

Abstract
We present an angiofibroma of soft tissue with the karyotype 46,XY,t(4;5)(q24;q31),t(5;8;17)(p15;q13;q21) [8]/46,XY,t(1;14)(p31;q32)[2]/46,XY[3]. RNA-sequencing showed that the t(4;5)(q24;q31) resulted in recombination of the genes TBCK on 4q24 and P4HA2 on 5q31.1 with generation of an in-frame TBCK-P4HA2 and the reciprocal but out-of-frame P4HA2-TBCK fusion transcripts. The putative TBCK-P4HA2 protein would contain the kinase, the rhodanese-like domain, and the Tre-2/Bub2/Cdc16 (TBC) domains of TBCK together with the P4HA2 protein which is a component of the prolyl 4-hydroxylase. The t(5;8;17)(p15;q13;q21) three-way chromosomal translocation targeted AHRR (on 5p15), NCOA2 (on 8q13), and ETV4 (on 17q21) generating the in-frame fusions AHRR-NCOA2 and NCOA2-ETV4 as well as an out-of-frame ETV4-AHRR transcript. In the AHRR-NCOA2 protein, the C-terminal part of AHRR is replaced by the C-terminal part of NCOA2 which contains two activation domains. The NCOA2-ETV4 protein would contain the helix-loop-helix, PAS_9 and PAS_11, CITED domains, the SRC-1 domain of NCOA2 and the ETS DNA-binding domain of ETV4. No fusion gene corresponding to t(1;14)(p31;q32) was found. Our findings indicate that, in spite of the recurrence of AHRR-NCOA2 in angiofibroma of soft tissue, additional genetic events (or fusion genes) might be required for the development of this tumor.
Keywords: angiofibroma of soft tissue, cytogenetics, fusion genes, AHRR-NCOA2, NCOA2-ETV4, TBCK-P4HA2, RNA-sequencing
Introduction
Angiofibroma of soft tissue is a recently described benign fibrovascular tumor of unknown cellular origin (1). It arises most commonly in the extremities of middle-aged adults but displays a broad anatomic and age distribution. Microscopically, it is characterized by bland, uniform, probably fibroblastic spindle cell set in an abundant fibromyxoid stroma, with a prominent and highly characteristic vascular pattern composed of innumerable branching, thin-walled blood vessels (1). Cytogenetic knowledge about angiofibroma of soft tissue is based on the analysis of six such tumors of which four showed a balanced t(5;8)(p15;q12) translocation and a fifth tumor showed a three-way t(5;8;8)(p15;q13;p11) (1). Molecular analysis of four tumors carrying the t(5;8)(p15;q12) showed in-frame AHRR-NCOA2 and NCOA2-AHHR fusion transcripts in all of them (2). A GTF2I-NCOA2 fusion gene was detected in a fifth tumor carrying a t(7;8;14)(q11;q13;q31) as the sole chromosome change (3). To the best of our knowledge, the above-mentioned tumors are the only angiofibromas of soft tissue which have been investigated both cytogenetically and molecularly for fusion genes. An additional angiofibroma of soft tissue with t(5;8)(p15;q12) was also reported but without molecular analysis (4). In three other studies, fluorescence in situ hybridization (FISH) was performed with probes for NCOA2 showing rearrangements of the NCOA2; however, no further investigation of fusion genes was performed (5–7).
We report here an angiofibroma of soft tissue which had the chromosome translocations t(4;5)(q24;q31) and t(5;8;17)(p15;q13;q21) and identified the fusion genes generated by the two translocations. Our data show that, in addition to the reported AHRR-NCOA2, the tumor carried also other fusion genes resulting from the chromosomal aberrations that might have contributed to tumorigenesis as well.
Materials and methods
Ethics statement
The study was approved by the regional Ethics Committee (Regional komité for medisinsk forskningsetikk Sør-Øst, Norge; http://helseforskning.etikkom.no), and written informed consent was obtained from the patient to publication of the case details. The Ethics Committee's approval included a review of the consent procedure. All patient information has been de-identified.
Case history
The patient was a 45-year-old male in whom MRI of the abdomen and pelvis showed a 53-mm tumor in the right inguinal region partially surrounding large vessels. The patient had been aware of the lesion for several years. Surgery was performed with removal of the entire tumor including part of the right deep femoral artery with immediate reconstruction of the vessel. The postoperative period was eventless and to date there is no sign of tumor relapse.
The specimen (58×45×45 mm) showed an encapsuled, well-circumscribed tumor with a homogenous gray/white cut surface. There were no signs of necrosis or bleeding. Routine microscopy showed a tumorous proliferation of small, spindled cells without atypia or mitotic activity (Fig. 1A–C). There were a lot of small, thin-walled blood vessels in the background (Fig. 1A–C). Immunohistochemical examination showed low proliferative activity (MIB1/Ki67 <5%) (Fig. 1D) and the vessels highlighted by the endothelial marker CD34 (Fig. 1E). The clinical setting as well as histopathological features fit well with a diagnosis of angiofibroma of soft tissue (1).
Figure 1.

Microscopic examination of the angiofibroma of soft tissue. (A) H&E-100×. (B) H&E-200×. (C) H&E-400×. (D) Immunoexpression of MIB1-200×. (E) Immunoexpression of CD34-200×.
G-banding and karyotyping
Fresh tissue from the tumor was processed for cytogenetic analysis as part of our diagnostic routine. The sample was disaggregated mechanically and enzymatically with collagenase II (Worthington Biochemical Corp., Freehold, NJ, USA). The resulting cells were cultured and harvested using standard techniques. Chromosome preparations were G-banded with Wright stain and examined. The karyotype was written according to the International System for Human Cytogenetic Nomenclature (ISCN) 2013 guidelines (8).
High-throughput paired-end RNA-sequencing
Total RNA was extracted using miRNeasy Mini Kit according to the manufacturer's instructions (Qiagen, Hilden, Germany). Tumor tissue was disrupted and homogenized in QIAzol Lysis Reagent (Qiagen) using a 5-mm stainless steel bead and TissueLyser II (Qiagen). Subsequently, total RNA was purified using QIAcube (Qiagen). The RNA quality was evaluated using the Experion Automated Electrophoresis System (Bio-Rad Laboratories, Oslo, Norway). The RNA quality indicator (RQI) was 8.5. Total RNA (3 µg) was sent for high-throughput paired-end RNA-sequencing at the Norwegian Sequencing Centre, Ullevål Hospital (http://www.sequencing.uio.no/). Detailed information about the high-throughput paired-end RNA-sequencing was given elsewhere (9). The software FusionCatcher (10) (https://github.com/ndaniel/fusioncatcher) was used for the discovery of fusion transcripts.
Molecular genetic analyses
The primers used for PCR amplification and sequencing are listed in Table I. The primer combinations, target fusion transcripts, and results of PCR amplifications are shown in Table II. cDNA was synthesized from 2 µg of total RNA in a 20-µl reaction volume using iScript Advanced cDNA Synthesis Kit for RT-qPCR according to the manufacturer's instructions (Bio-Rad Laboratories). cDNA was diluted to 100 µl and 2 µl were used as template in subsequent PCR assays. The 25-µl PCR volumes contained 12.5 µl of Premix Taq (Takara Bio Europe SAS, Saint-Germain-en-laye, France), 1 µl of diluted cDNA, and 0.4 µM of each of the forward and reverse primers (Table II). The quality of the cDNA synthesis was examined by amplification of a cDNA fragment of the TBCK gene using the primers TBCK-2558F1 and TBCK-2908R1. The PCRs were run on a C1000 Thermal cycler (Bio-Rad Laboratories) with the following cycling for the amplifications: an initial denaturation at 94°C for 30 sec, 35 cycles of 7 sec at 98°C, 7 sec at 60°C, 1 min at 72°C, and a final extension for 5 min at 72°C.
Table I.
Primers used for PCR amplification and Sanger sequencing analyses.
| Name | Sequence (5′→3′) | Position | Reference sequence | Gene |
|---|---|---|---|---|
| TBCK-2908R1 | TGGCGTGGATATGAAGAACTGTGC | 2931–2908 | NM_033115.4 | TBCK |
| TBCK-2558F1 | CCTGGTGGTTGACATCCGGAATAG | 2558–2581 | NM_033115.4 | TBCK |
| P4HA2-785R1 | AGCCAGGTAGCCCTCAGCATCAG | 807–785 | NM_004199.2 | P4HA2 |
| P4HA2-33F1 | CCGCGGGAGGTTCTGGAAAC | 33–52 | NM_001142598.1 | P4HA2 |
| NCOA2-intr14-R1 | CACCATGTCGAGACTGCTGGCTC | 71106777–71106799 | NC_018919.2 | NCOA2 |
| NCOA2-3364R1 | TCACTCGGAGACTCAGCTGCAGG | 3386–3364 | NM_006540.2 | NCOA2 |
| NCOA2-2858F1 | CTGGACCTTTCCCACCAATCAGAA | 2858–2881 | NM_006540.2 | NCOA2 |
| ETV4-1496R1 | GGGGCTCTCATCCAAGTGGGAC | 1517–1496 | NM_001986.2 | ETV4 |
| ETV4-863F1 | TGGGGTCAATGGGCACAGGTAC | 863–884 | NM_001986.2 | ETV4 |
| AHRR-1932R1 | TGCAGGGTGGAAAGGGGTCAG | 1952–1932 | NM_020731.4 | AHRR |
| AHRR-1503F1 | AGCAGACCCATGCGGGATGTC | 1503–1523 | NM_020731.4 | AHRR |
| AHRR-1425F1 | TGTGTCCAGGGCACTTTCAGGAA | 1425–1447 | NM_020731.4 | AHRR |
| EGFL7-353F1 | ACCCCAAAGCCACATCTGTAGCC | 353–375 | NM_016215.4 | EGFL7 |
| MCF2l-3271R1 | CGCCACGACCGTGTATTTACCTG | 3293–3271 | NM_024979.4 | MCF2L |
| CYP1B1-132F1 | TCAACGCTGTGAGGAAACCTCGA | 132–154 | NM_000104.3 | CYP1B1 |
| CLU-1164R1 | GACCTGGAGGGATTCGTCGAGC | 1185–1164 | NM_001831.3 | CLU |
Table II.
Primer combinations, target fusion transcripts and results of PCR amplification.
| Primer combination | Target fusion transcripts | Results |
|---|---|---|
| P4HA2-33F1/TBCK-2908R1 | P4HA2-TBCK | Positive |
| TBCK-2558F1/P4HA2-785R1 | TBCK-P4HA2 | Positive |
| AHRR-1503F1/NCOA2-intr14-R1 | AHRR-NCOA2 | Positive |
| AHRR-1425F1/NCOA2-3364R1 | AHRR-NCOA2 | Positive |
| ETV4-863F1/AHRR-1932R1 | ETV4-AHRR | Positive |
| NCOA2-2858F1/ETV4-1496R1 | NCOA2-ETV4 | Positive |
| EGFL7-353F1/MCF2L-3271R1 | EGFL7-MCF2L | Negative |
| CYP1B1-132F1/CLU-1164R1 | CYP1B1-CLU | Negative |
The PCR products were analyzed on a QIAxcel Advanced System according to the manufacturer's instructions (Qiagen). The remaining PCR products were purified using the QIAquick PCR Purification Kit or the QIAquick Gel Extraction Kit (both from Qiagen) and direct sequenced using the dideoxy procedure with the ABI Prism BigDye Terminator v1.1 Cycle Sequencing Kit (Applied Biosystems, Foster City, CA, USA) on the Applied Biosystems 3500 Genetic Analyzer sequencing system. The BLAST software (http://www.ncbi.nlm.nih.gov/BLAST/) was used for computer analysis of the sequence data.
Results
Cytogenetic analysis
The G-banding analysis showed that the tumor had two cytogenetically unrelated clones. The first clone, found in eight metaphases, had the t(4;5)(q24;q31) and t(5;8;17)(p15;q13;q21) chromosome aberrations (Fig. 2A). The second, found in two metaphases, had the t(1;14)(p31;q32) abnormality (Fig. 2B). This yielded the following karyotype: 46,XY,t(4;5)(q24;q31),t(5;8;17) (p15;q13;q21)[8]/46,XY,t(1;14)(p31;q32)[2]/46,XY[3].
Figure 2.
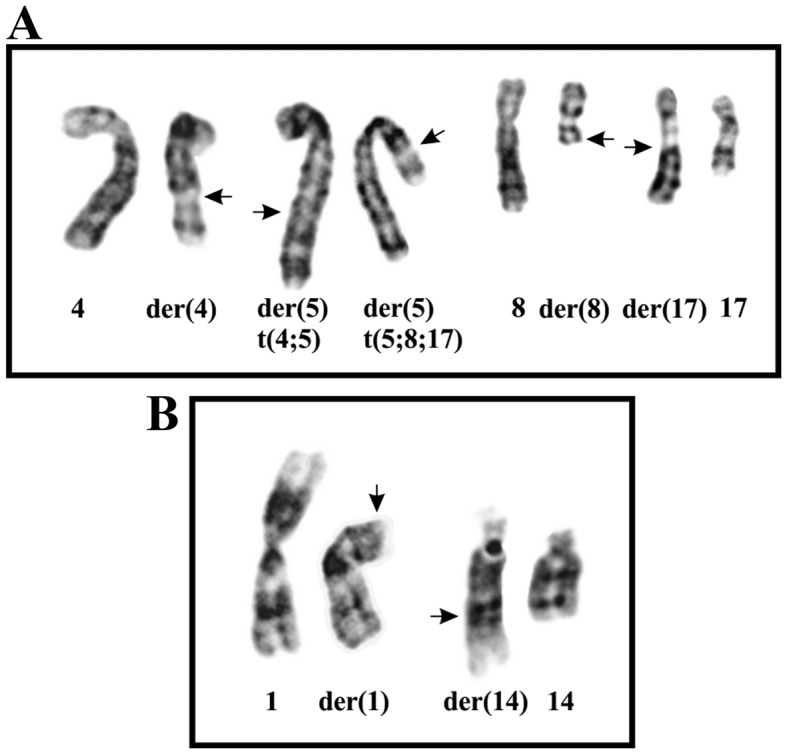
Cytogenetic analysis of the angiofibroma of soft tissue. (A) Partial karyotype showing from left to right the chromosomes 4, der(4)t(4;5) (q24;q31), der(5)t(4;5)(q24;q31), der(5)t(5;8;17)(p15;q13;q21), 8, der(8)t(5;8;17) (p15;q13;q21), der(17)t(5;8;17)(p15;q13;q21), and 17. (B) Partial karyotype showing the der(1)t(1;14)(p31;q32) and der(14)t(1;14)(p31;q32) together with the corresponding normal chromosome homologs. Breakpoint positions are indicated by arrows.
High-throughput paired-end RNA-sequencing analysis
Using the FusionCatcher software with the FASTQ files obtained from the Norwegian Sequencing Centre, Ullevål Hospital (http://www.sequencing.uio.no/), 39 potential fusions were found: 28 fusions were described as readthrough short-distance fusions and 5 as pseudogenes (Table III). Among the other fusions, the program detected the P4HA2-TBCK and the reciprocal TBCK-P4HA2. According to the UCSC genome Browser on Human, Feb. 2009, (GRCh37/hg19) assembly (http://genome-euro.ucsc.edu/cgi-bin/hggateway), P4HA2 maps on chromosome subband 5q31.1 and TBCK on band 4q24. Thus, the two fusions P4HA2-TBCK and the reciprocal TBCK-P4HA2 most probably were the result of the balanced chromosome translocation t(4;5)(q24;q31). FusionCatcher also detected AHRR-NCOA2 and ETV4-AHRR which correspond to the three-way t(5;8;17)(p15;q13;q21) found in the tumor. The three genes AHRR, NCOA2, and ETV4 map to chromosome subbands 5p15.33, 8q13.3, and 17q21.31, respectively (https://genome.ucsc.edu/). In the three-way t(5;8;17), the moving of 5p15 to 8q13 generated the AHRR-NCOA2 fusion whereas the translocation of 17q21 to 5p15 generated the ETV4-AHRR. We assume that the moving of 8q13 to 17q21 would have generated an NCOA2-ETV4 fusion but no such fusion was, for unknown reasons, detected by FusionCatcher. The fusion transcrips EGFL7-MCF2L and a CYP1B1-CLU were also detected by the analysis with FusionCatcher, in all likelihood generated by t(9;13)(q34;q34) and t(2;8)(p22.2;p21.1), respectively. No fusion gene corresponding to the cytogenetically detected t(1;14)(p31;q32) was found.
Table III.
Fusion transcripts detected using FusionCatcher.
| 5′-Partner gene | 3′-Partner gene | Fusion description | Fusion sequence |
|---|---|---|---|
| PCDP1 | TMEM177 | Readthrough | ATTCTAGAATGAAAGTCACCAGTAG*gaaagggaacatcacagaaaggtga |
| MIR155HG | JAM2 | Readthrough | CAAGGAGACGCTCCTGGCACTGCAG*atcataaggcctatgggttttctgc |
| GOLT1A | KISS1 | Readthrough | ATGATCTCCATCACCGAATGGCAGA*cctcaaggcacttctaggacctgcc |
| SHISA9 | U91319.1 | Readthrough | AAGTACGCCTCCTTAAAGGCAGTCG*agctggaacacccttcttctcctgc |
| VPS45 | PLEKHO1 | Readthrough | GCACCACAGTGCACAACACGAAAAG*ggacctcaggatggaaaccagcagc |
| P4HA2 | TBCK | AACGCCGGGAGCTGCGAGTGTCCAG*tttgcagctcaccttgtgaagatga | |
| TBCK | P4HA2 | GCATGTGGCAAAACACACAGCTGAG*acacttccctctgtgaccatgaaac | |
| ADCK4 | NUMBL | Readthrough | TCCAGCCTCTCAGTGTGTTGGAGAG*acggggcgggcaccatgaacaagtt |
| ETV4 | AHRR | AAGGTCAGAGAAGTGACTGTTGATG*ggggacctgtgtggtccgacgctgc | |
| FOSB | PPM1N | Readthrough | TCCACCCACCGCCGCCGCCTCCCAG*aaggggcaggatggggctgggaagt |
| MFSD7 | ATP5I | Readthrough | GGGGAGGATCCACTTGACTGGACAG*attacctaaaacctcgggcagaaga |
| DPY19L2 | DPY19L2P2 | Pseudogene | TTCTTCATCTTTGTTAATGACGTGG*ctaattcaaggtagtgcctggtggt |
| DPY19L2P2 | DPY19L2 | Pseudogene | TTCTTCATCTTTGTTAATGACATGG*ctaattcaaggtagtgcctggtggt |
| MATR3 | PAIP2 | Readthrough | CCGCGTCCCGCTCGCTGGGAGAGAG*gttaaaaacgacaaccaacatcagc |
| LINC00893 | LINC00894 | Antisense | AGGAAGCAGGAATGCTGGAGATGAG*acggagttttgctcttgttgcccag |
| PTPRG | C3ORF14 | Readthrough | GAGGCCTGGAGTATTCACAGACATT*ggcaagcactttaaccttttaagcc |
| SIX3 | AC012354.6 | Readthrough | AGACACCGGCACCTCCATCCTCTCG*acaaggccacctacatcccaagcca |
| CTBS | GNG5 | Readthrough | GCGGGCTCCTTATTATAACTATAAA*gtttcccaggcagctgcagacttga |
| CYP1B1 | CLU | Readthrough | CGAGTGGGAGTTAAAGCTTCCAGTG*aaggcgacgatgaccggactgtgtg |
| ZBTB16 | NNMT | Readthrough | CGGGACCCCCTCAGCCTCATTTCTG*aagggctgaactgatggaaggaatg |
| KB-1507C5.4 | ATP6V1C1 | Readthrough | TCCATGTCGTAAGTTACACAAGAAG*aatctctcttgatttttgaggaaat |
| PPP1R21 | STON1 | Readthrough | TGACACACTAAAGATGTCCAGTAAg*gagggagcgctctcccctcctctgg |
| SUZ12 | SUZ12P | Pseudogene | GAAACTCCAGAACAAACATCAAAAG*cttgtcagctcatttgcagcttaca |
| SUZ12P | SUZ12 | Pseudogene | AAATGACAGTATTTGATAAAAACAG*aggctgcctccattcgaaacatttt |
| TREM2 | TREML1 | Readthrough | CTGCTCATCTTACTCTTTGTCACAG*catccccttgatctggggtgctgtg |
| TRIM2 | MND1 | Readthrough | CGACTGGGGAAACAGCAGGATCCAG*tcaaagaaaaaaggactgagtgcag |
| AC015977.6 | CIB4 | Readthrough | GGTTCTGCCCAGAAGCCAGCTGCAG*gccctgaccttcctgaccagaaatga |
| AHRR | NCOA2 | GCAAGGTGTACCGATGCCTCCGGGG*ttcaacagaaaattatcttttggaa | |
| CHD4 | NOP2 | Readthrough | GGCACCCGAACCTACCCCACAGCAG*taccatggggcgcaagttggaccct |
| EGFL7 | MCF2L | GGGATGACTGATTCTCCTCCGCCAG*gttggagcaaaacgtcccactcact | |
| GPR65 | LINC01146 | Readthrough | AAACACATCACCGGAAGAAATATGG*atgatgcatatcataaattattact |
| HERC3 | FAM13A-AS1 | Readthrough | AATTCTACATGATTAAAGAATCCAT*ccctttacagaaaacaactgaccaa |
| KB-1572G7.2 | AP000347.4 | Readthrough | ACACCACTCTTCCTGTTGGCCCAAG*gtcagcccaagactaccccgtcggt |
| LCAT | PSMB10 | Readthrough | TGAATAAAGACCTTCCTTTGCTACC*agtacccagtgagcagcacagaggg |
| LSP1 | TNNT3 | Short-distance | CCGGCTCCCTAGGCGTCCCATCTCG*aaaccacccaccttcaccatgtctg |
| LTBP2 | NPC2 | Readthrough | GATGCGGCCCACATGGCCTGCGTAG*gttctgtggatggagttataaagga |
| OSBPL2 | ADRM1 | Readthrough | GGTTGCAAGCTGAGAACATCCAGAG*gaacccaagacagaccaggatgagg |
| PARL | MAP6D1 | Readthrough | ATCTTGGGGGAGCTCTTTTTGGAAT*acaggaattccaggcttggactgga |
| PTPN22 | RSBN1 | Readthrough | AACTCCAGCTCATTTCTGAATTTTG*aaacaccagatgaaaatggtaaaac |
We decided to investigate with molecular methods the described fusion transcripts. No other fusions were examined.
Molecular genetic confirmation of fusions
PCR with the primers TBCK-2558F1 and TBCK-2908R1 amplified a cDNA of the TBCK gene indicating that the synthesized cDNA was of good quality.
RT-PCR using cDNA from the tumor and subsequent direct Sanger sequencing verified the presence of the P4HA2-TBCK, TBCK-P4HA2, AHRR-NCOA2, ETV4-AHRR, and NCOA2-ETV4 fusion transcripts (Table II and Fig. 3). TBCK-P4HA2, AHRR-NCOA2, and NCOA2-ETV4 were in-frame fusions which would code for chimeric proteins. The detected ETV4-AHRR fusion, on the other hand, was out-of-frame and would not produce a chimeric protein, nor would the P4HA2-TBCK code for any functional protein. No EGFL7-MCF2L or CYP1B1-CLU fusion transcript was found by RT-PCR amplification (Table II).
Figure 3.
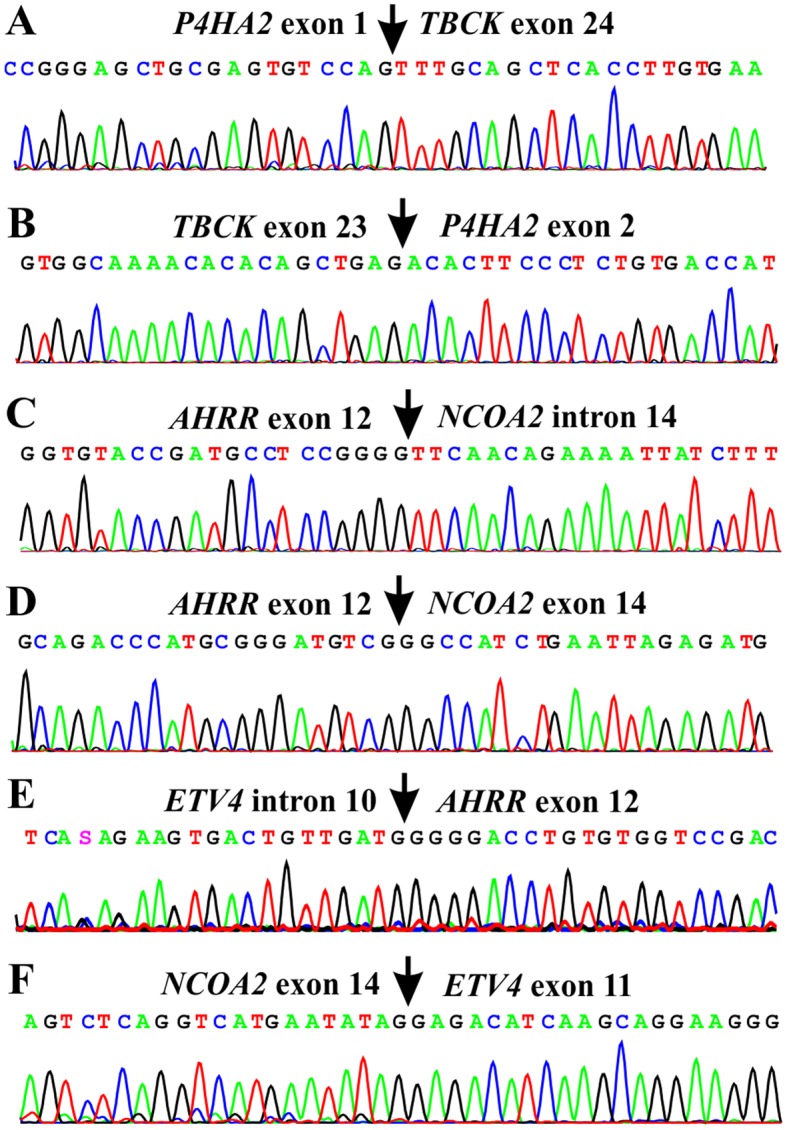
Partial sequence chromatogram of the amplified cDNA fragment showing the junction points of the fusion transcripts. (A) P4HA2-TBCK, (B) TBCK-P4HA2, (C) exon 12 of AHRR with sequence of intron 14 of NCOA2, (D) exon 12 of AHRR with exon 14 of NCOA2, (E) ETV4-AHRR, and (F) NCOA2-ETV4.
Discussion
The examined angiofibroma of soft tissue carried the recurrent AHRR-NCOA2 fusion transcript but lacked the reciprocal NCOA2-AHRR. This finding supports the initial suggestion that AHRR-NCOA2 is the pathogenetically significant fusion transcript in tumors carrying a t(5;8)(p15;q12) (2,3). While we were examining the current tumor, a report was published describing 13 cases of angiofibroma of soft tissue with an AHRR-NCOA2 but with only eight of them carrying the reciprocal NCOA2-AHRR (11). Current data therefore agree that the AHRR-NCOA2 fusion gene is recurrent in angiofibroma of soft tissue [(2,3,11), present case] and indicate that this is the pathogenetically crucial outcome of the t(5;8).
Using FISH on formalin-fixed, paraffin-embedded specimens, Sugita et al (5) found that 16–36% of the tumor cells showed NCOA2 rearrangement. A fairly small proportion of NCOA2 gene rearrangement-positive cells (4–12 split signals per 50 tumor cell nuclei) was recently reported also by Yamada et al (11). The split signals were mostly detected in relatively large, spindle-shaped nuclei, indicating that these were the ones belonging to the neoplastic parenchyma (11).
The present tumor had two cytogenetically unrelated clones: one (eight metaphases) with the translocations t(4;5)(q24;q31) and t(5;8;17)(p15;q13;q21) and another (2 cells) with t(1;14)(p31;q32) as the sole chromosome abnormality. Thus, our data not only are in agreement with previous observations that only a fraction of tumor cells carry the NCOA2 gene rearrangement, but also demonstrate genetic heterogeneity of uncertain pathogenetic significance within the tumor. Although no fusion gene was found corresponding to t(1;14)(p31;q32), this should not lead us to conclude that the translocation was pathogenetically unimportant. The t(1;14)(p31;q32) chromosome aberration may exert its influence through a position effect causing deregulation of a gene in the proximity of the breakpoints. Alternatively, the current methodology may be unable to detect a fusion gene as has been demonstrated (9).
So far, three types of AHRR-NCOA2 fusion transcripts have been described: in the first type, exon 9 of AHRR is joined with exon 16 of NCOA2, the second type shows exon 10 of AHRR being joined to exon 14 of NCOA2, and in the third type there is an insertion of an intronic sequence from the NCOA2 gene between exon 9 of AHRR and exon 14 of NCOA2 (2,11). In the present angiofibroma of soft tissue, two novel fusion transcripts were found with different fusion positions from those previously described: a fusion transcript in which nt 1670 (sequence with accession no. NM_020731) from exon 12 of the AHRR gene was fused with a sequence from intron 14 of NCOA2 and a transcript in which nt 1533 (also from exon 12) of AHRR was fused to exon 15 of NCOA2 (sequence with accession no. NM_006540.2). The resulting putative AHRR-NCOA2 protein would be similar to those reported (2) in as much as the C-terminal part of AHRR is replaced by the C-terminal part of NCOA2.
The involvement of NCOA2 in neoplasia was first reported in acute myeloid leukemia with the cytogenetic inversion inv(8)(p11q13) which resulted in a KAT6A-NCOA2, also known as MOZ-TIF2 fusion gene (12,13). Since then, NCOA2 has been implicated also in other malignancies. A fusion between ETV6 (TEL) and NCOA2 was reported in childhood leukemia with the recurrent t(8;12)(q13;p13) (14). A PAX3-NCOA2 gene was found as a rare variant fusion in alveolar rhabdomyosarcoma; it was brought about by a t(2;8)(q35;q13) translocation (15). A HEY1-NCOA2 fusion gene was described in mesenchymal chondrosarcomas (16,17). Recently, SRF-NCOA2, TEAD1-NCOA2, and VGLL2-NCOA2 fusions were reported in rhabdomyosarcomas (18,19). In all the above-mentioned fusions, NCOA2 is the 3′-partner gene and all fusion proteins contain the two C-terminal activation domains AD1/CID (activation domain 1/CREB-binding protein interacting domain) and AD2 (2,3,12–19). The transforming activities of KAT6A-NCOA2 and PAX3-NCOA2 have been demonstrated experimentally (15,20). In addition, KAT6A-NCOA2 was shown to induce acute myeloid leukemia in transgenic fish (21). Deguchi et al (20) showed that the KAT6A-NCOA2 interaction with CREBBP through AD1/CID is essential for transformation. Similarly, Sumegi et al (15) showed that while deletion of the AD2 portion of PAX3-NCOA2 fusion protein reduced the transforming activity, deletion of the AD1/CID domain fully abrogated the transforming activity of the chimeric protein. Thus, the AD1/CID and AD2 domains of NCOA2 seem to be essential for the transformation ability of the various fusion proteins.
The three-way translocation t(5;8;17)(p15;q13;q21) of the present case not only generated an AHRR-NCOA2 resulting from the translocation of 5p15 to 8q13, but also two additional fusion genes: an NCOA2-ETV4, stemming from the moving of 8q13 to 17q21, and an ETV4-AHRR, generated by the moving of 17q21 to 5p15. The detected ETV4-AHRR fusion transcript is out-of-frame and so cannot produce a chimeric protein. The NCOA2-ETV4 fusion transcript is in-frame coding for a chimeric NCOA2-ETV4 protein, the oncogenetic potential of which cannot be ruled out. Based on the NCOA2 and ETV4 proteins with accession nos. NP_006531.1 and NM_001986.2, respectively, the chimeric NCOA2-ETV4 would contain 1,175 amino acids. The NCOA2 N-terminal part of the protein would contain the helix-loop-helix, PAS_9 and PAS_11, the CITED, and the SRC-1 domains. The ETV4 C-terminal part would contain the ETS DNA-binding domain of ETV4 (Fig. 4).
Figure 4.

Illustration of the proteins AHRR, NCOA2, AHRR-NCOA2, NCOA2-ETV4, and TBCK-P4H2A. The domains, their accession nos., and intervals are also shown. Arrows in the AHRR protein indicate the known fusion points for the published AHRR-NCOA2 proteins. Arrows in the AHRR-NCOA2, NCOA2-ETV4, and TBCK-P4H2A show the fusion points in the present angiofibroma of soft tissue.
ETV4 was reported to contribute the 3′-part of the oncogenic protein in the subset of Ewing's sarcomas characterized by a t(17;22)(q12;q12) translocation (22,23). The EWSR1-ETV4 protein, in which the N-terminal part of EWSR1 is fused to the ETS DNA-binding domain of ETV4, has an oncogenetic potential similar to that of the EWSR1-FLI1, EWSR1-ERg, EWSR1-FEV, and EWSR1-ETV1 fusion proteins which may also be found in Ewing's sarcoma (24). The ETV4 gene was also described as the 3′-partner in fusion genes found in prostate carcinoma (25–27). ETV4 was found to fuse with the TMPRSS2, KLK2, CANT1, and DDX5 (25–27). All these fusions genes, TMPRSS2-ETV4, KLK2-ETV4, CANT1-ETV4, and DDX5-ETV4, contain (like the present NCOA2-ETV4) the part of ETV4 coding for the ETS DNA-binding domain.
The chromosome translocation t(4;5)(q24;q31) generated the P4HA2-TBCK and TBCK-P4HA2 fusion transcripts. P4HA2-TBCK does not encode any functional protein, whereas TBCK-P4HA2 encodes a chimeric 1,335-amino acid protein. TBCK-P4HA2 would contain the first 794 out of 830 amino acids of the TBCK protein (accession no. NP_149106.2), 6 amino acids from the untranslated region of exon 2 of P4HA2 (accession no. NM_004199.2), and the entire 535 amino acid-P4HA2 protein (NP_004190.1). The function of this putative chimeric protein is difficult to predict since it would contain both the protein kinase domain, the Rhodanese-like domain, and the Tre-2/Bub2/Cdc16 (TBC) domain of TBCK together with the P4HA2 protein which is a component of the prolyl 4-hydroxylase. The TBCK protein is thought to play a role in actin organization, cell growth, and cell proliferation by regulating the mammalian target of the rapamycin (mTOR) signaling pathway. This protein may also be involved in the transcriptional regulation of the components of the mTOR complex (http://www.ncbi.nlm.nih.gov/gene/93627). Depletion of TBCK significantly inhibits cell proliferation, reduces cell size, and disrupts the organization of actin but not microtubule. Knockdown of TBCK induces a significant decrease in the protein levels of components of mTOR complex (mTORC) and suppresses the activity of mTOR signaling, but not the MAPK or PDK1/Akt pathway (28).
The protein encoded by the P4HA2 gene is one of several different types of α subunit of the prolyl 4-hydroxylase and provides the major part of the catalytic site of the active enzyme (http://www.ncbi.nlm.nih.gov/gene/8974). In collagen and related proteins, prolyl 4-hydroxylase catalyzes the formation of 4-hydroxyproline that is essential to the proper three-dimensional folding of newly synthesized procollagen chains. In breast cancer, P4HA2 was shown to promote progression and metastasis by regulating collagen deposition (29). In squamous cell carcinoma of the oral cavity, P4HA2 was identified as a metastasis associated protein (30).
In spite of the now repeatedly documented recurrence of AHRR-NCOA2 in angiofibroma of soft tissue [present case, (2,11)], our findings indicate that also additional genetic events, some of which lead to fusion genes, may be important in tumor development. Worthy of mention is that of the eight hitherto cytogenetically reported tumors, including the present case, three had three-way translocations (1–3). What lies behind this highly unusual feature is unknown. Obviously, more such tumors must be studied cytogenetically and molecularly before all important aspects of their pathogenesis are laid bare.
Acknowledgments
The authors would like to thank Hege Kilen Andersen and Nina Øino for their excellent technical assistance. This study was supported by grants from the Norwegian Radium Hospital Research Foundation.
References
- 1.Mariño-Enríquez A, Fletcher CD. Angiofibroma of soft tissue: Clinicopathologic characterization of a distinctive benign fibrovascular neoplasm in a series of 37 cases. Am J Surg Pathol. 2012;36:500–508. doi: 10.1097/PAS.0b013e31823defbe. [DOI] [PubMed] [Google Scholar]
- 2.Jin Y, Möller E, Nord KH, Mandahl N, Von Steyern FV, Domanski HA, Mariño-Enríquez A, Magnusson L, Nilsson J, Sciot R, et al. Fusion of the AHRR and NCOA2 genes through a recurrent translocation t(5;8)(p15;q13) in soft tissue angiofibroma results in upregulation of aryl hydrocarbon receptor target genes. Genes Chromosomes Cancer. 2012;51:510–520. doi: 10.1002/gcc.21939. [DOI] [PubMed] [Google Scholar]
- 3.Arbajian E, Magnusson L, Mertens F, Domanski HA, Vult von Steyern F, Nord KH. A novel GTF2I/NCOA2 fusion gene emphasizes the role of NCOA2 in soft tissue angiofibroma development. Genes Chromosomes Cancer. 2013;52:330–331. doi: 10.1002/gcc.22033. [DOI] [PubMed] [Google Scholar]
- 4.Schoolmeester JK, Sukov WR, Aubry MC, Folpe AL. Angiofibroma of soft tissue: Core needle biopsy diagnosis, with cytogenetic confirmation. Am J Surg Pathol. 2012;36:1421–1423. doi: 10.1097/PAS.0b013e3182627ccb. [DOI] [PubMed] [Google Scholar]
- 5.Sugita S, Aoyama T, Kondo K, Keira Y, Ogino J, Nakanishi K, Kaya M, Emori M, Tsukahara T, Nakajima H, et al. Diagnostic utility of NCOA2 fluorescence in situ hybridization and Stat6 immunohistochemistry staining for soft tissue angiofibroma and morphologically similar fibrovascular tumors. Hum Pathol. 2014;45:1588–1596. doi: 10.1016/j.humpath.2013.12.022. [DOI] [PubMed] [Google Scholar]
- 6.Fukuda Y, Motoi T, Kato I, Ikegami M, Funata N, Ohtomo R, Horiguchi S, Goto T, Hishima T. Angiofibroma of soft tissue with fibrohistiocytic features and intratumor genetic heterogeneity of NCOA2 gene rearrangement revealed by chromogenic in situ hybridization: A case report. Pathol Int. 2014;64:237–242. doi: 10.1111/pin.12163. [DOI] [PubMed] [Google Scholar]
- 7.Edgar MA, Lauer SR, Bridge JA, Rizzo M. Soft tissue angiofibroma: Report of 2 cases of a recently described tumor. Hum Pathol. 2013;44:438–441. doi: 10.1016/j.humpath.2012.08.021. [DOI] [PubMed] [Google Scholar]
- 8.Schaffer LG, McGowan-Jordan J, Schmid M, editors. ISCN 2013: An International System for Human Cytogenetic Nomenclature (2013): Recommendations of the International Standing Committee on Human Cytogenetic Nomenclature. 1st edition. S. Karger AG; Basel: 2013. [Google Scholar]
- 9.Panagopoulos I, Gorunova L, Bjerkehagen B, Heim S. The ʻgrepʼ command but not FusionMap, FusionFinder or ChimeraScan captures the CIC-DUX4 fusion gene from whole transcriptome sequencing data on a small round cell tumor with t(4;19)(q35;q13) PLoS One. 2014;9:e99439. doi: 10.1371/journal.pone.0099439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Nicorici D, Satalan M, Edgren H, Kangaspeska S, Murumagi A, Kallioniemi O, Virtanen S, Kilkku O. FusionCatcher - a tool for finding somatic fusion genes in paired-end RNA-sequencing data. bioRxiv 011650. http://dx.doi.org/10.1101/011650.
- 11.Yamada Y, Yamamoto H, Kohashi K, Ishii T, Iura K, Maekawa A, Bekki H, Otsuka H, Yamashita K, Tanaka H, et al. Histological spectrum of angiofibroma of soft tissue: Histological and genetic analysis of 13 cases. Histopathology. 2016 Feb 4; doi: 10.1111/his.12943. Epub ahead of print. [DOI] [PubMed] [Google Scholar]
- 12.Carapeti M, Aguiar RC, Goldman JM, Cross NC. A novel fusion between MOZ and the nuclear receptor coactivator TIF2 in acute myeloid leukemia. Blood. 1998;91:3127–3133. [PubMed] [Google Scholar]
- 13.Carapeti M, Aguiar RC, Watmore AE, Goldman JM, Cross NC. Consistent fusion of MOZ and TIF2 in AML with inv(8)(p11q13) Cancer Genet Cytogenet. 1999;113:70–72. doi: 10.1016/S0165-4608(99)00007-2. [DOI] [PubMed] [Google Scholar]
- 14.Strehl S, Nebral K, König M, Harbott J, Strobl H, Ratei R, Struski S, Bielorai B, Lessard M, Zimmermann M, et al. ETV6-NCOA2: A novel fusion gene in acute leukemia associated with coexpression of T-lymphoid and myeloid markers and frequent NOTCH1 mutations. Clin Cancer Res. 2008;14:977–983. doi: 10.1158/1078-0432.CCR-07-4022. [DOI] [PubMed] [Google Scholar]
- 15.Sumegi J, Streblow R, Frayer RW, Dal Cin P, Rosenberg A, Meloni-Ehrig A, Bridge JA. Recurrent t(2;2) and t(2;8) translocations in rhabdomyosarcoma without the canonical PAX-FOXO1 fuse PAX3 to members of the nuclear receptor transcriptional coactivator family. Genes Chromosomes Cancer. 2010;49:224–236. doi: 10.1002/gcc.20731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Panagopoulos I, Thorsen J, Gorunova L, Micci F, Heim S. Sequential combination of karyotyping and RNA-sequencing in the search for cancer-specific fusion genes. Int J Biochem Cell Biol. 2014;53:462–465. doi: 10.1016/j.biocel.2014.05.018. [DOI] [PubMed] [Google Scholar]
- 17.Wang L, Motoi T, Khanin R, Olshen A, Mertens F, Bridge J, Dal Cin P, Antonescu CR, Singer S, Hameed M, et al. Identification of a novel, recurrent HEY1-NCOA2 fusion in mesenchymal chondrosarcoma based on a genome-wide screen of exon-level expression data. Genes Chromosomes Cancer. 2012;51:127–139. doi: 10.1002/gcc.20937. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Alaggio R, Zhang L, Sung YS, Huang SC, Chen CL, Bisogno G, Zin A, Agaram NP, LaQuaglia MP, Wexler LH, et al. A molecular study of pediatric spindle and sclerosing rhabdomyosarcoma: Identification of novel and recurrent VGLL2-related fusions in infantile cases. Am J Surg Pathol. 2016;40:224–235. doi: 10.1097/PAS.0000000000000538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Mosquera JM, Sboner A, Zhang L, Kitabayashi N, Chen CL, Sung YS, Wexler LH, LaQuaglia MP, Edelman M, Sreekantaiah C, et al. Recurrent NCOA2 gene rearrangements in congenital/infantile spindle cell rhabdomyosarcoma. Genes Chromosomes Cancer. 2013;52:538–550. doi: 10.1002/gcc.22050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Deguchi K, Ayton PM, Carapeti M, Kutok JL, Snyder CS, Williams IR, Cross NC, Glass CK, Cleary ML, Gilliland DG. MOZ-TIF2-induced acute myeloid leukemia requires the MOZ nucleosome binding motif and TIF2-mediated recruitment of CBP. Cancer Cell. 2003;3:259–271. doi: 10.1016/S1535-6108(03)00051-5. [DOI] [PubMed] [Google Scholar]
- 21.Zhuravleva J, Paggetti J, Martin L, Hammann A, Solary E, Bastie JN, Delva L. MOZ/TIF2-induced acute myeloid leukaemia in transgenic fish. Br J Haematol. 2008;143:378–382. doi: 10.1111/j.1365-2141.2008.07362.x. [DOI] [PubMed] [Google Scholar]
- 22.Kaneko Y, Yoshida K, Handa M, Toyoda Y, Nishihira H, Tanaka Y, Sasaki Y, Ishida S, Higashino F, Fujinaga K. Fusion of an ETS-family gene, EIAF, to EWS by t(17;22)(q12;q12) chromosome translocation in an undifferentiated sarcoma of infancy. Genes Chromosomes Cancer. 1996;15:115–121. doi: 10.1002/(SICI)1098-2264(199602)15:2<115::AID-GCC6>3.0.CO;2-6. [DOI] [PubMed] [Google Scholar]
- 23.Urano F, Umezawa A, Hong W, Kikuchi H, Hata J. A novel chimera gene between EWS and E1A-F, encoding the adenovirus E1A enhancer-binding protein, in extraosseous Ewing's sarcoma. Biochem Biophys Res Commun. 1996;219:608–612. doi: 10.1006/bbrc.1996.0281. [DOI] [PubMed] [Google Scholar]
- 24.Braunreiter CL, Hancock JD, Coffin CM, Boucher KM, Lessnick SL. Expression of EWS-ETS fusions in NIH3T3 cells reveals significant differences to Ewing's sarcoma. Cell Cycle. 2006;5:2753–2759. doi: 10.4161/cc.5.23.3505. [DOI] [PubMed] [Google Scholar]
- 25.Tomlins SA, Mehra R, Rhodes DR, Smith LR, Roulston D, Helgeson BE, Cao X, Wei JT, Rubin MA, Shah RB, et al. TMPRSS2:ETV4 gene fusions define a third molecular subtype of prostate cancer. Cancer Res. 2006;66:3396–3400. doi: 10.1158/0008-5472.CAN-06-0168. [DOI] [PubMed] [Google Scholar]
- 26.Han B, Mehra R, Dhanasekaran SM, Yu J, Menon A, Lonigro RJ, Wang X, Gong Y, Wang L, Shankar S, et al. A fluorescence in situ hybridization screen for E26 transformation-specific aberrations: Identification of DDX5-ETV4 fusion protein in prostate cancer. Cancer Res. 2008;68:7629–7637. doi: 10.1158/0008-5472.CAN-08-2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Hermans KG, Bressers AA, van der Korput HA, Dits NF, Jenster G, Trapman J. Two unique novel prostate-specific and androgen-regulated fusion partners of ETV4 in prostate cancer. Cancer Res. 2008;68:3094–3098. doi: 10.1158/0008-5472.CAN-08-0198. [DOI] [PubMed] [Google Scholar]
- 28.Liu Y, Yan X, Zhou T. TBCK influences cell proliferation, cell size and mTOR signaling pathway. PLoS One. 2013;8:e71349. doi: 10.1371/journal.pone.0071349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Xiong G, Deng L, Zhu J, Rychahou PG, Xu R. Prolyl-4-hydroxylase α subunit 2 promotes breast cancer progression and metastasis by regulating collagen deposition. BMC Cancer. 2014;14:1. doi: 10.1186/1471-2407-14-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Chang KP, Yu JS, Chien KY, Lee CW, Liang Y, Liao CT, Yen TC, Lee LY, Huang LL, Liu SC, et al. Identification of PRDX4 and P4HA2 as metastasis-associated proteins in oral cavity squamous cell carcinoma by comparative tissue proteomics of microdissected specimens using iTRAQ technology. J Proteome Res. 2011;10:4935–4947. doi: 10.1021/pr200311p. [DOI] [PubMed] [Google Scholar]