

Abstract
Delivering vaccine antigens to mucosal surfaces is potentially very attractive, especially as protection from mucosal infections may be mediated by local immune responses. However, to date mucosal immunization has had limited successes, with issues of both safety and poor immunogenicity. One approach to improve immunogenicity is to develop adjuvants that are effective and safe at mucosal surfaces. Differences in immune responses between mice and men have overstated the value of some experimental adjuvants which have subsequently performed poorly in the clinic. Due to their closer similarity, non-human primates can provide a more accurate picture of adjuvant performance. In this study we immunised rhesus macaques (Macaca mulatta) using a unique matrix experimental design that maximised the number of adjuvants screened while reducing the animal usage. Macaques were immunised by the intranasal, sublingual and intrarectal routes with the model protein antigens keyhole limpet haemocyanin (KLH), β-galactosidase (β-Gal) and ovalbumin (OVA) in combination with the experimental adjuvants Poly(I:C), Pam3CSK4, chitosan, Thymic Stromal Lymphopoietin (TSLP), MPLA and R848 (Resiquimod). Of the routes used, only intranasal immunization with KLH and R848 induced a detectable antibody response. When compared to intramuscular immunization, intranasal administration gave slightly lower levels of antigen specific antibody in the plasma, but enhanced local responses. Following intranasal delivery of R848, we observed a mildly inflammatory response, but no difference to the control. From this we conclude that R848 is able to boost antibody responses to mucosally delivered antigen, without causing excess local inflammation.
Keywords: adjuvant, inflammation, intranasal, Mucosal, Macaque, TLR ligand
Introduction
Mucosal immunization offers an attractive prospect for many of the more intractable infections for which we have yet to develop vaccines, especially those that infect via mucosal surfaces, such as HIV, RSV and tuberculosis. In principle, mucosal immunization may lead to responses at the sites of infection – either the respiratory or the genital tracts, improving protective efficacy.1 However, immune responses to mucosally delivered antigens are often limited for various reasons including biochemical and mechanical degradation of the antigen, or immune tolerance at mucosal sites.2 An important consideration for the mucosal administration of vaccines is the selection of the route of administration. While targeting the genital or rectal mucosa may theoretically induce more specific local responses, there are cultural, biomechanical and immunological reasons why these routes may not be the most effective. It has, however, been suggested that mucosal immunization at one site can induce responses at other distal mucosal sites.3 Specifically, immunological linkage between the upper respiratory tract and lower genital tract has been proposed, based predominantly on studies performed in mice.4
Another approach to overcome the poor immune response at mucosal sites is to develop effective mucosal adjuvants, reviewed by Newsted el al.5 To date there is no licensed adjuvant for mucosal use. Safety is of paramount importance as perturbations of the tightly regulated immune responses at mucosal surfaces can cause unwanted reactions.6 Likewise the proximity of the nervous system to mucosal surfaces can lead to complications such as Bell's palsy.7 A number of adjuvants have been suggested and tested in mouse studies.8 Many of the adjuvants that have been tested have been based upon agents that are known ligands for toll like receptors (TLR), a highly conserved family of pattern recognition receptors that activate the innate immune response. While the murine model is highly effective for the screening of compounds, there are limitations in the translation of these compounds from the mouse to humans. Non-human primates, because of their genetic proximity to man, larger size and the similarities in anatomy provide a more effective platform to confirm the efficacy of potential adjuvants. However, there are ethical considerations in the use of large numbers of these animals and approaches to reduce animal numbers are required.
In the current study we used a matrix design to compare mucosal immunization with 3 different antigens by the intranasal, sublingual or intrarectal routes in combination with 6 different experimental adjuvants Poly(I:C), Pam3CSK4, chitosan, Thymic Stromal Lymphopoietin (TSLP), MPLA and R848 (Resiquimod), in order to select the best adjuvant for future clinical trials. Of the combinations tested, only antigen delivered intranasally in combination with the TLR7/8 agonist R848 induced significant antibody responses. When the R848/ antigen combination was delivered by either the intramuscular or intranasal routes, intramuscular delivery induced greater systemic responses but intranasal delivery induced a slightly greater nasal response. In a separate study intranasal R848 gave a very similar cytokine and cell profile to intranasal PBS (control) except for a delayed TNF signal, suggesting that it is safe for intranasal delivery. In summary, R848 appears to be a highly effective in promoting local and systemic immune responses by the nasal route of administration.
Results
Intranasal but not other routes induce an immune response
Macaques (Macaca mulatta) were immunised by the intranasal, sublingual and intrarectal routes (Table 1) with the model protein antigens keyhole limpet haemocyanin (KLH), β-galactosidase (β-Gal) or ovalbumin (OVA) in combination with the experimental adjuvants Pam3CSK4, Poly(I:C), Chitosan, Thymic Stromal Lymphopoietin (TSLP), Monophosphoryl Lipid A (MPLA) or Resiquimod (R848). Adjuvant selection was based on preclinical studies that identified Pam3CSK4 (TLR1/2), chitosan and TSLP as effective adjuvants in mice.8,9 R848 was inefficient in the murine model but was seen to be effective in previous macaque studies10 and was therefore selected as a comparator. This study was designed as a preliminary study to select adjuvants for future clinical studies and therefore a range of experimental adjuvants targeting a broad spectrum of pattern recognition receptors were selected. To maximise the screening potential of this study, different adjuvants were used at different sites. Different antigens were used to allow the overlapping study of the different routes of delivery while reducing animal usage, the antigens selected are common model antigens.
Table 1.
Dosing schedule
| WK −4 | WK 0 | WK 1 | WK 2 | WK 3 | WK 4 | WK 5 | WK 6 | WK 7 | WK 8 | WK 9 | WK 10 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Depo-Provera | √ | |||||||||||
| Immunization | ||||||||||||
| Nasal | √ | √ | √ | |||||||||
| Rectal | √ | √ | √ | |||||||||
| Sublingual | √ | √ | √ |
In each vaccination group, 4 macaques were immunised with a range of antigens and adjuvants by the intranasal, sublingual and intrarectal routes (Table 2). No responses were detected in sera or mucosally after sublingual ovalbumin or intrarectal β-galactosidase delivery (data not depicted). Intranasal immunization with KLH in PBS alone, or co-administered with either Pam3CSK4 or chitosan, failed to induce any detectable local or systemic responses (Fig. 1A). The lack of responsiveness in the presence of Pam3CSK4 or chitosan was in complete contrast to that seen in mice where they promoted a potent immune response. A single animal (IE71) in the TSLP group induced detectable but low systemic IgG and IgA (Fig. 1B) responses with transient IgG responses in the vagina (Fig. 1E) but not nasal mucosa (1 out of 4). Because they failed to induce a systemic IgG response further analysis was not performed on antigen alone, Pam3CSK4 or chitosan adjuvanted samples.
Table 2.
Antigen, adjuvant and route
| Animal ID | IE57, IE58, IE59, IE60 | IE61, IE62, IE63, IE64 | IE65, IE66, IE67, IE68 | IE69, IE70, IE71, IE72 | IE73, IE74, GK02, IF53 | |
|---|---|---|---|---|---|---|
| Antigen | Route | Adjuvant | ||||
| Keyhole Limpet Haemocyanin (KLH) | Intranasal | PBS | Pam3CSK4 | Chitosan | TSLP | R848 |
| Ovalbumin (Ova) | Sublingual | Poly(I:C) | Chitosan | R848 | PBS | Pam3CSK4 |
| Beta-galactosidase (β-Gal) | Intrarectal | R848 | Poly(I:C) | PBS | Pam3CSK4 | MPLA |
Figure 1.
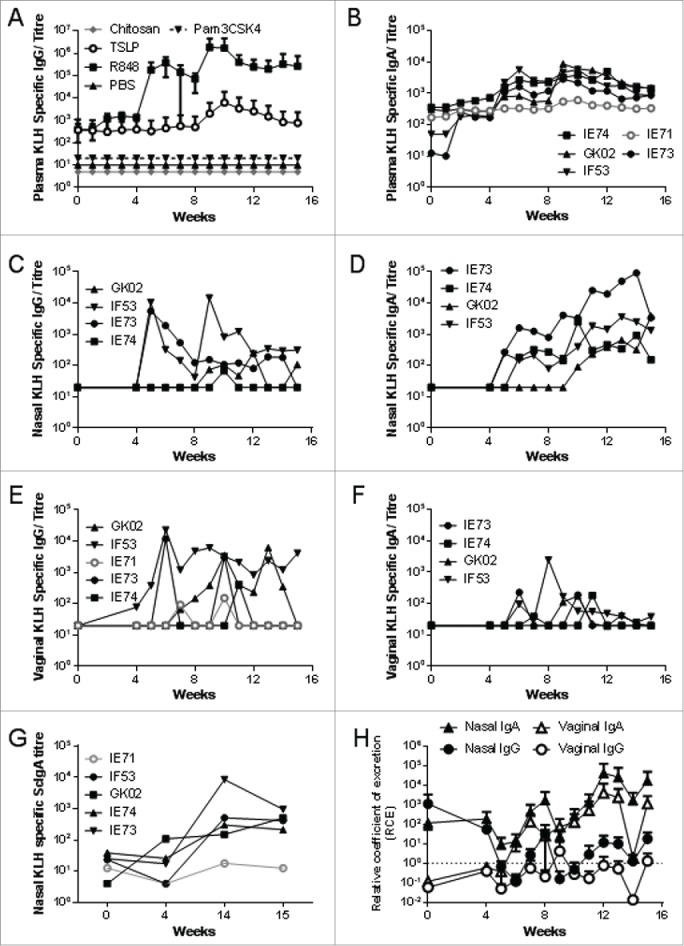
R848 is a potent mucosal adjuvant. Macaques were immunised intranasally with 200 μg KLH in combination with 500 μg of the adjuvants R848, Pam3SCK4 or chitosan, or 50 μg of TSLP or in PBS alone (n = 4 per group). Immunizations were administered at weeks 0, 4 and 8. KLH specific IgG was measured in plasma for all groups (A). Further analysis was performed for animals IE73, IE74, GK02 and IF53 from the R848/ KLH group and IE71 from the TSLP group. KLH specific ELISA were performed for IgG (C, E) and IgA (B, D, F) on plasma (B), nasal wash (C, D) and vaginal Weck-cels (E, F). KLH specific scIgA was measured in nasal samples (G). Relative coefficients of excretion (RCE) compared to albumin in nasal and vaginal samples (H). Data is presented as mean +/− SD of n = 4 animals (A, H) or individual animals (B-G).
Potent immune responses were induced on co-administration of KLH with R848 (Fig. 1). Here systemic specific IgG levels (Fig. 1A) one week after the third immunization had reached mean titer of 1.8×106 (+/− 2.6 × 106). Systemic specific IgA levels (Fig. 1B) were lower with a mean titer of 4.9 × 103 (+/− 1.3 × 103) at the same time point, one week after the final immunization. In contrast, nasal specific IgG (Fig. 1C) and IgA (Fig. 1D) responses were equivalent. Vaginal specific IgG levels (Fig. 1E) were higher than those for IgA (Fig. 1F) reflecting the normal predominance of IgG in the lower genital tract. Sporadic specific IgG responses were observed in rectal secretions of 3/4 animals, rectal responses were low and not positive over concurrent weeks (data not shown). One animal (IF53) had detectable titres of rectal IgG over weeks 11–13 with a peak titer of 632 at week 12. Only one animal had detectable rectal antigen specific IgA (GK02) at a single time-point (Wk11) where it was also positive for IgG. No responses were detected in saliva collected sublingually.
We measured KLH specific secretory IgA (scIgA) levels in nasal samples at weeks 0, 4, 14 and 15 following immunization with KLH. All animals in the R848 group showed induction of high levels of scIgA, in contrast the one animal in the TSLP group (IE71) that had shown a specific immune response to KLH immunization showed lower levels of scIgA (Fig. 1G). Direct comparison in specific scIgA and IgA cannot be made due to the different configuration of the 2 assays. Secretion rates of specific IgG and IgA were evaluated by calculation of their relative coefficients of excretion (RCE) relative to albumin levels. A theoretical value of 1 assumes equal distribution between serum and mucosal compartments. An RCE value significantly higher than 1 indicates that Ig detected in the secretion is locally produced or selectively transported from serum across the mucosal barrier (or both), but does not exclude that a part of the Ig detected is also transudated from serum.11 RCE values for nasal IgA and vaginal IgA levels were greater than 1 indicating that the IgA is either actively transported or locally produced (Fig. 1H), supporting the scIgA data. From this we conclude that R848 was the most effective adjuvant of those tested when delivered intranasally.
Comparison of responses induced by intramuscular and intranasal immunization
As intranasal immunization using R848 as the adjuvant gave good local and systemic IgG and IgA responses, we decided to investigate how the responses would compare to intramuscular immunization. Eight animals from the initial study that had no detectable KLH responses were randomized and reused. KLH and R848 were administered intranasally to 4 animals (IE58, IE60, IE63, IE64) and another 4 animals intramuscularly (IE57, IE59, IE61, IE62) at the same doses as above. Intramuscular immunization with KLH and R848 resulted in consistently high titres for plasma IgG (Fig. 2A) and IgA (Fig. 2B), with titres reaching a peak 2 weeks after the second and third immunization for IgG and 2 weeks after each immunization for IgA. As seen before, intranasal immunization induced both antigen specific IgG (Fig. 2C) and IgA (Fig. 2D). While intramuscular immunization induced higher levels of KLH specific plasma IgG than intranasal (Fig. 2E), similar levels of plasma IgA were observed (Fig. 2F).
Figure 2.
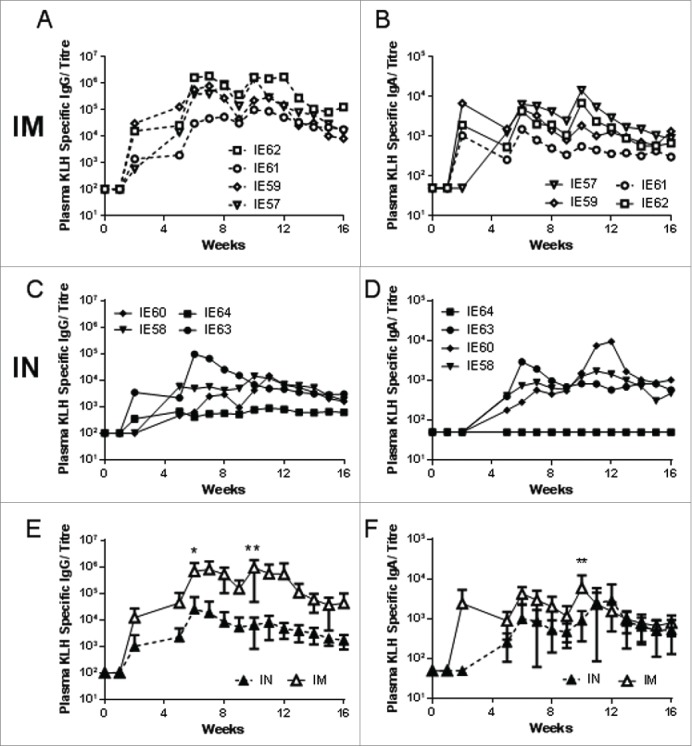
R848 induces antibody responses after mucosal or systemic immunization. Macaques were immunised intramuscularly (IM, open symbols, A, B) or intranasally (IN, closed symbols, C, D) with 200 μg KLH in combination with 500 μg R848 (n = 4 per group). Immunizations were administered at weeks 0, 4 and 8. KLH specific ELISA were performed for IgG (A, C) and IgA (B, D) in plasma, data is presented as individual animals. Pooled data for each route is presented for IgG (E) and IgA (F), where each point represents mean of the n = 4 animals presented in A-D. * p < 0.05, ** p < 0.01 by multiple weighted t-test.
We then examined specific IgG and IgA titres in nasal and vaginal samples from the same animals. Low levels of KLH specific nasal IgG was induced by both the intranasal (Fig. 3A) and intramuscular routes (Fig. 3B), the levels were similar between the 2 routes peaking after the third immunization at week 8 (Fig. 3C). Of note, specific IgA responses were considerably enhanced by IN immunization, peaking after the 3rd immunization, whereas IM immunization failed to induce any IgA responses in nasal fluids (Fig. 3E-G). Low levels of KLH specific vaginal IgG were induced by both the intranasal (Fig. 3I) and intramuscular routes (Fig. 3J), the levels were similar between the 2 routes peaking after the third immunization (Fig. 3K). After IN immunization, but not IM immunization, the RCE for KLH specific nasal IgA (Fig. 3H), but not nasal IgG (Fig. 3D) or vaginal IgG (Fig. 3L) was significantly greater than 1 for the IN immunised animals.
Figure 3.

R848 induces local antibody responses after mucosal or systemic immunization. Macaques were immunised intranasally (A, E, I) or intramuscularly (B, F, J) with 200 μg KLH in combination with 500 μg R848 (n = 4 per group). Immunizations were administered at weeks 0, 4 and 8. KLH specific ELISA were performed for IgG (A, B, C) and IgA (E, F, G) in nasal lavage and IgG in vaginal samples (I, J, K). Relative coefficients of excretion (RCE) compared to albumin in nasal and vaginal samples (D, H, L). Data is presented as individual animals. Pooled data for each route is presented for each sample (C, F, I), where each point represents mean of n = 4 animals.
R848 delivered intranasally induces mild local inflammatory response
To assess safety, macaques were inoculated intranasally with R848 or PBS and nasal swabs collected after inoculation. Nasal swabs were examined for cellular infiltrates by flow cytometry and manual cytospin counts. The introduction of fluid alone (either PBS or R848) intranasally both lead to an acute recruitment of neutrophils (Fig. 4A) and monocytes (Fig. 4B), however the increase in neutrophils was greater with R848, peaking at 6 hours post immunization. R848 also led to a slightly enhanced recruitment of CD3+ cells into the nasal airways over PBS alone, increasing to 96 hours after inoculation (Fig. 4C). Nasal fluids were assessed for cytokine/chemokine responses by cytokine bead array (Fig. 4D-I). Inoculation with fluid alone, and/or the repeat sampling appeared to induce an acute local response, regardless of the agent given. Similar responses were seen in R848 and PBS treated animals for IL-1β (Fig. 4D), CCL2 (Fig. 4E), IL-6 (Fig. 4F) and CXCL8 (Fig. 4G), with an acute peak within 12 hours of exposure and then a return to baseline levels, matching the cell recruitment profile. Other mediators including IL-2, CCL2 and GMCSF were undetectable (data not depicted). Only CCL5 (Fig. 4H) and TNF (Fig. 4I) followed a slightly different kinetic, with sustained detection of both after 24 hours, and a significantly greater level of TNF detectable in the R848 inoculated animals at 96 hours after inoculation. This suggests that R848 induces a mild local inflammation above that induced by exposure to fluid alone in the nasal airway, which may contribute to the adjuvant activity.
Figure 4.

R848 induces mild local inflammation. Macaques were immunised intranasally with R848 or PBS and Nasal swabs collected at time 0, 1, 2, 3, 4, 6, 8, 24, 48, and 96 hrs. Nasal swabs examined for cellular infiltrates, neutrophils by cytospin (A), monocytes (B) and CD3 cells (C) by flow cytometry and fluids for cytokine/chemokine responses by CBA (D–I). Points represent n = 15 animals in the R848 group and 5 animals in the PBS control group +SEM, * p < 0.05 by multiple weighted t test.
Discussion
In this study we assessed a range of adjuvants for mucosal immunization. We compared the adjuvants in combination with model antigens via a range of routes. Of the mucosal routes tested only intranasal immunization induced an antibody response and this only occurred when R848 was used as an adjuvant. It is not clear why the other adjuvants did not boost the response, in a previous study chitosan did have a slight adjuvant effect in cynomologous monkeys,12 though a larger dose of both antigen (ovalbumin) and adjuvant was used, furthermore there was no statistical increase in antibody titer in the antigen plus chitosan groups.12 Chitosan is believed to act as an adjuvant,13 has been delivered in clinical trials intranasally with diphtheria14 and Neisseria meningitidis serogroup C polysaccharide15 boosting the response, but it may be that these antigens are more immunogenic than the ones used in the current study. TSLP has not been tested as an adjuvant in macaques before. One possibility for the absence of a strong adjuvant effect for TSLP in the macaques was that since recombinant human not macaque TSLP was used and there may be species specific differences, however one study has suggested that human TSLP can activate DC from cynomolgus macaques.16
Previous studies have investigated the role of R848 as a mucosal adjuvant in macaques using a prime boost regime,10 though it was not compared to antigen alone in the published study. A recently published study investigating the mechanism of R848 action after systemic delivery demonstrated that R848 led to the release of TNF by macrophages and neutrophils which in turn led to the maturation of Langerhans cells.17 In the current study we observed limited local inflammation after R848 compared to PBS alone. However, there was a trend toward increased neutrophil recruitment after intranasal R848 delivery and increased TNF at 96 hours after immunization, which suggests a similar mechanism works at mucosal surfaces. Apart from the delayed TNF signature, R848 administration gave a very similar cytokine profile to PBS alone, administration of fluid intranasally followed by repeat sampling led to an acute peak of pro-inflammatory cytokines and cells. In the current study, the R848 was given without antigen, a different profile may have been observed in the presence of antigen. These studies suggest that R848 is safe and it is currently in use in a number of clinical trials as a vaccine adjuvant and has been shown to have some effect in combination with GMCSF and Poly(I:C) when administered with a tumor antigen intradermally.18 R848 is also a licensed product for use as a topical cream (Resiquimod), which we have also previously shown induces increased TNF responses when administered intranasally to macaques.19 A study looking at oral administration of R848 to control Hepatitis C virus found adverse reactions in patients given 0.02mg/kg twice weekly for 4 weeks,20 in the current study we use a final dose equivalent to half the safe dose (0.5 mg total in the current study, equivalent to 0.005 mg/kg in the clinical study), this dose in mg/ kg of the macaque was considerably more than used in the previous human study, but for a single administration. Adverse effects from R848 (and other adjuvants) are most likely associated with inflammation and there is a critical balance (“the goldilocks effect”) required in adjuvants between unreactive and hyper inflammatory response. Thus, although R848 did not appear to be toxic in this study we cannot exclude hyper inflammatory responses and associated adverse effects when used at high doses in humans.
We only saw responses following nasal immunization, but not sublingual or intrarectal. To our knowledge this is the first study that has explored the use of sublingual vaccination in macaques using protein alone, though it has been used for DNA vaccines21 and an Adenovirus-protein boost regime.22 While sublingual delivery has been shown to be highly effective in small animal models,23 in our study, we saw no response following sublingual delivery of ovalbumin. It is possible that the success of sublingual delivery in the mouse system in some way reflective of the much smaller size of the murine mouth, and that the delivered antigen either coats a greater surface area of responsive cells, or is swallowed or recirculated to the adenoids or tonsillar lymphoid tissues at the back of the mouth. It is of note that most trials of sublingually delivered antigen are used in the context of immunotherapy with a view to induce tolerance.24 In a clinical trial sublingually delivered human papilloma virus vaccine only induced an immune response in 3 out of 12 volunteers.25 Intrarectal immunization of macaques with peptides adjuvanted with heat labile entertoxin has been demonstrated to induce a cellular response,26,27 but no responses were seen following intrarectal DNA immunization,28 none of these studies assessed the antibody response. Intrarectal delivery of canarypox virus vaccines has been tested in a clinical trial, but failed to induce a response.29 While intrarectal immunization is a conceptually attractive route for the induction of local immune response for sexually transmitted and gastrointestinal infections, the lack of efficacy and possible issues around cultural acceptability could potentially rule this route out. Of the routes tested only intranasal delivered antigen induced an immune response, and the use of this route has been well explored in a range of pre-clinical and clinical studies and it is currently used for the delivery of live attenuated influenza vaccine. There are a number of possible reasons why intranasal is better than other mucosal routes for the induction of both local and systemic immune responses, including longer retention times than the rectal route (particularly in animal models), a kinder environment for protein antigens in terms of pH and digestive enzymes and a higher level of antigen presenting cells.6 The frequency and level of microbacterial colonisation in the different mucosal compartments may also contribute to the levels of immunosuppression at different mucosal surfaces.30
There is an ongoing drive to replace, reduce and refine the use of animals, particularly higher species. Using a novel matrix design we were able to screen a larger number of routes and adjuvants in a species that is more predictive of the responses in the clinic, thereby reducing animal usage and refining the quality of data produced. The importance of using an animal species closer to man is underlined by the difference between the immune response to mucosal vaccination and adjuvants observed in macaques and previously published data from mouse models. In our previous studies we have observed that R848 was a less potent adjuvant than the synthetic TLR4 ligand GLA31 and range of other TLR based adjuvants8 when delivered intranasally in mice, though other groups have observed a boost to antibody responses when R848 was delivered intranasally.32 Likewise as described above, sublingual immunization is extremely effective in mice, but appears to have limited immunogenicity in the macaque study we performed here. The difference between species will be driven by a range of factors including anatomy, pattern recognition receptor expression patterns and response, for example mice do not have a functional TLR8 molecule.33 We believe there is still a role for mouse models in the initial screen for vaccines, particularly for the dissection of immune response and challenge models, but they are poorly predictive of adjuvant strategies effective for mucosal immunization in non-human primates and most likely also in humans.
This study was designed to maximise the number of adjuvants screened and the routes tested, while minimising the number of animals used in order to select products for clinical trials. As such there some limitations to the interpretation of the results, which would need to be addressed in order to fully define mechanisms of action. The adjuvants were selected based on our previous small animal studies, the availability of GMP products (to accelerate clinical trials) and literature review5 but ideally, more adjuvants would have been tested, for example comparing intranasal Poly(I:C) and R848 as both work via endocytic TLR. Due to repeat usage in the same animals, different antigens had to be used for the different routes. The antigens were chosen as they are common experimental antigens, widely used in other studies, but they may have different immunogenicity in macaques. Our previous experience with intrarectal immunization in mice suggests this route is poorly immunogenic.34 The results from this study suggest sublingual delivery of antigen is poorly immunogenic, but further studies with different antigens delivered sublingually are required to confirm our findings. Another limitation to the design of the study is that the close proximity of the immunizations may have altered the outcomes of subsequent immunizations, we don't anticipate original antigenic sin as the antigens are structurally diverse, but there may have been some hangover effect of the inflammation from the adjuvants, which we have seen in previous studies.35 However, given the lack of response to sublingual or intrarectal delivered antigen, this seems to have been minimal. Finally in the follow up studies comparing intramuscular with intranasal R848, the animals used had been previously exposed to KLH intranasally with the adjuvants that had not induced a response (4 with PBS, 4 with Pam3CSK4). The animals were randomized into the follow up study, but there may have been some priming. In conclusion, in the current study, we successfully screened a number of mucosal adjuvants, demonstrating that R848 was safe and effective, suggesting it could be taken forward into phase I clinical trials for intranasally delivered antigen after appropriate toxicity studies.
Methods and Materials
Reagents
Adjuvants: TLR ligands Pam3CSK4 (TLR1/2), Poly(I:C) (TLR3), R848 (TLR7/8) (InvivoGen), Monophosphoryl Lipid A (MPLA, TLR4) (Sigma-Aldrich), human Thymic Stromal Lymphopoietin (ProSpec) and chitosan (NovaMatrix).
Antigens: EndoGrade Keyhole Limpet Haemocyanin (Calbiochem), Ovalbumin (Hyglos GmbH) and β-galactosidase (ProZyme Inc.).
Animals and ethics statement
Rhesus macaques (Macaca mulatta) were obtained from and housed at the Tulane National Primate Research Center. The Institutional Animal Care and Use Committee (IACUC) of Tulane University approved all macaque procedures described (protocol permit number P0031). In this study all procedures were carried out in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health (NIH) and with the recommendations of the Weatherall report; “The use of non-human primates in research.” All procedures were performed under anesthesia using ketamine, and all efforts were made to minimise stress, improve housing conditions, and to provide enrichment opportunities (e.g., objects to manipulate in cage, varied food supplements, foraging and task-oriented feeding methods, interaction with caregivers and research staff).
Macaque plasma samples were shipped to the UK, under strict accordance of The Convention on International Trade in Endangered Species (CITES). Permits were obtained from the US Fish and Wildlife Service and the Department of Environment, Food, and Rural Affairs (DEFRA), UK. The samples were stored at −80°C until further use.
Antigen and route of immunization
Female Rhesus macaques (n = 4 per group) were treated with an intramuscular injection of depomedroxyprogesterone acetate (Depo-Provera) (30 mg) 4 weeks prior to first administration. For each route, immunizations were performed in a staggered regime every 4 weeks according to the schedule in Table 1. The same animals were used for all 3 routes tested, with one week gaps between each route of delivery. Animals were immunised intranasally (IN), with KLH at a dose of 200 µg in a volume of 200 μl per nostril (400 μl total), intrarectally (IR) with 200 µg β-Gal in a volume of 4 ml or sublingually (s.l.) with 225 µg OVA in a volume of 200 μl per each side of the tongue (400 μl total) (Table 2).
Six different adjuvants were assessed (Table 2); PBS (control), Pam3CSK4, MPLA, Poly(I:C), chitosan and R848. Pam3CSK4, MPLA, Poly(I:C), TSLP, and R848 (Resiquimod) were used at 500 µg per dose for all routes of administration, chitosan was used at 1%. All immunizations were administered in PBS.
Following the initial study, animals (in 2 groups of 4 animals, n = 4), were immunised either IN or intramuscularly (IM) with KLH+R848 at the same concentrations as in the previous study, at weeks 0, 4 and 8.
Macaque samples
Blood samples were taken once a week from week 0 to week 16 for determination of systemic antibody levels. Ten ml EDTA anti-coagulated blood was collected at each timepoint and separated into plasma. Cervical, nasal, rectal and sublingual/saliva fluid samples were taken to determine mucosal antibody levels at week 0, then once weekly from week 4 to week 16. All fluid samples were frozen at − 80°C and centrifuged before testing, in addition the plasma samples were heat-treated at 56°C for 30 min prior to centrifugation to remove any non-specific complement activation.
For mucosal fluid collections, animals were first sedated using ketamine hydrochloride, and then secretions were sampled from all tissues using pre-weighed, pre-wet Weck-cel surgical spears (Medtronic Ophthalmics) placed in each site for 5 min. For vaginal secretions, 2 pre-wet Weck-cel spears were placed in the vaginal vault; for sublingual saliva samples, a pre-moistened sponge was placed sublingually; for rectal samples 2 sponges were gently inserted into the rectum and nasal samples were collected by inserting one Weck-cel sponge into each nare.
For all samples, Weck-cel sponges were removed after 5 min, reweighed, and secretions were eluted from the sponges by placing each spear into the upper chamber cup of a Spin-X tube (Corning) to which 300 µl of a hypertonic extraction buffer containing sodium azide (preservative) and protease inhibitors (protease inhibitor cocktail set 1, Calbiochem Merck) was added. Samples were incubated for 10 min on ice, then spun for 15 min at 15,000 RPM, after which the filter cup and sponge were discarded, and the fluid in the bottom chamber was frozen and stored at −80°C until analysis.
Detection of specific and total immunoglobulins
Specific immunoglobulin concentrations in plasma and mucosal samples were measured by sandwich ELISA, adapted from a gp-140 specific ELISA developed by our laboratory.36 96-well plates (medium binding, Greiner Bio-One, UK) were coated with specific antigen (KLH, OVA or β-gal), (5 µg/ml). After washing with 0.05% PBS-Tween 20 (PBST), (Tween-20, Fisher Scientific) and blocking with assay buffer (10% FBS-PBST) (FBS, Gibco®-Life Technologies), plasma samples were added at 1/100 and mucosal at 1/20 dilutions, in triplicate. Bound immunoglobulin was detected by addition of goat anti- monkey IgG (Fc-specific) HRP conjugate (AbD Serotec) or goat anti-monkey (α-specific) – biotin conjugate (ACRIS) followed by avidin – peroxidise detection antibody. Plates were read at 450 nm on a VersaMax™ microplate reader, after addition of SureBlue TMB substrate (KPL) followed by 1N H2SO4 to stop the colorimetric reaction. Endpoint titres were calculated from raw data using SoftMax Pro® software (Molecular Devices) and GraphPad Prism as the reciprocal of the highest dilution giving an absorbance value equal or higher to the background (normal rhesus macaque plasma) plus 2 standard deviations. Cut-off value was set at 0.2.
Relative coefficients of excretion
Albumin levels were detected in plasma, nasal and vaginal samples using a Human Albumin Elisa kit as per manufacturer's instructions (Bethyl Labs Inc.). The relative coefficients of excretion (RCE) was calculated according to the formula: [(Immunoglobulin (Ig) in fluid)/ (Human serum albumin (HAS) in fluid)]/ [(Ig in serum)/(HSA in serum)].11 Calculations were performed based on titres of Ig, not absolute concentration values.
Secretory IgA levels
Secretory IgA (scIgA) levels were determined using a sandwich ELISA, as detailed above for the specific antigen ELISA. Plates were coated with specific antigen, prior to the addition of samples. Detection of scIgA was achieved using a biotin-conjugated IgG antibody specific to monkey secretory component, (Nordic Immunological Laboratories), at 1/20 dilution, followed by detection by HRP-streptavidin.
Inflammatory profile of intranasal R848
Following completion of the above experiments, 20 of the animals selected from the above experiment were briefly re-used in experiments to assess safety of nasal R848 administration. Then, 15 animals received an intranasal inoculation with 500 µg R848 in 400 μl PBS (200 μl per nostril), and 5 controls were treated with PBS alone. One nasal swab was collected from each nare at time 0, 1, 2, 3, 4, 6, 8, 24, 48, and 96 hrs. One swab was immersed and eluted in 1 ml sterile PBS for determining cellular infiltrates by flow cytometry and manual cytospin counts, and the other was processed in the extraction buffer as described above for assessing cytokine/chemokine responses by multiplex bead array.
For cell counts, 200 μl aliquots of cells that were eluted in PBS were cytospun onto glass slides and stained with a Wrights stain for manual cell counts, and counted by a pathologist. The remaining cells were stained with anti-CD3, CD4, CD8, and CCR5 monoclonal antibodies, and analyzed by flow cytometry.
For multiplex cytokine bead arrays (CBA, Becton Dickinson), nasal fluid was incubated with beads pre-conjugated to anti-cytokine/chemokine antibodies against IL-1β, IL-2, IL-6, CXCL8/IL-8, CCL3/MIP-1α, CCL4/MIP-1β, G-CSF, GM-CSF, CCL5/RANTES, CCL2/MCP-1, TNF, and IFN-γ and analyzed using a FACS Array bioanalyzer (Becton Dickinson).
Statistical analysis
Comparisons of 2 groups were performed using Student's t tests. Comparisons of multiple groups were performed using one- or 2-way ANOVA with appropriate post-tests. All statistical tests were performed using GraphPad Prism version 6.01 for Windows (GraphPad Software).
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
We also wish to thank Megan Gardner, Meagan Watkins, and Kelsi and Terri Rasmussen for assistance with the nonhuman primate studies.
Funding
This work was funded by a grant to RJS by the Center for HIV/AIDS Vaccine Immunology (CHAVI) # U19 AI067854–05. We gratefully acknowledge Dormeur Investment Service Ltd for providing funds to purchase equipment used in these studies.
References
- 1.Lycke N. Recent progress in mucosal vaccine development: potential and limitations. Nat Rev Immunol 2012; 12:592-605; PMID:22828912; http://dx.doi.org/ 10.1038/nri3251 [DOI] [PubMed] [Google Scholar]
- 2.Neutra MR, Kozlowski PA. Mucosal vaccines: the promise and the challenge. Nat Rev Immunol 2006; 6:148-58; PMID:16491139; http://dx.doi.org/ 10.1038/nri1777 [DOI] [PubMed] [Google Scholar]
- 3.Mestecky J, McGhee JR, Michalek SM, Arnold RR, Crago SS, Babb JL. Concept of the local and common mucosal immune response. Adv Exp Med Biol 1978; 107:185-92 [DOI] [PubMed] [Google Scholar]
- 4.Holmgren J, Czerkinsky C. Mucosal immunity and vaccines. Nat Med 2005; 11:S45-S53; PMID:15812489; http://dx.doi.org/ 10.1038/nm1213 [DOI] [PubMed] [Google Scholar]
- 5.Newsted D, Fallahi F, Golshani A, Azizi A. Advances and challenges in mucosal adjuvant technology. Vaccine 2015; 33:2399-405; PMID:25865473; http://dx.doi.org/ 10.1016/j.vaccine.2015.03.096 [DOI] [PubMed] [Google Scholar]
- 6.Riese P, Sakthivel P, Trittel S, Guzman CA. Intranasal formulations: promising strategy to deliver vaccines. Expert opin Drug Deliv 2014; 11:1619-34; PMID:24962722; http://dx.doi.org/ 10.1517/17425247.2014.931936 [DOI] [PubMed] [Google Scholar]
- 7.Lewis DJ, Huo Z, Barnett S, Kromann I, Giemza R, Galiza E, Woodrow M, Thierry-Carstensen B, Andersen P, Novicki D, et al.. Transient facial nerve paralysis (Bell's palsy) following intranasal delivery of a genetically detoxified mutant of Escherichia coli heat labile toxin. PloS one 2009; 4:e6999; PMID:19756141; http://dx.doi.org/ 10.1371/journal.pone.0006999 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Buffa V, Klein K, Fischetti L, Shattock RJ. Evaluation of TLR Agonists as Potential Mucosal Adjuvants for HIV gp140 and Tetanus Toxoid in Mice. PloS one 2012; 7:e50529; PMID:23272062; http://dx.doi.org/ 10.1371/journal.pone.0050529 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Van Roey GA, Arias MA, Tregoning JS, Rowe G, Shattock RJ. Thymic stromal lymphopoietin (TSLP) acts as a potent mucosal adjuvant for HIV-1 gp140 vaccination in mice. Eur J Immunol 2012; 42:353-63; PMID:22057556; http://dx.doi.org/ 10.1002/eji.201141787 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Fouda GG, Amos JD, Wilks AB, Pollara J, Ray CA, Chand A, Kunz EL, Liebl BE, Whitaker K, Carville A, et al.. Mucosal immunization of lactating female rhesus monkeys with a transmitted/founder HIV-1 envelope induces strong Env-specific IgA antibody responses in breast milk. J Virol 2013; 87:6986-99; PMID:23596289; http://dx.doi.org/ 10.1128/JVI.00528-13 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Li Y, To J, Verdia-Baguena C, Dossena S, Surya W, Huang M, Paulmichl M, Liu DX, Aguilella VM, Torres J. Inhibition of the human respiratory syncytial virus small hydrophobic protein and structural variations in a bicelle environment. J Virol 2014; 88:11899-914; PMID:25100835; http://dx.doi.org/ 10.1128/JVI.00839-14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Kobayashi T, Fukushima K, Sannan T, Saito N, Takiguchi Y, Sato Y, Hasegawa H, Ishikawa K. Evaluation of the effectiveness and safety of chitosan derivatives as adjuvants for intranasal vaccines. Viral Immunol 2013; 26:133-42; PMID:23509985; http://dx.doi.org/ 10.1089/vim.2012.0057 [DOI] [PubMed] [Google Scholar]
- 13.Vasiliev YM. Chitosan-based vaccine adjuvants: incomplete characterization complicates preclinical and clinical evaluation. Expert Rev Vaccines 2015; 14:37-53; PMID:25262982; http://dx.doi.org/ 10.1586/14760584.2015.956729 [DOI] [PubMed] [Google Scholar]
- 14.McNeela EA, Jabbal-Gill I, Illum L, Pizza M, Rappuoli R, Podda A, Lewis DJ, Mills KH. Intranasal immunization with genetically detoxified diphtheria toxin induces T cell responses in humans: enhancement of Th2 responses and toxin-neutralizing antibodies by formulation with chitosan. Vaccine 2004; 22:909-14; PMID:15161067; http://dx.doi.org/ 10.1016/j.vaccine.2003.09.012 [DOI] [PubMed] [Google Scholar]
- 15.Huo Z, Sinha R, McNeela EA, Borrow R, Giemza R, Cosgrove C, Heath PT, Mills KH, Rappuoli R, Griffin GE, et al.. Induction of protective serum meningococcal bactericidal and diphtheria-neutralizing antibodies and mucosal immunoglobulin A in volunteers by nasal insufflations of the Neisseria meningitidis serogroup C polysaccharide-CRM197 conjugate vaccine mixed with chitosan. Infect Immun 2005; 73:8256-65; PMID:16299322; http://dx.doi.org/ 10.1128/IAI.73.12.8256-8265.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Robinson M, Strickland I, Coyle A, Sleeman M, Anderson I, May RM. Thymic Stromal Lymphopoeitin (TSLP) Activates Human And Cynomolgus Macaque Dendritic Cells And Induces Th1 As Well As Th2 Responses In Human Total T Helper Cells. ATS: American Thoracic Society, 2010:A3794-A. [Google Scholar]
- 17.Epaulard O, Adam L, Poux C, Zurawski G, Salabert N, Rosenbaum P, Dereuddre-Bosquet N, Zurawski S, Flamar AL, Oh S, et al.. Macrophage- and neutrophil-derived TNF-α instructs skin langerhans cells to prime antiviral immune responses. J Immunol 2014; 193:2416-26; http://dx.doi.org/ 10.4049/jimmunol.1303339 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Morse MA, Chapman R, Powderly J, Blackwell K, Keler T, Green J, Riggs R, He LZ, Ramakrishna V, Vitale L, et al.. Phase I study utilizing a novel antigen-presenting cell-targeted vaccine with Toll-like receptor stimulation to induce immunity to self-antigens in cancer patients. Clin Cancer Res 2011; 17:4844-53; PMID:21632857; http://dx.doi.org/ 10.1158/1078-0432.CCR-11-0891 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Clejan S, Mandrea E, Pandrea IV, Dufour J, Japa S, Veazey RS. Immune responses induced by intranasal imiquimod and implications for therapeutics in rhinovirus infections. J Cell Mol Med 2005; 9:457-61; PMID:15963264; http://dx.doi.org/ 10.1111/j.1582-4934.2005.tb00370.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Pockros PJ, Guyader D, Patton H, Tong MJ, Wright T, McHutchison JG, Meng TC. Oral resiquimod in chronic HCV infection: Safety and efficacy in 2 placebo-controlled, double-blind phase IIa studies. J Hepatol 2007; 47:174-82; PMID:17532523; http://dx.doi.org/ 10.1016/j.jhep.2007.02.025 [DOI] [PubMed] [Google Scholar]
- 21.McCluskie MJ, Brazolot Millan CL, Gramzinski RA, Robinson HL, Santoro JC, Fuller JT, Widera G, Haynes JR, Purcell RH, Davis HL, et al.. Route and method of delivery of DNA vaccine influence immune responses in mice and non-human primates. Mol Med 1999; 5:287-300; PMID:10390545; http://dx.doi.org/ 10.1007/s0089490050287 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Xiao P, Patterson LJ, Kuate S, Brocca-Cofano E, Thomas MA, Venzon D, Zhao J, DiPasquale J, Fenizia C, Lee EM, et al.. Replicating adenovirus-simian immunodeficiency virus (SIV) recombinant priming and envelope protein boosting elicits localized, mucosal IgA immunity in rhesus macaques correlated with delayed acquisition following a repeated low-dose rectal SIV(mac251) challenge. J Virol 2012; 86:4644-57; PMID:22345466; http://dx.doi.org/ 10.1128/JVI.06812-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Kraan H, Vrieling H, Czerkinsky C, Jiskoot W, Kersten G, Amorij JP. Buccal and sublingual vaccine delivery. J Control Release 2014; 190:580-92; PMID:24911355; http://dx.doi.org/ 10.1016/j.jconrel.2014.05.060 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Larche M. Immune mechanisms of sublingual immunotherapy: are oral Langerhans cells the masters of tolerance? J Allergy Clin Immunol 2010; 126:646-7; PMID:20816196; http://dx.doi.org/ 10.1016/j.jaci.2010.07.027 [DOI] [PubMed] [Google Scholar]
- 25.Huo Z, Bissett SL, Giemza R, Beddows S, Oeser C, Lewis DJ. Systemic and mucosal immune responses to sublingual or intramuscular human papilloma virus antigens in healthy female volunteers. PloS One 2012; 7:e33736; PMID:22438987; http://dx.doi.org/ 10.1371/journal.pone.0033736 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Belyakov IM, Isakov D, Zhu Q, Dzutsev A, Berzofsky JA. A novel functional CTL avidity/activity compartmentalization to the site of mucosal immunization contributes to protection of macaques against simian/human immunodeficiency viral depletion of mucosal CD4+ T cells. J Immunol 2007; 178:7211-21; http://dx.doi.org/ 10.4049/jimmunol.178.11.7211 [DOI] [PubMed] [Google Scholar]
- 27.Belyakov IM, Hel Z, Kelsall B, Kuznetsov VA, Ahlers JD, Nacsa J, Watkins DI, Allen TM, Sette A, Altman J, et al.. Mucosal AIDS vaccine reduces disease and viral load in gut reservoir and blood after mucosal infection of macaques. Nat Med 2001; 7:1320-6; PMID:11726972; http://dx.doi.org/ 10.1038/nm1201-1320 [DOI] [PubMed] [Google Scholar]
- 28.Sharpe S, Hanke T, Tinsley-Bown A, Dennis M, Dowall S, McMichael A, Cranage M. Mucosal immunization with PLGA-microencapsulated DNA primes a SIV-specific CTL response revealed by boosting with cognate recombinant modified vaccinia virus Ankara. Virology 2003; 313:13-21; PMID:12951017; http://dx.doi.org/ 10.1016/S0042-6822(03)00282-4 [DOI] [PubMed] [Google Scholar]
- 29.Wright PF, Mestecky J, McElrath MJ, Keefer MC, Gorse GJ, Goepfert PA, Moldoveanu Z, Schwartz D, Spearman PW, El Habib R, et al.. Comparison of systemic and mucosal delivery of 2 canarypox virus vaccines expressing either HIV-1 genes or the gene for rabies virus G protein. J Infect Dis 2004; 189:1221-31; PMID:15031791; http://dx.doi.org/ 10.1086/382088 [DOI] [PubMed] [Google Scholar]
- 30.Spasova DS, Surh CD. Blowing on embers: commensal microbiota and our immune system. Front Immunol 2014; 5:318; PMID:25120539; http://dx.doi.org/ 10.3389/fimmu.2014.00318 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Arias MA, Van Roey GA, Tregoning JS, Moutaftsi M, Coler RN, Windish HP, Reed SG, Carter D, Shattock RJ. Glucopyranosyl Lipid Adjuvant (GLA), a Synthetic TLR4 Agonist, Promotes Potent Systemic and Mucosal Responses to Intranasal Immunization with HIVgp140. PloS one 2012; 7:e41144; PMID:22829921; http://dx.doi.org/ 10.1371/journal.pone.0041144 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Gwinn WM, Johnson BT, Kirwan SM, Sobel AE, Abraham SN, Gunn MD, Staats HF. A comparison of non-toxin vaccine adjuvants for their ability to enhance the immunogenicity of nasally-administered anthrax recombinant protective antigen. Vaccine 2013; 31:1480-9; PMID:23352329; http://dx.doi.org/ 10.1016/j.vaccine.2013.01.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Ketloy C, Engering A, Srichairatanakul U, Limsalakpetch A, Yongvanitchit K, Pichyangkul S, Ruxrungtham K. Expression and function of Toll-like receptors on dendritic cells and other antigen presenting cells from non-human primates. Vet Immunol Immunopathol 2008; 125:18-30; PMID:18571243; http://dx.doi.org/ 10.1016/j.vetimm.2008.05.001 [DOI] [PubMed] [Google Scholar]
- 34.Klein K, Mann JF, Rogers P, Shattock RJ. Polymeric penetration enhancers promote humoral immune responses to mucosal vaccines. J Control Release 2014; 183:43-50; PMID:24657807; http://dx.doi.org/ 10.1016/j.jconrel.2014.03.018 [DOI] [PubMed] [Google Scholar]
- 35.Tregoning JS, Buffa V, Oszmiana A, Klein K, Walters AA, Shattock RJ. A “prime-pull” vaccine strategy has a modest effect on local and systemic antibody responses to HIV gp140 in mice. PloS one 2013; 8:e80559; PMID:24260419; http://dx.doi.org/ 10.1371/journal.pone.0080559 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Lewis DJ, Fraser CA, Mahmoud AN, Wiggins RC, Woodrow M, Cope A, Cai C, Giemza R, Jeffs SA, Manoussaka M, et al.. Phase I Randomised Clinical Trial of an HIV-1 CN54, Clade C, Trimeric Envelope Vaccine Candidate Delivered Vaginally. PloS one 2011; 6:e25165; PMID:21984924; http://dx.doi.org/ 10.1371/journal.pone.0025165 [DOI] [PMC free article] [PubMed] [Google Scholar]