

Abstract
Evaluation of the radio‐transmission pH‐measurement system for monitoring the ruminal pH and subacute ruminal acidosis (SARA) in cattle is described. This is done in order to reveal the possible application of this system for detection and pathophysiological research of SARA by continuous ruminal pH measurement. The possibility of using this system for assessment of the ruminal pH in SARA cattle, and the presence of negative correlation between the ruminal pH and ruminal temperature in heathy and SARA cattle were determined. In addition, the 16S rRNA gene pyrosequencing analysis showed that the ruminal microbial community was simpler in SARA cattle, and the bacterial numbers in SARA cattle were lower than those in healthy hay‐fed cattle. Concentrate feeding might have reduced the diversity of the ruminal microbial community. Changes in the ruminal microbial community of SARA cattle might be related to the changes in ruminal pH followed by the decrease in the number of some bacteria. Continuous monitoring of the ruminal pH using the radio‐transmission pH‐measurement system would be applied for detection and prevention of SARA in the field and pathophysiological research of SARA, including ruminal zymology and bacteriology, which have been determined previously by sampling of the ruminal fluid and measuring of ruminal pH.
Keywords: cattle, continuous monitoring, ruminal pH, subacute ruminal acidosis (SARA)
Introduction
Cows with subacute ruminal acidosis (SARA) often develop complications of laminitis, reduced and erratic feed intake, low body condition score, low milk fat syndrome, abomasal displacement, ulceration and ruminitis (Enemark 2008; Plaizier et al. 2009). Furthermore, SARA is associated with immunosuppression (Kleen et al. 2003) and inflammation (Khafipour et al. 2009a,2009b; Plaizier et al. 2009). Free lipopolysaccharide (LPS) increases in the rumen during SARA (Gozho et al. 2007; Emmanuel et al. 2008), and the LPS translocates from the rumen into blood circulation, which activates an inflammatory response (Khafipour et al. 2009a,2009b) affecting peripheral leukocyte populations and production of pro‐inflammatory cytokines. Therefore, monitoring ruminal pH and prevention of acidic pH are very important challenges in the dairy industry.
SARA is characterized by repeated bouts of low ruminal pH (Enemark et al. 2004; Enemark 2008; Plaizier et al. 2009), but accurate measurement of the ruminal pH is very difficult. In order to obtain the ruminal fluid for pH measurement, oro‐ruminal probe and rumenocentesis are used (Duffield et al. 2004; Bramley et al. 2008). Recently, an indwelling ruminal pH system was developed and placed in the rumen of fistulated cattle (Marden et al. 2005; AlZahal et al. 2007; Penner et al. 2006, 2009). Although an indwelling ruminal pH system has made continuous recording of ruminal pH possible (Keunen et al. 2002; Nocek et al. 2002a,2002b; Rustome et al. 2006), this system can be used only for experimentally fistulated cows. Therefore, this system does not allow the field application of SARA evaluation. Based on these backgrounds, we recently have invented a new radio‐transmission pH‐measurement system, and assessed its performance and usefulness in a continuous evaluation of ruminal pH (Sato et al. 2012a, b).
In the cows exposed to SARA challenge, the ruminal pH remained very low during the first day of SARA challenge, but mid‐lactating cows fed a 60% forage diet recovered to the normal pH more quickly than early lactating cows fed a 45% forage diet (Dohme et al. 2008). Decrease in ruminal pH on the day of SARA challenge was getting severer with each successive challenge. Authors speculated that the increased severity of SARA with repeated challenges might be due to short recovery time provided to the experimental cows between challenge periods. However, changes in the ruminal pH and volatile fatty acid (VFA) of cattle with repeated acidosis challenges are not adequately known. Furthermore, several studies have been conducted on the rumen microbial community in cows with SARA (Chen et al. 2012; Hernandez‐Sanabria et al. 2012; Mao et al. 2013; Golder et al. 2014). The most common bacteria detected in the rumen of cattle with SARA are Lactobacillus spp. and Streptococcus bovis (Nagaraja & Titgemeyer 2007). The dominant bacterial phyla include Firmicutes, Bacteroidetes and all subgroups of Proteobacteria. Recently, pyrosequencing has been used for assessment of microbial composition in the rumen of cattle (Jami & Mizrahi 2012; Rice et al. 2012). These studies supported the notion that pyrosequencing facilitated greater understanding of microbial diversity. Most of the studies about microbial communities in SARA cattle are based on single induction of SARA. The microbial community in cattle with repeated induction of SARA is unknown.
In this review, our radio‐transmission pH‐measurement system for continuous monitoring of ruminal pH and SARA in cattle is reported, to reveal the possible application of this system for detection and pathophysiological research of SARA. In addition, our recent research about the changes in the ruminal pH, VFA and microbial communities in cattle associated with repeatedly induced SARA, as well as the effects of Propionibacterium‐based probiotics on the ruminal pH and VFA in cattle are presented. The experimental designs described in this review were approved by the Iwate University Laboratory Animal Care and Use committee.
Continuous monitoring of ruminal pH by a radio transmission measurement system
The radio‐transmission pH‐measurement system consists of a wireless pH sensor, a data measurement receiver, a relay unit and a personal computer with special software (YCOW‐S; DKK‐Toa Yamagata) (Sato et al. 2012b) installed. The wireless system utilized a specified low‐power radio transmission system (429 MHz band), which could be operated without a license in accordance with Japanese Radio Law. The mean variations of the measurements by the glass pH electrode were + 0.20 after 2 months of continuous recording, compared to the values confirmed by a standard pH solution at the start of the recording. The mean lifetime of the internal battery was 3 months when measurements were continuously transmitted every 10 min. There was a good correlation between continuous recording and spot sampling (r = 0.986, P < 0.01), when the ruminal pH recorded by our new system was compared to that of the spot sampling of ruminal fluid (Sato et al. 2012b). We have demonstrated the possible application of the radio‐ transmission pH‐measurement system for assessment and monitoring of the ruminal pH in cows. Our new system might contribute to accurate assessment and prevention of SARA. In this chapter, our experimental results of basic tests for performance of radio‐transmission pH‐measurement system are described.
Changes in ruminal pH in cattle fed different diets
The ruminal pH was measured continuously by the radio‐transmission pH‐measurement system in cattle, which were fed three kinds of diets, 1 week for each. Four rumen‐cannulated Holstein steers, aged 7 months, were used. Cattle were fed orchard grass mixed hay and concentrate (50% flaked barley and 50% corn). The ratios of roughage and concentrate were 10:0 (A‐diet), 2:8 (B‐diet) and 5:5 (C‐diet), respectively. The pH sensor was placed in the ventral sac of the rumen through rumen‐fistula, and pH was measured at 10‐min intervals for 4 weeks.
The ruminal pH decreased markedly and rapidly when feeding changed from A‐ to B‐diet, but it returned to the basal level when feeding changed from B‐ to C‐diet (Fig. 1). Thereafter, another pattern of the ruminal pH changes was observed when B‐diet was fed the second time showing moderate decrease in pH compared to that of the first B‐diet feeding. Within 1 week of B‐diet feeding, the decreased ruminal pH recovered slightly. In addition, the changes in the ruminal pH after feeding the same diet varied among individual cows.
Figure 1.
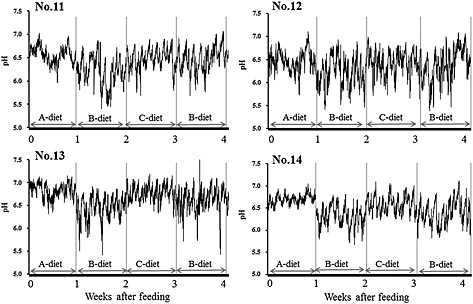
Continuous changes in ruminal pH of cattle fed a different diet 1 week for each. The ruminal pH was measured continuously at 10 min intervals. Ratio of roughage and concentrate in the three diets were 8:2 (A‐diet), 2:8 (B‐diet) and 5:5 (C‐diet), respectively.
These results indicated that the SARA could be induced successfully by feeding the B‐diet. Changes of the pH values after the morning feeding in this study were similar to previous studies (Krause & Oetzel 2005; Keunen et al. 2002). These results demonstrated the possible use of the radio‐transmission pH‐measurement system for the assessment and monitoring of ruminal pH of cows in field conditions. It is suggested that the ruminal pH could be recoverable after rapid feeding changes by adaptation of ruminal fermentation.
Relations between pH and temperature, VFA in ruminal fluid
Ruminal temperature rises during the early phase of disease, estrus and onset of calving (Cooper‐Prado et al. 2011; Rose‐Dye et al. 2011). In addition, the ruminal temperature exceeds rectal temperature presumably due to accelerated microbial fermentation (AlZahal et al. 2008, 2009). Ruminal temperature is negatively correlated with the ruminal pH in cows with SARA, and a predictive equation for aiding the diagnosis of SARA has been sought based on the negative correlation between the lowest ruminal pH and high rumen temperature (AlZahal et al. 2008, 2009). However, it is still controversial with regard to whether an accurate ruminal pH can be predicted by measuring ruminal temperature because it is not clear when the lowest pH and the highest temperature appear.
To assess the relationship between pH and temperature in the ruminal bottom fluid, circadian changes were monitored using cattle fed a control diet (C‐diet) or rumen acidosis‐inducing (RAI) diet by using the radio‐transmission pH‐measurement system (Kimura et al. 2012a). Eight primiparous non‐lactating, rumen‐cannulated Holstein cattle weighting 620‐720 kg were used. Both diets were composed of orchard grass and timothy mixed hay and concentrate (equivalent amount of flaked barley and corn). Each cow was given 5.5‐6.5 kg twice daily on a dry matter (DM) basis for 2 weeks. The ratios of roughage to concentrate for C‐diet and RAI‐diet were 9:1 and 2:8. The pH sensor was placed in the ventral sac of the rumen, and two parameters were measured simultaneously at 10‐min intervals for 14 days after commencement of feeding.
Compared to the mean ruminal pH for 60 min immediately after the morning feeding (0 h), significantly lower pH was noted 3‐13 h later (P < 0.05) and 4‐19 h later (P < 0.01) in cattle fed C‐ and RAI‐diets, respectively, although the reduction in the latter was much higher than that in the former (Fig. 2). In contrast, significantly higher ruminal temperature was found at 8 and 12 14 h later (P < 0.05) and 6, 8 and 10‐19 h later (P < 0.01) in cattle fed C‐ and RAI‐diets, respectively. A significant negative correlation was observed between the lowest ruminal pH and its corresponding ruminal temperature in cattle fed C‐ and RAI‐diets (r = ‐0.722 and ‐0.650, respectively, P < 0.01), suggesting active fermentation and VFA production in the rumen. However, the ruminal pH profiles may not be predictable by measuring only ruminal temperature because decreases in pH were preceded by increases in temperature, and the circadian changes in pH and temperature were associated with ruminal fermentation. Meanwhile, ruminal lactic acid and total VFA concentrations increased significantly or tended to increase in cattle fed an RAI‐diet, whereas ruminal ammonia N concentrations decreased significantly in cows fed an RAI‐diet, although there was variation. Significant (P < 0.05) negative correlations were observed between the ruminal pH and concentrations of total VFA (r = −0.681) or its components such as acetic acid (r = −0.699), propionic acid (r = −0.575) and butyric acid (r = −0.638) in cows fed a C‐diet. A significant (P < 0.05) negative correlation was observed between the ruminal temperature and lactic acid concentration (r = −0.582) in cows fed an RAI‐diet.
Figure 2.
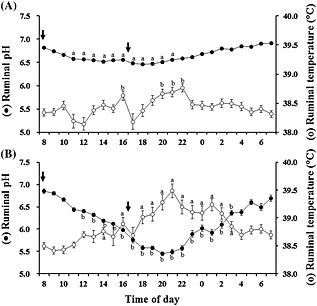
Circadian changes in 1‐h mean ruminal pH (black circles) and temperature (white circles) in cattle fed the control diet (C‐diet, A) or the rumen acidosis‐inducing diet (RAI‐diet, B). Arrows indicate the feeding times. Values represent mean ± SE (n = 8). a P < 0.05, b P < 0.01 (significant difference compared to the 1‐h mean at 08.00‐09.00 hours in each group).
Ruminal lactic acid is not the cause of pH reduction in SARA model cows (Keunen et al. 2002). Some SARA cows have normal ruminal lactate concentrations, indicating that elevated VFA concentration is one of the main causes of low ruminal pH. In our study, a negative correlation has been observed between the ruminal pH and ruminal temperature in cattle fed the RAI‐diet, as previously described (AlZahal et al. 2008). In addition, significant negative correlations were observed between the ruminal pH and VFA concentrations in cattle fed the C‐diet. The negative correlation was also observed between the ruminal temperature and lactic acid concentration in cattle fed the RAI‐diet. Elevated total VFA concentration is the main cause of low ruminal pH in cows with SARA (Keunen et al. 2002). Therefore, the ruminal temperature might increase and the ruminal pH might decrease simultaneously by rapid fermentation and VFA production after RAI‐diet feeding.
Simultaneous estimation of pH in the rumen and reticulum
The reliability of ruminal pH depends on the sampling site (bottom vs. middle of the rumen and rumen vs. reticulum) and sampling procedure (rumenocentesis vs. an oro‐ruminal probe) (Duffield et al. 2004; Tafaj et al. 2004; Bramley et al. 2008). An indwelling wired system has been used to measure pH at the bottom of the rumen (Marden et al. 2005; AlZahal et al. 2007; Penner et al. 2009), but there have been no reports on simultaneous pH measurements of fluids at the bottom and in the middle of the rumen and in the reticulum (i.e. three sites) of cows. Furthermore, a few reports had examined the pH of reticulum fluid (Bryant 1964; Lane et al. 1968). In these reports the highest pH in the reticulum and the lowest pH at the top or middle of the rumen were reported (Lane et al. 1968).
To assess the site difference of pH in the rumen and reticulum, circadian pH changes in the fluid of the rumen (bottom and middle) and reticulum were assessed simultaneously using wireless and wired radio‐transmission pH‐measurement systems in cattle fed the control diet (C‐diet) or the RAI diet (Kimura et al. 2012b; Sato et al. 2012c). Cattle (n = 8) and diets (C‐diet and RAI‐diet) used in this study were the same as a previous study, which investigated the relation between ruminal pH and temperature. Wireless pH sensor was placed in the ventral sac at the bottom of the rumen and in the reticulum. Wired pH sensor was fixed 10‐20 cm below the ruminal mat surface (middle of the rumen) by a stout wire through a rumen cannula.
The pH in the three sites decreased following the morning and evening feedings (Fig. 3). In cattle fed the C‐diet, the bottom‐rumen and reticular pH reverted to the basal level by the next morning, while the middle‐rumen pH did not recover completely, suggesting that active fermentation occurred in the middle of the rumen. The mean pH at 1‐h intervals was higher in the reticulum than at the bottom and in the middle of the rumen. The relatively stable reticular pH may result from dilution due to salivation and drinking water, as previously described (Dado & Allen 1993). In cows fed the RAI‐diet, the bottom‐rumen pH fell to approximately 5.2 after the evening feeding, but returned to the basal level by the next morning. In contrast, the middle‐rumen pH did not return to the basal level (6.5) within 24 h, presumably owing to continuous and vigorous fermentation. Significant (P < 0.01) positive correlations were observed in the pH of cattle fed the RAI‐diet between in the middle and at the bottom of the rumen (r = 0.894), in the middle of the rumen and in the reticulum (r = 0.764) and at the bottom of the rumen and in the reticulum (r = 0.797).
Figure 3.
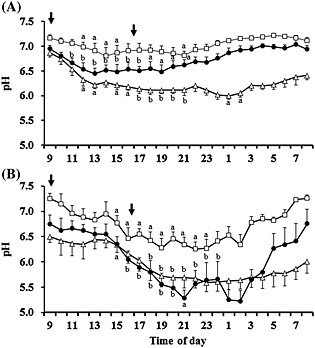
Circadian pH changes at the bottom (closed circles) and in the middle (open triangles) of the rumen and in the reticulum (open squares) of cattle fed control diet (C‐diet, A) or the rumen acidosis‐inducing diet (RAI‐diet, B). Arrows show the feeding times. Values are expressed as mean ± SE (n = 4). a P < 0.05, b P < 0.01 (significant difference compared to the corresponding basal pH (taken at 09.00 hours)).
Pathophysiological evaluation of SARA by continuous monitoring of ruminal pH
SARA is characterized by ruminal pH (Enemark 2008; Plaizier et al. 2009) lower than 5.6, and pH 5.8 is considered as the marginal threshold for SARA (Kleen et al. 2003; Gozho et al. 2005, 2007). In an experiment of successive SARA induction, decreased ruminal pH was much severer in the day of challenge feeding (Dohme et al. 2008). Increased severity of SARA might have been induced by instability of the ruminal microflora (Nagaraja & Titgemeyer 2007), and decreased absorptive capacity of the ruminal epithelium might also have contributed to VFA accumulation in the rumen. A short‐term, severe insult of acute acidosis decreased ruminal VFA absorption, presumably caused by damage to the ruminal wall (Krehbiel et al. 1995). Decreased ruminal absorptive capacity as a result of acidosis challenge may increase the severity of subsequent acidosis challenges. However, changes in the ruminal pH and VFA of cattle with repeated acidosis challenges are unknown. In addition, molecular techniques have shown that the ruminal microbial community is highly diverse (Mao et al. 2013), but the factors influencing its composition remain unknown. The ruminal microbial community in SARA cows has been reported (Chen et al. 2012; Hernandez‐Sanabria et al. 2012; Golder et al. 2014), but the relationship between the ruminal pH, VFA and microbiota composition in SARA cattle is unclear.
Bacteria‐based probiotics are known to have positive implications on animal health and performance. Bacterial probiotics have been shown to improve the rumen‐predominant microorganisms (Ghorbani et al. 2002; Chiquette et al. 2012; Qadis et al. 2014a) and ruminal performance in cattle (Qadis et al. 2014b, 2014c). Steers that received probiotics containing both lactate‐utilizing Propionibacterium and lactate‐producing Enterococcus had higher ruminal concentrations of acetate, and the blood variables indicated a reduced risk of metabolic acidosis (Ghorbani et al. 2002). However, little information is available about the effect of probiotics containing Propionibacterium on ruminal pH and VFA in cattle. Changes in the ruminal pH, VFA and microbial community in cattle with repeatedly induced SARA and effects of Propionibacterium‐based probiotics on the ruminal pH and VFA in SARA cattle should be assessed pathophysiologically for the possible association with SARA by continuous monitoring of ruminal pH.
Changes in ruminal pH, VFA and microbiota composition in cattle with repeatedly induced SARA
The ruminal pH, VFA and microbial composition in cattle with repeatedly induced SARA were investigated to reveal the ruminal adaptation to acute and short‐term changes in feeding. The 16S rRNA gene pyrosequencing and real‐time PCR were used to analyze the ruminal microbial community in SARA cattle, and to compare the composition of microbial community in cattle fed hay or high‐concentrate diet.
Eight rumen‐cannulated Holstein steers (age, 8‐10 months; body weight, 180‐200 kg) were used and fed hay or RAI diet (20% hay and 80% concentrate) each for 7 days. Each steer was fed either hay or the RAI‐diet for 7 days, 2‐3 kg of feed twice daily on a DM basis, followed by the other diet for an additional 7 days without a resting period. This experiment was continuously performed twice. The pH sensor was placed in the ventral sac of the rumen and pH was measured every 10 min. Ruminal fluid was collected three times (08.00, 14.00 and 20.00 hours) at 7 days after starting each feeding period. Total VFA and VFA components were separated and quantified by gas chromatography. The microbial community of the ruminal fluids was analyzed by 16S rRNA arrangement analysis (four of eight cattle in the first experimental period) and real time PCR (four of eight cattle in both experimental periods) using bacterial DNA extraction from fluids collected at 20.00 hours.
The 24‐h mean ruminal pH values in the concentrate‐fed (CF) period were significantly (P < 0.05) lower than those in the hay‐fed (FF) period, and the pH values obtained at 3 days after concentrate feeding were slightly lower in the first experiment compared to the second experiment (Fig. 4). The 1‐h mean pH values of the ruminal fluids decreased gradually following morning and evening feedings in both FF and CF periods, and the pH values in the CF period were lower overall than those in the FF period. Ruminal total VFA, propionic acid and butyric acid concentrations, and LPS activity in the CF period were slightly higher than those in the FF period (Fig. 5). However, no significant difference was observed in the ruminal total VFA, concentration of each VFA and lactic acid, as well as LPS activity between the first and second experiments.
Figure 4.
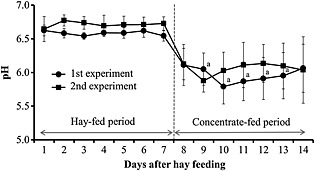
Changes in 24‐h mean ruminal pH of cattle fed hay and concentrate at the first (black circles) and second (black squares) experiment. Values represent mean ± SE (n = 8). a P < 0.05 (significant difference compared to the 24‐h mean at 7 days after hay feeding in the first experiment).
Figure 5.
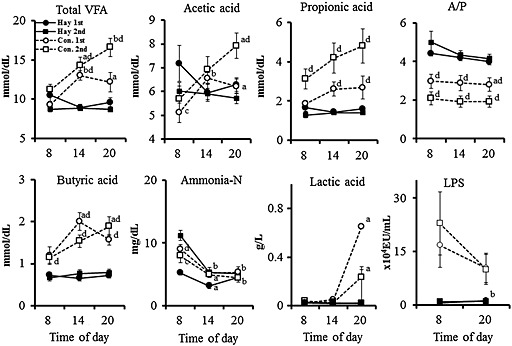
Circadian changes in total volatile fatty acids (VFA), each VFA component, ammonia N and lactic acid concentrations, and lipopolysaccharide (LPS) activity of cattle fed hay and concentrate in the first and second experiment. Values represent mean ± SE (n = 8). a P < 0.05, b P < 0.01 (significant difference compared to the value at 08:00 hours in each feeding group). c P < 0.05, d P < 0.05 (significant difference compared to the value of first experiment in each feeding).
The 16S rRNA gene pyrosequencing analysis showed that the ruminal microbial community was simpler in the CF period than in the FF period. Detected numbers of the bacterial phylum, genus and species in the CF period were significantly (P < 0.05) lower than those in the FF period (Table 1). At the phylum level, Firmicutes, Bacteroidetes and Actinobacteria were dominant in during both FF and CF periods (Fig. 6). The percentages of Bacteroidetes and Proteobacteria were reduced by concentrate feeding, whereas Firmicutes was more abundant in the CF period than the FF period. At the genus level, Prevotella, Clostridium, Butyrivibrio, Eubacterium and Ruminococcus were dominant in both feeding periods. As compared with the FF period, the ratios of Prevotella and Eubacterium were lower, while the percentages of Ruminococcus, Clostridium and Butyrivibrio increased in the CF period. The real‐time PCR analysis showed that the bacterial numbers of total methanogens, Fibrobacter succinogenes and Selenomonas ruminantium, significantly (P < 0.05) decreased, and those of Streptococcus bovis and Megasphaera elsdenii significantly (P < 0.05) increased in the CF period compared to the FF period (Fig. 7). In the FF period, the bacterial numbers of total bacteria, total methanogens and Selenomonas ruminantium in the second experimental period were significantly (P < 0.05) lower than those in the first experiment.
Table 1.
Number of bacterial phylum, genus and species in cattle fed hay and concentrate in the first experiment by the 16S rRNA gene pyrosequencing
| Phylum1 | Genus1 | Species1 | ||||
|---|---|---|---|---|---|---|
| Cattle No. | Hay‐fed | Concentrate‐fed | Hay‐fed | Concentrate‐fed | Hay‐fed | Concentrate‐fed |
| 1 | 16 | 11 | 144 | 97 | 371 | 229 |
| 2 | 16 | 7 | 145 | 75 | 325 | 165 |
| 3 | 11 | 9 | 132 | 77 | 317 | 177 |
| 4 | 10 | 5 | 97 | 81 | 237 | 163 |
| mean | 13.3 | 8a | 129.5 | 82.5a | 312.5 | 183.5a |
| SE | 1.6 | 1.3 | 11.2 | 5.0 | 27.8 | 15.5 |
Bacterial number detected by 16S rRNA pyrosequence
P < 0.05 (significant difference compared to the hay‐fed period
Figure 6.
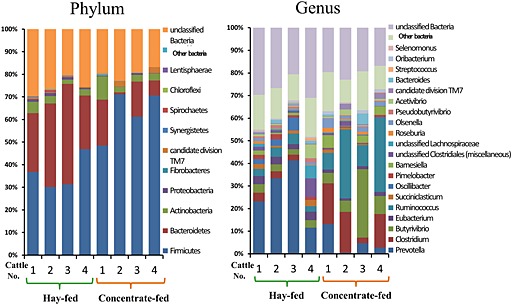
Comparison of rumen microbial community in cattle (n = 4) fed hay and concentrate in the first experiment by the 16S rRNA gene pyrosequencing. The microbial composition was analyzed by the 16S rRNA arrangement analysis using bacterial DNA extraction from fluids collected at 20.00 hours.
Figure 7.
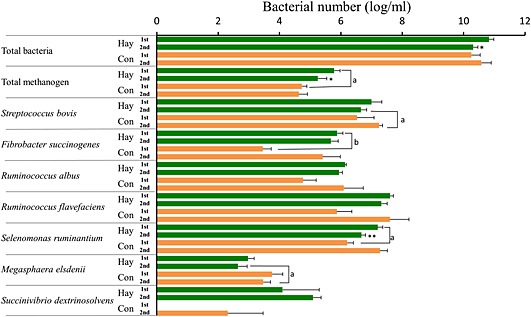
Comparison of rumen bacterial numbers in cattle (n = 4) fed hay and concentrate (Con) in the first and second experiment by real‐time PCR. The bacterial number was analyzed using bacterial DNA extraction from fluids collected at 20.00 hours. Values represent mean ± SE (n = 4). * P < 0.05, ** P < 0.01 (significant difference compared to the first experiment in the same feeding), a P < 0.05, b P < 0.01 (significant difference between the hay‐ and concentrate‐feeding within the same experimental period).
In the CF period, both ruminal pH and bacterial diversity were lower and the rumen microbial composition was simpler than those in the FF period. Concentrate feeding might have reduced the diversity of the rumen microbial community. Changes in the rumen microbial composition of SARA cattle might be related to the marked changes in ruminal pH followed by the rapid decrease in numbers of bacteria. Changes in ruminal pH and rumen microbial community were observed markedly by acute and short‐term change in feeding, which indicated ruminal adaptation (Mao et al. 2013). However, further research is needed to reveal the relation between ruminal pH and microbial community.
Effects of Propionibacterium‐based probiotic on ruminal pH and VFA in cattle
Propionibacteria are natural inhabitants of the rumen and they produce propionate, a major precursor for glucose production, through hepatic gluconeogenesis (Sauer et al. 1989). Feeding of propionibacteria may increase hepatic glucose production and positively influence host metabolism (Francisco et al. 2002). The effects of propionibacteria feeding on the lactational performance of dairy cows in early lactation have been reported (Stein et al. 2006; Lehloenya et al. 2008; Weiss et al. 2008), and the cattle that received probiotics containing lactate‐utilizing Propionibacterium had higher ruminal concentrations of acetate (Ghorbani et al. 2002). However, little information is available regarding the effect of probiotics containing Propionibacterium on the ruminal pH and VFA in cattle. To evaluate the preventive effect of Propionibacterium on ruminal pH and SARA, changes in the ruminal pH and VFA of cattle administered Propionibacterium‐based probiotic were investigated.
Four primiparous non‐lactating, rumen‐cannulated Holstein cattle weighing 620‐700 kg were used, and the experiment was performed three times at 4‐week intervals. Cattle were fed hay over the 2 weeks before the experiment. Thereafter, the RAI‐diet consisting of 20% hay and 80% concentrate was fed for 2 weeks. Each cow was given 6‐7 kg feed twice daily on a DM basis. During the RAI‐died feeding, two groups of cattle were fed simultaneously either with a Propionibacterium‐based probiotic (5×1010 colony forming units) (A‐diet), or with a Propionibacterium‐based probiotic plus fermented products of Lactobacillus (10 g) and lactose (23.5 g) (B‐diet) once a day. Cattle fed the RAI‐diet without probiotic served as the control. The ruminal pH was measured at 10‐min intervals using the pH sensor, and the ruminal fluids for VFA analysis were collected at 14 days after RAI‐diet feeding.
The mean 24‐h ruminal pH values decreased markedly and showed significantly (P < 0.01) low levels after RAI‐diet feeding in the control and two probiotic feeding groups. However, the pH values in the B‐diet group at 1‐3 and 11 days after RAI‐diet feeding were significantly (P < 0.01) higher than those of the control (Fig. 8). No significant difference was observed in the ruminal VFA concentrations between both feeding groups and the control, whereas the ruminal total VFA and VFA concentrations including propionic acid in cattle fed Propionibacterium‐based probiotic (A‐ and B‐diet) were slightly higher than those in the control (Fig. 9).
Figure 8.
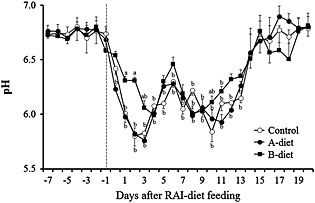
Changes in 24‐h mean ruminal pH of cattle fed the rumen acidosis‐inducing diet (RAI)‐diet (Control), RAI‐diet plus Propionibacterium‐based probiotic (A‐diet), or RAI‐diet plus Propionibacterium‐based probiotic, fermented products of Lactobacillus, and lactose (B‐diet) once a day. Values represent mean ± SE (n = 4). a P < 0.01 (significant difference compared to the control on the same day), b P < 0.01 (significant difference compared to the day ‐1 within the same group).
Figure 9.
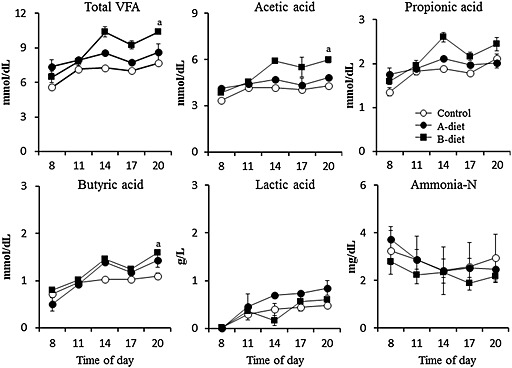
Circadian changes in total volatile fatty acids (VFA), each VFA component, lactic acid and ammonia N concentrations of cattle fed the rumen acidosis‐inducing diet (RAI)‐diet (Control), RAI‐diet plus Propionibacterium‐based probiotic (A‐diet), or RAI‐diet plus Propionibacterium‐based probiotic, fermented products of Lactobacillus, and lactose (B‐diet) once a day. Values represent mean ± SE (n = 4). a P < 0.01 (significant difference compared to the 08.00 hours within the same group).
These results indicate that the decreased ruminal pH resulting from feeding the RAI‐diet was relieved by the administration of Propionibacterium‐based probiotic, fermented products of Lactobacillus and lactose. Propionibacterium‐based additive might improve the ruminal fermentation and reduce ruminal pH in SARA cattle.
Conclusion
Continuous monitoring of the ruminal pH using a newly invented radio‐transmission pH‐measurement system could be applied for detection and prevention of SARA in the field and pathophysiological research of SARA, including the ruminal zymology and bacteriology, which have been determined previously by sampling of the ruminal fluid and measuring of ruminal pH.
Acknowledgments
The author would like to express sincere thanks to Meiji Feed CO., LTD for the kind gift of Propionibacterium‐based probiotic and fermented products of Lactobacillus.
Sato, S. (2016) Pathophysiological evaluation of subacute ruminal acidosis (SARA) by continuous ruminal pH monitoring. Anim Sci J, 87: 168–177. doi: 10.1111/asj.12415.
References
- AlZahal O, Rustomo B, Odongo NE, Duffield TF, McBride BW. 2007. Technical note: A system for continuous recording of ruminal pH in cattle. Journal of Animal Science 85, 213–217. [DOI] [PubMed] [Google Scholar]
- AlZahal O, Kebreab E, France J, Froetschel M, McBride BW. 2008. Ruminal temperature may aid in the detection of subacute ruminal acidosis. Journal of Dairy Science 91, 202–207. [DOI] [PubMed] [Google Scholar]
- AlZahal O, Steele MA, Valdes EV, McBride BW. 2009. Technical note: The use of a telemetric system to continuously monitor ruminal temperature and to predict ruminal pH in cattle. Journal of Dairy Science 92, 5697–5701. [DOI] [PubMed] [Google Scholar]
- Bramley E, Lean IJ, Fulkerson WJ, Stevenson MA, Rabiee AR, Costa ND. 2008. The definition of acidosis in dairy herds predominantly fed on pasture and concentrates. Journal of Dairy Science 91, 308–321. [DOI] [PubMed] [Google Scholar]
- Bryant AM. 1964. Variations in the pH and volatile fatty acid concentration within the bovine reticulo‐rumen. New Zealand Journal of Agricultural Research 7, 694–706. [Google Scholar]
- Chen Y, Oba M, Guan LL. 2012. Variation of bacterial communities and expression of toll‐like receptor genes in the rumen of steers differing in susceptibility to subacute ruminal acidosis. Veterinary Microbiology 159, 451–459. [DOI] [PubMed] [Google Scholar]
- Chiquette J, Allison MJ, Rasmussen M. 2012. Use of Prevotella bryantii 25A and a commercial probiotic during subacute acidosis challenge in midlactation dairy cows. Journal of Dairy Science 95, 5985–5995. [DOI] [PubMed] [Google Scholar]
- Cooper‐Prado MJ, Long NM, Wright EC, Goad CL, Wettemann RP. 2011. Relationship of ruminal temperature with parturition and estrus of beef cows. Journal of Animal Science 89, 1020–1027. [DOI] [PubMed] [Google Scholar]
- Dado RG, Allen MS. 1993. Continuous computer acquisition of feed and water intakes, chewing, reticular motility, and ruminal pH of cattle. Journal of Animal Science 76, 1589–1600. [Google Scholar]
- Dohme F, DeVries TJ, Beauchemin KA. 2008. Repeated ruminal acidosis challenges in lactating dairy cows at high and low risk for developing acidosis: Ruminal pH. Journal of Dairy Science 91, 3554–3567. [DOI] [PubMed] [Google Scholar]
- Duffield T, Plaizier JC, Fairfield A, Bagg R, Vessie G, Dick P, et al. 2004. Comparison of techniques for measurement of rumen pH in lactating dairy cows. Journal of Dairy Science 87, 59–66. [DOI] [PubMed] [Google Scholar]
- Emmanuel DGV, Dunn SM, Ametaj BN. 2008. Feeding high proportions of barley grain stimulates an inflammatory response in dairy cows. Journal of Dairy Science 91, 606–614. [DOI] [PubMed] [Google Scholar]
- Enemark JMD. 2008. The monitoring, prevention and treatment of sub‐acute ruminal acidosis (SARA): A review. Veterinary Journal 176, 32–43. [DOI] [PubMed] [Google Scholar]
- Enemark JMD, Jørgensen RJ, Kristensen NB. 2004. An evaluation of parameters for the detection of subclinical rumen acidosis in dairy herds. Veterinary Research Communication 28, 687–709. [DOI] [PubMed] [Google Scholar]
- Francisco CC, Chamberlain CS, Waldner DN, Wettemann RP, Spicer LJ. 2002. Propionibacteria fed to dairy cows: Effects on energy balance, plasma metabolites and hormones, and reproduction. Journal of Dairy Science 85, 1738–1751. [DOI] [PubMed] [Google Scholar]
- Ghorbani GR, Morgavi DP, Beauchemin KA, Leedle JAZ. 2002. Effects of bacterial direct‐fed microbials on ruminal fermentation, blood variables, and the microbial populations of feedlot cattle. Journal of Animal Science 80, 1977–1985. [DOI] [PubMed] [Google Scholar]
- Golder HM, Denman SE, McSweeney C, Celi P, Lean IJ. 2014. Ruminal bacterial community shifts in grain‐, sugar‐, and histidine‐challenged dairy heifers. Journal of Animal Science 97, 5131–5150. [DOI] [PubMed] [Google Scholar]
- Gozho GN, Plaizier JC, Krause DO, Kennedy AD, Wittenberg KM. 2005. Subacute ruminal acidosis induces ruminal lipopolysaccharide endotoxin release and triggers an inflammatory response. Journal of Dairy Science 88, 1399–1403. [DOI] [PubMed] [Google Scholar]
- Gozho GN, Krause DO, Plaizier JC. 2007. Ruminal lipopolysaccharide concentration and inflammatory response during grain‐induced subacute ruminal acidosis in dairy cows. Journal of Dairy Science 90, 856–866. [DOI] [PubMed] [Google Scholar]
- Hernandez‐Sanabria E, Goonewardene LA, Wang Z, Durunna ON, Moore SS, Guan LL. 2012. Impact of feed efficiency and diet on adaptive variations in the bacterial community in the rumen fluid of cattle. Applied and Environmental Microbiology 78, 1203–1214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jami E, Mizrahi I. 2012. Composition and similarity of bovine rumen microbiota across individual animals. PLoS One 7, e33306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keunen JE, Plaizier JC, Kyriazakis L, Duffield TF, Widowski TM, Lindinger MI, McBride BW. 2002. Effects of a subacute ruminal acidosis model on the diet selection of dairy cows. Journal of Dairy Science 85, 3304–3313. [DOI] [PubMed] [Google Scholar]
- Khafipour E, Krause DO, Plaizier JC. 2009a. A grain‐based subacute ruminal acidosis challenge causes translocation of lipopolysaccharide and triggers inflammation. Journal of Dairy Science 92, 1060–1070. [DOI] [PubMed] [Google Scholar]
- Khafipour E, Krause DO, Plaizier JC. 2009b. Alfalfa pellet‐induced subacute ruminal acidosis in dairy cows increases bacterial endotoxin in the rumen without causing inflammation. Journal of Dairy Science 92, 1712–1724. [DOI] [PubMed] [Google Scholar]
- Kimura K, Sato S, Goto H, Yamagishi N, Okada K, Mizuguchi H, Ito K. 2012a. Simultaneous Estimation of the pH of Rumen and Reticulum Fluids of Cows using a Radio‐Transmission pH‐Measurement System. Journal of Veterinary Medical Science 74, 531–535. [DOI] [PubMed] [Google Scholar]
- Kimura A, Sato S, Kato T, Ikuta K, Yamagishi N, Okada K, et al. 2012b. Relationship between pH and temperature in the ruminal fluid of cows, based on a radio‐transmission pH‐measurement system. Journal of Veterinary Medical Science 74, 1023–1028. [DOI] [PubMed] [Google Scholar]
- Kleen JL, Hooijer GA, Rehage J, Noordhuizen JP. 2003. Subacute ruminal acidosis (SARA): A Review. Journal of Veterinary Medical Science 50, 406–414. [DOI] [PubMed] [Google Scholar]
- Krause KM, Oetzel GR. 2005. Inducing subacute ruminla acidosis in lactating dairy cows. Journal of Dairy Science 88, 3633–3639. [DOI] [PubMed] [Google Scholar]
- Krehbiel CR, Britton RA, Harmon DL, Wester TJ, Stock RA. 1995. The effects of ruminal acidosis on volatile fatty acid absorption and plasma activities of pancreatic enzymes in lambs. Journal of Animal Science 73, 3111–3121. [DOI] [PubMed] [Google Scholar]
- Lane GT, Noller CH, Colenbrander VF, Cummings KR, Harrington RB. 1968. Apparatus for obtaining ruminoreticular samples and the effect of sampling location on pH and volatile fatty acids. Journal of Dairy Science 51, 114–116. [DOI] [PubMed] [Google Scholar]
- Lehloenya KV, Krehbiel CR, Mertz KJ, Rehberger TG, Spicer LJ. 2008. Effects of propionibacteria and yeast culture fed to steers on nutrient intake and site and extent of digestion. Journal of Dairy Science 91, 653–662. [DOI] [PubMed] [Google Scholar]
- Mao SY, Zhang RY, Wang DS, Zhu WY. 2013. Impact of subacute ruminal acidosis (SARA) adaptation on rumen microbiota in dairy cattle using pyrosequencing. Anaerobe 24, 12–19. [DOI] [PubMed] [Google Scholar]
- Marden JP, Bayourthe C, Enjalbert F, Moncoulon R. 2005. A new device for measuring kinetics of ruminal pH and redox potential in dairy cattle. Journal of Dairy Science 88, 277–281. [DOI] [PubMed] [Google Scholar]
- Nagaraja TG, Titgemeyer EC. 2007. Ruminal acidosis in beef cattle: The current microbiological and nutritional outlook. Journal of Dairy Science 90 (E‐Suppl), E17–E38. [DOI] [PubMed] [Google Scholar]
- Nocek JE, Allman JG, Kautz WP. 2002a. Evaluation of an indwelling ruminal probe methodology and effect of grain level on diurnal pH variation in dairy cattle. Journal of Dairy Science 85, 422–428. [DOI] [PubMed] [Google Scholar]
- Nocek JE, Kautz WP, Leedle JAZ, Allman JG. 2002b. Ruminal supplementation of direct‐fed microbials on diurnal pH variation and in situ digestion in dairy cattle. Journal of Dairy Science 95, 5985–5995. [DOI] [PubMed] [Google Scholar]
- Penner GB, Beauchemin KA, Mutsvangwa T. 2006. An evaluation of the accuracy and precision of a stand‐alone submersible continuous ruminal pH measurement system. Journal of Dairy Science 89, 2132–2140. [DOI] [PubMed] [Google Scholar]
- Penner GB, Aschenbach JR, Gäbel G, Oba M. 2009. Technical note:Evaluation of a continuous ruminal pH measurement system for use in noncannulated small ruminants. Journal of Animal Science 87, 2363–2366. [DOI] [PubMed] [Google Scholar]
- Plaizier JC, Krause DO, Gozho GN, McBride BW. 2009. Subacute ruminal acidosis in dairy cows: the physiological causes, incidence and consequences. Veterinary Journal 176, 21–31. [DOI] [PubMed] [Google Scholar]
- Qadis AQ, Goya S, Ikuta K, Yatsu M, Kimura A, Nakanishi S, Sato S. 2014a. Effects of a bacteria‐based probiotic on ruminal pH, volatile fatty acids and bacterial flora of Holstein calves. Journal of Veterinary Medical Science 76, 877–885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qadis AQ, Goya S, Yatsu M, Yoshida Y, Ichijo T, Sato S. 2014b. Effects of a bacteria‐based probiotic on subpopulations of peripheral leukocytes and their interleukin mRNA expression in calves. Journal of Veterinary Medical Science 76, 189–195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qadis AQ, Goya S, Yatsu M, Yoshida Y, Ichijo T, Sato S. 2014c. Immune‐stimulatory effects of a bacteria‐based probiotic on peripheral leukocyte subpopulations and cytokine mRNA expression levels in scouring Holstein calves. Journal of Veterinary Medical Science 76, 677–684. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rice WC, Galyean ML, Cox SB, Dowd SE, Cole NA. 2012. Influence of wet distillers grains diets on beef cattle fecal bacterial community structure. BMC Microbiology 12, 25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rose‐Dye TK, Burciaga‐Robles LO, Krehbiel CR, Step DL, Fulton RW, Confer AW, Richards CJ. 2011. Rumen temperature change monitored with remote rumen temperature boluses after challenges with bovine viral diarrhea virus and Mannheimia haemolytica . Journal of Animal Science 89, 1193–1200. [DOI] [PubMed] [Google Scholar]
- Rustome B, AlZahal O, Cant JP, Fan MZ, Duffield TF, Odongo NE, McBride BW. 2006. Acidogenic value of feeds. II. Effects of rumen acid load from feeds on dry matter intake, ruminal pH, fiber degradability and milk production in the lactating dairy cow. Canadian Journal of Animal Science 86, 119–126. [Google Scholar]
- Sato S, Ikeda A, Tsuchiya Y, Ikuta K, Murayama I, Kanehira M, et al. 2012a. Diagnosis of subacute ruminal acidosis (SARA) by continuous reticular pH measurements in cows. Veterinary Research Communication 36, 85–89. [DOI] [PubMed] [Google Scholar]
- Sato S, Kimura A, Anan T, Yamagishi N, Okada K, Mizuguchi H, Ito K. 2012b. A radio transmission pH measurement system for continuous evaluation of fluid pH in the rumen of cows. Veterinary Research Communication 36, 85–89. [DOI] [PubMed] [Google Scholar]
- Sato S, Mizuguchi H, Ito K, Ikuta K, Kimura A, Okada K. 2012c. Development and testing of a radio transmission pH measurement system for continuous monitoring of ruminal pH in cows. Preventive Veterinary Medicine 103, 274–279. [DOI] [PubMed] [Google Scholar]
- Sauer FD, Kramer JKG, Cantwell WJ. 1989. Antiketogenic effects of monensin in early lactation. Journal of Dairy Science 72, 436–442. [DOI] [PubMed] [Google Scholar]
- Stein DR, Allen DT, Perry EB, Bruner JC, Gates KW, Rehberger TG. 2006. Effects of feeding Propionibacteria to dairy cows on milk yield, milk components, and reproduction. Journal of Dairy Science 89, 111–125. [DOI] [PubMed] [Google Scholar]
- Tafaj M, Junck B, Maulbetsch A, Steingass H, Piepho HP, Drochner W. 2004. Digesta characteristics of dorsal, middle and ventral rumen of cows fed with different hay qualities and concentrate levels. Archives of Animal Nutrition 58, 325–342. [DOI] [PubMed] [Google Scholar]
- Weiss WP, Wyatt DL, McKelvey TR. 2008. Effects of feeding Propionibacteria on milk production by early lactation dairy cows. Journal of Dairy Science 91, 646–652. [DOI] [PubMed] [Google Scholar]