

Abstract
Purpose
Discovery of single nucleotide polymorphisms (SNPs) that predict a patient's risk of docetaxel-induced neuropathy would enable treatment individualization to maximize efficacy and avoid unnecessary toxicity. The objectives of this analysis were to discover SNPs associated with docetaxel-induced neuropathy and mechanistically validate these associations in preclinical models of drug-induced neuropathy.
Experimental Design
A genome-wide association study was conducted in metastatic castrate-resistant prostate cancer patients treated with docetaxel, prednisone and randomized to bevacizumab or placebo on CALGB 90401. SNPs were genotyped on the Illumina HumanHap610-Quad platform followed by rigorous quality control. The inference was conducted on the cumulative dose at occurrence of grade 3+ sensory neuropathy using a cause-specific hazard model that accounted for early treatment discontinuation. Genes with SNPs significantly associated with neuropathy were knocked down in cellular and mouse models of drug-induced neuropathy.
Results
498,081 SNPs were analyzed in 623 Caucasian patients, 50 (8%) of whom experienced grade 3+ neuropathy. The 1000 SNPs most associated with neuropathy clustered in relevant pathways including neuropathic pain and axonal guidance. A SNP in VAC14 (rs875858) surpassed genome-wide significance (p=2.12×10-8 adjusted p=5.88×10-7). siRNA knockdown of VAC14 in stem cell derived peripheral neuronal cells increased docetaxel sensitivity as measured by decreased neurite processes (p=0.0015) and branches (p<0.0001). Prior to docetaxel treatment VAC14 heterozygous mice had greater nociceptive sensitivity than wild-type litter mate controls (p=0.001).
Conclusions
VAC14 should be prioritized for further validation of its potential role as a predictor of docetaxel-induced neuropathy and biomarker for treatment individualization.
Keywords: docetaxel, paclitaxel, pharmacogenetics, genome-wide association study, heritability, single nucleotide polymorphism, chemotherapy-induced peripheral neuropathy, VAC14, peripheral neuronal cell model, animal model
Introduction
The taxane class includes three US Food and Drug Administration (FDA)-approved chemotherapeutic agents, paclitaxel, docetaxel and cabazitaxel, that have activity in a variety of solid tumors including lung, breast, ovarian, gastric, and prostate cancers. Taxanes bind to and stabilize microtubules, ultimately inhibiting the mitotic phase of cell cycle development(1). The taxanes and other microtubule-targeting chemotherapeutic agents (e.g., vinca alkaloids) commonly induce peripheral neuropathy of varying severity(2). Chemotherapy-induced peripheral neuropathy (CIPN) often presents as a combination of paresthesia and dysesthesia, and can progress to irreversible loss of balance and dexterity with continued treatment(3). Due to the progressive nature of the toxicity and its potential effect on long-term quality of life(4), CIPN can necessitate treatment discontinuation, even in the context of active tumor control.
Patient demographic (i.e., race and history of neuropathy) and treatment (i.e., cumulative dose and drug exposure) factors have been associated with CIPN(5-8), but there are currently no effective methods for predicting CIPN risk and/or severity. Genome-wide association studies (GWAS) of paclitaxel and vincristine-induced CIPN confirm that genetic variation modifies patients' risk(9-12). The discovery and validation of single nucleotide polymorphisms (SNPs) that influence risk of docetaxel-induced peripheral neuropathy would be of clinical benefit, enabling identification of high-risk patients in whom taxane use should be avoided if possible. SNPs that modulate risk of docetaxel-induced neuropathy have been reported from candidate gene studies(13-15); however, no study has taken a genome-wide approach. Based on the previous success of GWAS in discovering SNPs that affect paclitaxel-induced neuropathy, a similar analysis was conducted in a cohort of docetaxel-treated patients to identify genetic predictors of docetaxel-induced neuropathy.
GWAS was performed in a large, prospectively enrolled, chemotherapy-naive cohort of metastatic castrate-resistant prostate cancer (mCRPC) patients who were treated with up to two years of docetaxel and prednisone, with half randomized to concurrent bevacizumab(16). As these metastatic patients are at high risk of early treatment discontinuation due to disease progression and/or death, which precludes occurrence of neuropathy, a competing-risks adjusted statistical model was utilized(17, 18). Following discovery of a SNP that surpassed genome-wide significance, pharmacogenetic replication between the results of a previously conducted GWAS of paclitaxel-induced neuropathy(9) and this GWAS of docetaxel-induced neuropathy and mechanistic validation of that gene in cellular and animal models were attempted to determine whether genetic predictors of CIPN were similar between the taxanes and to potentially validate clinically useful genetic predictors of docetaxel-induced sensory peripheral neuropathy
Methods
Patients and toxicity
Cancer and Leukeumia Group B (CALGB/Alliance) 90401 was a double-blinded phase III trial that equally randomized men with hormone-refractory prostate cancer to receive docetaxel and prednisone with or without bevacizumab for up to two years(16). All patients enrolled in the CALGB 90401 parent study who provided IRB-approved informed consent for a pharmacogenomic substudy (CALGB 60404) were eligible for this GWAS. Briefly, patient eligibility included histologically documented adenocarcinoma of the prostate that had progressed while the patient was on hormone deprivation therapy. Relevant exclusion criteria included prior chemotherapy or anti-angiogenesis therapy or clinically significant (grade 2+) peripheral neuropathy. Toxicity data were collected by the Alliance Statistics and Data Center at each treatment cycle on standardized forms that mandated reporting of grade 3+ peripheral sensory neuropathy, as defined by National Cancer Institute Common Toxicity Criteria for Adverse Events (NCI CTCAE) version 3.0. Additionally, one case was reported as a grade 3 cranial neuropathy event described as “sensory-facial.” Only neuropathy that was considered by the clinician to be possibly, probably, or definitely attributable to treatment, and occurred within 30 days of docetaxel administration, was included in the analysis.
Docetaxel treatment
All patients received docetaxel 75 mg/m2 infused over 1 hour on day 1 of each 21-day cycle with 8 mg oral dexamethasone 12, 3, and 1 hour prior to infusion. Patients on both arms also received 5 mg oral prednisone twice daily and were randomized to 15 mg/kg bevacizumab or placebo by intravenous infusion on day 1 of each cycle. Use of growth factor, aspirin, anti-emetics and luteinizing-hormone releasing hormone agonists was under the discretion of the treating physician. Docetaxel administration was held for neutropenia (absolute neutrophil count <1,500 cells/mm3) and the dose was decreased in increments of 10 mg/m2 for hepatic dysfunction, neurotoxicity, gastrointestinal toxicity or febrile neutropenia. The protocol mandated discontinuation of docetaxel treatment if the patient required more than two docetaxel dose decreases or in the event of specific toxicities or confirmed cancer progression.
Genotyping
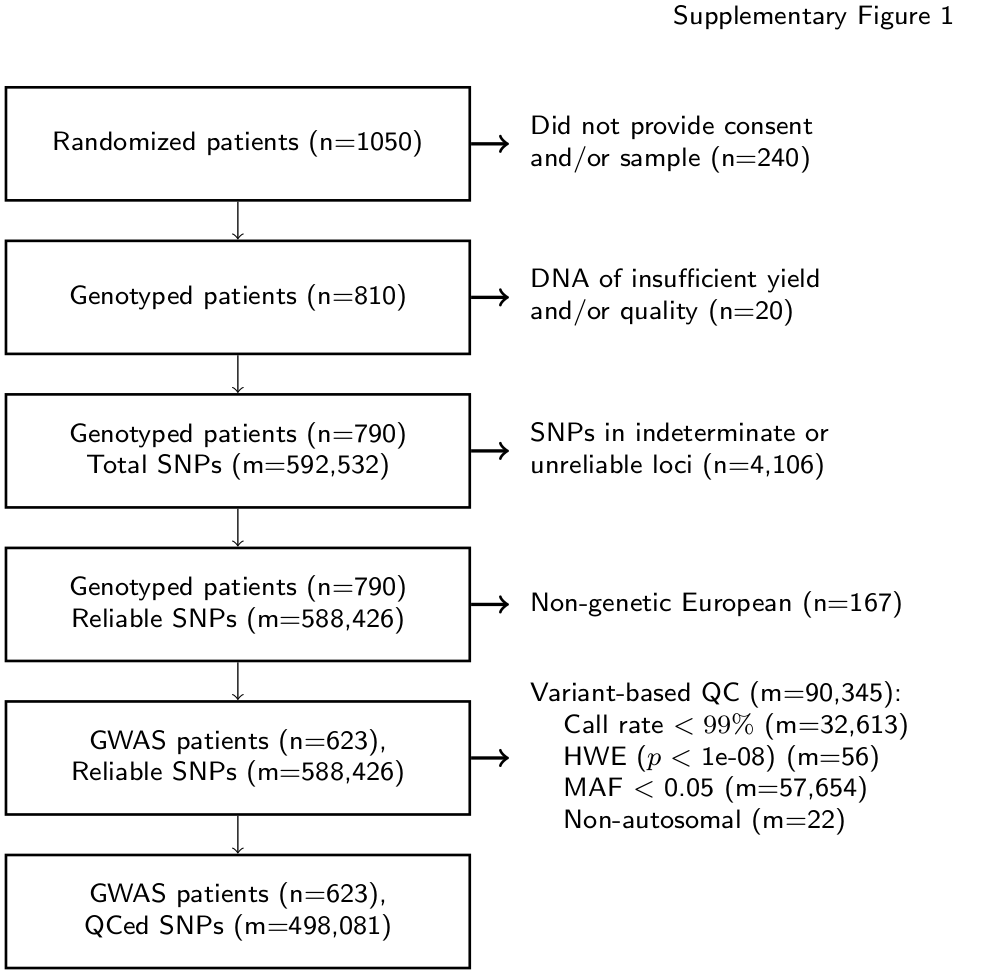
A 10-mL sample of whole blood was collected from all patients enrolling on the pharmacogenomic substudy prior to initiation of protocol treatment. Genotyping and genetic quality control were similar to that previously reported in the CALGB 40101 GWAS(9). Genotyping was performed on the HumanHap610-Quad Genotyping BeadChip (Illumina Inc., CA, USA) at the RIKEN Center for Genomic Medicine (Yokohama City, Japan). Genotype data is available at dbGap (http://www.ncbi.nlm.nih.gov/projects/gap/cgi-bin/study.cgi?study_id=phs001002.v1.p1) SNPs that were known indeterminate (n=4,106) or unreliable loci (Tech Note: Infinium® Genotyping Data Analysis, 2007) and patients with SNP call rate <95% were excluded. Eigensoft version 3.0 was used to visualize genetic ancestry for the 790 evaluable patients to identify a genetic European population (n=623) in which the analysis was conducted. Additional SNPs were excluded for low call rate (<99%), low minor allele frequency (<0.05), p-value of Hardy-Weinberg distribution <10-8 or non-autosomal loci, leaving 498,081 SNPs for analysis (Supplementary Figure 1).
Cellular Sensitivity Experiments
Commercially available human induced pluripotent stem cell (iPSC)-derived peripheral neurons (Peri.4U) were purchased from Axiogenesis (Cologne, Germany) and plated at 1 × 104 cells/well in 100 μL media as per manufactures' instructions. Figure 3A illustrates the scheme for this experiment. Dharmacon Accell technology (GE Dharmacon, Lafayette, CO, USA) was applied at 4 hours post-plating using 1 μM human siVAC14 SMARTpool or the non-targeting control (NTC). 24 hours later, transfection media was exchanged for media containing 0.17% DMSO vehicle control and either docetaxel (LKT Laboratories Inc., St. Paul, MN) or paclitaxel (Sigma-Aldrich, St. Louis, MO). The drugs were prepared in a darkened hood by dissolving powder in 100% DMSO to obtain a stock solution of 58.4 mM. Stock drug was serially diluted in media for final dosing concentrations ranging from 10 pM to 1 μM, increasing by factors of ten. Control wells were treated with 0.17% final concentration of DMSO to match drug treatments. After 24 or 48 hour drug treatments, neurons were stained with 0.3 ng/mL Hoechst 33342 (Sigma-Aldrich) and 1.5 μg/mL Calcein AM (Molecular Probes, Life Technologies Inc.) and imaged at 10× magnification using an ImageXpress Micro imaging microscope (Molecular Devices, LLC, Sunnyvale, CA) at the University of Chicago Cellular Screening Center. Neurite changes were determined by individual cell measurements of relative total neurite outgrowth, relative number of processes, relative number of branches, mean/median/max process length, relative cell body area, relative straightness and relative mean outgrowth intensity. The data represents triplicate experiments making measurements on more than 1000 cells per treatment as previously described(19). Two wells of transfected cells were collected per timepoint using the Cells-to-Ct kit (Ambion, Life Technologies Inc., Carlsbad, CA) and reversed transcribed as per kit instructions. Quantitative real time reverse transcription polymerase chain reaction (qRT-PCR) was performed using Taqman primers (Fisher Scientific LLC, Hanover Park, IL) for VAC14 (Hs00947931_m1) and compared to human beta-2-microglobulin (NM_004048.2) housekeeping gene at 24, 48 and 72 hours post-transfection (as previously described; (19).
Figure 3. VAC14 Knockdown increases neuronal cell sensitivity to docetaxel but decreases sensitivity to paclitaxel.
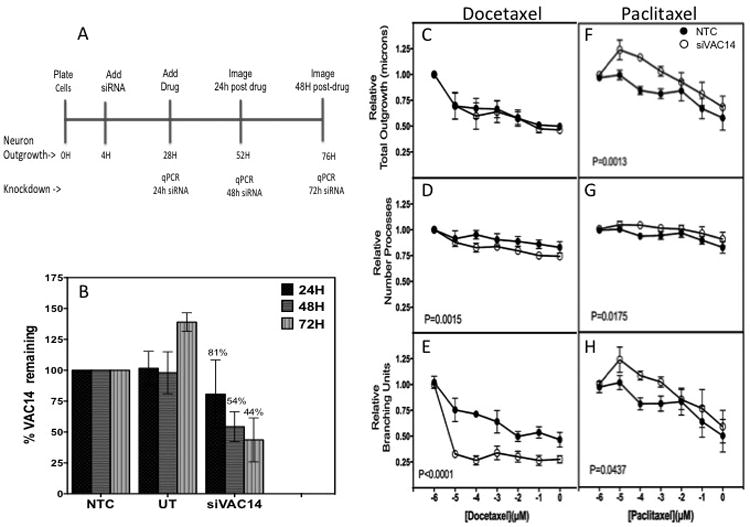
A) Peripheral neuron (Peri.4U) experimental scheme illustrating timing of treatment and imaging. B) Relative VAC14 expression at 24, 48 and 72 hours post-transfection with siVAC14 compared to non-targeting control (NTC) produced a steady decline of VAC14 expression over time. The neurite outgrowth changes after 24 hours docetaxel (48 hours knockdown) showed no significance as measured by C) total outgrowth but did result in greater sensitivity to docetaxel in D) relative number of processes and E) relative number of branches. Alternatively, VAC14 knockdown decreased peripheral neuronal cell sensitivity to paclitaxel, 3F-3H. Open circle represent data from VAC14 siRNA and closed circle from NTC siRNA. Neurite parameters measured with MetaXPress® software were averaged from 3 independent experiments. Error bars represent the standard error of the mean from three independent transfection experiments. NTC = non-targeting control, UT= untransfected control.
Mouse Sensitivity Experiments
Mice bred for VAC14 heterozygosity (VAC14+/-) and VAC14 wild-type litter mates (VAC14+/+) on C57/BL6 background were obtained from Lois Weisman at University of Michigan and acclimated to living conditions at University of Maryland(20). Mice 3-5 months of age were randomly assigned to docetaxel or placebo treatment, with 6-7 mice in each treatment arm. The entire experiment was conducted separately in male and female mice. Intraperitoneal administration of docetaxel 2 mg/kg or saline vehicle was administered on days 1, 3, 5, and 7.
Behavioral testing for the development of increased mechanical sensitivity took place at baseline (prior to treatment), day 5 (after two doses), day 8 (24 hours after the fourth dose), and weekly for 3 additional assessments. Mechanical sensitivity was measured in mice from both treatment arms simultaneously by a blinded individual using von Frey filaments, as previously described(21). Briefly, mice were placed in individual Plexiglas® cubes on an elevated wire mesh platform and allowed to acclimate for approximately one hour, when grooming and exploration behaviors ended. A series of von Frey filaments (Touch Test Sensory Evaluator Kit, myNeurolab.com, St. Louis, MO), with bending forces of 0.04-2.0 g, were applied to the plantar surface of the left hind paw until the filament just bent and was held in place for 5 seconds or until the mouse withdrew its paw. A positive response was defined as a brisk withdrawal, with or without shaking or licking, of the hind paw during or immediately upon removal of the filament application. Each filament was tested 5 times on each hind paw starting with the 0.4 g filament. If the 0.4 g filament elicited 3 positive responses out of 5 trials, then testing continued moving downward through the series to the 0.04 g filament and the number of withdrawals was recorded for each filament. If the 0.4 g filament did not elicit 3 positive responses, then testing continued moving upward through the series to the 2.0 g filament and the number of withdrawals was recorded for each filament. Threshold was defined as the filament with the lowest bending force that elicited at least 3 positive responses out of 5 trials.
Statistics
The primary endpoint for GWAS analysis was the cumulative docetaxel dose (mg/m2) at first report of treatment-related grade 3+ sensory peripheral neuropathy. Patients who did not complete two years of therapy due to progression/death, treatment-terminating adverse event, or any other reason were informatively censored at the cumulative docetaxel dose received; therefore, the statistical analysis was conducted within a competing-risk model framework with the inference conducted on the basis of cause-specific hazard(17, 18). Cumulative incidence curves were constructed to visualize the influence of each of the top three hits with neuropathy incidence. The relationship between each SNP and neuropathy was adjusted for clinical covariates with putative relevance to neuropathy risk: diabetes (reported history of diabetes or current diabetes treatment vs. none), age (continuous), body mass index (BMI) (≤ 30 kg/m2 vs. > 30 kg/m2), and treatment arm (bevacizumab vs. placebo), regardless of whether the covariate was significantly associated with neuropathy in this cohort.
Cross-study clinical replication of the SNP with the greatest association with neuropathy was attempted in a cohort of Caucasian breast cancer patients treated with paclitaxel on the CALGB 40101 clinical trial(22). GWAS of the cumulative paclitaxel dose at occurrence of grade 2+ neuropathy has previously been published(9). Additionally, a SNP predictive of paclitaxel-induced neuropathy risk (rs10771973, FGD4) discovered and replicated in the CALGB 40101 GWAS was interrogated in the GWAS results from the docetaxel-treated Caucasian patients on CALGB 90401 to assess whether the SNPs that influence neuropathy risk are similar between the two taxanes.
Statistical analyses were performed by Alliance statisticians on a database locked on August 15, 2012. All analyses were conducted using the R [R]statistical environment version 3.1.1(23) or higher, using extension packages survival (version 2.37-7)(24), GenABEL (version 1.8-0)(25), cmprsk (version 2.2-7)(26) and interval (version 1.1-0.1)(27). To search for cumulative genetic effects, the genes that mapped to the top 1000 SNPs were included in a pathway enrichment analysis using the Ingenuity Pathway Analysis version 17199142 (Ingenuity® Systems, Inc, www.ingenuity.com) internal algorithm. Peripheral neuronal cell phenotypes (outgrowth, processes, branching, etc.) measured after treatment with increasing docetaxel and paclitaxel concentrations were compared between VAC14 siRNA and non-targeted control siRNA using two-way ANOVA.
Mouse mechanical sensitivity thresholds were compared prior to docetaxel treatment between VAC14+/+ and VAC14+/- mice using an interval-censored analysis with the minimum fiber size inducing a response as the phenotype. The genotype groups in the untreated mice were compared again on day 28. The same method was used to compare sensitivity thresholds of docetaxel treated and untreated mice following treatment on day 8. To assess whether VAC14+/- mice were more sensitive to docetaxel-induced changes in mechanical stimuli, parametric survival regression was used to test for the effect of VAC14 by treatment interaction on withdrawal threshold on day 8, after treatment was completed.
Results
Genome-wide Association Study
The CALGB/Alliance 90401 parent study enrolled 1,050 patients, of whom 863 provided consent and registered for the pharmacogenomics substudy CALGB 60604 (CALGB is now a part of the Alliance for Clinical Trials in Oncology). The GWAS included 790 patients, of whom only the 616 self-reported, genetically defined Europeans receiving treatment were included in the analysis to minimize confounding from population substructure (Supplementary Figure 1). Demographic characteristics for the discovery cohort including baseline covariates potentially relevant to treatment-induced neuropathy risk are reported in Table 1. The median age was 69 years, body mass index (BMI) was 29 kg/m2, and 97 patients (16%) had a history of diabetes. Randomization was nearly even to the bevacizumab (314 patients, 51%) and placebo (302, 49%) arms.
Table 1.
Demographic Data for Patients Included in GWAS (n=616).
| Age | Median (years) | 68.9 |
| Range (years) | 41.7-93.5 | |
| Body mass index | Median (kg/m2) | 29.0 |
| Range (kg/m2) | 16.1-53.3 | |
| Diabetes | Yes | 97 (15.8%) |
| No | 519 (84.3%) | |
| Treatment arm | Bevacizumab | 314 (51.0%) |
| Placebo | 302 (49.0%) | |
| Cumulative docetaxel dose | Median (mg/m2) | 626 |
| Range (mg/m2) | 72-2,942 | |
| ECOG performance status | 0 | 340 (55.2%) |
| 1 | 258 (41.9%) | |
| 2 | 18 (2.9%) | |
| Disease type | Measurable disease | 310 (50.3%) |
The overall incidence of grade 3+ sensory neuropathy was 8.1% (50/616). The 566 patients who did not experience neuropathy were classified as having either completed treatment without neuropathy (3.9%) or categorized based on their reason for discontinuation: death/progression (41.1%), non-CIPN treatment terminating adverse event (30.2%) or withdrawal/other (16.7%) (Supplementary Table 1). The risk of neuropathy was not significantly different in the bevacizumab and placebo arms (p=0.11), which were pooled for analysis.
GWAS analysis was performed on the cumulative docetaxel dose (mg/m2) at first report of treatment-induced grade 3+ sensory peripheral neuropathy (Figure 1). The ten most significant SNPs, ranked according to the unadjusted p-value for cause-specific association with neuropathy, are listed in Table 2 with rsID, gene annotation, and hazard ratios before and after adjustment (Top 1000 SNPs in Supplementary Table 2). One SNP surpassed Bonferonni-corrected significance (0.05/498,022=1.004×10-7). This is an intronic SNP in the VAC14 gene (rs875858, minor allele frequency=0.056), which increased neuropathy risk (HR=3.60, 95%CI: 2.21-5.84, p=2.12×10-8, Figure 2A). The second and third most strongly associated SNPs were an intergenic SNP (rs11017056, MAF=0.223, HR=2.61, 95%CI: 1.77-3.84, p=3.84×10-7) and an intronic SNP in the ATP8A2 gene (rs1326116, MAF=0.194, HR=2.77, 95%CI: 1.79-4.27, p=1.77×10-6), both of which increased neuropathy risk (Figures 2B and 2C).
Figure 1. Genome-wide association results.
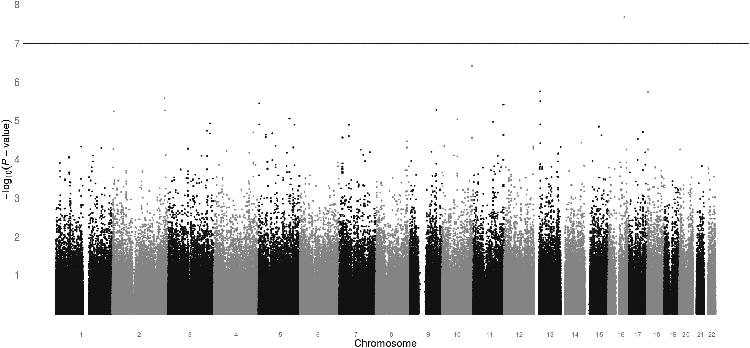
Manhattan plot displaying the unadjusted p-value for the association of cumulative docetaxel dose at grade 3+ sensory neuropathy for all SNPs that passed quality control (n=498,022). One SNP (rs875858) surpassed the Bonferonni-corrected significance threshold of 1.0×10-7.
Table 2. Ten SNPs Most Associated with Neuropathy in GWAS.
| rsID | Chr | Position | Gene | MAF | P-value | HR | 95% CI | Adjusted* | |
|---|---|---|---|---|---|---|---|---|---|
| P-value | HR | ||||||||
| rs875858 | 16 | 69332956 | VAC14 | 0.056 | 2.12E-08 | 3.60 | 2.21-5.84 | 5.88E-07 | 3.85 |
| rs11017056 | 10 | 131720630 | Intergenic | 0.223 | 3.84E-07 | 2.61 | 1.77-3.84 | 6.36E-07 | 2.58 |
| rs1326116 | 13 | 25273046 | ATP8A2 | 0.194 | 1.77E-06 | 2.77 | 1.79-4.27 | 4.57E-06 | 2.99 |
| rs8089250 | 18 | 4762807 | Intergenic | 0.175 | 1.83E-06 | 2.72 | 1.78-4.17 | 2.51E-05 | 2.49 |
| rs6436559 | 2 | 226061042 | KIAA1486 | 0.063 | 2.62E-06 | 3.34 | 1.95-5.71 | 3.26E-05 | 3.19 |
| rs11840337 | 13 | 25287298 | ATP8A2 | 0.191 | 3.16E-06 | 2.71 | 1.75-4.18 | 9.26E-06 | 2.90 |
| rs1566691 | 5 | 3780179 | Intergenic | 0.057 | 3.63E-06 | 3.64 | 2.03-6.52 | 4.45E-05 | 3.41 |
| rs1027796 | 11 | 132262880 | OPCML | 0.305 | 3.90E-06 | 2.38 | 1.62-3.48 | 2.41E-05 | 2.32 |
| rs2213920 | 9 | 117311405 | Intergenic | 0.128 | 5.26E-06 | 2.63 | 1.70-4.06 | 2.11E-05 | 2.66 |
| rs4438470 | 2 | 228722963 | SPHKAP | 0.137 | 5.47E-06 | 2.71 | 1.73-4.25 | 7.79E-05 | 2.47 |
95% CI: 95% Confidence interval around hazard ratio estimate
Chr: Chromosome
HR: Hazard ratio
MAF: minor allele frequency in the 90401 Caucasian cohort
Adjusted for patient BMI, age, history of diabetes, and treatment arm.
Figure 2. Incidence of Neuropathy by Genotype for Highly Associated SNPs.
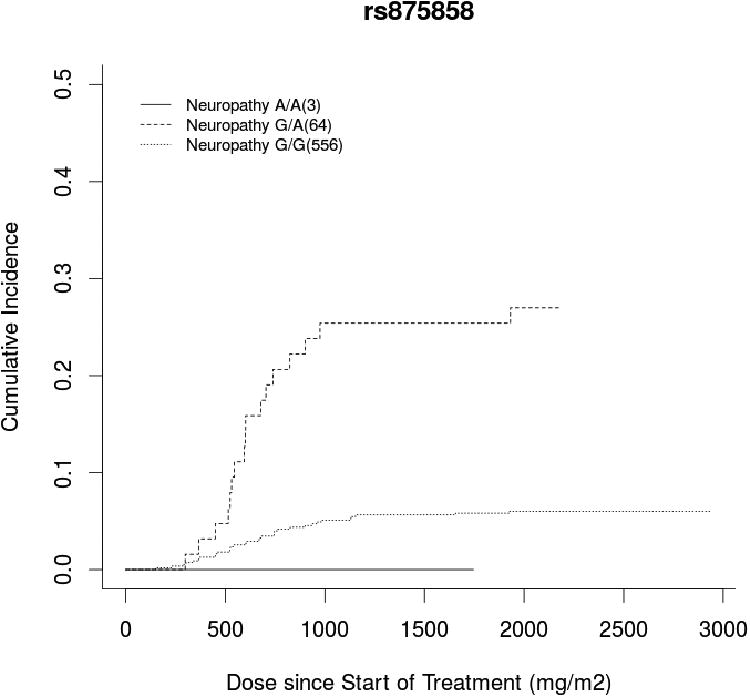
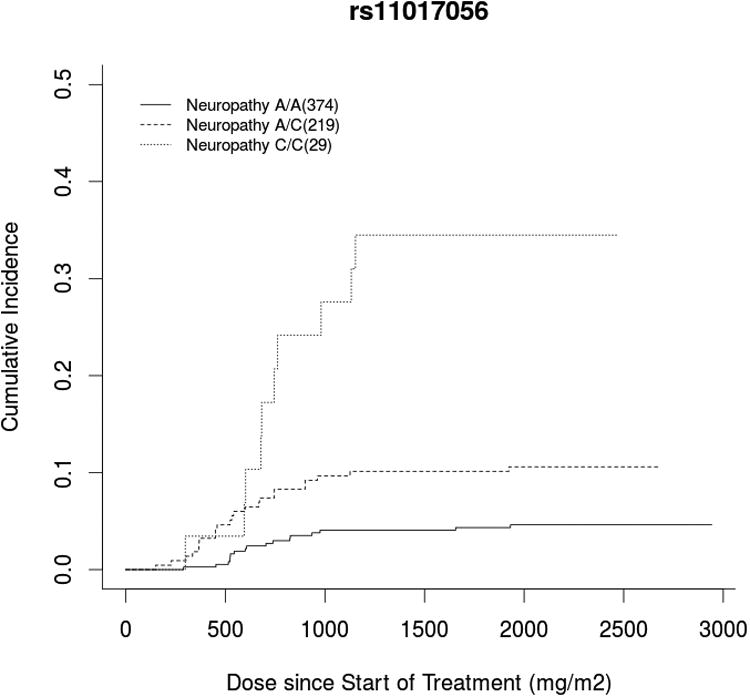
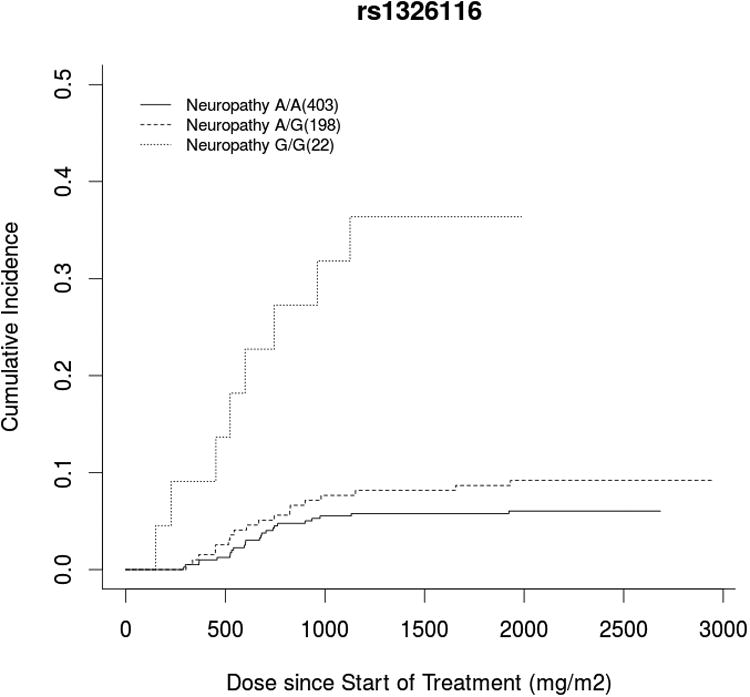
Cumulative incidence curve of docetaxel dose at grade 3+ neuropathy occurrence stratified by genotype for the top three hits: A) a VAC14 intronic SNP (rs875858), B) an intergenic SNP (rs11017056), and C) an ATP8A2 intronic SNP (rs1326116). In each case, carrying the variant allele increased neuropathy risk compared to wild-type patients, though there were too few patients who were homozygous for the rare VAC14 variant (n=3) to draw meaningful conclusions in this group.
The associations of all SNPs were then adjusted for treatment arm and several clinical covariates previously reported to be relevant to neuropathy risk including diabetes, age, and BMI. In this particular cohort, among the clinical covariates only increasing age (p=0.0003) was significantly associated with neuropathy, though increasing BMI showed a trend toward association (p=0.05). Covariate adjustment did not substantially affect the results, though the association of VAC14 fell below the Bonferonni-corrected significance threshold after adjustment (p=5.88 ×10-7). The top 1000 SNPs mapped to 240 unique genes, which were included in the pathway analysis. The three most significantly enriched pathways in the GWAS are displayed in Supplementary Table 3. Notably, two of these enriched pathways are relevant to CIPN; Axonal Guidance (p=1.20×10-4) and Neuropathic Pain Signaling in Dorsal Horn Neurons (p=6.03×10-3).
Cross-GWAS Pharmacogenetic Replication
Due to the absence of a large cohort of docetaxel-treated patients with prostate cancer in which to perform replication, and our interest in assessing whether taxanes share genetic predictors of neuropathy risk, cross-study pharmacogenetic replication was attempted in a cohort of paclitaxel-treated Caucasian patients with breast cancer enrolled on CALGB (Alliance 40101). The VAC14 SNP (rs875858) discovered in our docetaxel-induced neuropathy GWAS was not associated with paclitaxel-induced neuropathy in the 40101 study (p=0.70)(9). In addition, SNPs that were discovered in previously published GWAS of paclitaxel-induced neuropathy (FGD4 (rs10771973) p=0.39, EPHA4 (rs17348202) p=0.76, EPHA5 (rs7349683) p=0.15, intergenic (rs3125923) p=0.057, FCAMR (rs1856746) p=0.25) (9, 10, 12) or candidate SNP studies of docetaxel-induced neuropathy (GSTP1 Ala114Val (rs1138272) p=0.98, ABCB1 1236C>T (rs1128503) p=0.62, ABCB1 (rs4148738) p=0.90, ABCB1 3435C>T (rs1045642) p=0.80) (13-15) were not associated with docetaxel-induced neuropathy occurrence in the CALGB 90401 patients. Finally, when comparing the top SNPs from taxane-induced neuropathy GWAS performed within CALGB 90401 (docetaxel) and 40101 (paclitaxel), there was no statistical evidence that the degree of overlap was greater than would be expected by random chance (data not shown).
Peripheral Neuronal Cell Sensitivity
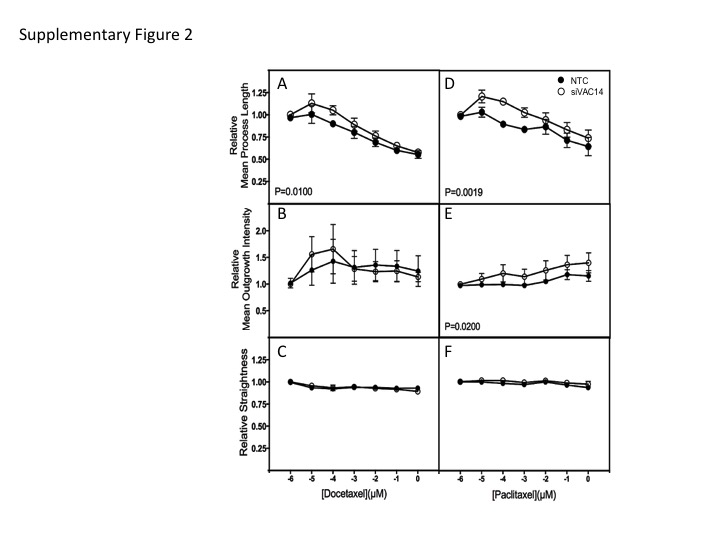
To determine the effect of VAC14 knockdown on neuronal sensitivity to docetaxel and paclitaxel, commercially available human iPSC-derived peripheral neurons (Peri4.U) were utilized. At the time of image analysis, which is 24 and 48 hours post drug treatment (48 and 72 h post transfection), expression of VAC14 in neurons treated with 1 μM siRNA had decreased to 54 and 44%, respectively, as compared to control cells treated with a non-targeting siRNA (Figure 3B). The VAC14 siRNA knockdown at 24 hours post-docetaxel treatment did not affect relative total outgrowth (Figure 3C) in the peripheral neurons but resulted in significantly greater sensitivity to docetaxel as measured by relative number of processes (p=0.0015, Figure 3D) and relative number of branches (p<0.0001, Figure 3E) Similar effects were found in preliminary studies of VAC14 knockdown in a human IPSC-derived cortical neuron, iCell® Neurons measured 48 hours after treatment (Cellular Dynamics International, Madison, WI) (data not shown). In contrast, VAC14 siRNA knockdown significantly decreased peripheral neuronal cell sensitivity to paclitaxel as measured by these same cellular phenotypes (all p<0.05, Figure 3F-3H). Other morphological characteristics including mean process length, mean outgrowth intensity and relative straightness were also measured for docetaxel and paclitaxel (Supplementary Figure 2). We found a decreased sensitivity to docetaxel and paclitaxel when measuring mean process length and for mean outgrowth intensity for paclitaxel only. There was no difference in measurements of relative straightness (data not shown).
Mouse Mechanical Sensitivity
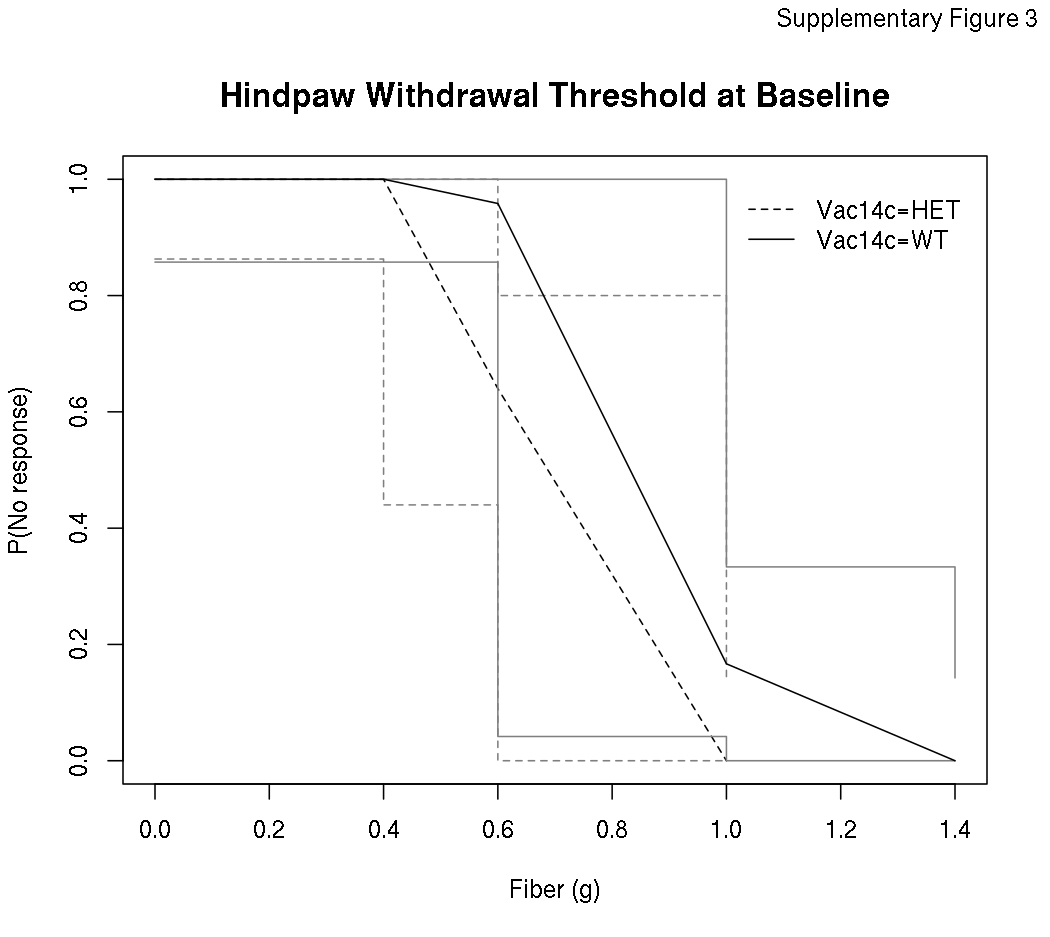
Prior to receiving docetaxel treatment, VAC14+/- heterozygous mice were more sensitive than wild type VAC14+/+ mice to mechanical stimuli using von Frey filaments (p=0.001, Supplementary Figure 3). When stratified by gender this effect was significant in male mice (p=0.002) but not female mice (p=0.16), however, there was no statistically significant gender interaction (p=0.10). The difference in sensitivity to mechanical stimuli between genotype groups at baseline was attenuated over the course of the experiment in untreated mice, as there was no evidence of a difference between these groups from at the end of the experiment (p=0.23). As expected, 8 days of docetaxel treatment increased sensitivity to mechanical stimuli compared to untreated control mice, regardless of mouse genotype (p<0.0001, Figure 4). Comparing across genotype groups, there was no difference between VAC14+/- and VAC14+/+ mice in docetaxel-induced increase in threshold sensitivity to mechanical stimuli at day 8 (p=0.18) or day 14 (p=0.80) when the neuropathy was at its maximum.
Figure 4. Docetaxel Treatment Increases Nociceptive Sensitivity in VAC14 Heterozygous and Wild-type Mice.
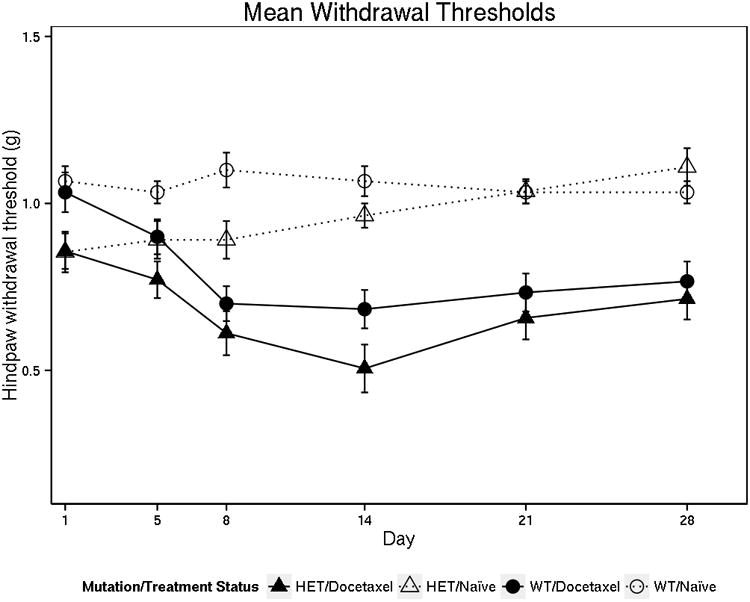
The withdrawal threshold is defined as the lightest fiber weight that caused each mouse to withdraw their hind paw at least three times out of five. Each mouse has a withdrawal threshold at each time-point in the study. The mean withdrawal threshold is the average of 12-14 mice (6-7 for each gender) for each treatment condition (VAC14 genotype and docetaxel treatment). Docetaxel- treated mice of either genotype had greater hindpaw withdrawal sensitivity following the end of treatment at day 8 compared to the untreated control mice (p<0.0001). There was no significant difference in the docetaxel-induced increase in withdrawal sensitivity comparing VAC14 heterozygous and wild-type mice (p=0.18).
Discussion
GWAS in a large cohort of mCRPC patients receiving docetaxel treatment identified an intronic SNP (rs875858) within VAC14 that was associated with increase neuropathy risk (HR=3.60). VAC14 has been directly linked to neurodegeneration in animal models(20) and linked to hereditary neuropathy conditions in humans(28). The lack of clinical replication in a cohort of paclitaxel-treated breast cancer patients is perhaps due to known differences between the two taxanes, including neuropathy profile(29). Distinct genetic modifiers of cellular sensitivity to paclitaxel and docetaxel have been previously reported in in vitro models(30) and the results of cellular sensitivity experiments demonstrating that VAC14 knockdown enhances sensitivity of peripheral neurons to docetaxel but not paclitaxel for relative number of processes and relative branching, further support this hypothesis. Given the distinct genetic effects on neuronal sensitivity to docetaxel and paclitaxel it is conceivable that the polymorphisms that affect neuropathy risk also differ between the two taxanes. This would be consistent with the lack of cross-study replication, including the lack of association with docetaxel-induced neuropathy occurrence in this analysis for SNPs (FGD4, EPHA4 and EPHA5) that have been previously discovered, and in some cases replicated, in paclitaxel-induced neuropathy GWAS (9, 10, 12, 31). Alternatively, heterogeneity between the patient cohorts, including differences in gender, age, and functional status, or differences in the phenotype (grade 2+ vs. grade 3+), may explain the lack of replication across the clinical GWAS. This GWAS was also not able to replicate previously reported associations between docetaxel-induced neuropathy and SNPs in ABCB1(15) or GSTP1(13, 14) reported in previous candidate SNP studies, similar to the failure of paclitaxel GWAS to replicate associations with candidate SNPs including CYP2C8*3(32).
The mechanistic importance of VAC14 in docetaxel-induced neuropathy was tested in two pre-clinical models of CIPN. Knockdown of VAC14 in the peripheral neuronal cell model (Peri.4U) increased sensitivity to docetaxel treatment as measured by decreased neurite number of processes and neurite branching. Interestingly, knockdown of VAC14 seemed to decrease cellular sensitivity to paclitaxel treatment for these morphological characteristics suggesting that this gene may play an important role in the damage to processes and branching induced by docetaxel but not paclitaxel. A similar model of human neuronal cell sensitivity that utilizes induced pluripotent cortical neurons (iCell® Neurons) has previously been used for mechanistic validation of GWAS hits that increases risk of paclitaxel-induced neuropathy(19, 33) and vincristine-induced neuropathy(11). These human model systems have several advantages over the use of rodent cell lines such as rat pheochromocytoma cell lines, PC12 or NS1. One major limitation of rodent cell lines is the required treatment with nerve growth factor to initiate nerve outgrowth, which confounds the evaluation of drug treatment effects(34-36).
To our knowledge this is the first study to attempt mechanistic validation of a finding from a clinical GWAS in an animal model of CIPN. The Von Frey Filament Test is a well-established rodent model to test sensitivity to nociceptive stimuli, as a surrogate for peripheral neuropathy, that is commonly used for mechanistic investigation of neuropathy(21, 37, 38). Unlike the cell experiment, VAC14 heterozygous mice were not more sensitive than wild-type littermates to the effect of docetaxel, but were more sensitive to mechanical stimuli prior to treatment. The enhanced sensitivity in the heterozygous mice at baseline was attenuated over time in the untreated cohort, in both male and female mice. It is possible that these mice acclimate to the discomfort of the mechanical stimuli or they may be experiencing numbness, another characteristic symptom of peripheral neuropathy in taxane-treated patients, though additional experiments would be required to test either hypothesis. It is unclear from these findings whether patients with VAC14 polymorphisms have subclinical neuropathy at baseline that is expressed by treatment, as suggested by the mouse data, or whether they are similar at baseline and have greater neuropathy development from docetaxel treatment. Subjective testing that is sensitive for subclinical neuropathy, perhaps with electrophysiological or quantitative sensory assessment(39), in untreated patients with wild-type and polymorphic VAC14 may help resolve this conundrum.
The VAC14 protein provides a scaffold for the lipid kinase PIKfyve and its positive regulator, FIG4. These proteins reside in a complex that is responsible for the generation of the signaling lipids phosphatidylinositol (3,5)-bisphosphate (PI(3,5)P2) and PI5P. These lipids are essential in multiple tissues(28). The inability of VAC14-/- homozygous mice to survive beyond 1-2 days after birth necessitated the use of VAC14+/- heterozygous mice(20). On autopsy AC14-/- homozygous mice exhibit massive neurodegeneration with cell bodies in the central and peripheral nervous system containing large vacuoles. This neurodegeneration and vacuolization are similar to that found in FIG4 null mice, suggesting that variation in these genes is likely to phenocopy in humans as well as in mice(40). A rare mutation in the FIG4 gene is known to cause a subtype of Charcot-Marie-Tooth disease, a hereditary neuropathy syndrome(41). Interestingly, the CALGB 40101 paclitaxel-induced neuropathy GWAS discovered and replicated an association for a SNP in FGD4 (rs10771973), another gene known to cause Charcot-Marie-Tooth disease(42), though this variant was not associated with docetaxel-induced neuropathy in our CALGB 90401 cohort.
The SNP (rs875858) discovered in this GWAS lies within an intron of the VAC14 gene. It is unclear what effect, if any, this individual SNP has on VAC14 expression or activity in the peripheral neuron. In the GTEx database there is no association between rs875858 and VAC14 gene expression in the tibial nerve tissue, though the power of this analysis is limited by the relatively small number of samples (n=256) and low minor allele frequency (MAF=0.06) of this SNP http://www.gtexportal.org/home/eqtls/bySnp?snpId=rs875858&tissueName=Nerve_Tibial. In the larger DGN cohort (n=922), rs875858 is highly associated with IL34 (p=4e-7) expression in whole blood. IL-34 is structurally similar to colony-stimulating factor 1 and both are ligands of the colony-stimulating factor 1 receptor(43), which has recently been identified as an efficacious target in mouse models of Charcot-Marie-Tooth disease(44), providing an alternative plausible biological mechanism for this association. Finally, it is conceivable that this SNP is not functionally relevant but instead is a genetic marker of a different causative SNP; however, rs875858 lies in a region of low linkage disequilibrium.
The SNP (rs1326116) with the third strongest association with docetaxel-induced neuropathy is an intronic polymorphism in the ATP8A2 gene, which encodes ATP8A2, a P4 ATPase. The P-type ATPases are a superfamily of proteins that translocate phospholipids between the two leaflets of cellular lipid bilayers(45). Several lines of evidence link ATP8A2 to neurological development and function; in vitro data that ATP8A2 overexpression enhances neurite outgrowth(46), animal data that ATP8A2 mutations cause overt progressive neurodegeneration in Wabbler-lethal mice by disrupting axonal transport and polarity(47), and case reports of ATP8A2 mutations causing severe neurological phenotypes in patients(48, 49).
The three pathways most highly enriched in the GWAS results include axonal guidance and neuropathic pain signaling, both of which have obvious relevance to taxane-induced peripheral neuropathy. Growth and maintenance of neuronal axons requires organization of microtubules into a cytoskeleton(50). Taxanes stabilize microtubules, potentially interfering with cytoskeletal organization leading to the overt neuropathic symptoms reported by patients. Enrichment in a similar pathway, axonal outgrowth, was reported in a secondary analysis of a paclitaxel-induced neuropathy GWAS, suggesting that these two studies are identifying similar genetic signatures(51), despite the lack of overlap of individual SNPs in the top 1000 hits. Neuropathic pain is one of the characteristic clinical manifestations of taxane-induced neuropathy, in addition to sensory and motor components(2). Neuropathic pain signaling in the dorsal horn neurons may contribute to taxane-induced peripheral neuropathy via a higher propensity for development of central sensitization(52).
The primary limitation of this study is the lack of availability of an independent clinical trial cohort of docetaxel-treated patients for pharmacogenetic replication. Another limitation is the reliance on the protocol-specified collection of severe peripheral neuropathy (grade 3 or higher), rather than systematic collection across all toxicity grades. It is possible that some patients experienced grade 2 neuropathy without progressing to grade 3, and thus were not included as neuropathy events in this analysis. Furthermore, ordinal analysis across all neuropathy grades (0-4) would have brought greater statistical power and may have identified a distinct set of SNPs that predict neuropathy severity, similar to the paclitaxel-induced neuropathy GWAS findings in CALGB (Alliance) 40101(9). The lack of grade 1 or 2 neuropathy data collection may explain the absence of events in the patients homozygous for the rs875858 SNP. However, two of these patients discontinued docetaxel treatment quite early, after 6 (442 mg/m2) and 8 (561 mg/m2) cycles for progression and grade 3 nausea/vomiting, respectively. The third patient was discontinued after 26 cycles (1,746 mg/m2) for a constellation of grade 3 toxicities including cough, dyspnea, fatigue, febrile neutropenia, and pneumonitis, without any documentation of peripheral neuropathy during treatment. The collection of DNA and comprehensive baseline and outcomes data, including accurate assessment of timing and severity of peripheral neuropathy, in other large clinical cohorts will allow for independent replication of our findings.
In conclusion, GWAS of a prospectively enrolled patient cohort identified a SNP in VAC14 that predicts sensitivity to docetaxel-induced peripheral neuropathy. VAC14 heterozygous mice were subsequently shown to be more sensitive to nociceptive stimuli at baseline, and peripheral neuronal cell (Peri.4U) sensitivity to docetaxel is enhanced after VAC14 knockdown, providing mechanistic validation of the relevance of VAC14 to docetaxel-induced neuropathy. However, the relatively low hazard ratio of this SNP suggests that no single variant or gene is responsible for dictating neuropathy sensitivity or will be clinically useful for personalizing docetaxel treatment. This is consistent with the multifactorial nature of neuropathy that depends on both drug exposure and a variety of patient factors, including age and BMI, which were associated with neuropathy occurrence in this cohort. Replication of the influence of our top candidate SNPs on neuropathy risk in a similar clinical trial cohort of docetaxel-treated mCRPC patients is of great interest to validate the role of these genes and provide support for further investigation into this mechanism for chemotherapy-induced neuropathy prevention or treatment.
Supplementary Material
{kind=link}
{kind=link}
{kind=link}
Statement of Translational Relevance.
The taxanes are a class of chemotherapeutic agents that are commonly associated with treatment-limiting peripheral sensory neuropathy. Genetic predictors of paclitaxel-induced neuropathy have been discovered but little is known about whether similar genetic factors are associated with risk of docetaxel-induced neuropathy. Discovery of genetic predictors of docetaxel-induced neuropathy could inform personalized treatment decisions to maximize efficacy and avoid unnecessary toxicity. We conducted a genome-wide association study in a large cohort of Caucasian men with hormone refractory prostate cancer treated with docetaxel. One single nucleotide polymorphism in VAC14 that was significantly associated with docetaxel-induced neuropathy was then mechanistically validated in cellular and animal models of neurotoxicity. These finding suggest that distinct genetic variants influence risk of peripheral neuropathy from paclitaxel and docetaxel. The genetic variant identified in this study could be useful for understanding the mechanism of docetaxel-induced neuropathy and may be informative for avoiding docetaxel treatment in patients at elevated neuropathy risk.
Acknowledgments
Susan Halabi received research funding from Sanofi. Mark Ratain has served as a paid consultant for Sanofi-Aventis and Genentech. Howard McLeod serves on the Scientific Advisory Board for Cancer Genetics Inc. M. Eileen Dolan has received research funding from Insys Therapeutics. Phillip Febbo received honoraria from Sanofi and is now an employee of Genomic Health.
Support: This study was supported by the NIH National Institute of General Medical Sciences Grant U01 GM61390, NIH/National Institute of General Medical Sciences (NIGMS) Pharmacogenomics of Anticancer Agents grant U01GM61393, NIH/National Institute of Nursing Research (NINR) P30NR014129, ALLIANCE grants (CA31946 and CA33601) and ECOG-ACRIN grants (CA21115 and CA16116). This work is part of the NIH PGRN-RIKEN Center for Genomic Medicine Global Alliance. Daniel Hertz was an American Foundation for Pharmaceutical Education Pre-Doctoral Fellow in Clinical Pharmaceutical Science when working on this analysis. Also supported in part by funds from U10CA031983, U10CA060138, U10CA047577, U10CA180838, U10CA180857 and Genentech. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
Footnotes
Conflict of Interest: The authors declare the following relevant conflicts of interest.
Previous Reports: This study was presented in part at the 2013 American Society of Clinical Oncology Annual Meeting. This report contains original material that has not been previously reported.
References
- 1.Manfredi JJ, Parness J, Horwitz SB. Taxol binds to cellular microtubules. J Cell Biol. 1982 Sep;94(3):688–96. doi: 10.1083/jcb.94.3.688. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Lee JJ, Swain SM. Peripheral Neuropathy Induced by Microtubule-Stabilizing Agents. J Clin Oncol. 2006 Apr 01;24(10):1633–42. doi: 10.1200/JCO.2005.04.0543. [DOI] [PubMed] [Google Scholar]
- 3.Postma TJ, Vermorken JB, Liefting AJM, Pinedo HM, Heimans JJ. Paclitaxel-induced neuropathy. Ann Oncol. 1995 May 1;6(5):489–94. doi: 10.1093/oxfordjournals.annonc.a059220. [DOI] [PubMed] [Google Scholar]
- 4.Mols F, Beijers T, Lemmens V, van den Hurk CJ, Vreugdenhil G, van de Poll-Franse LV. Chemotherapy-induced neuropathy and its association with quality of life among 2- to 11-year colorectal cancer survivors: results from the population-based PROFILES registry. J Clin Oncol. 2013 Jul 20;31(21):2699–707. doi: 10.1200/JCO.2013.49.1514. [DOI] [PubMed] [Google Scholar]
- 5.van Gerven JMA, Moll JWB, van den Bent MJ, Bontenbal M, van der Burg MEL, Verweij J, et al. Paclitaxel (Taxol) induces cumulative mild neurotoxicity. Eur J Cancer. 1994;30(8):1074–7. doi: 10.1016/0959-8049(94)90459-6. [DOI] [PubMed] [Google Scholar]
- 6.Mielke S, Sparreboom A, Steinberg SM, Gelderblom H, Unger C, Behringer D, et al. Association of Paclitaxel Pharmacokinetics with the Development of Peripheral Neuropathy in Patients with Advanced Cancer. Clin Cancer Res. 2005 Jul 1;11(13):4843–50. doi: 10.1158/1078-0432.CCR-05-0298. [DOI] [PubMed] [Google Scholar]
- 7.Akerley W, Herndon JE, Egorin MJ, Lyss AP, Kindler HL, Savarese DM, et al. Weekly, high-dose paclitaxel in advanced lung carcinoma. Cancer. 2003;97(10):2480–6. doi: 10.1002/cncr.11375. [DOI] [PubMed] [Google Scholar]
- 8.Rowinsky EK, Eisenhauer EA, Chaudhry V, Arbuck SG, Donehower RC. Clinical toxicities encountered with paclitaxel (Taxol) Semin Oncol. 1993 Aug;20(4 Suppl 3):1–15. [PubMed] [Google Scholar]
- 9.Baldwin RM, Owzar K, Zembutsu H, Chhibber A, Kubo M, Jiang C, et al. A genome-wide association study identifies novel loci for paclitaxel-induced sensory peripheral neuropathy in CALGB 40101. Clin Cancer Res. 2012 Sep 15;18(18):5099–109. doi: 10.1158/1078-0432.CCR-12-1590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Leandro-Garcia LJ, Inglada-Perez L, Pita G, Hjerpe E, Leskela S, Jara C, et al. Genome-wide association study identifies ephrin type A receptors implicated in paclitaxel induced peripheral sensory neuropathy. J Med Genet. 2013 Sep;50(9):599–605. doi: 10.1136/jmedgenet-2012-101466. [DOI] [PubMed] [Google Scholar]
- 11.Diouf B, Crews KR, Lew G, Pei D, Cheng C, Bao J, et al. Association of an inherited genetic variant with vincristine-related peripheral neuropathy in children with acute lymphoblastic leukemia. JAMA. 2015 Feb 24;313(8):815–23. doi: 10.1001/jama.2015.0894. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Schneider BP, Li L, Radovich M, Shen F, Miller KD, Flockhart DA, et al. Genome-Wide Association Studies for Taxane-Induced Peripheral Neuropathy in ECOG-5103 and ECOG-1199. Clin Cancer Res. 2015 Nov 15;21(22):5082–91. doi: 10.1158/1078-0432.CCR-15-0586. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Mir O, Alexandre J, Tran A, Durand J, Pons G, Treluyer J, et al. Relationship between GSTP1 Ile105Val polymorphism and docetaxel-induced peripheral neuropathy: clinical evidence of a role of oxidative stress in taxane toxicity. Ann Oncol. 2009 Apr 1;20(4):736–40. doi: 10.1093/annonc/mdn698. [DOI] [PubMed] [Google Scholar]
- 14.Eckhoff L, Feddersen S, Knoop AS, Ewertz M, Bergmann TK. Docetaxel-induced neuropathy: a pharmacogenetic case-control study of 150 women with early-stage breast cancer. Acta Oncol. 2015 Apr;54(4):530–7. doi: 10.3109/0284186X.2014.969846. [DOI] [PubMed] [Google Scholar]
- 15.Sissung TM, Baum CE, Deeken J, Price DK, Aragon-Ching J, Steinberg SM, et al. ABCB1 Genetic Variation Influences the Toxicity and Clinical Outcome of Patients with Androgen-Independent Prostate Cancer Treated with Docetaxel. Clin Cancer Res. 2008 Jul 15;14(14):4543–9. doi: 10.1158/1078-0432.CCR-07-4230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Kelly WK, Halabi S, Carducci M, George D, Mahoney JF, Stadler WM, et al. Randomized, Double-Blind, Placebo-Controlled Phase III Trial Comparing Docetaxel and Prednisone With or Without Bevacizumab in Men With Metastatic Castration-Resistant Prostate Cancer: CALGB 90401. J Clin Oncol. 2012 May 01;30(13):1534–40. doi: 10.1200/JCO.2011.39.4767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Fine JP, Gray RJ. A Proportional Hazards Model for the Subdistribution of a Competing Risk. JASA. 1999 Jun 01;94(446):496–509. [Google Scholar]
- 18.Prentice RL, Kalbfleisch JD, Peterson AV, Jr, Flournoy N, Farewell VT, Breslow NE. The analysis of failure times in the presence of competing risks. Biometrics. 1978 Dec;34(4):541–54. [PubMed] [Google Scholar]
- 19.Wheeler HE, Wing C, Delaney SM, Komatsu M, Dolan ME. Modeling chemotherapeutic neurotoxicity with human induced pluripotent stem cell-derived neuronal cells. PLoS One. 2015 Feb 17;10(2):e0118020. doi: 10.1371/journal.pone.0118020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Zhang Y, Zolov SN, Chow CY, Slutsky SG, Richardson SC, Piper RC, et al. Loss of Vac14, a regulator of the signaling lipid phosphatidylinositol 3,5-bisphosphate, results in neurodegeneration in mice. Proc Natl Acad Sci U S A. 2007 Oct 30;104(44):17518–23. doi: 10.1073/pnas.0702275104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Renn CL, Carozzi VA, Rhee P, Gallop D, Dorsey SG, Cavaletti G. Multimodal assessment of painful peripheral neuropathy induced by chronic oxaliplatin-based chemotherapy in mice. Mol Pain. 2011 Apr 26;7:29. doi: 10.1186/1744-8069-7-29. 8069-7-29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Shulman LN, Cirrincione CT, Berry DA, Becker HP, Perez EA, O'Regan R, et al. Six Cycles of Doxorubicin and Cyclophosphamide or Paclitaxel Are Not Superior to Four Cycles As Adjuvant Chemotherapy for Breast Cancer in Women With Zero to Three Positive Axillary Nodes: Cancer and Leukemia Group B 40101. J Clin Oncol. 2012 Nov 20;30(33):4071–6. doi: 10.1200/JCO.2011.40.6405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.R Development Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing; Vienna, Austria: 2008. URL http://www.R-project.org. [Google Scholar]
- 24.Therneau T, Grambsch P. Modeling Survival Data: Extending the Cox Model. New York, New York: Springer-Verlag; 2000. [Google Scholar]
- 25.Aulchenko YS, Ripke S, Isaacs A, van Duijn CM. GenABEL: an R library for genome-wide association analysis. Bioinformatics. 2007 May 15;23(10):1294–6. doi: 10.1093/bioinformatics/btm108. [DOI] [PubMed] [Google Scholar]
- 26.Filleron T, Laplanche A, Boher JM, Kramar A. An R function to non-parametric and piecewise analysis of competing risks survival data. Comput Methods Programs Biomed. 2010 Oct;100(1):24–38. doi: 10.1016/j.cmpb.2010.02.004. [DOI] [PubMed] [Google Scholar]
- 27.Fay MP, Shaw PA. Exact and Asymptotic Weighted Logrank Tests for Interval Censored Data: The interval R package. J Stat Softw. 2010 Aug;36(2):i02. doi: 10.18637/jss.v036.i02. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Zolov SN, Bridges D, Zhang Y, Lee W, Riehle E, Verma R, et al. In vivo, Pikfyve generates PI(3,5)P2, which serves as both a signaling lipid and the major precursor for PI5P. Proc Natl Acad Sci U S A. 2012 Oct 23;109(43):17472–7. doi: 10.1073/pnas.1203106109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Takemoto S, Ushijima K, Honda K, Wada H, Terada A, Imaishi H, et al. Precise evaluation of chemotherapy-induced peripheral neuropathy using the visual analogue scale: a quantitative and comparative analysis of neuropathy occurring with paclitaxel-carboplatin and docetaxel-carboplatin therapy. Int J Clin Oncol. 2012 Aug;17(4):367–72. doi: 10.1007/s10147-011-0303-6. [DOI] [PubMed] [Google Scholar]
- 30.Niu N, Schaid D, Abo R, Kalari K, Fridley B, Feng Q, et al. Genetic association with overall survival of taxane-treated lung cancer patients - A genome-wide association study in human lymphoblastoid cell lines followed by a clinical association study. BMC Cancer. 2012 Sep 24;12(1):422. doi: 10.1186/1471-2407-12-422. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Abraham JE, Guo Q, Dorling L, Tyrer J, Ingle S, Hardy R, et al. Replication of Genetic Polymorphisms Reported to be Associated with Taxane-Related Sensory Neuropathy in Early Breast Cancer Patients treated with Paclitaxel. Clinical Cancer Research. 2014 Mar 05; doi: 10.1158/1078-0432.CCR-13-3232. [DOI] [PubMed] [Google Scholar]
- 32.Hertz DL, Roy S, Motsinger-Reif AA, Drobish A, Clark LS, McLeod HL, et al. CYP2C8*3 increases risk of neuropathy in breast cancer patients treated with paclitaxel. Ann Oncol. 2013 Jun;24(6):1472–8. doi: 10.1093/annonc/mdt018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Komatsu M, Wheeler HE, Chung S, Low SK, Wing C, Delaney SM, et al. Pharmacoethnicity in Paclitaxel-Induced Sensory Peripheral Neuropathy. Clin Cancer Res. 2015 May 26; doi: 10.1158/1078-0432.CCR-15-0133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Rovini A, Carre M, Bordet T, Pruss RM, Braguer D. Olesoxime prevents microtubule-targeting drug neurotoxicity: selective preservation of EB comets in differentiated neuronal cells. Biochem Pharmacol. 2010 Sep 15;80(6):884–94. doi: 10.1016/j.bcp.2010.04.018. [DOI] [PubMed] [Google Scholar]
- 35.Verstappen CC, Postma TJ, Geldof AA, Heimans JJ. Amifostine protects against chemotherapy-induced neurotoxicity: an in vitro investigation. Anticancer Res. 2004 Jul-Aug;24(4):2337–41. [PubMed] [Google Scholar]
- 36.Kawashiri T, Egashira N, Itoh Y, Shimazoe T, Ikegami Y, Yano T, et al. Neurotropin reverses paclitaxel-induced neuropathy without affecting anti-tumour efficacy. Eur J Cancer. 2009 Jan;45(1):154–63. doi: 10.1016/j.ejca.2008.10.004. [DOI] [PubMed] [Google Scholar]
- 37.Hoke A, Ray M. Rodent models of chemotherapy-induced peripheral neuropathy. ILAR J. 2014;54(3):273–81. doi: 10.1093/ilar/ilt053. [DOI] [PubMed] [Google Scholar]
- 38.Chaplan SR, Bach FW, Pogrel JW, Chung JM, Yaksh TL. Quantitative assessment of tactile allodynia in the rat paw. J Neurosci Methods. 1994 Jul;53(1):55–63. doi: 10.1016/0165-0270(94)90144-9. [DOI] [PubMed] [Google Scholar]
- 39.Arezzo JC, Seto S, Schaumburg HH. Sensory-motor assessment in clinical research trials. Handb Clin Neurol. 2013;115:265–78. doi: 10.1016/B978-0-444-52902-2.00016-3. [DOI] [PubMed] [Google Scholar]
- 40.Lenk GM, Meisler MH. Mouse models of PI(3,5)P2 deficiency with impaired lysosome function. Methods Enzymol. 2014;534:245–60. doi: 10.1016/B978-0-12-397926-1.00014-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Lenk GM, Ferguson CJ, Chow CY, Jin N, Jones JM, Grant AE, et al. Pathogenic mechanism of the FIG4 mutation responsible for Charcot-Marie-Tooth disease CMT4J. PLoS Genet. 2011 Jun;7(6):e1002104. doi: 10.1371/journal.pgen.1002104. doi:10.1371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Delague V, Jacquier A, Hamadouche T, Poitelon Y, Baudot C, Boccaccio I, et al. Mutations in FGD4 encoding the Rho GDP/GTP exchange factor FRABIN cause autosomal recessive Charcot-Marie-Tooth type 4H. Am J Hum Genet. 2007 Jul;81(1):1–16. doi: 10.1086/518428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Nakamichi Y, Udagawa N, Takahashi N. IL-34 and CSF-1: similarities and differences. J Bone Miner Metab. 2013 Sep;31(5):486–95. doi: 10.1007/s00774-013-0476-3. [DOI] [PubMed] [Google Scholar]
- 44.Klein D, Patzko A, Schreiber D, van Hauwermeiren A, Baier M, Groh J, et al. Targeting the colony stimulating factor 1 receptor alleviates two forms of Charcot-Marie-Tooth disease in mice. Brain. 2015 Nov;138(Pt 11):3193–205. doi: 10.1093/brain/awv240. [DOI] [PubMed] [Google Scholar]
- 45.van der Mark VA, Elferink RP, Paulusma CC. P4 ATPases: Flippases in Health and Disease. Int J Mol Sci. 2013 Apr 11;14(4):7897–922. doi: 10.3390/ijms14047897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Xu Q, Yang GY, Liu N, Xu P, Chen YL, Zhou Z, et al. P4-ATPase ATP8A2 acts in synergy with CDC50A to enhance neurite outgrowth. FEBS Lett. 2012 Jun 21;586(13):1803–12. doi: 10.1016/j.febslet.2012.05.018. [DOI] [PubMed] [Google Scholar]
- 47.Zhu X, Libby RT, de Vries WN, Smith RS, Wright DL, Bronson RT, et al. Mutations in a P-type ATPase gene cause axonal degeneration. PLoS Genet. 2012;8(8):e1002853. doi: 10.1371/journal.pgen.1002853. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Cacciagli P, Haddad MR, Mignon-Ravix C, El-Waly B, Moncla A, Missirian C, et al. Disruption of the ATP8A2 gene in a patient with a t(10;13) de novo balanced translocation and a severe neurological phenotype. Eur J Hum Genet. 2010 Dec;18(12):1360–3. doi: 10.1038/ejhg.2010.126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Onat OE, Gulsuner S, Bilguvar K, Nazli Basak A, Topaloglu H, Tan M, et al. Missense mutation in the ATPase, aminophospholipid transporter protein ATP8A2 is associated with cerebellar atrophy and quadrupedal locomotion. Eur J Hum Genet. 2013 Mar;21(3):281–5. doi: 10.1038/ejhg.2012.170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Prokop A. The intricate relationship between microtubules and their associated motor proteins during axon growth and maintenance. Neural Dev. 2013 Sep 8;8:17. doi: 10.1186/1749-8104-8-17. 8104-8-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Chhibber A, Mefford J, Stahl EA, Pendergrass SA, Baldwin RM, Owzar K, et al. Polygenic inheritance of paclitaxel-induced sensory peripheral neuropathy driven by axon outgrowth gene sets in CALGB 40101 (Alliance) Pharmacogenomics J. 2014 Aug;14(4):336–42. doi: 10.1038/tpj.2014.2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Woolf CJ. Central sensitization: implications for the diagnosis and treatment of pain. Pain. 2011 Mar;152(3 Suppl):S2–15. doi: 10.1016/j.pain.2010.09.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.