

ABSTRACT
The spike proteins of coronaviruses are capable of binding to a wide range of cellular targets, which contributes to the broad species tropism of coronaviruses. Previous reports have demonstrated that Middle East respiratory syndrome coronavirus (MERS-CoV) predominantly utilizes dipeptidyl peptidase 4 (DPP4) for cell entry. However, additional cellular binding targets of the MERS-CoV spike protein that may augment MERS-CoV infection have not been further explored. In the current study, using the virus overlay protein binding assay (VOPBA), we identified carcinoembryonic antigen-related cell adhesion molecule 5 (CEACAM5) as a novel cell surface binding target of MERS-CoV. CEACAM5 coimmunoprecipitated with the spike protein of MERS-CoV in both overexpressed and endogenous settings. Disrupting the interaction between CEACAM5 and MERS-CoV spike with anti-CEACAM5 antibody, recombinant CEACAM5 protein, or small interfering RNA (siRNA) knockdown of CEACAM5 significantly inhibited the entry of MERS-CoV. Recombinant expression of CEACAM5 did not render nonpermissive baby hamster kidney (BHK21) cells susceptible to MERS-CoV infection. Instead, CEACAM5 overexpression significantly enhanced the attachment of MERS-CoV to the BHK21 cells. More importantly, the entry of MERS-CoV was increased when CEACAM5 was overexpressed in permissive cells, which suggested that CEACAM5 could facilitate MERS-CoV entry in conjunction with DPP4 despite not being able to support MERS-CoV entry independently. Taken together, the results of our study identified CEACAM5 as a novel cell surface binding target of MERS-CoV that facilitates MERS-CoV infection by augmenting the attachment of the virus to the host cell surface.
IMPORTANCE Infection with the Middle East respiratory syndrome coronavirus (MERS-CoV) is associated with the highest mortality rate among all known human-pathogenic coronaviruses. Currently, there are no approved vaccines or therapeutics against MERS-CoV infection. The identification of carcinoembryonic antigen-related cell adhesion molecule 5 (CEACAM5) as a novel cell surface binding target of MERS-CoV advanced our knowledge on the cell binding biology of MERS-CoV. Importantly, CEACAM5 could potentiate the entry of MERS-CoV by functioning as an attachment factor. In this regard, CEACAM5 could serve as a novel target, in addition to dipeptidyl peptidase-4 (DPP4), in the development of antiviral strategies for MERS-CoV.
INTRODUCTION
Coronaviruses are enveloped, positive-sense, single-stranded RNA viruses with genome sizes of approximately 30 kb. They belong to the family Coronaviridae in the order Nidovirales and are currently classified into four major genera, Alphacoronavirus, Betacoronavirus, Gammacoronavirus, and Deltacoronavirus (1). Coronaviruses can infect a wide range of mammals, as well as birds (2). The broad species tropism is predominantly attributed to the high diversity in receptor usage across different coronaviruses. To date, six coronaviruses are known to infect humans, and they utilize different surface molecules for cell entry. In particular, human coronavirus 229E (HCoV-229E) binds aminopeptidase N (APN) (3), and human coronavirus OC43 (HCoV-OC43) binds O-acetylated sialic acid (4). Severe acute respiratory syndrome coronavirus (SARS-CoV) (5) and human coronavirus NL63 (HCoV-NL63) (6) both bind angiotensin I converting enzyme 2 (ACE2). The receptor for human coronavirus HKU1 (HCoV-HKU1) has not been defined. However, O-acetylated sialic acid has been suggested as an attachment factor that contributes to the binding of HCoV-HKU1 to the cell surface (7). Middle East respiratory syndrome coronavirus (MERS-CoV) is the sixth coronavirus known to cause infection in humans (8). Intriguingly, MERS-CoV utilizes a unique cellular receptor, dipeptidyl peptidase 4 (DPP4), for virus entry (9). The host cell receptors for a number of animal coronaviruses have also been identified. For instance, porcine transmissible gastroenteritis coronavirus (TGEV) binds APN (10), and the prototype betacoronavirus mouse hepatitis virus (MHV) uses carcinoembryonic antigen-related cell adhesion molecule 1 (CEACAM1) for entry (11).
Coronaviruses have evolved complex receptor recognition patterns. In addition to the defined receptors essential for virus entry into host cells, multiple coreceptors or attachment factors have been reported to play critical roles in the propagation of coronaviruses. In this regard, sialic acids (12) and glycoproteins (13) facilitate the binding of TGEV to target cells, in addition to APN. Similarly, HCoV-NL63 utilizes heparan sulfate proteoglycans for attachment to target cells (14). Apart from ACE2, SARS-CoV can also enter cells through liver/lymph node-specific intercellular adhesion molecule-3-grabbing integrin (L-SIGN) (15) and dendritic cell-specific intercellular adhesion molecule 3-grabbing nonintegrin (DC-SIGN) (16). Additionally, we have previously identified major histocompatibility complex class I C (HLA-C) as an attachment factor for HCoV-HKU1 that facilitates the entry of the virus (17).
The emerging MERS-CoV is associated with the highest mortality rate (more than 30%) in infected patients among all known human-pathogenic coronaviruses, and there is as yet no approved treatment regimen or vaccine for MERS (18, 19). As of 16 May 2016, MERS-CoV had caused 1,733 laboratory-confirmed cases of human infection, including at least 628 deaths (20). Clinical features of severe MERS include high fever, pneumonia, and acute respiratory distress syndrome (ARDS), as well as extrapulmonary manifestations, including gastrointestinal symptoms, lymphopenia, acute kidney injury, hepatic inflammation, and pericarditis (21). In agreement with these clinical observations, recent in vitro and in vivo studies have highlighted the extraordinarily wide range of tissue and cell tropism of MERS-CoV, which is unparalleled by other coronaviruses (22–24). Following the identification of DPP4 as the receptor of MERS-CoV, the broad tissue tropism of MERS-CoV infection was in part explained by the ubiquitous cellular expression of DPP4. However, alternative factors may exist and potentiate the infection of MERS-CoV either in conjunction with or independently of DPP4. In this study, we employed the virus overlay protein binding assay (VOPBA), followed by the liquid chromatography-tandem mass spectrometry (LC–MS-MS) approach, to identify novel cell surface binding targets of MERS-CoV. Our data demonstrated carcinoembryonic antigen-related cell adhesion molecule 5 (CEACAM5) to be a novel attachment factor that facilitated MERS-CoV entry. Importantly, interrupting the interaction between CEACAM5 and MERS-CoV spike (S) protein inhibited virus entry. Overexpression of CEACAM5 did not allow entry but significantly increased the attachment of MERS-CoV on nonpermissive cells. Collectively, our study identified CEACAM5 as an important cell surface binding target for MERS-CoV spike that facilitates host cell entry for MERS-CoV.
MATERIALS AND METHODS
Cells.
A549, AD293, Huh7, Caco2, and VeroE6 cells were maintained in Dulbecco's modified Eagle medium (DMEM) supplemented with 10% heat-inactivated fetal bovine serum (FBS), 100 U/ml penicillin, and 100 μg/ml streptomycin. BEAS2B and Calu3 cells were maintained in DMEM/F-12 supplemented with 10% heat-inactivated FBS, 100 U/ml penicillin, and 100 μg/ml streptomycin. BHK21 cells were maintained in minimum essential medium (MEM) supplemented with 10% heat-inactivated FBS, 100 U/ml penicillin, and 100 μg/ml streptomycin. L929 cells were maintained in MEM supplemented with 20% heat-inactivated FBS, 100 U/ml penicillin, and 100 μg/ml streptomycin. Human primary T cells were isolated from peripheral blood mononuclear cells (PBMCs) by negative selection using the Dynabeads Untouched Human T cells kit (Invitrogen, USA), as we previously described (25). The isolated T cells were maintained in RPMI 1640 supplemented with 10% FBS, 100 μg/ml streptomycin, 100 U/ml penicillin, 1% sodium pyruvate, and 1% nonessential amino acids and were used immediately for infection.
Virus.
The EMC/2012 strain of MERS-CoV (passage 8; referred to as MERS-CoV) was provided by Ron Fouchier (Erasmus Medical Center) and cultured with VeroE6 cells in serum-free DMEM supplemented with 100 U/ml penicillin and 100 μg/ml streptomycin. Three days after virus inoculation, the culture supernatants were collected, aliquoted, and stored at −80°C. To determine the virus titer, aliquots of MERS-CoV were used for plaque assays on confluent VeroE6 cells in 24-well plates. In brief, MERS-CoV stocks were 10-fold serially diluted in DMEM. The diluted MERS-CoV was then added to duplicate wells of 24-well plates at a volume of 200 μl. After inoculation for 1 h at 37°C, the inoculum was removed from the wells, and an agarose overlay was added to the cells. The cells were then incubated for approximately 72 h. To count plaques, the cells were fixed with 4% paraformaldehyde for 5 h and stained with crystal violet.
Antibodies.
Rabbit anti-human DPP4, rabbit anti-human CEACAM5, rabbit control IgG (Abcam, USA), and mouse anti-human β-actin (Immunoway, USA) were used in the relevant experiments. MERS-CoV NP was detected with guinea pig anti-MERS-CoV NP serum as we previously described (25). MERS-CoV spike was detected either with the in-house mouse anti-MERS-CoV spike immune serum or with a rabbit anti-MERS-CoV spike antibody (Sino Biological, China). Secondary antibodies, including Alexa Fluor 488/647 goat anti-guinea pig and Alexa Fluor 488/647 goat anti-rabbit (Life Technologies, USA), were used for flow cytometry. Goat anti-mouse horseradish peroxidase (HRP) (Abcam, USA) was used for Western blotting.
Plasmid construction.
The synthetic human codon-optimized S gene was used as a template for the construction of all MERS-CoV S plasmids. For pcDNA-MERS-CoV S used in producing MERS-S-pseudotyped virus, the full-length S from the N-terminal KpnI site to the C-terminal XhoI site was subcloned to generate full-length S in pcDNA 3.1(+). For the construction of the S1 plasmid (amino acids 1 to 925), the 5′ forward primer sequence containing a BSSHII site (5′GCGCGCCACCATGATACACTCAGTGTTTCTACTGATGTTC) was used, together with the 3′ reverse primer (5′-GGGCCCATCACCGTCTTCCCACACAGTGGATG) with an ApaI site. The fragment was PCR amplified and cloned into the pSFV1 vector (kindly provided by P. Liljestrom) with the C terminus fused in frame with the FLAG sequence (DYKDDDDK), resulting in the plasmid pSFV-MERS-CoV S1-FLAG. For the construction of the pSFV-DPP4-6×His plasmid, a forward primer containing a BSSHII site (5′-GCGCGCCACCATGAAGACACCGTGGAAGG) and a reverse primer with an ApaI site (5′-GGGCCCAGGTAAAGAGAAACATTGTTTTATG) were used. The fragment was cloned into the pSFV1 vector with the 6×His sequence (HHHHHH) fused in frame at the C terminus. For the construction of pcDNA-DPP4, the forward primer containing a KpnI site (5′-TAAGCAGGTACCGCCACCATGAAGACACCGTGGAAGGTTCT) and the reverse primer containing a EcoRI site (5′-TGCTTAGAATTCCTAAGGTAAAGAGAAACATTGTTTTATGAAGTGGC) were used to insert the full-length human DPP4 into the pcDNA3.1(+) vector between the KpnI and EcoRI sites. For the construction of pcDNA-CEACAM5-V5, the forward primer (5′GATATCCACCATGGAGTCTCCCTCGGCCCCTCCCCAC) contained the N-terminal signal sequence, the EcoRV site, and the Kozak sequence. The reverse primer (5′-CTCGAGTATCAGAGCAACCCCAACCAGCACTCC) contained the XhoI site in the C-terminal cytoplasmic domain. The amplified fragment was used to generate full-length CEACAM5 in pcDNA 3.1(+) with a V5 tag (GKPIPNPLLGLDST) in frame at the C terminus, resulting in pcDNA-CEACAM5-V5.
Production of MERS-S-pseudotyped virus.
Lentivirus-based MERS-S-pseudotyped virus was generated by cotransfecting 293FT cells with pcDNA-MERS full-length spike in combination with pNL4-3-R−.E−, which is an HIV backbone plasmid bearing the luciferase reporter gene. pNL4-3-R−.E− was obtained through the NIH AIDS Research and Reference Reagents program. The viral particles in the supernatant were harvested 48 h posttransfection by ultracentrifugation in 30% sucrose solution in a Beckman SW32Ti rotor at 32,000 rpm for 1 h at 4°C (17). The p24 concentrations were quantified with a p24 antigen enzyme-linked immunoassay kit (Cell Biolabs, USA) and stored in aliquots at −80°C. The pseudovirus titer was quantified in lentiviral particles (LP) per milliliter according to the manufacturer's instructions.
VOPBA and Western blotting.
Confluent A549 cells in 75-cm2 tissue flasks were washed three times in chilled phosphate-buffered saline (PBS), surface biotinylated, and extracted using a Pierce Cell Surface Protein Isolation kit (Thermo Scientific, USA) according to the manufacturer's protocol. Biotinylated membrane extracts were bound onto neutrAvidin agarose resin (Thermo Scientific, USA), washed, and eluted in LDS sample buffer (Invitrogen, USA). After electrophoresis in a 4 to 12% gradient NuPAGE SDS-PAGE gel (Invitrogen, USA) under reducing conditions, the biotinylated surface extracts were electrotransferred onto Hybond polyvinylidene difluoride (PVDF) membranes (GE Healthcare, UK) for 2 h at a constant voltage of 30 V at 4°C. The membranes were blocked in 10% skim milk (Oxoid, UK) in PBS at 4°C overnight on a rotator. The membranes were then incubated with 1% skim milk solution containing MERS-S-pseudotyped virus at 109 lentiviral particles/ml for 2 h. The membranes were washed twice in 10% skim milk and once in PBS and incubated with rabbit antiserum raised against MERS-CoV spike protein at 1 μg/ml for 2 h at room temperature. After incubation with the spike-specific antibody, we washed the membranes twice in 10% skim milk and once in PBS and incubated them with PBS containing 1:5,000 goat anti-rabbit 800 infrared dye (Li-Cor, USA). The membranes were scanned with an Odyssey Imaging System (Li-Cor, USA). The visualized reactive protein detected by VOPBA on a Western blot was excised from the gel. The gel piece was inserted into dialysis tubing and filled with 1× MOPS (morpholinepropanesulfonic acid) SDS running buffer. The protein contents were electroeluted at a constant voltage of 50 V for 6 h, collected, dialyzed overnight at 4°C against PBS in a Slide-A-Lyzer Cassette (Thermo Scientific, USA), and concentrated using Microcon columns (Millipore, USA). The eluted contents were electrophoresed and confirmed by VOPBA. The gel fragment was excised for LC–MS-MS analysis, carried out in the Center for Genomic Sciences at the University of Hong Kong, as we previously described (26).
Co-IP.
Coimmunoprecipitation (co-IP) was performed using anti-FLAG M2 affinity gel beads (Sigma, USA) or V5 antibody preadsorbed to protein A+G Sepharose (Thermo Scientific, USA). Cell lysates were harvested in lysis buffer (20 mM Tris-HCl [pH 7.4], 150 mM NaCl, 1% Triton X-100, 0.1% NP-40, protease and phosphatase inhibitor cocktails [Roche]). The lysed cells were incubated on ice for 15 to 30 min and then centrifuged. The supernatant was added to antibody-coupled beads or anti-FLAG affinity gel beads and incubated overnight at 4°C. The beads were washed three times with lysis buffer, and Western blotting was performed. The immunoprecipitated products or cell lysates were mixed with SDS loading buffer and heated at 95°C for 10 min. Samples were fractionated in SDS-10% PAGE and then transferred onto PVDF membranes (GE Healthcare, UK). After blocking overnight with 10% skim milk, the membranes were incubated with the corresponding primary antibodies and then with infrared-dye-conjugated secondary antibodies (Li-Cor, USA). The membranes were scanned with an Odyssey Imaging System (Li-Cor, USA).
Quantitative RT-PCR.
Cells were lysed in RLT buffer with 40 mM dithiothreitol (DTT) and extracted with an RNeasy minikit (Qiagen, USA). Viral RNA in the supernatant was extracted with a PureLink Viral RNA/DNA minikit (Life Technologies, USA). Reverse transcription (RT) and quantitative PCR (qPCR) were performed with the Transcriptor First Strand cDNA synthesis kit and LightCycler 480 master mix from Roche, as we previously described (27, 28). In the RT reactions, reverse primers against the NP gene of MERS-CoV were used to detect cDNA complementary to the positive strands of viral genomes. The following sets of primers were used to detect NP in qPCR: F, 5′-CAAAACCTTCCCTAAGAAGGAAAAG-3′; R, 5′-GCTCCTTTGGAGGTTCAGACAT-3′; and probe, 6-carboxyfluorescein (FAM) 5′-ACAAAAGGCACCAAAAGAAGAATCAACAGACC-3′ black hole quencher 1 (BHQ1).
Antibody blocking assays.
The antibody blocking assay based on the MERS-S-pseudotyped virus was performed by measuring the infection of target cells with the use of luciferase as a reporter gene (17). In brief, the antibody was diluted to 2 μg/ml in serum-free culture medium and incubated with Calu3 cells for 1 h at 37°C. Pseudotyped virus was added to the target cells at a ratio of 100 LP per cell, and the inoculum was replaced with fresh medium with 10% FBS after 1 h. Luciferase activity was determined at 48 h postinfection. For the antibody blocking assay based on the infectious MERS-CoV, Calu3 cells were preincubated with rabbit polyclonal anti-CEACAM5 at different concentrations ranging from 0.5 to 4 μg/ml. Rabbit polyclonal anti-DPP4 at 4 μg/ml and rabbit IgG at 4 μg/ml were included as controls. After preincubation with the antibodies for 1 h at 37°C, the cells were challenged with MERS-CoV at a multiplicity of infection (MOI) of 1 for 1 h at 37°C in the presence of antibodies. The cells were subsequently washed with PBS and lysed with RLT buffer with 40 mM DTT. To determine the impact of antibody blocking on MERS-CoV multicycle replication, Huh7 cells were preincubated with 2 μg/ml anti-CEACAM5 antibody, anti-DPP4 antibody, or rabbit IgG. After the preincubation, the cells were infected with MERS-CoV at an MOI of 0.0005 for 1 h at 37°C in the presence of antibodies. The cells were then washed in PBS and replenished with DMEM with blocking antibodies. The infected cells were incubated at 37°C and harvested at 1, 24, and 48 h postinfection. The virus copy number was determined by qPCR.
Human recombinant protein blocking assays.
Human recombinant proteins (DPP4, CEACAM5, and control IgG) were obtained from Sino Biological. MERS-CoV was preincubated with human recombinant proteins (5 μg/ml to 40 μg/ml for CEACAM5, 40 μg/ml for DPP4, and 40 μg/ml for control IgG) for 1 h before inoculation onto Calu3 and Huh7 cells. The protein-virus complexes were then incubated with the Calu3 and Huh7 cells for 2 h at 37°C. After the incubation period, the inoculum was discarded and the cells were washed with PBS, followed by lysing in RLT with 40 mM DTT. The virus copy number was determined by qPCR.
siRNA knockdown.
Silencer Select siRNA human CEACAM5 small interfering RNA (siRNA), Silencer Select siRNA human DPP4 siRNA, and Silencer Select siRNA negative control were obtained from Life Technologies. Transfection of siRNA on Huh7 cells was performed using Lipofectamine 3000 (Thermo Fisher, USA) following the manufacturer's manual. In brief, Huh7 cells were transfected with 100 nM siRNA for two consecutive days. At 24 h after the second siRNA transfection, the cells were challenged with MERS-CoV at an MOI of 1 for 2 h at 37°C. Following the incubation, the cells were washed with PBS and lysed in RLT buffer with 40 mM DTT. The virus copy number was determined by qPCR.
Flow cytometry.
All samples were detached with 10 mM EDTA and fixed in 4% paraformaldehyde. Cell permeabilization for intracellular staining was performed with 0.1% Triton X-100 in PBS. Immunostaining for flow cytometry was performed following standard procedures as we previously described (29). The flow cytometry was performed using a BD FACSCanto II flow cytometer (BD Biosciences, USA), and the data were analyzed using FlowJo vX (Tree Star, USA).
Flow cytometry of BHK21 cells with overexpression of DPP4 and CEACAM5.
BHK21 cells were transfected with pSFV-DPP4 or pcDNA-CEACAM5. The transfected cells were inoculated with MERS-CoV at 48 h posttransfection. To determine virus entry, the cells were inoculated with MERS-CoV at an MOI of 5 at 37°C for 2 h. After 2 h, the cells were washed with PBS and incubated for another 4 h. At 6 h postinfection, the cells were washed extensively with PBS, fixed in 4% paraformaldehyde, and immunolabeled for flow cytometry. To determine virus attachment, the cells were inoculated with MERS-CoV at an MOI of 30 at 4°C for 2 h. After 2 h, the cells were washed with PBS, fixed in 4% paraformaldehyde, and immunolabeled for flow cytometry.
Confocal microscopy.
This study was approved by the Institutional Review Board of the University of Hong Kong/Hospital Authority Hong Kong West Cluster. Normal human lung sections were deparaffinized and rehydrated following standard procedures. Antigen unmasking was performed by boiling tissue sections with the antigen-unmasking solution from Vector Laboratories. Goat anti-DPP4 was obtained from R&D, and rabbit anti-CEACAM5 was obtained from Abcam. Secondary antibodies were obtained from Life Technologies. Mounting and DAPI (4′,6-diamidino-2-phenylindole) staining were performed with Vectashield mounting medium (Vector Laboratories, USA). Images were acquired with a Carl Zeiss LSM 780 system.
Statistical analysis.
Data are presented as means and standard deviations. Statistical comparison between different groups was performed by Student's t test using GraphPad Prism 6. Differences were considered statistically significant when the P value was <0.05.
RESULTS
Identification of CEACAM5 as a cell surface binding protein of MERS-CoV.
To probe for potential cell surface factors that can interact with MERS-CoV, we employed the VOPBA, which we have previously utilized to identify host cell surface binding proteins of influenza A virus (26). In brief, membrane proteins on A549 cells were selectively biotinylated and separated from nonbiotinylated intracellular proteins by binding to avidin agarose (23). The biotinylated A549 surface extracts separated in a 4 to 12% gradient SDS NuPAGE gel were transferred to PVDF membranes before incubation with MERS-S-pseudotyped viruses for 2 h at room temperature (Fig. 1A). Subsequently, reactive signals were revealed with a rabbit polyclonal antibody against MERS-CoV spike. As shown in Fig. 1B, VOPBA revealed several reactive bands from A549 surface extracts, including two prominent bands at molecular masses of approximately 120 and 90 kDa (lane 1 from left) but not from the control NIH 3T3 cell (lane 2), which is not permissive for MERS-CoV infection (30). In order to confirm the immunoreactivity of these two proteins to MERS-S-pseudotyped virus, the relevant fractions from the SDS-PAGE gel were excised, electroeluted, and hybridized with MERS-S-pseudotyped virus. Although both proteins reacted with the virus, the protein at around 90 kDa returned a much stronger signal and was selected for subsequent analyses (Fig. 1C). Tryptic peptide mass fingerprints derived from the excised band revealed the protein to be human CEACAM5, using the MASCOT search engine against the NCBI and Swiss-Prot databases (Fig. 1D).
FIG 1.

CEACAM5 is a cell surface binding protein of MERS-CoV. (A) A549 and NIH 3T3 biotinylated cell membrane proteins were extracted, separated by 4 to 12% gradient gel, and transferred to PVDF membranes. WB, Western blotting. (B) Membrane proteins were probed with MERS-S-pseudotyped virus followed by detection of virus binding by incubation with immune serum directed against the MERS-CoV spike protein (left lane). Nonsusceptible cell membrane extracts from NIH 3T3 was included as a negative control (right lane). (C) The gel fractions corresponding to the positive signal at approximately 90 kDa were cut, electroeluted, and confirmed by VOPBA prior to submission for MS protein identification. (B and C) The position of CEACAM5 is indicated with asterisks. (D) Identified amino acid sequences and their corresponding positions.
CEACAM5 is expressed on cell types that are highly susceptible to MERS-CoV.
Most CEACAMs are considered modulators of general cellular processes, including proliferation, motility, apoptosis, and attachment, as well as cell-cell interaction. In addition, CEACAM5 may also play roles in innate immune defense by binding and trapping microorganisms (31). To verify the relevance of CEACAM5 in MERS-CoV entry, we investigated the expression of CEACAM5 on a number of cell types. To this end, different cell types in 6-well plates were detached with EDTA, washed, and fixed in 4% paraformaldehyde. Immunostaining for surface CEACAM5 and DPP4 expression was performed without cell permeabilization. The percentage of positive cells and the surface mean fluorescent intensities (MFIs) were determined by flow cytometry. In agreement with previously published reports, DPP4 was detected ubiquitously on different human cell lines (Fig. 2A), as well as human primary T cells (Fig. 2B). In addition, the polyclonal antibody against human DPP4 also cross-reacted with DPP4 on a number of nonhuman mammalian cells, which included VeroE6 (monkey), BHK21 (hamster), and L929 (mouse) cells (Fig. 2C). Notably, despite being recognized by the human DPP4 antibody, the DPP4 on hamster and mouse cells did not support MERS-CoV entry (30). In contrast to DPP4, our data demonstrated that the expression of CEACAM5 was more restricted. Among the six human cell lines we analyzed, CEACAM5 expression was detected on A549, Calu3, and Huh7 cells (Fig. 2A). At the same time, little to no CEACAM5 expression was detected on AD293, BEAS2B, and Caco2 cells (Fig. 2A). Importantly, CEACAM5 expression was also detected on human primary T cells (Fig. 2B). However, no human CEACAM5 ortholog was expressed on VeroE6, BHK21, and L929 cells (Fig. 2C). Quantitatively, CEACAM5 expression was detected predominantly on cells that are highly susceptible to MERS-CoV infection, which include Calu3, Huh7, A549, and T cells (Fig. 2D and E). Thus, our data supported the notion that CEACAM5 might be an important surface binding protein that could facilitate MERS-CoV entry.
FIG 2.

Surface expression of CEACAM5 on mammalian cells. (A) The indicated human cell lines were fixed in 4% paraformaldehyde and immunolabeled for surface DPP4 and CEACAM5 expression. The shaded curves and the open curves represent isotype- and antigen-specific staining, respectively. Horizontal lines represent the gating for DPP4+ or CEACAM5+ cells. The numbers represent the percentage of positive cells. (B and C) The same fixation and immunostaining procedures were performed for human primary T cells (B) and a number of nonhuman mammalian cells (C). (D) Average percentages of DPP4/CEACAM5-positive cells. (E) Average DPP4/CEACAM5 MFIs were quantified. Rabbit isotype IgG was used in place of antigen-specific antibodies for the controls. (D and E) Shown are means and standard deviations of the results of three independent experiments.
CEACAM5 is coexpressed with DPP4 in human lung tissues.
To further verify the physiological relevance of CEACAM5 in MERS-CoV infection, we evaluated the expression of CEACAM5 in human lung tissues and compared the expression with that of DPP4 by immunofluorescence microscopy. Our result revealed that CEACAM5 could be readily detected in the epithelial cells in various regions of the human lung tissues (Fig. 3). In particular, colocalization between CEACAM5 and DPP4 was observed in the bronchial epithelium (Fig. 3A) but appeared to be more extensive in the epithelium of small airways (Fig. 3B). The colocalization between CEACAM5 and DPP4 was also detected in alveoli (Fig. 3C), including the alveolar macrophages (Fig. 3C, arrowheads). Collectively, the detection of CEACAM5 in the epithelial cells of the human lung tissues and its coexpression with DPP4 in these samples demonstrated the potential capacity of CEACAM5 to facilitate MERS-CoV infection in the lower respiratory tract.
FIG 3.

Expression of CEACAM5 in human lung tissues. Immunostaining of CEACAM5 and DPP4 was performed on paraffin slides of normal human lung tissues. Representative images of CEACAM5 and DPP4 expression in various regions of the human lung are shown, including the bronchi (A), small airways (B), and alveoli (C). The arrowheads indicate alveolar macrophages with CEACAM5 and DPP4 coexpression. Bars, 50 μm.
CEACAM5 interacts with MERS-CoV spike.
To validate our VOPBA-LC-MS/MS findings, the direct interaction between MERS-CoV S1 and CEACAM5 was characterized by co-IP. BHK21 cells were transfected with either a CEACAM5-V5 expression vector or a control vector tagged with V5. The surface and intracellular expression of transfected CEACAM-V5 was verified by flow cytometry (Fig. 4A). The transfected BHK21 cells were then immunoprecipitated with either MERS-CoV S1-FLAG or Escherichia coli bacterial alkaline phosphatase (BAP)-FLAG preadsorbed onto anti-FLAG M2 agarose beads. The bound protein complexes were spun down, washed, and subjected to Western blot detection with anti-FLAG (Fig. 4B, top) or the anti-V5 antibody (Fig. 4B, bottom). As demonstrated in Fig. 4B (bottom, lane 1 from left), CEACAM5 bound specifically to MERS-CoV S1. In contrast, CEACAM5 did not bind to the control bait protein (Fig. 4B, bottom, lane 2). At the same time, CEACAM5 was not detected in samples transfected with the empty vector (Fig. 4B, bottom, lane 3). In parallel, we performed reciprocal co-IP experiments in which CEACAM5-V5 was used as the bait instead of MERS-CoV S1-FLAG (Fig. 4C). To this end, CEACAM5-V5- or empty-vector-transfected BHK21 cell lysates were immunoprecipitated with anti-V5 preadsorbed protein A/G Sepharose, spun down, washed, and incubated with purified MERS-CoV S1-FLAG or BAP-FLAG. Western blot detection with anti-FLAG showed that CEACAM5-V5 interacted with MERS-CoV S1-FLAG (Fig. 4C, top, lane 1 from left), but not BAP-FLAG (Fig. 4C, top, lane 2). As a negative control, the V5 empty vector did not pull down MERS-CoV S1 (Fig. 4C, top, lane 3). In line with the flow cytometry data from Fig. 2C, the antibody against human CEACAM5 did not pick up any signal from the BHK21 cell lysates (Fig. 4C, bottom, lane 3). To further validate our findings and to avoid the use of overexpressed or tagged fusion proteins, we performed endogenous co-IP on nontransfected Huh7 cells infected with MERS-CoV using antibody against CEACAM5 or MERS-CoV spike. As demonstrated in Fig. 4D, MERS-CoV spike was detected in infected samples immunoprecipitated with the anti-CEACAM5 antibody. In contrast, MERS-CoV spike was not detected from uninfected samples or from infected samples immunoprecipitated with nonspecific rabbit IgG. The interaction between MERS-CoV spike and CEACAM5 was confirmed by reciprocal co-IP using antibody against MERS-CoV spike. In this regard, endogenous CEACAM5 was detected from infected samples immunoprecipitated with antibody against MERS-CoV spike but not from uninfected samples or from infected samples immunoprecipitated with nonspecific rabbit IgG (Fig. 4D). Taken together, our co-IP results demonstrated that MERS-CoV S1 could bind specifically to CEACAM5.
FIG 4.

CEACAM 5 interacts with MERS-CoV spike. (A) Surface and intracellular expression of CEACAM5-V5 were verified by flow cytometry. (B) For co-IP, BHK21 cells were transfected with CEACAM5-V5 (lanes 1 and 2 from left) or empty vector (lane 3). The cell lysate was immunoprecipitated with MERS-CoV S1-FLAG (lanes 1 and 3) or E. coli BAP-FLAG protein (lane 2) preadsorbed onto anti-FLAG M2 agarose beads prior to SDS-PAGE. The protein complex was detected by using the anti-FLAG antibody or the anti-V5 antibody. (C) Reciprocal co-IP was performed using CEACAM5-V5 as the bait. Purified MERS-CoV S1-FLAG (lanes 1 and 3) or BAP-FLAG (lane 2) proteins were immunoprecipitated with overexpressed CEACAM5-V5 or pcDNA-V5 protein preadsorbed onto anti-V5 Sepharose beads. Western blots were detected with the anti-FLAG or the anti-CEACAM5 antibody. (D) Endogenous co-IP was performed with MERS-CoV-infected or mock-infected Huh7 cell lysates using the rabbit anti-CEACAM5 antibody, the rabbit anti-MERS-CoV spike antibody, or the rabbit isotype IgG. Western blots were detected with the rabbit anti-MERS-CoV spike antibody or the rabbit anti-CEACAM5 antibody.
CEACAM5-specific antibody blocks MERS-CoV entry and spread.
After confirming the direct interaction between CEACAM5 and MERS-CoV S1, we next investigated the potential functional role of CEACAM5 in MERS-CoV entry. We first assessed the capacity of CEACAM5 antibody to block the entry of MERS-S pseudovirus. In this setting, Calu3 cells were preincubated with polyclonal antibody against human CEACAM5 or with rabbit IgG and polyclonal antibody against DPP4 as negative and positive controls, respectively. After the preincubation, MERS-S pseudoviruses were inoculated onto the cells for another hour in the presence of the CEACAM5 antibody. The inoculum was replaced with fresh medium with 10% FBS after 1 h, and the luciferase activity was determined at 48 h postinfection. As demonstrated in Fig. 5A, polyclonal antibody against human CEACAM5 at 2 μg/ml significantly reduced the entry of MERS-S pseudovirus. Next, the antibody blocking experiment was further validated using infectious MERS-CoV under a titration of different CEACAM5 antibody concentrations ranging from 0.5 μg/ml to 4 μg/ml. Our results demonstrated that CEACAM5 antibody blocked the infection of MERS-CoV in a dose-dependent manner (Fig. 5B). As a control, antibody added after MERS-CoV inoculation did not affect virus entry (Fig. 5C). At the same time, addition of the CEACAM5 antibody did not prevent the entry of MERS-CoV into Caco2 cells, which expressed a low level of CEACAM5 (Fig. 5D). To determine the impact of CEACAM5 inhibition on the multicycle spread of MERS-CoV, we extended the antibody blocking experiment to Huh7 cells. In this scenario, Huh7 cells were preincubated with the CEACAM5 antibody for 1 h and inoculated with MERS-CoV at a low MOI in the presence of the CEACAM5 antibody. The infected cells were then washed and incubated in DMEM culture medium in the presence of the CEACAM5 antibody. Remarkably, our data suggested that the CEACAM5 antibody significantly reduced MERS-CoV propagation in both cell lysates (Fig. 5E) and supernatants (Fig. 5F) of infected Huh7 cells. Overall, we demonstrated that CEACAM5 was important for MERS-CoV entry, as well as in multicycle viral spread.
FIG 5.

CEACAM5-specific antibody blocks MERS-CoV entry and replication. (A) The antibody blocking assay was performed in Calu3 cells using MERS-S-pseudotyped virus. Antibodies were diluted to 2 μg/ml and incubated with Calu3 cells for 1 h at 37°C. The pseudotyped viruses were then added at a ratio of 100 LP per cell for 1 h. Luciferase activity was determined at 48 h postinfection and was normalized to that of the mock-treated cells. (B) The antibody blocking assay was performed in Calu3 cells using MERS-CoV. Calu3 cells were preincubated with antibodies at the indicated concentrations for 1 h at 37°C. The cells were then inoculated with MERS-CoV at an MOI of 1 for 1 h at 37°C in the presence of the antibodies. After 1 h, the cells were washed and harvested. MERS-CoV entry was assessed by qPCR, and the result was normalized to that of the mock-treated cells. (C) Calu3 cells were treated with CEACAM5 antibody for a total of 2 h during preincubation and virus inoculation (−1 to 1) or after virus inoculation (1 to 3). MERS-CoV entry was assessed by qPCR. (D) The antibody blocking assay was performed in Caco2 cells. (E and F) The impact of CEACAM5 inhibition on MERS-CoV replication was investigated in Huh7 cells. Huh7 cells were preincubated with antibodies at 2 μg/ml for 1 h at 37°C. The cells were then inoculated with MERS-CoV at an MOI of 0.0005 for 1 h at 37°C in the presence of the antibodies. At the end of the inoculation period, the inoculum was replaced with culture medium containing the indicated antibodies. Cell lysates (E) and supernatants (F) were harvested at 1, 24, and 48 h postinfection. The virus genome copy number was determined with qPCR. ND, virus was not detected. The data are represented as means and standard deviations of the results of three independent experiments. The results for the anti-DPP4-, anti-CEACAM5-, and control IgG-treated samples were compared with those for the mock-treated samples. Statistical analyses were carried out using Student's t test. Statistical significance is indicated by the asterisks (P < 0.05).
CEACAM5 recombinant protein and siRNA knockdown of CEACAM5 inhibit MERS-CoV entry.
In addition to antibody blocking, we sought to further verify the impact of CEACAM5 inhibition on MERS-CoV infection with alternative approaches, including protein blocking and siRNA knockdown. In protein blocking assays, MERS-CoV was preincubated with human recombinant CEACAM5 protein at concentrations ranging from 5 μg/ml to 40 μg/ml. Human recombinant DPP4 protein and human recombinant IgG were included as controls. Calu3 or Huh7 cells were subsequently inoculated for 2 h with the virus-protein mixtures. After inoculation, the cells were washed and harvested for viral-load quantification with qPCR analysis. Remarkably, our data demonstrated that the human CEACAM5 recombinant protein blocked the entry of MERS-CoV in a dose-dependent manner in both Calu3 (Fig. 6A) and Huh7 (Fig. 6B) cells. As a control, CEACAM5 protein added after MERS-CoV inoculation had no effect on virus entry (Fig. 6C). Next, we depleted the endogenous expression of CEACAM5 in Huh7 cells with siRNA. The reduction of CEACAM5 surface expression was examined by flow cytometry with no cell permeabilization (Fig. 6D). As illustrated in Fig. 6E and F, siRNA treatment significantly diminished the surface expression of CEACAM5 in terms of the percentage of CEACAM5-positive cells (Fig. 6E), as well as the MFI (Fig. 6F). Notably, when the CEACAM5 siRNA-treated Huh7 cells were challenged with MERS-CoV, we detected a significant decrease in MERS-CoV entry (Fig. 6G). Overall, our data from protein blocking and siRNA knockdown experiments corroborated the findings of the antibody blocking assays, which supported CEACAM5 as an important cell surface protein in MERS-CoV infection.
FIG 6.

CEACAM5 recombinant protein and siRNA knockdown of CEACAM5 inhibit MERS-CoV entry. (A and B) MERS-CoV at an MOI of 0.1 was preincubated with human recombinant proteins at the indicated concentrations for 1 h at 37°C. After the preincubation, the protein-virus mixture was added to Calu3 cells (A) or Huh7 cells (B) for 2 h at 37°C. The cell lysates were subsequently harvested for qPCR analysis. Human recombinant DPP4 and human recombinant IgG were included as positive and negative controls, respectively. Results from the human recombinant DPP4-, human recombinant CEACAM5-, and control human recombinant IgG-treated samples were compared with those of the mock-treated samples. (C) Huh7 cells were treated with CEACAM5 protein for a total of 3 h during preincubation and virus inoculation (−1 to 2) or after virus inoculation (2 to 5). MERS-CoV entry was assessed by qPCR. (D) Huh7 cells were treated with 100 nM gene-specific or scrambled siRNA for two consecutive days. (E and F) Summary of the reduction of surface CEACAM5 and DPP4 expression. (G) siRNA-treated Huh7 cells were subjected to MERS-CoV infection at an MOI of 1 for 2 h at 37°C. The cell lysates were subsequently harvested for qPCR analysis, and the result was normalized to those of the mock-treated cells. The results from the DPP4 siRNA-, CEACAM5 siRNA-, and scrambled-siRNA-treated samples were compared with those of the mock-treated samples. The data are represented as means and standard deviations of the results of three independent experiments. Statistical analyses were carried out using Student's t test. Statistical significance is indicated by the asterisks (P < 0.05).
CEACAM5 is an attachment factor for MERS-CoV.
To further define the role of CEACAM5 in MERS-CoV infection, we examined the capacity of CEACAM5 to support MERS-CoV entry and attachment. To assess the role of CEACAM5 in MERS-CoV entry, CEACAM5-overexpressing BHK21 cells were inoculated with MERS-CoV at 37°C for 2 h. The cells were then washed and incubated at 37°C for 4 additional hours before being harvested for flow cytometry. Next, to assess the role of CEACAM5 in MERS-CoV attachment, CEACAM5-overexpressing BHK21 cells were inoculated with high-MOI MERS-CoV at 4°C for 2 h. The infected cells were subsequently washed, harvested, and immunolabeled for flow cytometry. In both cases, DPP4-overexpressing BHK21 cells were included as controls. The result of the entry assessment recapitulated the finding that human DPP4 overexpression could support MERS-CoV entry in the otherwise nonpermissive BHK21 cells. In stark contrast, human CEACAM5-expressing BHK21 cells failed to sustain the entry of MERS-CoV (Fig. 7A and B). In the attachment assessment, our data suggested that while MERS-CoV could attach to the surfaces of BHK21 cells, human DPP4 overexpression increased the amount of attached virus particles by around 3-fold. Intriguingly, in human CEACAM5-expressing BHK21 cells, the number of attached virus particles was also significantly increased, by more than 2-fold (Fig. 7C and D). To further verify the potential contribution of CEACAM5 in MERS-CoV entry, we overexpressed CEACAM5 in BHK21 and AD293 cells, followed by MERS-CoV infection. In corroboration of the results shown in Fig. 7A, our data showed that while CEACAM5 enhanced virus attachment to BHK21 cells (Fig. 8B), it could not function as an independent receptor for MERS-CoV entry in the nonpermissive BHK21 cells (Fig. 8A). Perhaps most importantly, overexpression of CEACAM5 increased the entry of MERS-CoV in permissive cells (Fig. 8C), in addition to enhancing the attachment of the virus (Fig. 8D), which suggested that CEACAM5 could contribute to MERS-CoV entry in conjunction with DPP4. To further investigate if CEACAM5 could facilitate the entry of other human coronaviruses, we challenged CEACAM5-transfected VeroE6 cells (Fig. 9A) with SARS-S pseudovirus and compared the pseudovirus entry with that of the empty-vector-transfected VeroE6 cells. Our results demonstrated that CEACAM5 overexpression did not significantly enhance the entry of SARS-S pseudovirus (Fig. 9B). Taken together, our data identified CEACAM5 as an important cell surface binding protein for MERS-CoV that could facilitate the entry of MERS-CoV by acting as an attachment factor.
FIG 7.

CEACAM5 is an attachment factor for MERS-CoV. (A and B) To assess whether CEACAM5 is important for MERS-CoV entry, human CEACAM5 was overexpressed in BHK21 cells. Human CEACAM5-expressing BHK21 cells were then challenged with MERS-CoV at an MOI of 5 for 2 h at 37°C. The inoculum was replaced with culture medium, and the cells were incubated for another 4 h before being harvested for flow cytometry. (C and D) To assess whether CEACAM5 is important for MERS-CoV attachment, human CEACAM5-expressing BHK21 cells were challenged with MERS-CoV at an MOI of 30 for 2 h at 4°C before being harvested for flow cytometry. Human DPP4-expressing BHK21 cells were included as controls for both entry and attachment assays. (B and D) The data are represented as the percentages of MERS-CoV NP-positive BHK21 cells after infection with or without DPP4/CEACAM5 overepression. The means and standard deviations were derived from the results of three independent experiments. Statistical analyses were carried out using Student's t test. Statistical significance is indicated by the asterisks (P < 0.05).
FIG 8.
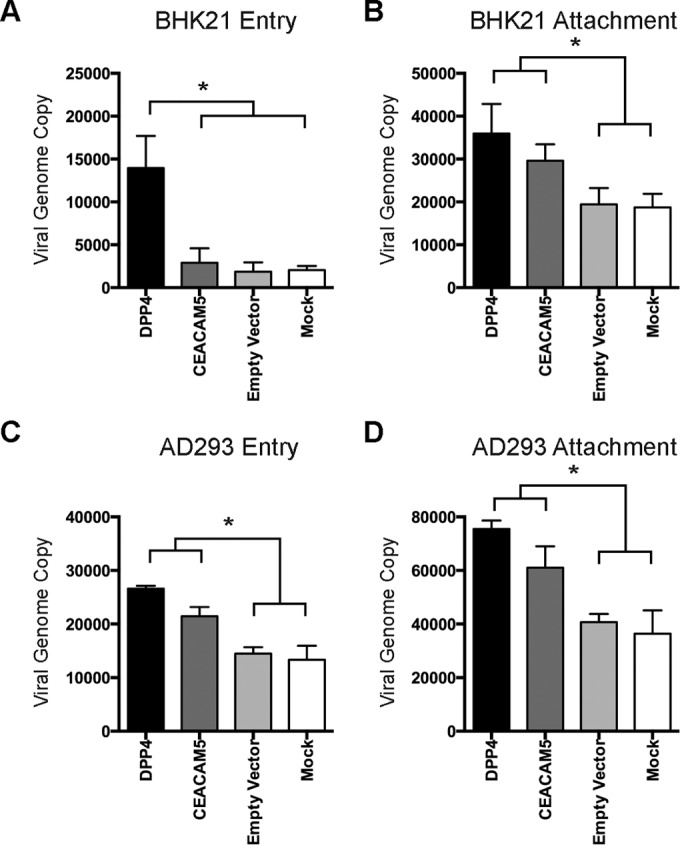
CEACAM5 overexpression does not confer infectivity to nonpermissive cells on MERS-CoV but enhances MERS-CoV entry into permissive cells. (A and C) To verify the role of CEACAM5 in MERS-CoV entry, human CEACAM5 was overexpressed in BHK21 cells or AD293 cells. The cells were then challenged with MERS-CoV at an MOI of 1 for 2 h at 37°C. After 2 h, the cells were washed extensively and harvested for qPCR analysis. (B and D) To verify the role of CEACAM5 in MERS-CoV attachment, human CEACAM5-expressing BHK21 cells or human CEACAM5-expressing AD293 cells were challenged with MERS-CoV at an MOI of 1 for 2 h at 4°C. The cell lysates were then harvested for qPCR analysis. DPP4- and empty-vector (pcDNA3.1)-expressing BHK21 cells were included as controls. The data are represented as means and standard deviations of the results of three independent experiments. The results from the human DPP4-, human CEACAM5-, and empty-vector-overexpressing samples were compared with those of the mock-treated samples. Statistical analyses were carried out using Student's t test. Statistical significance is indicated by the asterisks (P < 0.05).
FIG 9.
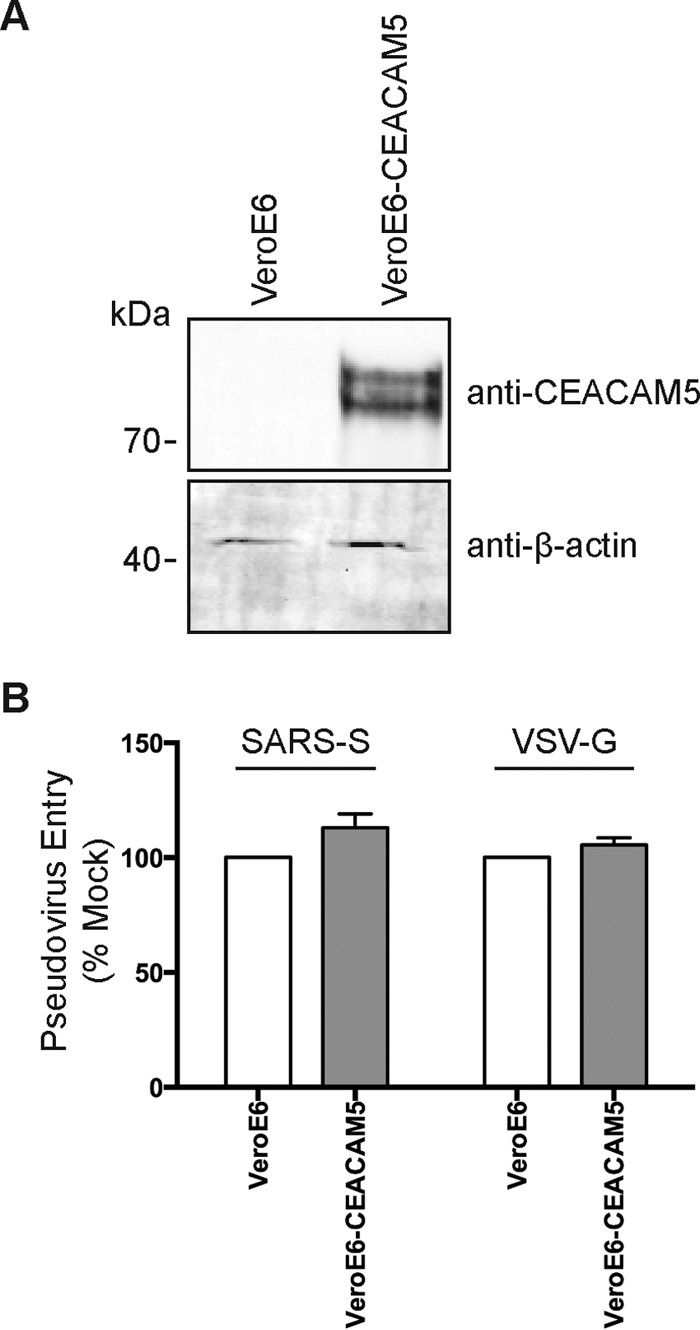
CEACAM5 overexpression does not facilitate SARS-CoV entry in VeroE6 cells. (A) VeroE6 cells were transfected with a CEACAM5-expressing plasmid or an empty vector. (B) The VeroE6 cells were challenged with SARS-S pseudovirus or VSV-G pseudovirus at a ratio of 100 LP per cell for 1 h. Luciferase activity from CEACAM5-transfected VeroE6 cells was determined at 48 h after pseudovirus challenge and was normalized to that of the empty-vector-transfected VeroE6 cells. (B) Means and standard deviations were derived from the results of three independent experiments. Statistical analyses were carried out using Student's t test. Statistical significance is indicated by the asterisks (P < 0.05).
DISCUSSION
The spike proteins of coronaviruses are known to bind a broad range of cellular targets, including sialic acids, sugars, and proteins (32). It is now clear that MERS-CoV predominantly utilizes DPP4 for cell entry (9). However, additional cellular binding targets of the MERS-CoV spike protein that may augment MERS-CoV entry have not been further explored. In the current study, using the VOPBA/LC–MS-MS approach, we unveiled CEACAM5 as a novel cell surface binding protein of MERS-CoV (Fig. 1). Intriguingly, expression analysis revealed that CEACAM5 was expressed on cell types that are highly permissive to MERS-CoV infection, including Calu3, Huh7, A549, and primary T cells (Fig. 2). CEACAM5 and DPP4 were coexpressed in the human lung, which highlighted the physiological relevance of CEACAM5 in MERS-CoV entry (Fig. 3). Subsequently, a series of co-IP studies demonstrated the capacity of CEACAM5 to bind to the spike protein of MERS-CoV in either overexpressed or endogenous settings (Fig. 4). Remarkably, when the interaction between CEACAM5 and MERS-CoV spike was perturbed with anti-CEACAM5 antibody (Fig. 5), recombinant CEACAM5 protein (Fig. 6), or CEACAM5 siRNA knockdown (Fig. 6), the infection of MERS-CoV was significantly inhibited. Notably, recombinant expression of CEACAM5 did not confer susceptibility to infection by MERS-CoV on nonpermissive BHK21 cells (Fig. 7A). Instead, CEACAM5 overexpression significantly enhanced the attachment of MERS-CoV to the BHK21 cells (Fig. 7B). Perhaps most importantly, the entry of MERS-CoV was increased when CEACAM5 was overexpressed in AD293 cells, which suggested that CEACAM5 could facilitate MERS-CoV entry in conjunction with DPP4 despite being unable to support MERS-CoV entry independently (Fig. 8). The interaction between CEACAM5 and MERS-CoV appeared to be specific, since CEACAM5 did not facilitate the entry of SARS-CoV (Fig. 9). Taken together, the results of our study identified CEACAM5 as a novel cell surface binding protein of MERS-CoV that facilitates MERS-CoV infection by augmenting the attachment of the virus to the cell surface.
CEACAM family members, including CEACAM5, are heavily glycosylated cell surface proteins (33, 34). Currently, 12 members of the CEACAM family have been identified in humans. Most CEACAMs are considered modulators of general cellular processes, including proliferation, motility, apoptosis, and attachment, as well as cell-cell interaction (35). Notably, aberrant CEACAM function is associated with tumor progression, and a number of CEACAMs are utilized as clinical biomarkers. In addition, members of the CEACAM family have been implicated in signal transduction and the binding of bacteria to host target cells. In particular, certain CEACAMs, including CEACAM5, are expressed on the apical membranes of epithelial cells and are exploited as receptors or binding factors for a number of pathogenic bacteria (35). Engagement of surface CEACAMs by these bacteria not only provides a portal of attachment, but also triggers internalization and entry of the bacteria into epithelial cells.
In the context of coronaviruses, previous reports have identified murine CEACAM1 as the principal cellular receptor of MHV (11). The discovery of CEACAM5 as a cellular binding target of MERS-CoV spike suggested that perhaps cell surface CEACAMs are common binding targets of coronavirus spike proteins that can be exploited for attachment or entry. It is tempting to speculate that other members of the coronavirus family can also make use of certain CEACAMs to facilitate infection. Notably, the spike proteins of coronaviruses are capable of binding to multiple cellular targets. As an example, SARS-CoV binds to ACE2 with the spike S1 C-terminal domain (CTD) (36). At the same time, the SARS-CoV spike can also bind DC-SIGN (16), DC-SIGNR (a closely related homolog of DC-SIGN) (16), and L-SIGN (15). Similarly, TGEV binds to APN with the spike S1 CTD (10), but the virus is also capable of binding to cell surface sugars and sialic acids using the N-terminal domain (NTD) of S1 (37). Since MHV binds CEACAM1 at the first 330 amino acids of the S1 NTD (38), one may postulate that MERS-CoV spike may similarly bind CEACAM5 with the NTD of S1. In this regard, the CEACAM5-S1 NTD interaction may orchestrate the DPP4-S1 CTD interaction (36, 39) and facilitate the infection of MERS-CoV. Importantly, the epitopes from the receptor-binding domain of MERS-CoV S1 are known to trigger the production of potent neutralization antibodies. It is potentially feasible to include additional epitopes derived from the CEACAM5-binding domain in future vaccine designs, which may further enhance the efficiency of the neutralizing antibodies produced while at the same time avoiding potential immunopathological effects observed when using the full-length spike protein (40).
By definition, a receptor is a cellular factor essential for viral entry and bound by the viral glycoproteins, whereas an attachment factor is a cellular factor that can increase the efficiency of virus entry but is not sufficient to render otherwise refractory cells susceptible to infection. In line with the definitions, CEACAM5 expression was predominantly detected in cell types highly susceptible to MERS-CoV infection, including Huh7, Calu3, and T cells. Therefore, the expression of CEACAM5 may in part contribute to the permissiveness of a certain cell type in addition to DPP4. Our data showed that the expression of CEACAM5 did not confer infectivity of MERS-CoV for the nonpermissive BHK21 cells. However, the expression of CEACAM5 markedly increased the number of virus particles that were bound to the cell surface. Subsequent analyses confirmed that CEACAM5 expression could enhance MERS-CoV entry in AD293 cells, which expressed endogenous DPP4. Therefore, mechanistically, CEACAM5 may help to concentrate the virus particles at the cell surface and provide the virus particles with greater accessibility to DPP4, which can then be used to initiate the cell entry process.
In conclusion, in this study, we identified CEACAM5 as a novel surface binding protein for MERS-CoV that could facilitate MERS-CoV entry by serving as an important attachment factor. This novel cell surface factor could be a potential antiviral target in the fight against MERS-CoV infection.
ACKNOWLEDGMENTS
We thank Suet Yi Leung for providing the normal human lung sections. We thank the staff at the Core Facility, Li Ka Shing Faculty of Medicine, University of Hong Kong, for facilitation of the study.
This work was partly supported by the Providence Foundation Ltd. in memory of the late Lui Hac Minh; the Hong Kong Health and Medical Research Fund (14131392 and 15140762); the NSFC/RGC Joint Research Scheme (N_HKU728/14) and the Theme-Based Research Scheme (T11/707/15) of the Research Grants Council, Hong Kong Special Administrative Region; and funding from the Ministry of Education of China for the Collaborative Innovation Center for Diagnosis and Treatment of Infectious Diseases.
The funding sources had no role in the study design, data collection, analysis, interpretation, or writing of the report. We report no conflicts of interest.
Funding Statement
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
REFERENCES
- 1.Chan JF, Lau SK, To KK, Cheng VC, Woo PC, Yuen KY. 2015. Middle East respiratory syndrome coronavirus: another zoonotic betacoronavirus causing SARS-like disease. Clin Microbiol Rev 28:465–522. doi: 10.1128/CMR.00102-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Chan JF, To KK, Tse H, Jin DY, Yuen KY. 2013. Interspecies transmission and emergence of novel viruses: lessons from bats and birds. Trends Microbiol 21:544–555. doi: 10.1016/j.tim.2013.05.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Yeager CL, Ashmun RA, Williams RK, Cardellichio CB, Shapiro LH, Look AT, Holmes KV. 1992. Human aminopeptidase N is a receptor for human coronavirus 229E. Nature 357:420–422. doi: 10.1038/357420a0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Vlasak R, Luytjes W, Spaan W, Palese P. 1988. Human and bovine coronaviruses recognize sialic acid-containing receptors similar to those of influenza C viruses. Proc Natl Acad Sci U S A 85:4526–4529. doi: 10.1073/pnas.85.12.4526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Li W, Moore MJ, Vasilieva N, Sui J, Wong SK, Berne MA, Somasundaran M, Sullivan JL, Luzuriaga K, Greenough TC, Choe H, Farzan M. 2003. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature 426:450–454. doi: 10.1038/nature02145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Hofmann H, Pyrc K, van der Hoek L, Geier M, Berkhout B, Pohlmann S. 2005. Human coronavirus NL63 employs the severe acute respiratory syndrome coronavirus receptor for cellular entry. Proc Natl Acad Sci U S A 102:7988–7993. doi: 10.1073/pnas.0409465102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Huang X, Dong W, Milewska A, Golda A, Qi Y, Zhu QK, Marasco WA, Baric RS, Sims AC, Pyrc K, Li W, Sui J. 2015. Human coronavirus HKU1 spike protein uses O-acetylated sialic acid as an attachment receptor determinant and employs hemagglutinin-esterase protein as a receptor-destroying enzyme. J Virol 89:7202–7213. doi: 10.1128/JVI.00854-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Zaki AM, van Boheemen S, Bestebroer TM, Osterhaus AD, Fouchier RA. 2012. Isolation of a novel coronavirus from a man with pneumonia in Saudi Arabia. N Engl J Med 367:1814–1820. doi: 10.1056/NEJMoa1211721. [DOI] [PubMed] [Google Scholar]
- 9.Raj VS, Mou H, Smits SL, Dekkers DH, Muller MA, Dijkman R, Muth D, Demmers JA, Zaki A, Fouchier RA, Thiel V, Drosten C, Rottier PJ, Osterhaus AD, Bosch BJ, Haagmans BL. 2013. Dipeptidyl peptidase 4 is a functional receptor for the emerging human coronavirus-EMC. Nature 495:251–254. doi: 10.1038/nature12005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Delmas B, Gelfi J, L'Haridon R, Vogel LK, Sjostrom H, Noren O, Laude H. 1992. Aminopeptidase N is a major receptor for the entero-pathogenic coronavirus TGEV. Nature 357:417–420. doi: 10.1038/357417a0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Williams RK, Jiang GS, Holmes KV. 1991. Receptor for mouse hepatitis virus is a member of the carcinoembryonic antigen family of glycoproteins. Proc Natl Acad Sci U S A 88:5533–5536. doi: 10.1073/pnas.88.13.5533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Schultze B, Krempl C, Ballesteros ML, Shaw L, Schauer R, Enjuanes L, Herrler G. 1996. Transmissible gastroenteritis coronavirus, but not the related porcine respiratory coronavirus, has a sialic acid (N-glycolylneuraminic acid) binding activity. J Virol 70:5634–5637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Schwegmann-Wessels C, Zimmer G, Schroder B, Breves G, Herrler G. 2003. Binding of transmissible gastroenteritis coronavirus to brush border membrane sialoglycoproteins. J Virol 77:11846–11848. doi: 10.1128/JVI.77.21.11846-11848.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Milewska A, Zarebski M, Nowak P, Stozek K, Potempa J, Pyrc K. 2014. Human coronavirus NL63 utilizes heparan sulfate proteoglycans for attachment to target cells. J Virol 88:13221–13230. doi: 10.1128/JVI.02078-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Jeffers SA, Tusell SM, Gillim-Ross L, Hemmila EM, Achenbach JE, Babcock GJ, Thomas WD Jr, Thackray LB, Young MD, Mason RJ, Ambrosino DM, Wentworth DE, Demartini JC, Holmes KV. 2004. CD209L (L-SIGN) is a receptor for severe acute respiratory syndrome coronavirus. Proc Natl Acad Sci U S A 101:15748–15753. doi: 10.1073/pnas.0403812101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Marzi A, Gramberg T, Simmons G, Moller P, Rennekamp AJ, Krumbiegel M, Geier M, Eisemann J, Turza N, Saunier B, Steinkasserer A, Becker S, Bates P, Hofmann H, Pohlmann S. 2004. DC-SIGN and DC-SIGNR interact with the glycoprotein of Marburg virus and the S protein of severe acute respiratory syndrome coronavirus. J Virol 78:12090–12095. doi: 10.1128/JVI.78.21.12090-12095.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Chan CM, Lau SK, Woo PC, Tse H, Zheng BJ, Chen L, Huang JD, Yuen KY. 2009. Identification of major histocompatibility complex class I C molecule as an attachment factor that facilitates coronavirus HKU1 spike-mediated infection. J Virol 83:1026–1035. doi: 10.1128/JVI.01387-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Zumla A, Chan JF, Azhar EI, Hui DS, Yuen KY. 2016. Coronaviruses—drug discovery and therapeutic options. Nat Rev Drug Discov 15:327–347. doi: 10.1038/nrd.2015.37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Yeung ML, Yao Y, Jia L, Chan JF, Chan KH, Cheung KF, Chen HL, Poon VK, Tsang AK, To KK, Yiu MK, Teng JL, Chu H, Zhou J, Zhang Q, Deng W, Lau SK, Lau JY, Woo PC, Chan TM, Yung S, Zheng BJ, Jin DY, Mathieson PW, Qin C, Yuen KY. 2016. MERS coronavirus induces apoptosis in kidney and lung by upregulating Smad7 and FGF2. Nat Microbiol 1:16004. doi: 10.1038/nmicrobiol.2016.4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.WHO. 16 May 2016. Middle East respiratory syndrome coronavirus (MERS-CoV)—Saudi Arabia. WHO, Geneva, Switzerland: http://www.who.int/csr/don/16-may-2016-mers-saudi-arabia/en/. [Google Scholar]
- 21.Assiri A, Al-Tawfiq JA, Al-Rabeeah AA, Al-Rabiah FA, Al-Hajjar S, Al-Barrak A, Flemban H, Al-Nassir WN, Balkhy HH, Al-Hakeem RF, Makhdoom HQ, Zumla AI, Memish ZA. 2013. Epidemiological, demographic, and clinical characteristics of 47 cases of Middle East respiratory syndrome coronavirus disease from Saudi Arabia: a descriptive study. Lancet Infect Dis 13:752–761. doi: 10.1016/S1473-3099(13)70204-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Muller MA, Raj VS, Muth D, Meyer B, Kallies S, Smits SL, Wollny R, Bestebroer TM, Specht S, Suliman T, Zimmermann K, Binger T, Eckerle I, Tschapka M, Zaki AM, Osterhaus AD, Fouchier RA, Haagmans BL, Drosten C. 2012. Human coronavirus EMC does not require the SARS-coronavirus receptor and maintains broad replicative capability in mammalian cell lines. mBio 3:e00515-12. doi: 10.1128/mBio.00515-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Chan JF, Chan KH, Choi GK, To KK, Tse H, Cai JP, Yeung ML, Cheng VC, Chen H, Che XY, Lau SK, Woo PC, Yuen KY. 2013. Differential cell line susceptibility to the emerging novel human betacoronavirus 2c EMC/2012: implications for disease pathogenesis and clinical manifestation. J Infect Dis 207:1743–1752. doi: 10.1093/infdis/jit123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Zhou J, Chu H, Chan JF, Yuen KY. 2015. Middle East respiratory syndrome coronavirus infection: virus-host cell interactions and implications on pathogenesis. Virol J 12:218. doi: 10.1186/s12985-015-0446-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Chu H, Zhou J, Wong BH, Li C, Chan JF, Cheng ZS, Yang D, Wang D, Lee AC, Li C, Yeung ML, Cai JP, Chan IH, Ho WK, To KK, Zheng BJ, Yao Y, Qin C, Yuen KY. 2016. Middle East respiratory syndrome coronavirus efficiently infects human primary T lymphocytes and activates the extrinsic and intrinsic apoptosis pathways. J Infect Dis 213:904–914. doi: 10.1093/infdis/jiv380. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Chan CM, Chu H, Zhang AJ, Leung LH, Sze KH, Kao RY, Chik KK, To KK, Chan JF, Chen H, Jin DY, Liu L, Yuen KY. 2016. Hemagglutinin of influenza A virus binds specifically to cell surface nucleolin and plays a role in virus internalization. Virology 494:78–88. doi: 10.1016/j.virol.2016.04.008. [DOI] [PubMed] [Google Scholar]
- 27.Zhou J, Chu H, Li C, Wong BH, Cheng ZS, Poon VK, Sun T, Lau CC, Wong KK, Chan JY, Chan JF, To KK, Chan KH, Zheng BJ, Yuen KY. 2014. Active replication of Middle East respiratory syndrome coronavirus and aberrant induction of inflammatory cytokines and chemokines in human macrophages: implications for pathogenesis. J Infect Dis 209:1331–1342. doi: 10.1093/infdis/jit504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Chan JF, Yao Y, Yeung ML, Deng W, Bao L, Jia L, Li F, Xiao C, Gao H, Yu P, Cai JP, Chu H, Zhou J, Chen H, Qin C, Yuen KY. 2015. Treatment with lopinavir/ritonavir or interferon-beta1b improves outcome of MERS-CoV infection in a nonhuman primate model of common marmoset. J Infect Dis 212:1904–1913. doi: 10.1093/infdis/jiv392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Chu H, Zhou J, Wong BH, Li C, Cheng ZS, Lin X, Poon VK, Sun T, Lau CC, Chan JF, To KK, Chan KH, Lu L, Zheng BJ, Yuen KY. 2014. Productive replication of Middle East respiratory syndrome coronavirus in monocyte-derived dendritic cells modulates innate immune response. Virology 454-455:197–205. doi: 10.1016/j.virol.2014.02.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.van Doremalen N, Miazgowicz KL, Milne-Price S, Bushmaker T, Robertson S, Scott D, Kinne J, McLellan JS, Zhu J, Munster VJ. 2014. Host species restriction of Middle East respiratory syndrome coronavirus through its receptor, dipeptidyl peptidase 4. J Virol 88:9220–9232. doi: 10.1128/JVI.00676-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Hammarstrom S. 1999. The carcinoembryonic antigen (CEA) family: structures, suggested functions and expression in normal and malignant tissues. Semin Cancer Biol 9:67–81. doi: 10.1006/scbi.1998.0119. [DOI] [PubMed] [Google Scholar]
- 32.Graham RL, Donaldson EF, Baric RS. 2013. A decade after SARS: strategies for controlling emerging coronaviruses. Nat Rev Microbiol 11:836–848. doi: 10.1038/nrmicro3143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Han SU, Kwak TH, Her KH, Cho YH, Choi C, Lee HJ, Hong S, Park YS, Kim YS, Kim TA, Kim SJ. 2008. CEACAM5 and CEACAM6 are major target genes for Smad3-mediated TGF-beta signaling. Oncogene 27:675–683. doi: 10.1038/sj.onc.1210686. [DOI] [PubMed] [Google Scholar]
- 34.Roda G, Jianyu X, Park MS, DeMarte L, Hovhannisyan Z, Couri R, Stanners CP, Yeretssian G, Mayer L. 2014. Characterizing CEACAM5 interaction with CD8alpha and CD1d in intestinal homeostasis. Mucosal Immunol 7:615–624. doi: 10.1038/mi.2013.80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Tchoupa AK, Schuhmacher T, Hauck CR. 2014. Signaling by epithelial members of the CEACAM family-mucosal docking sites for pathogenic bacteria. Cell Commun Signal 12:27. doi: 10.1186/1478-811X-12-27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Lu G, Wang Q, Gao GF. 2015. Bat-to-human: spike features determining ‘host jump’ of coronaviruses SARS-CoV, MERS-CoV, and beyond. Trends Microbiol 23:468–478. doi: 10.1016/j.tim.2015.06.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Li F. 2015. Receptor recognition mechanisms of coronaviruses: a decade of structural studies. J Virol 89:1954–1964. doi: 10.1128/JVI.02615-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Kubo H, Yamada YK, Taguchi F. 1994. Localization of neutralizing epitopes and the receptor-binding site within the amino-terminal 330 amino acids of the murine coronavirus spike protein. J Virol 68:5403–5410. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Du L, Zhao G, Kou Z, Ma C, Sun S, Poon VK, Lu L, Wang L, Debnath AK, Zheng BJ, Zhou Y, Jiang S. 2013. Identification of a receptor-binding domain in the S protein of the novel human coronavirus Middle East respiratory syndrome coronavirus as an essential target for vaccine development. J Virol 87:9939–9942. doi: 10.1128/JVI.01048-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Du L, Tai W, Zhou Y, Jiang S. 2016. Vaccines for the prevention against the threat of MERS-CoV. Expert Rev Vaccines 67603:1–12. [DOI] [PMC free article] [PubMed] [Google Scholar]