

Abstract
Purpose
Retinopathy of prematurity (ROP) is a vision-threatening disease associated with abnormal retinal vascular development. Proteins from the insulin-like growth factor pathway are related to ROP. However, there is a paucity of research on the role of other proteins in ROP. The aim of this study was to identify plasma proteins related to clinically significant ROP.
Methods
We measured 1121 plasma proteins in the early neonatal period in infants at risk for ROP using an aptamer-based proteomic technology. The primary aim of the study was to compare plasma protein concentrations in infants who did (n = 12) and did not (n = 23) subsequently develop clinically significant ROP using logistic regression. As a secondary aim, we examined patterns in the proteins across categories of clinically significant, low-grade, and no ROP groups.
Results
Lower levels of 16 proteins were associated with an increased risk of clinically significant ROP. In this group, superoxide dismutase (Mn), mitochondrial (MnSOD), and chordin-like protein 1 (CRDL1) were highly ranked. Other proteins in this group included: C-C motif chemokine 14 (HCC-1), prolactin, insulin-like growth factor-binding protein 7 (IGFBP-7), and eotaxin. Higher levels of 12 proteins were associated with a higher risk for ROP. Fibroblast growth factor 19 (FGF-19) was the top-ranked protein target followed by hepatocyte growth factor-like protein (MSP), luteinizing hormone (LH), cystatin M, plasminogen, and proprotein convertase subtilisin/kexin type 9 (PCSK9). We also noted different patterns in the trend of concentrations of proteins across the clinically significant, low-grade, and no ROP groups.
Conclusions
We discovered plasma proteins with novel associations with clinically significant ROP (MnSOD, CRDL1, PCSK9), proteins with links to established ROP signaling pathways (IGFBP-7), and proteins such as MnSOD that may be a target for future therapeutic interventions.
Keywords: retinopathy of prematurity, protein biomarkers, early neonatal period
Retinopathy of prematurity (ROP) is a vision-threatening disease associated with abnormal retinal vascular development1–3 and is a leading cause of childhood blindness.1,2,4 Risk factors for ROP include early gestational age at delivery, very low birth weight, and higher or fluctuating oxygen levels in the extrauterine environment.2 Moreover, the premature loss of the maternal–fetal interaction specifically, a lack of maternal growth factors, has been shown to play a role in ROP.2 Despite advances in our understanding of the pathogenesis of ROP,1 we still do not fully understand why some infants who have a similar pattern of risk factors develop ROP and others do not, suggesting that additional risk factors may be linked with ROP. Proteins from the insulin-like growth factor (IGF) pathway2,3 have been shown to be related to ROP; however, there is a paucity of research on the role of other protein pathways in ROP.
The objective of this study was to identify plasma proteins related to ROP using a biomarker discovery approach. We were specifically focused on clinically significant ROP. To address this objective we measured 1121 proteins in plasma samples from the early neonatal period using a multiplexed aptamer-based proteomic technology.5
Methods
This study is an analysis of data collected as part of a multisite Institutional Review Board–approved study to investigate the role of angiogenesis-related markers in preterm infants who developed bronchopulmonary dysplasia (BPD). The study is described elsewhere.6 The research followed the tenets of the Declaration of Helsinki. In brief, infants were enrolled into the study within a week of birth. Informed consent was obtained from the parents with explanation of the nature and possible consequences of the study. An EDTA–plasma tube was obtained. The plasma sample was centrifuged after phlebotomy, and the supernatant was removed, aliquoted, and placed in a freezer at −80°C. At the conclusion of the study, a proteomic analysis was conducted on an aliquot using the SOMAscan assay at the laboratories of SomaLogic, Inc. (Boulder, CO, USA).5
From this dataset, we included nonanomalous premature infants enrolled at Children's Hospital Colorado (CHCO) or the University of Colorado Hospital (UCH) who were born at <31 weeks gestation or with a birth weight less than 1500 g per 2013 ROP screening criteria.4 We merged these records (n = 84) to the corresponding record in our ROP registry7 containing the results of the ROP screening examinations on all infants who received care at CHCO or UCH (2006 to present).
Definition of ROP
Clinically significant (high grade) ROP was defined as type 1 or type 2 ROP. Type 1 ROP was defined as stage 1 or 2 ROP in zone I with plus disease, stage 3 in zone I with or without plus disease, or stage 2 or 3 ROP in zone II with plus disease. Type 2 ROP was defined as stage 1 or 2 ROP in zone I without plus disease, or stage 3 ROP in zone II with no plus disease. Low-grade ROP was defined as ROP not meeting type 1 or type 2 definitions.8 The screening of infants for ROP followed standard guidelines4,9 and was defined by the International Classification of Retinopathy of Prematurity.9 Ultimately, 12 cases of clinically significant ROP, 27 cases of low-grade ROP, and 23 controls with no ROP were included in the final analytic dataset.
SomaLogic Proteomic Technology and Analysis
The SOMAscan proteomic assay is described in detail elsewhere.5,10 In brief, a biological sample in each well of a 96-well plate was incubated with a mixture of the 1121 SOMAmer reagents. Two sequential bead-based immobilization and washing steps eliminated unbound or nonspecifically bound proteins and the unbound SOMAmer reagents, leaving only protein target-bound SOMAmer reagents. These remaining SOMAmer reagents were isolated, and each reagent was quantified simultaneously on a custom Agilent (Santa Clara, CA, USA) hybridization array. The amount of each SOMAmer measured was quantitatively proportional to the protein concentration in the original sample.
Statistical Analysis
Descriptive statistics were calculated and compared across groups using t-tests or χ2 tests for variables such as gestational age and birth weight at delivery, infant age at blood draw, sex, and select inflammation-related neonatal complications including culture-positive sepsis and necrotizing enterocolitis.
In the first stage of the analysis, we concentrated on infants with the extreme phenotype of ROP. Concentrations for each of 1121 proteins were log (base 2) transformed and compared between infants who did (n = 12) and did not develop clinically significant ROP (n = 23) using a logistic regression (P < 0.05). Comparisons were both unadjusted and adjusted for gestational age. Adjustments for multiple comparisons were not made given the exploratory focus of this study,11 but false discovery rate adjusted P values were calculated and are provided in the online supplement. In the second stage of the analysis, we compared levels of the highest-ranking proteins across infants with clinically significant, low-grade ROP (n = 27), and no ROP using an analysis of variance.
Results
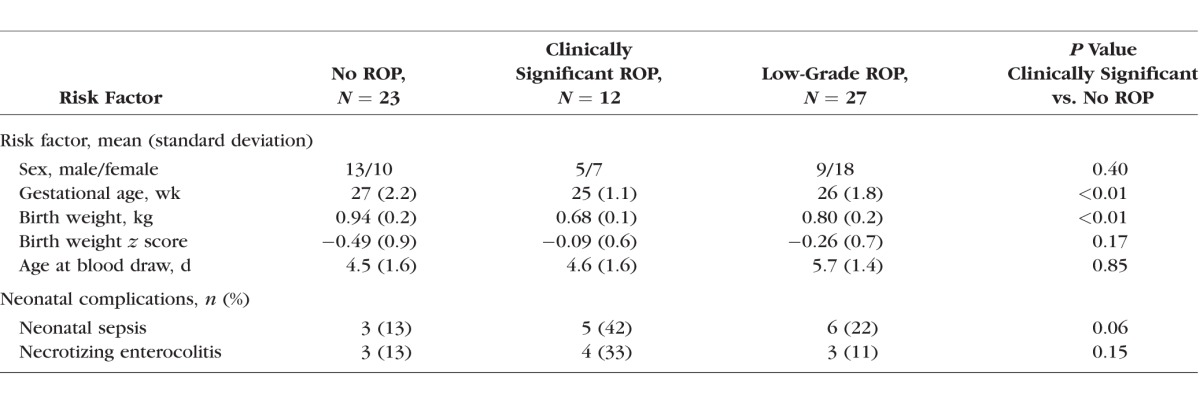
The overall characteristics of the cohort and the differences in select variables across the ROP groups are shown in Table 1. Gestational age and birth weight at delivery were significantly different between the infants across the ROP categories. However, the birth weight z score (birth weight adjusted for gestational age) was not significantly different across the ROP categories.
Table 1.
Differences in Select Risk Factors and Neonatal Complications Among Infants Who Developed Clinically Significant, Low-Grade, and No Retinopathy of Prematurity
In the first stage of the analysis, the concentrations of 1121 proteins were compared in infants who developed clinically significant ROP to infants who did not develop ROP. The significance (−log10 P value) and the magnitude (odds ratio, OR) for all the proteins significantly associated with ROP are shown in Table 2 and graphically in Figure 1. Select proteins are named in Figure 1 (see legend for details). The results for all proteins (n = 1121 proteins) included in the SOMAscan analysis are shown in Supplementary Table S1.
Table 2.
The Unadjusted and the Adjusted Association of Select Proteins* Measured Early in the Neonatal Period for Clinically Significant† ROP
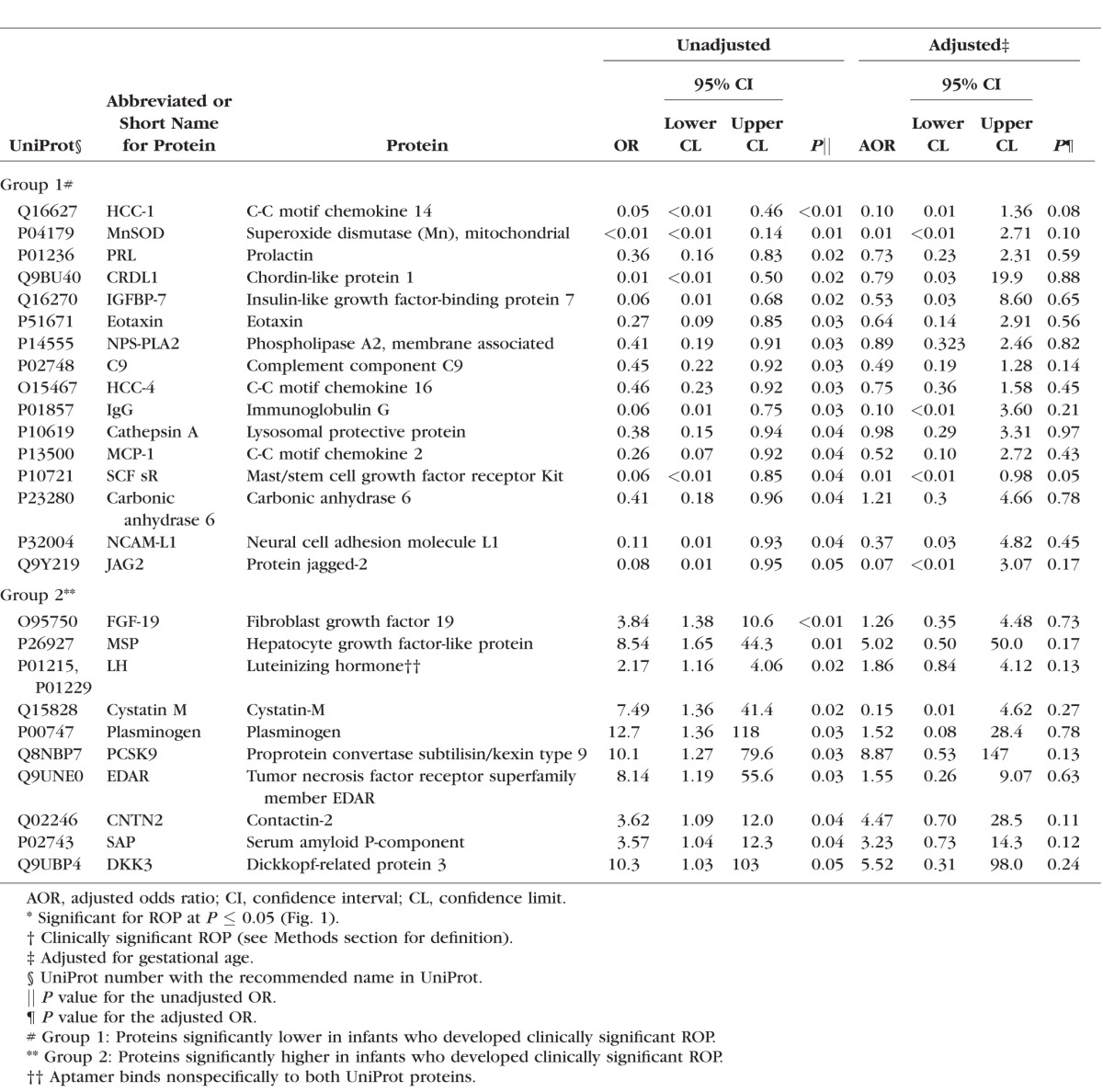
Figure 1.

Volcano plot displaying the results from the univariate tests comparing clinically significant ROP and no ROP for each protein. The significance (−log10 P value) and the magnitude (OR) for all the significant proteins are displayed graphically. Proteins with the largest magnitude of difference between groups and smallest P value are named in the figure. Proteins named in blue and in red were significantly lower and higher, respectively, in infants who developed ROP. Please see Table 2 for the full name of each protein.
We found that lower levels of 16 proteins were associated with a higher risk of an infant's developing clinically significant ROP (group 1, Table 2). Two of these proteins, superoxide dismutase (Mn), mitochondrial (MnSOD), and chordin-like protein 1 (CRDL1), were clearly set apart from the other proteins as seen in Figure 1. C-C motif chemokine 14 (HCC-1) was also a top-ranked protein in group 1. Additionally, prolactin (PRL), insulin-like growth factor-binding protein 7 (IGFBP-7), and eotaxin were similarly ranked high by P value and OR for ROP. The remaining proteins in group 1 (Table 2) were associated with a lower level of significance for ROP.
We found that higher levels of 12 proteins were associated with a higher risk for ROP (group 2, Table 2). Fibroblast growth factor 19 (FGF-19) was a top-ranked protein target. The next most highly ranked proteins were hepatocyte growth factor-like protein (MSP) and luteinizing hormone (LH). Cystatin M, plasminogen, and proprotein convertase subtilisin/kexin type 9 (PCSK9) had high unadjusted ORs for ROP associated with a lower rank. The remaining proteins in group 2 (Table 2) were associated with a further attenuation in the level of significance for ROP.
Adjusted for gestational age, HCC-1 and carbonic anhydrase 6 had a borderline association with ROP (Table 2). None of the other proteins were statistically significantly associated with ROP in the adjusted analysis (Table 2).
In the second stage of the analysis, we explored levels of the highly ranked proteins in infants who developed clinically significant, low-grade, or no ROP. We show the results in Table 3. The results are also shown in Figure 2 (box plots) for select proteins and for all of the highly ranked (Table 2) proteins in Supplementary Figure S1. We demonstrate in the figures (1) a gradient across the groups (e.g., HCC-1 and PRL); (2) a difference between the low-grade and clinically significant ROP groups (e.g., IGFBP-7 and MnSOD); and (3) a difference between the low-grade and no ROP groups (e.g., cystatin M and c-c-motif chemokine 2 [MCP-1]).
Table 3.
Concentrations of Select* Proteins in Infants With Clinically Significant, Low-Grade, and No ROP
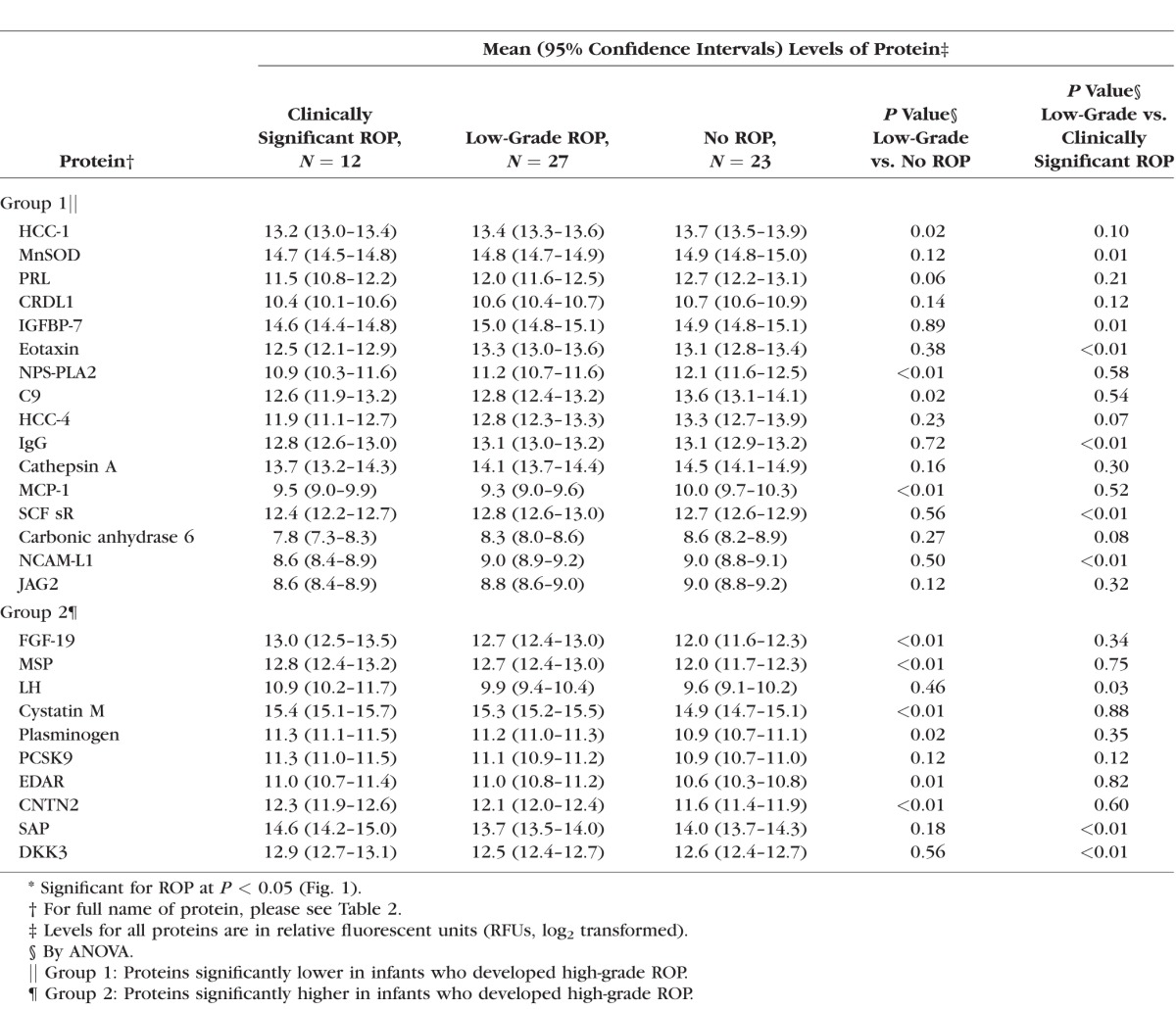
Figure 2.

Distribution of select proteins in infants with clinically significant, low-grade, and no ROP. The box indicates the IQR (25th–75th percentile), and the median and mean are indicated by the lines and diamonds, respectively. The whiskers indicate data within 1.5 times the IQR, and points are data outside 1.5 times the IQR. IQR, interquartile range; RFU, relative fluorescence unit; Clin Sig, clinically significant ROP.
Discussion
We used an innovative SOMAscan proteomic assay technology12 to determine the proteins from a blood sample in the first week of life that place an infant at a higher risk of developing clinically significant ROP. We studied over 1000 analytes simultaneously across many protein pathways.5 It is of interest that we found that lower levels of many proteins distinguished infants with and without clinically significant ROP. An advantage of using this aptamer-based technology is that concentrations of low-abundant proteins can be measured.5 Our study has initiated a new area of exploration of biomarkers for ROP. Indeed, we uncovered proteins early in the neonatal period that were novel in their association with clinically significant ROP (MnSOD, CRDL1, PCSK9), proteins with links to established signaling pathways for ROP (IGFBP-7),1,2 and some proteins such as MnSOD that may be a target for a future therapeutic interventions. We also noted different patterns in the trend of concentrations of proteins across the clinically significant, low-grade, and no ROP groups (Fig. 2). Aligned with the results of previous studies (reviewed in Ref. 2), we also found that infants at the lower extreme of gestational age and birth weight were at the highest risk of developing ROP.
Superoxide dismutase, mitochondrial (MnSOD), an antioxidant found in the mitochondria, was clearly distinguished from other proteins displayed in Figure 1: MnSOD has a role in scavenging oxygen radicals that result from oxidative stress and provides the cell with a powerful defense against the deleterious effect of elevated levels of reactive oxygen species (ROS) (reviewed in Ref. 13). An increased susceptibility to oxidative mitochondrial injury specifically in central nervous system neurons, cardiac myocytes, and other metabolically active tissues has been reported in a murine model of MnSOD deficiency, after postnatal exposure to ambient oxygen concentrations.14 Moreover, a role for oxidative stress has been proposed in ROP (reviewed in Ref. 1). Indeed, it has been demonstrated that an increase in SOD ameliorates oxygen-induced retinopathy in transgenic mice.15 Furthermore, supplementation with liposome-encapsulated SOD significantly increased retinal SOD activity and reduced oxygen-induced retinopathy in a rat model.16 The results of our study suggest that low levels of MnSOD may be an early neonatal marker of immaturity of the oxygen defense system that places the infant at risk for ROP.
Chordin-like protein 1, a bone morphogenic protein-4 (BMP-4) antagonist, is also a prominent protein as shown in Figure 1. Kane et al.17 investigated hypoxia inducible factor-1α-driven expression of CRDL1 in human pericytes and found that expression of CRDL1 and vascular endothelial growth factor (VEGF) was upregulated by hypoxia. The hypoxic state resulted in secretion of CRDL1 from the human pericytes. Chordin-like protein 1 complexed with BMP-4 to antagonize the antiangiogenic effects of BMP-4 on endothelial cells. This tipped the balance in favor of an angiogenic environment.17 We suggest that the low levels of CRDL1 observed in our study in the first week of life may contribute to an antiangiogenic state. Our suggestion would be consistent with the avascular state of the retina observed during the first phase of ROP. Indeed, ROP is understood to have two phases, possibly preceded by a prephase of antenatal sensitization via inflammation.2,3 These phases have links with dysregulation of both neuronal and vascular development of the retina.18,2,3 In phase 1 of ROP, there is a cessation in growth of the retinal blood vessels secondary to high oxygen levels that contribute to a downregulation in oxygen-regulated growth factors such as VEGF.2 In contrast, phase 2 of ROP is characterized by retinal neovascularization induced by hypoxia.2,19 During phase 2 of ROP, the compromised retinal blood vessels cannot supply enough oxygen to the developing retina. The metabolic demands of the retina increase, leading to increased hypoxia-driven local VEGF production and retinal neovascularization.2
In addition to hyperoxia, phase I of ROP is precipitated after a preterm delivery by the loss of factors normally provided by the mother in utero, including IGF-1, which promotes VEGF-mediated development of retinal blood vessels.2 In our study we found that lower levels of IGF-1 and VEGF placed an infant at risk for ROP, but neither of these relationships with ROP was statistically significant (Supplementary Table S1). However, another less well studied member of the IGF family of proteins, namely, IGFBP-7, emerged as important in our analysis. We found that levels of IGFBP-7 were lower in infants who subsequently developed ROP compared to infants with no ROP. Aligned with the results of our study, other authors have studied a murine model of oxygen-induced retinopathy and found that the IGFBP-7 gene was downregulated in expression in retinas removed from hyperoxia chambers when compared to retinas exposed to states of normal oxygen concentrations.20 Furthermore, other investigators have found ties between mutations of the IGFBP-7 gene with a human disease characterized by familial retinal artery macroaneurysms (FRAM) and supravalvular pulmonic stenosis, suggesting a role for IGFBP-7 in vasculogenesis and angiogenesis.21 Indeed, it is biologically plausible that the immaturity in levels of IGFBP-7 seen in our study may contribute to the abnormal vascular development seen in infants with ROP.
Other potentially interesting protein targets (Fig. 1) included PRL, a polypeptide hormone produced by the anterior pituitary that stimulates mammary gland development and lactation and has a role in angiogenesis and tumorigenesis.22 We also found lower levels of some proteins with ties to the inflammatory pathway, for example, HCC-123 and eotoxin.24
Several proteins found at elevated levels were associated with an increased likelihood of developing ROP (Fig. 1). Fibroblast growth factor 19, a member of the fibroblast growth factor family, is a key regulator of energy metabolism.25 Infants with elevated levels of MSP, a protein with links to inflammation, were over eight times more likely to develop ROP.26 Indeed, it is suggested that the presence of infection or inflammation may modify the risk of ROP.3 Lutenizing hormone, a hormone that stimulates the testes and ovaries to synthesize steroids, was also elevated in infants who developed clinically significant ROP.27 The significance of this finding is uncertain but perhaps represents an alteration in function of the hypothalamic–pituitary–gonadal axis. Other proteins identified in this group included cystatin-M, a cysteine protease inhibitor, which has a role in the process of desquamation or cell shedding,28 and plasminogen, the inactive precursor of the enzyme plasmin.29 Proprotein convertase subtilisin/kexin type 9 was discovered in 2003 and is distinguished as a target for pharmaceutical intervention to reduce low-density lipoprotein-cholesterol (LDL-C). Proprotein convertase subtilisin/kexin type 9 mutations are related to higher levels of LDL-C and an increased risk for cardiovascular disease.30 With definite links to vascular disease, this protein could potentially have a role in ROP.
There are some limitations to our study. The main limitation was the small sample size, which reduced our ability to conduct a more extensive analysis such as adequate adjustment for confounding variables, adjustment for multiple comparisons, and stratification by gestational age at delivery. The original study was powered for a different outcome (BPD) and utilized infants from additional sites for which we did not have ROP outcomes. As expected, given the small sample size, adjustment for gestational age resulted in the loss of statistical significance of our highly ranked proteins with ROP. We view gestational age as an outcome of pregnancy that is not modifiable once the infant is born. In contrast, the novel proteins discovered in this study are potential modifiable targets for therapeutic interventions in infants at risk for ROP. This was the focus of our study, and it is important to note that even with the small sample size, we uncovered novel proteins early in the neonatal period with a large magnitude of association related to the subsequent development of ROP. Although adjustment for gestational age affected statistical significance of risk for ROP, the magnitude of risk remained meaningful even if it was not statistically significant (Table 2). These observations are important and deserve future investigation in an adequately powered study of infants at risk for ROP to determine the independent relationship of gestational age and the individual proteins with ROP. An additional limitation was that we conducted the proteomic analysis only on a single sample from the early neonatal period. It will be important in future studies to characterize protein levels and profiles at the time of the ROP examination when the pathologic events in the retina are known to be different from those in the earlier phase of ROP.2
Notwithstanding these limitations, the results from this state-of-the-art proteomic analysis are informative, and we suggest validation of the results in a larger cohort of preterm infants. The findings of our pilot study related to the protein target MnSOD were especially compelling and deserve further investigation. Infants with lower levels of this protein may not be able to handle oxidative stress in the neonatal period. This result suggests that it is important to consider clinical interventions early in the neonatal period to attenuate the effects of oxygen stress in this vulnerable group of infants.
Supplementary Material
Acknowledgments
Supported by National Institutes of Health/National Heart, Lung, and Blood Institute R01 HL085703 (SHA).
Disclosure: A.M. Lynch, None; B.D. Wagner, None; N. Mandava, None; A.G. Palestine, None; P.M. Mourani, None; E.A. McCourt, None; S.C.N. Oliver, None; S.H. Abman None
References
- 1. Hartnett ME. Pathophysiology and mechanisms of severe retinopathy of prematurity. Ophthalmology. 2015; 122: 200–210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Smith LE,, Hard AL,, Hellstrom A. The biology of retinopathy of prematurity: how knowledge of pathogenesis guides treatment. Clin Perinatol. 2013; 40: 201–214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Hellstrom A,, Smith LE,, Dammann O. Retinopathy of prematurity. Lancet. 2013; 382: 1445–1457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Fierson WM. Screening examination of premature infants for retinopathy of prematurity. Pediatrics. 2013; 131: 189–195. [DOI] [PubMed] [Google Scholar]
- 5. Rohloff JC,, Gelinas AD,, Jarvis TC,, et al. Nucleic acid ligands with protein-like side chains: modified aptamers and their use as diagnostic and therapeutic agents. Mol Ther Nucleic Acids. 2014; 3: e201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Mourani PM,, Sontag MK,, Younoszai A,, et al. Early pulmonary vascular disease in preterm infants at risk for bronchopulmonary dysplasia. Am J Respir Crit Care Med. 2015; 191: 87–95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Cao JH,, Wagner BD,, McCourt EA,, et al. The Colorado-retinopathy of prematurity model (CO-ROP): postnatal weight gain screening algorithm. J AAPOS. 2016; 20: 19–24. [DOI] [PubMed] [Google Scholar]
- 8. Good WV. Final results of the Early Treatment for Retinopathy of Prematurity (ETROP) randomized trial. Trans Am Ophthalmol Soc. 2004; 102: 233–248 discussion 48–50. [PMC free article] [PubMed] [Google Scholar]
- 9. International Committee for the Classification of Retinopathy of Prematurity. The International Classification of Retinopathy of Prematurity revisited. Arch Ophthalmol. 2005; 123: 991–999. [DOI] [PubMed] [Google Scholar]
- 10. Lynch AM,, Wagner BD,, Deterding RR,, et al. The relationship of circulating proteins in early pregnancy with preterm birth. Am J Obstet Gynecol. 2016; 214: 517.e1–e8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Rothman KJ. No adjustments are needed for multiple comparisons. Epidemiology. 1990; 1: 43–46. [PubMed] [Google Scholar]
- 12. Ganz P,, Heidecker B,, Hveem K,, et al. Development and validation of a protein-based risk score for cardiovascular outcomes among patients with stable coronary heart disease. JAMA. 2016; 315: 2532–2541. [DOI] [PubMed] [Google Scholar]
- 13. Bresciani G,, da Cruz IB,, Gonzalez-Gallego J. Manganese superoxide dismutase and oxidative stress modulation. Adv Clin Chem. 2015; 68: 87–130. [DOI] [PubMed] [Google Scholar]
- 14. Lebovitz RM,, Zhang H,, Vogel H,, et al. Neurodegeneration, myocardial injury, and perinatal death in mitochondrial superoxide dismutase-deficient mice. Proc Natl Acad Sci U S A. 1996; 93: 9782–9787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Spierer A,, Rabinowitz R,, Pri-Chen S,, Rosner M. An increase in superoxide dismutase ameliorates oxygen-induced retinopathy in transgenic mice. Eye. 2005; 19: 86–91. [DOI] [PubMed] [Google Scholar]
- 16. Niesman MR,, Johnson KA,, Penn JS. Therapeutic effect of liposomal superoxide dismutase in an animal model of retinopathy of prematurity. Neurochem Res. 1997; 22: 597–605. [DOI] [PubMed] [Google Scholar]
- 17. Kane R,, Godson C,, O'Brien C. Chordin-like 1, a bone morphogenetic protein-4 antagonist, is upregulated by hypoxia in human retinal pericytes and plays a role in regulating angiogenesis. Mol Vis. 2008; 14: 1138–1148. [PMC free article] [PubMed] [Google Scholar]
- 18. Fulton AB,, Hansen RM,, Moskowitz A,, Akula JD. The neurovascular retina in retinopathy of prematurity. Prog Retin Eye Res. 2009; 28: 452–482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Smith LE. Pathogenesis of retinopathy of prematurity. Semin Neonatol. 2003; 8: 469–473. [DOI] [PubMed] [Google Scholar]
- 20. Ishikawa K,, Yoshida S,, Kadota K,, et al. Gene expression profile of hyperoxic and hypoxic retinas in a mouse model of oxygen-induced retinopathy. Invest Ophthalmol Vis Sci. 2010; 51: 4307–4319. [DOI] [PubMed] [Google Scholar]
- 21. Abu-Safieh L,, Abboud EB,, Alkuraya H,, et al. Mutation of IGFBP7 causes upregulation of BRAF/MEK/ERK pathway and familial retinal arterial macroaneurysms. Am J Hum Genet. 2011; 89: 313–319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Bernard V,, Young J,, Chanson P,, Binart N. New insights in prolactin: pathological implications. Nat Rev Endocrinol. 2015; 11: 265–275. [DOI] [PubMed] [Google Scholar]
- 23. Schulz-Knappe P,, Magert HJ,, Dewald B,, et al. HCC-1 a novel chemokine from human plasma. J Exp Med. 1996; 183: 295–299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Pease JE,, Williams TJ. Eotaxin and asthma. Curr Opin Pharmacol. 2001; 1: 248–253. [DOI] [PubMed] [Google Scholar]
- 25. Angelin B,, Larsson TE,, Rudling M. Circulating fibroblast growth factors as metabolic regulators--a critical appraisal. Cell Metab. 2012; 16: 693–705. [DOI] [PubMed] [Google Scholar]
- 26. Li J,, Chanda D,, Shiri-Sverdlov R,, Neumann D. MSP: an emerging player in metabolic syndrome. Cytokine Growth Factor Rev. 2015; 26: 75–82. [DOI] [PubMed] [Google Scholar]
- 27. Choi J,, Smitz J. Luteinizing hormone and human chorionic gonadotropin: distinguishing unique physiologic roles. Gynecol Endocrinol. 2014; 30: 174–181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Brocklehurst K,, Philpott MP. Cysteine proteases: mode of action and role in epidermal differentiation. Cell Tissue Res. 2013; 351: 237–244. [DOI] [PubMed] [Google Scholar]
- 29. Schuliga M. The inflammatory actions of coagulant and fibrinolytic proteases in disease. Mediators Inflamm. 2015; 2015: 437695. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Schulz R,, Schluter KD,, Laufs U. Molecular and cellular function of the proprotein convertase subtilisin/kexin type 9 (PCSK9). Basic Res Cardiol. 2015; 110: 4. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.