmidomycin selectively kills Staphylococcus species associated with companion animal infections. (a) A Mueller-Hinton agar plate containing 50 µg/ml fosmidomycin was streaked with overnight cultures of strains S. aureus 3964 (USA100), S. epidermidis (1151OP-12), S. pseudintermedius (11680P), and S. schleiferi (1360-13). (b) MIC and MBC values for 10 Staphylococcus strains.
Divergent isoprenoid biosynthesis in the genus Staphylococcus is associated with host species.
Although many Gram-negative bacteria have been described to use the nonmevalonate pathway (32, 36, 37), S. aureus and other Staphylococcus species have long been cited as examples of bacteria that use the mevalonate pathway (32, 33). This prompted us to expand our bioinformatics analysis beyond the five Staphylococcus species examined thus far to explore isoprenoid biosynthesis more broadly across the genus. The superoxide dismutase (sodA) gene was used to construct a maximum-likelihood phylogenetic tree, and manual curation with KEGG was used to determine which isoprenoid biosynthesis pathway was present in 29 staphylococcal species (Fig. 3). The sodA gene has previously been shown to be a good representative of Staphylococcus phylogeny (38). This analysis showed that the nonmevalonate pathway was frequently used by Staphylococcus species known to cause disease in companion animals and wildlife (Fig. 3, gray box). Use of the nonmevalonate pathway appears to be an ancestral trait, with use of the mevalonate pathway emerging later (Fig. 3, red dot). Staphylococcus species that possess the mevalonate pathway formed a monophyletic group that included many notable human- and primate-associated species, such as S. hominis, S. haemolyticus, S. simiae, and S. aureus (Fig. 3, yellow box). Also included in this clade were animal-associated species, including S. equorum, S. gallinarum, and S. xylosus, which are often associated with horses, chickens, and mice, respectively, as well as S. aureus strain ST398, which is an important cause of disease in livestock.
FIG 3 .
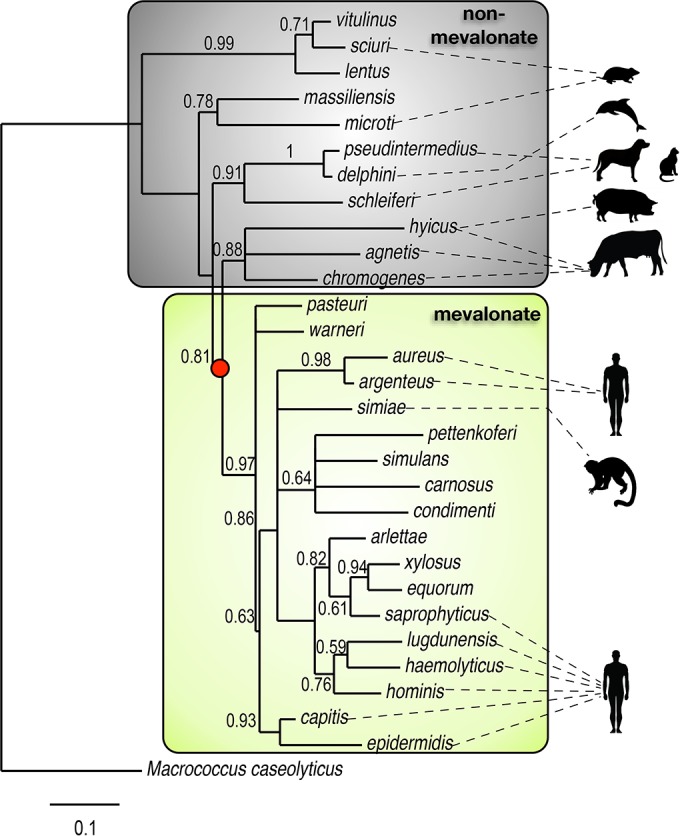
Evolution of isoprenoid biosynthesis in Staphylococcus. A maximum-likelihood phylogenetic tree constructed from the Staphylococcus superoxide dismutase gene (sodA) is shown. Gray and yellow boxes indicate Staphylococcus species that utilize the nonmevalonate or mevalonate pathway, respectively. Silhouettes show nonexclusive host associations. The red dot indicates the branch point in the tree where the mevalonate pathway emerged. The tree was rooted with Macrococcus caseolyticus as the outgroup.
Lateral gene transfer has been suggested to be common in the evolution of isoprenoid biosynthesis in bacteria (37), and this prompted us to examine the gene membership of both pathways for a range of Staphylococcus species. The mevalonate pathway begins with acetyl coenzyme A (CoA), which undergoes five sequential enzymatic reactions to generate isopentenyl pyrophosphate (IPP) and dimethylallyl pyrophosphate (DMAPP), which are then converted to isoprenoids (Fig. 4a, blue). In contrast, the nonmevalonate pathway involves a completely distinct set of seven sequential enzymatic steps to convert pyruvate or glucose-3-phosphate to IPP and DMAPP (Fig. 4a, red).
FIG 4 .

Isoprenoid pathway organization in Staphylococcus. (a) Schematic showing enzymes (italics) and intermediates (colored ovals) in the mevalonate (blue) and nonmevalonate (red) pathways. (b) A summary of pathway genes present (indicated by check marks) in Staphylococcus species, determined using the KEGG database. (c) Chromosomal location of the mevalonate (blue triangles) and nonmevalonate (red triangles) genes in staphylococcal genomes are shown. The genomes are presented as linear chromosomes oriented such that 0 Mbp is the origin of replication. S. aur, S. aureus; S. epi, S. epidermidis; S. lugd, S. lugdunensis; S. pseud, S. pseudintermedius; S. schl, S. schleiferi.
While S. aureus, S. epidermidis, and S. lugdunensis each possess every gene in the mevalonate pathway, they have also retained some nonmevalonate pathway enzymes, including 2-C-methyl-d-erythritol 4-phosphate cytidylyltransferase (IspD) and 4-diphosphocytidyl-2C-methyl-d-erythritol kinase (IspE) (Fig. 4b; see also Table S4 in the supplemental material). In contrast, canine-associated S. pseudintermedius and S. schleiferi possess all members of the nonmevalonate pathway and completely lack genes of the mevalonate pathway (Fig. 4b; see Table S4). The genomic location of functionally related genes can reflect their evolutionary history. Similar to the Staphylococcus mevalonate pathway (Fig. 4c, red triangles) (33), we found that the nonmevalonate pathway genes were dispersed across the chromosome (Fig. 4c, blue triangles). Taken together, these data argue against a recent lateral transfer of either pathway.
Isoprenoid biosynthesis genes in 14 Staphylococcus genomes (the mevalonate and nonmevalonate pathway gene names, abbreviations, enzyme catalog numbers, and locus identifiers of select Staphylococcus species are shown). Download Table S4, XLSX file, 0.05 MB (47.7KB, xlsx) .
Copyright © 2016 Misic et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
DISCUSSION
The majority of studies examining the metabolism and biology of Staphylococcus have focused on S. aureus. While S. aureus is an important pathogen in humans and animals, there are many other Staphylococcus species of great importance to veterinary medicine. Early biochemical experiments conducted primarily with S. aureus helped to establish a dogma that Staphylococcus uses the mevalonate pathway to synthesize essential isoprenoids. Our data challenge this dogma and show that isoprenoid biosynthesis differs between Staphylococcus species. Given that mammals use the same pathway, it has been assumed that targeting isoprenoid biosynthesis is not a viable strategy to treat staphylococcal bacterial infections in humans or other animals. These data point to inhibitors of the nonmevalonate pathway, such as fosmidomycin, as potential antimicrobials to treat certain Staphylococcus infections in animals, in particular in companion animals where species utilizing the nonmevalonate pathway are a major cause of skin and ear infections.
The main target of fosmidomycin is the Dxr protein, but there is evidence that it can also inhibit a downstream enzyme, IspD, in vitro and in vivo (39). As is the case with any antibiotic, resistance to fosmidomycin could develop in Staphylococcus, either by blocking entry or accumulation of the drug in the bacterium or via mutations in the Dxr binding site, both of which have been reported in other pathogens (40–42). Despite these potential problems, the current clinical literature suggests that fosmidomycin could be a promising drug to treat infections caused by Staphylococcus species that use the nonmevalonate pathway. Fosmidomycin is extremely well-tolerated and exhibits low toxicity in mammals (32). Plasmodium falciparum, the cause of malaria, also synthesizes isoprenoids via the nonmevalonate pathway, and fosmidomycin was shown to be effective in killing the parasite in culture and achieved cure rates of 85 to 100% in clinical trials when administered alone or in combination with clindamycin (43, 44).
Most Gram-negative bacteria synthesize isoprenoids via the nonmevalonate pathway. However, across different bacterial phyla, there are examples of species that use the mevalonate pathway, including Streptococcus, Lactobacillus, Myxococcus, and Borrelia. In addition, some Pseudomonas species, including Pseudomonas mevalonii, are known to use hydroxymethylglutaryl-CoA reductase (the third enzyme of the mevalonate pathway) for degradative functions (45, 46). Only Listeria monocytogenes and a few species of Streptomyces are known to possess both pathways, but in both organisms the nonmevalonate pathway plays the essential role in primary metabolism (37), while the mevalonate pathway is dispensable (47). Based on our data, Staphylococcus constitutes a unique example of a bacterial genus whose species utilize different isoprenoid biosynthesis pathways. Our phylogenetic analysis results (Fig. 3) are consistent with previous evolutionary studies that have suggested that the nonmevalonate pathway is the ancestral pathway in bacteria and the mevalonate pathway was acquired later through lateral gene transfer (34); this is further supported by our finding that deep-branching taxa (such as the S. sciuri and S. intermedius groups) use the nonmevalonate pathway, while the more recently branched taxa (including the S. aureus and S. epidermidis groups) use the mevalonate pathway.
An outstanding question is why Staphylococcus species evolved to use different pathways for isoprenoid synthesis and whether pathway usage influences host range selection. There is abundant literature on virulence factors influencing host range (48), but less is known about the role of bacterial metabolism. Interestingly, among the Staphylococcus species we analyzed, mevalonate pathway usage was associated with species that are found across human, nonhuman primate, and animal hosts. In contrast, nonmevalonate pathway usage was only associated with an animal host range. One possible explanation for this observation may lie in the secondary metabolites produced by these pathways and their interaction with the host immune system: 1-hydroxy-2-methyl-2-(E)-butenyl 4-diphosphate (HMB-PP) is an intermediate of the nonmevalonate pathway and a potent activator of Vγ2/Vδ2 (also called Vγ9/Vδ2) T cells (49). HMB-PP has 1,000 times greater immune stimulatory activity than IPP, the analogous intermediate produced by the mevalonate pathway (49, 50). Vγ2Vδ2 cells make up 1 to 5% of peripheral T cells but expand to >50% and rapidly traffic to barrier surfaces in response to pathogens that produce HMB-PP (51, 52), yet these cells are only found in humans and nonhuman primates. This could lead to a scenario in which mevalonate usage (i.e., by S. aureus) results in a relatively weak Vγ2Vδ2 signal, thereby allowing colonization of human and nonhuman primate skin. Moreover, spread to animals would not be impeded, because these hosts completely lack the Vγ2Vδ2-bearing cells. In contrast, nonmevalonate pathway usage (i.e., by S. schleiferi and S. pseudintermedius) would result in production of the potent Vγ2Vδ2 ligand HMB-PP, but this would only be of consequence if the bacteria were on human or nonhuman primate hosts. Thus, one plausible hypothesis is that nonmevalonate pathway usage by Staphylococcus may contribute to an animal host range restriction, but this remains to be tested. Interestingly, S. schleiferi and S. pseudintermedius were first identified in human infections and are occasionally reported to cause serious human disease, but such cases are primarily observed in infants, the elderly, immunocompromised patients, or as a consequence of medical complications (53–57), perhaps lowering the immunological barrier to transmission.
MATERIALS AND METHODS
Bacterial strains, media, and growth conditions.
Staphylococcus isolates used for biochemical analysis were collected at the Matthew J. Ryan Veterinary Hospital of the University of Pennsylvania. S. schleiferi biochemical identification was carried out on a Microscan Walkaway 40 PC20 Gram-positive combo-panel (Dade Behring, Sacramento, CA). Four banked S. schleiferi strains were selected for whole-genome sequencing: 1360-13, 2142-05, 2317-03, and 5909-02. Table S1 in the supplemental material contains a full list of S. schleiferi strains used in this study and their associated metadata. For biochemical assays, the following clinical isolates were used: S. aureus 3964, S. aureus 3899, S. pseudintermedius 5317, S. pseudintermedius 1168OP, S. epidermidis 1151OP-12, and S. lugdunensis F36680.
DNA purification and sequencing.
S. schleiferi genomic DNA was purified, sequenced, and assembled into complete genomes as previously described (23). Briefly, DNA was extracted from overnight cultures of S. schleiferi isolates by using the Qiagen Genomic Tips kit (Valencia, CA, USA). DNA quantity and quality were assessed using a NanoDrop 1000 spectrophotometer (Thermo Scientific, Pittsburgh, PA) and a Qubit fluorometer (Life Technologies, Grand Island, NY, USA). Agarose gel electrophoresis was used to confirm high-molecular-weight DNA (>50 kb) for single-molecule real-time sequencing on a Pacific Biosciences RSII platform. SMRTbell adapters were ligated, and each strain of S. schleiferi was sequenced on 1 cell with one 120-min movie. A hierarchical genome assembly process (HGAP) was performed for each strain using the HGAP.3 module (58). The genome was closed using manual refinement.
Comparative genomics.
For comparative genome analyses, 14 sequences from GenBank were retrieved for the following organisms: S. schleiferi 1360-13 (accession number CP009470), S. schleiferi 2142-05 (CP009762), S. schleiferi 5909-02 (CP009676), S. schleiferi 2317-03 (GenBank: CP010309), S. pseudintermedius HKU10-03 (NC_014925.1), S. pseudintermedius ED99 (NC_017568), S. lugdunensis HKU09-01 (CP001837), S. lugdunensis N920143 (FR870271.1), S. epidermidis ATCC 12228 (NC_004461), S. epidermidis RP62A (NC_002976.3), S. aureus subspecies aureus ST398 (NC_017333), S. aureus subspecies aureus USA300_FPR3757 (NC_007793), S. aureus subspecies aureus COL (NC_002951), and S. aureus subspecies aureus NCTC 8325 (NC_007795) were downloaded from NCBI (ftp://ftp.ncbi.nlm.nih.gov). Five circular, whole genomes were aligned using the BLAST Ring Image Generator (BRIG) version 0.95 (26) and the Basic Local Alignment Search Tool (BLAST, v. 2/2/22) (25). Functional gene categories were determined with the Rapid Annotation using Subsystem Technology (RAST) v. 2.0 (27) and FigFam v. 70 programs. Subsystems with a standard deviation of zero among species were removed. The remaining differentially abundant subsystems were clustered by a Pearson correlation, and the bacterial species were clustered by a Spearman correlation using the hclust function in R (v. 3.2.0) (59) to reveal species-specific subsystem clusters. The cutree function in R was used to identify groups with similar subsystem abundance profiles.
Antibiotic susceptibility assays.
The MICs of various antimicrobials (amoxicillin-clavulanic acid, ampicillin, cefazolin, chloramphenicol, ciprofloxacin, clindamycin, erythromycin, gentamicin, imipenem, oxacillin, penicillin, rifampin, tetracycline, trimethoprim-sulfamethoxazole, and vancomycin) were tested via broth microdilution on a Microscan Walkaway 40 PC20 Gram-positive combo-panel (Dade Behring, Sacramento, CA). Fosmidomycin MIC assays were performed following clinical laboratory standards (60). Fosmidomycin (Sigma, St. Louis, MO, USA) was filter sterilized with a 0.2-mm filter (Pall Corporation, Ann Arbor, MI, USA) and was serially 2-fold diluted for a concentration range of 40.96 to 0.25 µg/ml. Tubes containing 1 ml Mueller-Hinton broth (Sigma) were inoculated with 5 × 105 CFU/ml. A 50-μl aliquot of the appropriate fosmidomycin concentration was added to each tube containing the starter culture and was incubated at 37°C overnight with shaking at 250 rpm for 18 to 20 h. The MICs were determined by visual inspection. The MBC was determined by plating 200 µl of the drug-treated cultures onto Mueller-Hinton agar plates (Remel, Lenexa, MA, USA). The lowest concentration at which there was no growth after a 24-h incubation at 37°C was determined to be the MBC. For the plate-based fosmidomycin growth assay, overnight cultures of Staphylococcus species grown in Mueller-Hinton broth were subcultured to Mueller-Hinton agar plates containing 50 µg/ml fosmidomycin and were incubated for 24 h at 37°C. All assays were performed and the results were interpreted using Clinical and Laboratory Standards Institute guidelines (60).
Phylogenetic tree construction and metabolic pathway comparisons.
Gene sequences of sodA from 29 staphylococcal species were downloaded from NCBI. The Web-based tool Phylogeny.fr (61) was used to construct a phylogenetic tree from a sodA gene multiple-sequence alignment, and Macrococcus caseolyticus was set as the outgroup. Sequences were aligned with Muscle v3.7 (62), poorly aligned regions were removed using Gblocks v0.91b (63), and the phylogenetic tree was reconstructed using the maximum likelihood method implemented in PhyML v3.0 (64). Tree rendering was performed using TreeDyn v198.3 (65), and bootstrap values are indicated on the branches. Metabolic pathway reconstructions of each strain were compared in silico using the terpenoid backbone biosynthesis pathway from KEGG (66, 67).
ACKNOWLEDGMENTS
We thank Meghan Davis, Stephen Cole, Kathleen O’Shea, and Kristen Constantine for assistance with bacterial growth and helpful discussions.
A.M.M. purified genomic DNA, assembled the genome sequences, analyzed sequence and subsystems data, performed comparative genomics, performed all bacterial growth and antibiotic assays, and wrote the manuscript. C.L.C. collected S. schleiferi clinical samples. S.C.R. isolated the Staphylococcus clinical isolates, performed all phenotypic testing of the strains, and archived them in a collection held at −80°C. D.P.B. analyzed subsystems data and wrote the manuscript. A.M.M., C.L.C., D.O.M., S.C.L., and D.P.B. conceived of the study, edited the manuscript, gave approval for publication, and agreed to be accountable for the work.
Funding Statement
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.
REFERENCES
- 1.Kawakami T, Shibata S, Murayama N, Nagata M, Nishifuji K, Iwasaki T, Fukata T. 2010. Antimicrobial susceptibility and methicillin resistance in Staphylococcus pseudintermedius and Staphylococcus schleiferi subsp. coagulans isolated from dogs with pyoderma in Japan. J Vet Med Sci 72:1615–1619. doi: 10.1292/jvms.10-0172. [DOI] [PubMed] [Google Scholar]
- 2.Cain CL, Morris DO, Rankin SC. 2011. Clinical characterization of Staphylococcus schleiferi infections and identification of risk factors for acquisition of oxacillin-resistant strains in dogs: 225 cases (2003–2009). J Am Vet Med Assoc 239:1566–1573. doi: 10.2460/javma.239.12.1566. [DOI] [PubMed] [Google Scholar]
- 3.Cain CL. 2013. Antimicrobial resistance in staphylococci in small animals. Vet Clin North Am Small Anim Pract 43:19–40. doi: 10.1016/j.cvsm.2012.09.003. [DOI] [PubMed] [Google Scholar]
- 4.Centers for Disease Control and Prevention 2014. ABCs Report: methicillin-resistant Staphylococcus aureus, 2014 Centers for Disease Control and Prevention, Atlanta, GA. [Google Scholar]
- 5.Weese JS. 2010. Methicillin-resistant Staphylococcus aureus in animals. ILAR J 51:233–244. doi: 10.1093/ilar.51.3.233. [DOI] [PubMed] [Google Scholar]
- 6.Loeffler A, Pfeiffer DU, Lindsay JA, Soares Magalhães RJ, Lloyd DH. 2011. Prevalence of and risk factors for MRSA carriage in companion animals: a survey of dogs, cats and horses. Epidemiol Infect 139:1019–1028. doi: 10.1017/S095026881000227X. [DOI] [PubMed] [Google Scholar]
- 7.Davis MF, Misic AM, Morris DO, Moss JT, Tolomeo P, Beiting DP, Nachamkin I, Lautenbach E, Rankin SC. 2015. Genome sequencing reveals strain dynamics of methicillin-resistant Staphylococcus aureus in the same household in the context of clinical disease in a person and a dog. Vet Microbiol 180:304–307. doi: 10.1016/j.vetmic.2015.09.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.May ER, Hnilica KA, Frank LA, Jones RD, Bemis DA. 2005. Isolation of Staphylococcus schleiferi from healthy dogs and dogs with otitis, pyoderma, or both. J Am Vet Med Assoc 227:928–931. doi: 10.2460/javma.2005.227.928. [DOI] [PubMed] [Google Scholar]
- 9.Davis MF, Cain CL, Brazil AM, Rankin SC. 2013. Two coagulase-negative staphylococci emerging as potential zoonotic pathogens: wolves in sheep’s clothing? Front Microbiol 4:123. doi: 10.3389/fmicb.2013.00123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Foster AP. 2012. Staphylococcal skin disease in livestock. Vet Dermatol 23:342–351. doi: 10.1111/j.1365-3164.2012.01093.x. [DOI] [PubMed] [Google Scholar]
- 11.Maier K, Fischer D, Hartmann A, Kershaw O, Prenger-Berninghoff E, Pendl H, Schmidt MJ, Lierz M. 2015. Vertebral osteomyelitis and septic arthritis associated with Staphylococcus hyicus in a juvenile peregrine falcon (Falco peregrinus). J Avian Med Surg 29:216–223. doi: 10.1647/2014-036. [DOI] [PubMed] [Google Scholar]
- 12.Pyörälä S, Taponen S. 2009. Coagulase-negative staphylococci-emerging mastitis pathogens. Vet Microbiol 134:3–8. doi: 10.1016/j.vetmic.2008.09.015. [DOI] [PubMed] [Google Scholar]
- 13.Taponen S, Supré K, Piessens V, Van Coillie E, De Vliegher S, Koort JM. 2012. Staphylococcus agnetis sp. nov., a coagulase-variable species from bovine subclinical and mild clinical mastitis. Int J Syst Evol Microbiol 62:61–65. doi: 10.1099/ijs.0.028365-0. [DOI] [PubMed] [Google Scholar]
- 14.Bramble M, Morris D, Tolomeo P, Lautenbach E. 2011. Potential role of pet animals in household transmission of methicillin-resistant Staphylococcus aureus: a narrative review. Vector Borne Zoonotic Dis 11:617–620. doi: 10.1089/vbz.2010.0025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Rutland BE, Weese JS, Bolin C, Au J, Malani AN. 2009. Human-to-dog transmission of methicillin-resistant Staphylococcus aureus. Emerg Infect Dis 15:1328–1330. doi: 10.3201/eid1508.081635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Verkade E, Kluytmans-van den Bergh M, van Benthem B, van Cleef B, van Rijen M, Bosch T, Schouls L, Kluytmans J. 2014. Transmission of methicillin-resistant Staphylococcus aureus CC398 from livestock veterinarians to their household members. PLoS One 9:e100823. doi: 10.1371/journal.pone.0100823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Loeffler A, Boag AK, Sung J, Lindsay JA, Guardabassi L, Dalsgaard A, Smith H, Stevens KB, Lloyd DH. 2005. Prevalence of methicillin-resistant Staphylococcus aureus among staff and pets in a small animal referral hospital in the UK. J Antimicrob Chemother 56:692–697. doi: 10.1093/jac/dki312. [DOI] [PubMed] [Google Scholar]
- 18.Morris DO, Rook KA, Shofer FS, Rankin SC. 2006. Screening of Staphylococcus aureus, Staphylococcus intermedius, and Staphylococcus schleiferi isolates obtained from small companion animals for antimicrobial resistance: a retrospective review of 749 isolates (2003–04). Vet Dermatol 17:332–337. doi: 10.1111/j.1365-3164.2006.00536.x. [DOI] [PubMed] [Google Scholar]
- 19.Van Duijkeren E, Wolfhagen MJ, Box AT, Heck ME, Wannet WJ, Fluit AC. 2004. Human-to-dog transmission of methicillin-resistant Staphylococcus aureus. Emerg Infect Dis 10:2235–2237. doi: 10.3201/eid1012.040387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Fitzgerald JR. 2012. Livestock-associated Staphylococcus aureus: origin, evolution and public health threat. Trends Microbiol 20:192–198. doi: 10.1016/j.tim.2012.01.006. [DOI] [PubMed] [Google Scholar]
- 21.Fluit AC. 2012. Livestock-associated Staphylococcus aureus. Clin Microbiol Infect 18:735–744. doi: 10.1111/j.1469-0691.2012.03846.x. [DOI] [PubMed] [Google Scholar]
- 22.Chambers HF, Deleo FR. 2009. Waves of resistance: Staphylococcus aureus in the antibiotic era. Nat Rev Microbiol 7:629–641. doi: 10.1038/nrmicro2200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Misic AM, Cain CL, Morris DO, Rankin SC, Beiting DP. 2015. Complete genome sequence and methylome of Staphylococcus schleiferi, an important cause of skin and ear infections in veterinary medicine. Genome Announc 3:e01011-15. doi: 10.1128/genomeA.01011-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Hill CW. 1999. Large genomic sequence repetitions in bacteria: lessons from rRNA operons and Rhs elements. Res Microbiol 150:665–674. doi: 10.1016/S0923-2508(99)00125-4. [DOI] [PubMed] [Google Scholar]
- 25.Camacho C, Coulouris G, Avagyan V, Ma N, Papadopoulos J, Bealer K, Madden TL. 2009. BLAST+: architecture and applications. BMC Bioinformatics 10:421. doi: 10.1186/1471-2105-10-421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Alikhan NF, Petty NK, Ben Zakour NL, Beatson SA. 2011. BLAST ring image generator (Brig): simple prokaryote genome comparisons. BMC Genomics 12:402. doi: 10.1186/1471-2164-12-402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Aziz RK, Bartels D, Best AA, DeJongh M, Disz T, Edwards RA, Formsma K, Gerdes S, Glass EM, Kubal M, Meyer F, Olsen GJ, Olson R, Osterman AL, Overbeek RA, McNeil LK, Paarmann D, Paczian T, Parrello B, Pusch GD, Reich C, Stevens R, Vassieva O, Vonstein V, Wilke A, Zagnitko O. 2008. The RAST server: rapid annotations using subsystems technology. BMC Genomics 9:75. doi: 10.1186/1471-2164-9-75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Powers ME, Bubeck Wardenburg J. 2014. Igniting the fire: Staphylococcus aureus virulence factors in the pathogenesis of sepsis. PLoS Pathog 10:e1003871. doi: 10.1371/journal.ppat.1003871. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Lowy FD. 1998. Staphylococcus aureus infections. N Engl J Med 339:520–532. doi: 10.1056/NEJM199808203390806. [DOI] [PubMed] [Google Scholar]
- 30.Schlievert PM. 1993. Role of superantigens in human disease. J Infect Dis 167:997–1002. doi: 10.1093/infdis/167.5.997. [DOI] [PubMed] [Google Scholar]
- 31.Merchant C, Villanueva DD, Lalani I, Eng M, Kang Y. 2016. PBP-2 negative methicillin resistant Staphylococcus schleiferi bacteremia from a prostate abscess: an unusual occurrence. Case Rep Infect Dis 2016:8979656. doi: 10.1155/2016/8979656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Odom AR. 2011. Five questions about non-mevalonate isoprenoid biosynthesis. PLoS Pathog 7:e1002323. doi: 10.1371/journal.ppat.1002323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Wilding EI, Brown JR, Bryant AP, Chalker AF, Holmes DJ, Ingraham KA, Iordanescu S, So CY, Rosenberg M, Gwynn MN. 2000. Identification, evolution, and essentiality of the mevalonate pathway for isopentenyl diphosphate biosynthesis in gram-positive cocci. J Bacteriol 182:4319–4327. doi: 10.1128/JB.182.15.4319-4327.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Lange BM, Rujan T, Martin W, Croteau R. 2000. Isoprenoid biosynthesis: the evolution of two ancient and distinct pathways across genomes. Proc Natl Acad Sci U S A 97:13172–13177. doi: 10.1073/pnas.240454797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Steinbacher S, Kaiser J, Eisenreich W, Huber R, Bacher A, Rohdich F. 2003. Structural basis of fosmidomycin action revealed by the complex with 2-C-methyl-d-erythritol 4-phosphate synthase (IspC). Implications for the catalytic mechanism and anti-malaria drug development. J Biol Chem 278:18401–18407. doi: 10.1074/jbc.M300993200. [DOI] [PubMed] [Google Scholar]
- 36.Heuston S, Begley M, Gahan CG, Hill C. 2012. Isoprenoid biosynthesis in bacterial pathogens. Microbiology 158:1389–1401. doi: 10.1099/mic.0.051599-0. [DOI] [PubMed] [Google Scholar]
- 37.Boucher Y, Doolittle WF. 2000. The role of lateral gene transfer in the evolution of isoprenoid biosynthesis pathways. Mol Microbiol 37:703–716. doi: 10.1046/j.1365-2958.2000.02004.x. [DOI] [PubMed] [Google Scholar]
- 38.Poyart C, Quesne G, Boumaila C, Trieu-Cuot P. 2001. Rapid and accurate species-level identification of coagulase-negative staphylococci by using the sodA gene as a target. J Clin Microbiol 39:4296–4301. doi: 10.1128/JCM.39.12.4296-4301.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Zhang B, Watts KM, Hodge D, Kemp LM, Hunstad DA, Hicks LM, Odom AR. 2011. A second target of the antimalarial and antibacterial agent fosmidomycin revealed by cellular metabolic profiling. Biochemistry 50:3570–3577. doi: 10.1021/bi200113y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Armstrong CM, Meyers DJ, Imlay LS, Freel Meyers C, Odom AR. 2015. Resistance to the antimicrobial agent fosmidomycin and an FR900098 prodrug through mutations in the deoxyxylulose phosphate reductoisomerase gene (dxr). Antimicrob Agents Chemother 59:5511–5519. doi: 10.1128/AAC.00602-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Brown AC, Parish T. 2008. Dxr is essential in Mycobacterium tuberculosis and fosmidomycin resistance is due to a lack of uptake. BMC Microbiol 8:78. doi: 10.1186/1471-2180-8-78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Nair SC, Brooks CF, Goodman CD, Sturm A, Strurm A, McFadden GI, Sundriyal S, Anglin JL, Song Y, Moreno SN, Striepen B. 2011. Apicoplast isoprenoid precursor synthesis and the molecular basis of fosmidomycin resistance in Toxoplasma gondii. J Exp Med 208:1547–1559. doi: 10.1084/jem.20110039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Borrmann S, Issifou S, Esser G, Adegnika AA, Ramharter M, Matsiegui PB, Oyakhirome S, Mawili-Mboumba DP, Missinou MA, Kun JF, Jomaa H, Kremsner PG. 2004. Fosmidomycin-clindamycin for the treatment of Plasmodium falciparum malaria. J Infect Dis 190:1534–1540. doi: 10.1086/424603. [DOI] [PubMed] [Google Scholar]
- 44.Missinou MA, Borrmann S, Schindler A, Issifou S, Adegnika AA, Matsiegui PB, Binder R, Lell B, Wiesner J, Baranek T, Jomaa H, Kremsner PG. 2002. Fosmidomycin for malaria. Lancet 360:1941–1942. doi: 10.1016/S0140-6736(02)11860-5. [DOI] [PubMed] [Google Scholar]
- 45.Beach MJ, Rodwell VW. 1989. Cloning, sequencing, and overexpression of mvaA, which encodes Pseudomonas mevalonii 3-hydroxy-3-methylglutaryl coenzyme A reductase. J Bacteriol 171:2994–3001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Scher DS, Rodwell VW. 1989. 3-Hydroxy-3-methylglutaryl coenzyme A lyase from Pseudomonas mevalonii. Biochim Biophys Acta 1003:321–326. doi: 10.1016/0005-2760(89)90239-7. [DOI] [PubMed] [Google Scholar]
- 47.Begley M, Bron PA, Heuston S, Casey PG, Englert N, Wiesner J, Jomaa H, Gahan CG, Hill C. 2008. Analysis of the isoprenoid biosynthesis pathways in Listeria monocytogenes reveals a role for the alternative 2-C-methyl-d-erythritol 4-phosphate pathway in murine infection. Infect Immun 76:5392–5401. doi: 10.1128/IAI.01376-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Sung JM, Lloyd DH, Lindsay JA. 2008. Staphylococcus aureus host specificity: comparative genomics of human versus animal isolates by multi-strain microarray. Microbiology 154:1949–1959. doi: 10.1099/mic.0.2007/015289-0. [DOI] [PubMed] [Google Scholar]
- 49.Hintz M, Reichenberg A, Altincicek B, Bahr U, Gschwind RM, Kollas AK, Beck E, Wiesner J, Eberl M, Jomaa H. 2001. Identification of (E)-4-hydroxy-3-methyl-but-2-enyl pyrophosphate as a major activator for human gammadelta T cells in Escherichia coli. FEBS Lett 509:317–322. doi: 10.1016/S0014-5793(01)03191-X. [DOI] [PubMed] [Google Scholar]
- 50.Chen ZW. 2013. Multifunctional immune responses of HMBPP-specific Vγ2Vδ2 T cells in M. tuberculosis and other infections. Cell Mol Immunol 10:58–64. doi: 10.1038/cmi.2012.46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Huang D, Shen Y, Qiu L, Chen CY, Shen L, Estep J, Hunt R, Vasconcelos D, Du G, Aye P, Lackner AA, Larsen MH, Jacobs WR Jr., Haynes BF, Letvin NL, Chen ZW. 2008. Immune distribution and localization of phosphoantigen-specific Vgamma2Vdelta2 T cells in lymphoid and nonlymphoid tissues in Mycobacterium tuberculosis infection. Infect Immun 76:426–436. doi: 10.1128/IAI.01008-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Ali Z, Shao L, Halliday L, Reichenberg A, Hintz M, Jomaa H, Chen ZW. 2007. Prolonged (E)-4-hydroxy-3-methyl-but-2-enyl pyrophosphate-driven antimicrobial and cytotoxic responses of pulmonary and systemic Vgamma2Vdelta2 T cells in macaques. J Immunol 179:8287–8296. doi: 10.4049/jimmunol.179.12.8287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Kumar D, Cawley JJ, Irizarry-Alvarado JM, Alvarez A, Alvarez S. 2007. Case of Staphylococcus schleiferi subspecies coagulans endocarditis and metastatic infection in an immune compromised host. Transpl Infect Dis 9:336–338. doi: 10.1111/j.1399-3062.2007.00222.x. [DOI] [PubMed] [Google Scholar]
- 54.Jindal A, Shivpuri D, Sood S. 2015. Staphylococcus schleiferi meningitis in a child. Pediatr Infect Dis J 34:329. doi: 10.1097/INF.0000000000000561. [DOI] [PubMed] [Google Scholar]
- 55.Thawabi M, Jerome M, Slim J, Shamoon F, Boghossian J. 2015. Pericardial effusion and sepsis caused by Staphylococcus schleiferi. J Infect Public Health 8:392–393. doi: 10.1016/j.jiph.2015.01.006. [DOI] [PubMed] [Google Scholar]
- 56.Van Hoovels L, Vankeerberghen A, Boel A, Van Vaerenbergh K, De Beenhouwer H. 2006. First case of Staphylococcus pseudintermedius infection in a human. J Clin Microbiol 44:4609–4612. doi: 10.1128/JCM.01308-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Kuan EC, Yoon AJ, Vijayan T, Humphries RM, Suh JD. 2016. Canine Staphylococcus pseudintermedius sinonasal infection in human hosts. Int Forum Allergy Rhinol 6:710–715. doi: 10.1002/alr.21732. [DOI] [PubMed] [Google Scholar]
- 58.Chin CS, Alexander DH, Marks P, Klammer AA, Drake J, Heiner C, Clum A, Copeland A, Huddleston J, Eichler EE, Turner SW, Korlach J. 2013. Nonhybrid, finished microbial genome assemblies from long-read SMRT sequencing data. Nat Methods 10:563–569. doi: 10.1038/nmeth.2474. [DOI] [PubMed] [Google Scholar]
- 59.R Development Core Team 2010. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. [Google Scholar]
- 60.CLSI 2013. Performance standards for antimicrobial disk and dilution susceptibility tests for bacteria isolated from animals. Approved standard, 4th ed. Clinical and Laboratory Standards Institute, Wayne, PA. [Google Scholar]
- 61.Dereeper A, Guignon V, Blanc G, Audic S, Buffet S, Chevenet F, Dufayard JF, Guindon S, Lefort V, Lescot M, Claverie JM, Gascuel O. 2008. Phylogeny.fr: robust phylogenetic analysis for the non-specialist. Nucleic Acids Res 36:W465–W469. doi: 10.1093/nar/gkn180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Edgar RC. 2004. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res 32:1792–1797. doi: 10.1093/nar/gkh340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Castresana J. 2000. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol Biol Evol 17:540–552. doi: 10.1093/oxfordjournals.molbev.a026334. [DOI] [PubMed] [Google Scholar]
- 64.Guindon S, Dufayard JF, Lefort V, Anisimova M, Hordijk W, Gascuel O. 2010. New algorithms and methods to estimate maximum-likelihood phylogenies: assessing the performance of PhyML 3.0. Syst Biol 59:307–321. doi: 10.1093/sysbio/syq010. [DOI] [PubMed] [Google Scholar]
- 65.Chevenet F, Brun C, Bañuls AL, Jacq B, Christen R. 2006. TreeDyn: towards dynamic graphics and annotations for analyses of trees. BMC Bioinformatics 7:439. doi: 10.1186/1471-2105-7-439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Kanehisa M, Goto S. 2000. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res 28:27–30. doi: 10.1093/nar/28.1.27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Kanehisa M, Goto S, Sato Y, Kawashima M, Furumichi M, Tanabe M. 2014. Data, information, knowledge and principle: back to metabolism in KEGG. Nucleic Acids Res 42:D199–D205. doi: 10.1093/nar/gkt1076. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Long sequence reads assisted in assembling long repetitive regions. (a) Long reads generated by single-molecule real-time sequencing span a 5-kb rRNA operon and nearby coding regions. Red, yellow, and blue bars represent rRNA genes, tRNA genes, and protein-coding sequences, respectively. Black and gray lines indicate sequences that map to strain 5909-02 in the forward and reverse direction, respectively (total coverage in this region is ~90×). (b) Alignment of reads in the clustered regularly interspaced short palindromic repeat (CRISPR) region. Blue bars and pink triangles represent protein-coding cas genes and 36-bp CRISPR repeat sequences, respectively. Black and gray lines indicate sequences that map across the entire CRISPR repeat-spacer region in strain 1360-13 in the forward and reverse direction, respectively (total coverage in this region is ~80×). The genomes are ordered from smallest number of repeat sequences (4; strain 2142-05) to largest (36; strain 1360-13). Download Figure S1, PDF file, 0.8 MB (886.9KB, pdf) .
Copyright © 2016 Misic et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Antimicrobial susceptibility and metadata for S. schleiferi strains (the S. schleiferi strain metadata are shown, including date of collection, body site, and antibiotic susceptibility profiles; the antibiotic susceptibilities were determined using the Microscan Walkaway 40 PC20 Gram-positive combo-panel [Dade Behring, Sacramento, CA, USA]). Download Table S1, DOCX file, 0.02 MB (17.3KB, docx) .
Copyright © 2016 Misic et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
RAST-annotated subsystems of 14 Staphylococcus genomes (from a total of 382 genomes, 147 subsystems were conserved, with an equal number of genes present in each subsystem across all 14 strains examined; genes in 235 subsystems were differentially abundant among two or more species). Download Table S2, XLSX file, 0.03 MB (32.9KB, xlsx) .
Copyright © 2016 Misic et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Select adhesin and antimicrobial resistance genes found among 14 Staphylococcus genomes (the presence or absence of selected virulence genes in the respective genomes is indicated with a + or − symbol). Download Table S3, XLSX file, 0.02 MB (26.4KB, xlsx) .
Copyright © 2016 Misic et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Isoprenoid biosynthesis genes in 14 Staphylococcus genomes (the mevalonate and nonmevalonate pathway gene names, abbreviations, enzyme catalog numbers, and locus identifiers of select Staphylococcus species are shown). Download Table S4, XLSX file, 0.05 MB (47.7KB, xlsx) .
Copyright © 2016 Misic et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.