

Abstract
Multiple pluripotent cell populations, which together comprise the pluripotent cell lineage, have been identified. The mechanisms that control the progression between these populations are still poorly understood. The formation of early primitive ectoderm-like (EPL) cells from mouse embryonic stem (mES) cells provides a model to understand how one such transition is regulated. EPL cells form from mES cells in response to l-proline uptake through the transporter Slc38a2. Using inhibitors of cell signaling we have shown that Src family kinases, p38 MAPK, ERK1/2 and GSK3β are required for the transition between mES and EPL cells. ERK1/2, c-Src and GSK3β are likely to be enforcing a receptive, primed state in mES cells, while Src family kinases and p38 MAPK are involved in the establishment of EPL cells. Inhibition of these pathways prevented the acquisition of most, but not all, features of EPL cells, suggesting that other pathways are required. L-proline activation of differentiation is mediated through metabolism and changes to intracellular metabolite levels, specifically reactive oxygen species. The implication of multiple signaling pathways in the process suggests a model in which the context of Src family kinase activation determines the outcomes of pluripotent cell differentiation.
Introduction
The pluripotent cell lineage in the mouse embryo is founded in the forming blastocyst and develops through a series of functionally distinct intermediate populations before differentiating at gastrulation. Four identifiable pluripotent cell populations, or states, have been identified in vivo–the epiblast precursor cell, the epiblast of the Inner Cell Mass (ICM), and the early and late epiblast of the post-implantation embryo [1–3]. Culture equivalents of these populations have been established from mouse. Primed embryonic stem (mES) cells [4, 5] and epiblast stem cells (EpiSCs) [6–8], which represent the epiblast of the ICM and the late epiblast of the post-implantation embryo, respectively, have been isolated directly from the embryo. Naive ES cells, an in vitro equivalent of the early epiblast of the ICM, have been formed from primed mES cells in culture [9–12]. Lastly, the epiblast, or primitive ectoderm, of the early post-implantation embryo can be formed in culture through the differentiation of primed mES cells to early primitive ectoderm-like (EPL) cells [13],[14, 15]. EpiSC-like cells can also be derived from mES cells by culture in FGF and Activin A [16–18]. These populations of pluripotent cells are now well recognized, but the molecular mechanisms that regulate progression between them are not well understood.
EPL cell formation occurs when mES cells are cultured in MEDII, medium conditioned by HepG2 cells [13–15], or in medium containing the active component of MEDII, l-proline [19–22]. Expression of Oct4, Sox2 and alkaline phosphatase, and a differentiation potential in culture that includes mesoderm, endoderm and ectoderm, identifies EPL cells as pluripotent [13, 23–28]. The changes in colony morphology, gene expression [13, 14, 24, 29], proliferation rate [20], and developmental potential [15, 24, 25] that accompany EPL cell formation identify these cells as primitive ectoderm-like. EPL cell formation is dependent on elevated concentrations of l-proline within the medium (> 100 μM) [19, 20], and is inhibited by LIF [13]. The uptake of l-proline through the sodium-coupled neutral amino acid transporter 2 (Slc38a2, also known as SNAT2) on the surface of the cells is required for activity, and the inhibition of l-proline uptake through SNAT2 prevents EPL cell formation [19]. Collectively, these studies describe a system that models the transition from the epiblast of the ICM to early primitive ectoderm, and that can be used to understand the regulation of this event.
Little is known of how the internalization of l-proline by ES cells, when presented in MEDII or added exogenously, induces EPL cell formation. Changes in cell morphology characteristic of the system have been shown to require the metabolism of l-proline and generation of reactive oxygen species (ROS) [21]. Here we consider the role of signaling pathways in EPL cell formation. We describe the effect of pharmacologically inhibiting the Src family kinases and mitogen-activated protein kinase (MAPK) pathways (p38 MAPK and Extracellular signal-regulated kinases 1 and 2 (ERK1/2)) on the formation and maintenance of EPL cells. We show that inhibition of Src family kinases and p38 MAPK pathways, and pathways implicated in naïve cell formation, affected the formation and maintenance of EPL cells. Inhibition of a single pathway could not completely prevent EPL cell formation, suggesting the requirement of multiple signaling pathways in the process. These data have been used to develop a model for the process of pluripotent cell lineage progression and the formation of the primitive ectoderm based on a metabolic switch and increasing intracellular ROS.
Results
Inhibition of ERK1/2 signaling prevents EPL cell formation and maintenance in culture
EPL cells are routinely formed from primed mES cells in culture [13, 20]. The inhibition of ERK activity in mES cells promotes the transition of primed mES cells to the naive state [9, 11]. The ability of MEDII to induce EPL cells from a mES cell population in which MEK1 signalling had been inhibited, and therefore lacking phosphorylated ERK1/2, was tested. Phosphorylated (p)ERK2 was detected in ES cells; phosphorylation was lost in cells cultured with the MEK inhibitor PD0325901 [30](Table 1; Fig 1A). Addition of PD0325901 to ES cells in conjunction with MEDII or l-proline prevented the adoption of EPL cell-like colony morphology (Fig 1B and 1C; data not shown). The cells were analysed by qPCR for the expression of markers of pluripotency (Oct4, Sox2 and Nanog), the early epiblast of the ICM/ES cells (Rex1, Spp1 and Gbx2) and the primitive ectoderm/EPL cells (Dnmt3b, Otx2 and Fgf5; Fgf5 was only examined in cells cultured in MEDII [20]). Cells cultured in MEDII + PD0325901 failed to up regulated Fgf5 and Dnmt3b expression and maintained Nanog, Rex1 and Spp1 expression when compared to cells cultured in MEDII, consistent with maintenance of an ES cell-like state in the absence of MEK activity (Fig 1D).
Table 1. Summary of inhibitors used in this study.
| Compound | Chemical name | [μM] | Type of inhibition | Specificity | References1 |
|---|---|---|---|---|---|
| PD0325901 | N-[(2R)-2,3-dihydroxypropoxy]-3,4-difluoro-2-[(2-fluoro-4-iodophenyl)amino]-benzamide | 1 | Non-ATP competitive binding inhibition | MEK12,4 | [9, 11, 31–33] |
| PP2 | Pyrazolo-pyrimidine 4-amino-5-(4-chlorophenyl)-7-(t-butyl)pyrazolo[3,4-d]pyrimidine | 10 | ATP competitive binding inhibition | Src4, p56Lck,4, p59FynT,4, Hck4, EGFR5, RIP25, CK1δ5 and GAK5.3 | [34–39] |
| SB203580 | 4-(4´-fluorophenyl)-2-(4´-methylsulfinylphenyl)-5-(4´-pyridyl)-imidazole | 10 | ATP competitive binding inhibition | p38α MAPK4, p38β MAPK4 and CK1δ5. | [31–33, 40–44] |
| SU6656 | 2-oxo-3-(4,5,6,7-tetrahydro-1 H-indol-2-ylmethylene)-2,3-dihydro-1H-indole-5-sulfonic acid dimethylamide | ATP competitive binding inhibition | MAPKAP-K1a4, AMPK4, PHK5, p56Lck,4 and DYRK1A4. | [34, 45, 46]; |
1 Selected references, prioritized to include those reporting on role of inhibitor in ES cell renewal and differentiation.
2 Inhibitor is reported to have activity on MEK1 but not on ERK1, ERK2, p38αMAPK or p38βMAPK [34].
3 PP2 has been shown to have a modest impact on the activity of p38α MAPK, p38β MAPK [34].
4 Detected in ES cells.
5 Not reported in ES cells.
Fig 1. Inhibition of MEK1 prevents the formation of EPL cells in response to MEDII.

A. mES cells were pre-treated with 1 μM PD0325901 for 60 minutes. 200 μM l-proline was added, as denoted, and the cells incubated for a further 60 minutes. Cells were collected and analysed by western blot for the presence of phosphorylated ERK1 or ERK2. Total ERK1/2 was used as a loading control. The intensity of the pERK2 band was measured using Quantity One software (BioRad) and represented as a proportion of total ERK1/2. Error bars represent SEM; n = 4; * p ≤ 0.05 when compared to mES cells. B-C. mES cells were cultured in MEDII- and DMSO-containing medium (B) and MEDII- and 1μM PD0325901-contianing medium (C) for 3 days. Scale bar = 200 μm. D. MEDII- and DMSO-containing medium (■) and MEDII- and 1μM PD0325901-contianing medium (□) for 3 days. RNA from these cells was analyzed for expression of Oct4, Sox2, Nanog, Rex1, Spp1, Gbx2, Dnmt3b, Otx2 and Fgf5 by real-time PCR. Expression was normalized to β-actin and expressed relative to mES cells (Fgf5 has been expressed relative to MEDII + DMSO). Error bars represent SEM; n = 3. mES cells + MEDII + PD0325901 were compared to mES cells + MEDII + DMSO (** p ≤ 0.05) or mES cells (# p ≤ 0.05).
EPL cell formation requires signaling through SRC family kinases
There are nine members of the Src family kinases, seven of which have been shown to be expressed in mES cells; c-Src, p56Lck, p59FynT, Hck, Lyn, Fgr and Yes [30, 38, 47](JK, EW and JR, personal communication), and c-Src activity is required for the formation of a primitive ectoderm-like cell from mES cells in culture [38, 39]. PP2, at a concentration of 10 μM, was used to inhibit Src, p56Lck, p59FynT and Hck in mES cells (Table 1). At this concentration the phosphorylation of Src family members was reduced (Fig 2A) and cells did not adopt an EPL cell colony morphology in response to MEDII (S1 Fig). The addition of PP2 reduced colony size, which was likely a consequence of the inhibition of pluripotent cell proliferation (S1 Fig). The addition of PP2 maintained or increased the expression of pluripotent and mES cell markers, with the exception of Gbx2, in mES cells cultured in MEDII- or l-proline-containing medium, and reduced the expression of Dnmt3b and Fgf5, but not Otx2, (Fig 2D and S2 Fig).
Fig 2.

Src Family Kinases are required for the formation of EPL cells in response to MEDII A. mES cells were cultured with or without l-proline and PP2, as denoted. Total protein was extracted after 24 hours and analysed by western blot for the presence of phosphorylation of Src family proteins (pSFK). β-tubulin was used to normalise for protein loaded. B, C. mES cells were cultured in medium supplemented with MEDII and DMSO (A) or MEDII and 10 μM PP2 (B) for 3 days. Scale bar = 200 μm. D. mES cells were cultured in medium supplemented with MEDII and DMSO (■) or MEDII and 10 μM PP2 (□), for 3 days. RNA from these cells was analyzed for transcripts of Oct4, Sox2, Nanog, Rex1, Spp1, Gbx2, Dnmt3b, Otx2 and Fgf5 by real-time PCR. Expression was normalized to β-actin and expressed relative to ES cells (Fgf5 has been expressed relative to MEDII + DMSO). Error bars represent SEM; n = 3. ES cells in MEDII + PP2 were compared to cells cultured in MEDII + DMSO (** p ≤ 0.05) or ES cells (# p ≤ 0.05). E. mES cells were cultured in ES cell medium supplemented with MEDII, DMSO or MEDII and 10 μM PP2 for 3 days and formed into EBs. EBs were collected on days 2, 3 and 4, RNA was isolated and analyzed for expression of T, Mixl1 and Gapdh by RT-PCR. n = 3.
When differentiated as embryoid bodies (EBs), EPL cells increase expression of differentiation markers earlier than mES cells [20, 24]. mES cells and EPL cells were aggregated to form EBs and analysed on days 2, 3 and 4 for the expression of markers of the primitive streak (T, Mixl1, Fgf8, Wnt3 and Tgfβ1) and nascent mesoderm (BMP4) by RT-PCR (Fig 2D). Differentiation markers were up regulated approximately 48 hours earlier in EBs derived from EPL cells when compared to those derived from mES cells. EBs derived from mES cells cultured in MEDII supplemented with PP2 up regulated differentiation markers approximately 24 hours earlier than mES cell-derived EBs (Fig 2E, S1 Fig). The advancement of differentiated gene expression in EBs derived from cells cultured in l-proline has always been much less than in EBs derived from EPL cells [20]. Despite this, the addition of PP2 to the cells prior to the formation of EBs did impact differentiation kinetics, with a delay in the expression of the differentiation marker T (S2 Fig).
p38 MAP Kinase in EPL cell formation
A kinome screen of mES cells exposed to l-proline showed increased phosphorylation of p38α MAPK, p38β MAPK and Heat shock protein 2 (Hspb2, also known as Hsp27) (S3A Fig), suggesting that the p38 MAPK pathway could be activated [48, 49]. MAP kinase pathways have been implicated in early mES cell differentiation decisions and the formation of the germ lineages in culture [31–33, 40–44]. Western blot showed the presence of pHspb2 in mES cells and mES cell treated with l-proline (Fig 3A). Phosphorylated Hspb2 levels were significantly reduced in by the inhibitor SB203580 (Fig 3A).
Fig 3. The role of p38 MAPK in the formation of EPL cells in response to MEDII.

A. mES cells were pre-treated with 10 μM SB203580 for 60 minutes. 200 μM l-proline was added, as denoted and the cells incubated for a further 60 minutes. Cells were collected and analysed by western blot for the presence of phosphorylated pHspb2. Total Hspb2 was used as a loading control. B, C. mES cells were cultured medium supplemented with MEDII and DMSO (A) or MEDII and 10 μM SB203580 for 3 days. Scale bar = 200 μm. D. mES cells were cultured in medium supplemented with MEDII and DMSO (■) or MEDII and 10 μM SB203580 (□) for 3 days. RNA from these cells was analyzed for expression of Oct4, Sox2, Nanog Rex1, Spp1, Gbx2, Dnmt3b, Otx2 and Fgf5 by real-time PCR. Expression was normalized to β-actin and expressed relative to mES cells (Fgf5 has been expressed relative to MEDII + DMSO). Error bars represent SEM; n = 4. mES cells + MEDII + SB203580 were compared to mES cells + MEDII + DMSO (** p ≤ 0.05) or mES cells (# p ≤ 0.05). E. mES cells were cultured in ES cell medium supplemented with MEDII, DMSO or MEDII and 10 μM 10 μM SB203580 for 3 days and formed into EBs. EBs were collected on days 2, 3 and 4, RNA was isolated and analyzed for expression of T and Gapdh by RT-PCR. n = 3.
The addition of 10 μM SB203580 during EPL cell formation resulted in subtle changes in colony morphology across the population, with many colonies having a more rounded appearance and refractive edges, characteristics of mES cell colonies and not EPL cell colonies (Fig 3B and 3C; data not shown). The expression of Dnmt3b and Fgf5, but not Otx2, was reduced with the addition of SB203580 to cells cultured in MEDII-containing or l-proline-containing medium (Fig 3D and S2 Fig). These data suggest that p38 MAPK was required for the increased expression of these genes. The expression of Rex1, Spp1 and Gbx2 was generally unaffected by the addition of SB203580 and was not restored to levels comparable those in untreated mES cells. mES cells cultured in MEDII- or l-proline-containing medium and SB203580 were differentiated as EBs and the expression of T analysed (Fig 3E and S2 Fig). SB203580 prevented the acquisition of EPL cell differentiation kinetics.
mES cells that lack the p38α MAPK isoform have been established [50]. p38α-/-cells showed equivalent expression of Nanog, Rex1 and Spp1 when compared to the parental line, but higher expression of Gbx2 and significantly lower basal expression of Dnmt3b, Otx2 and Fgf5 (S3B Fig). The addition of MEDII to p38α-/- mES cells resulted in increased expression of the primitive ectoderm markers. These cells, however, failed to down-regulation the ES cell markers (S3C Fig). These data suggest that the loss of p38α affects the formation of EPL cells from mES cells but did not recapitulate the addition of SB203580.
The role of signaling pathways in the maintenance of EPL cells in culture
Signaling through the ERK2, Src family kinase and p38 MAPK all play a role in the formation of EPL cells from mES cells. Signaling inhibitors were added to extant EPL cells to assess the roles these pathways may play in EPL cell maintenance. Inhibition of MEK1/ERK2 in EPL cells resulted in a mixed population of pluripotent and differentiated cell colonies, with alkaline phosphatase positive cell colonies, adopting a compact, 3-dimensional structure rather than an epithelial structure (Fig 4A and 4B). The population showed a significant up regulation of Nanog, Rex1 and Spp1 expression, maintenance of Oct4 and Sox2 expression and a reduction in EPL cell marker expression when compared to EPL cells (Fig 4E). The addition of PP2 to EPL cells also disrupted colony morphology, with a number, but not all, colonies within the culture rounding up (Fig 4C); these colonies were positive for alkaline phosphatase activity. These cells expressed significantly more Nanog and Sox2 compared to EPL cells. Increased expression of Rex1 and Spp1, and decreased expression of Dnmt3b and Fgf5, in these cells was consistent with the cells acquiring a more ES cell-like gene expression profile (Fig 4F). These changes did not all reach parity with ES cell gene expression (data not shown). The expression of Gbx2 and Otx2 was unaffected by the addition of PP2. EPL cells in which p38 MAPK had been inhibited appeared to be more differentiated than controls and there were qualitatively fewer alkaline phosphatase cells (Fig 4D). Gene expression analysis detected no significant differences in the expression of the pluripotent, ES cell or EPL cell markers (Fig 4G), suggesting that an EPL cell gene expression profile was maintained. Despite the appearance of more differentiated cells within this population, gene expression analysis could not detect a role for p38 MAPK in EPL cell maintenance.
Fig 4. The role of ERK1/2, Src Family Kinases and p38 MAPK in the maintenance of EPL cells.
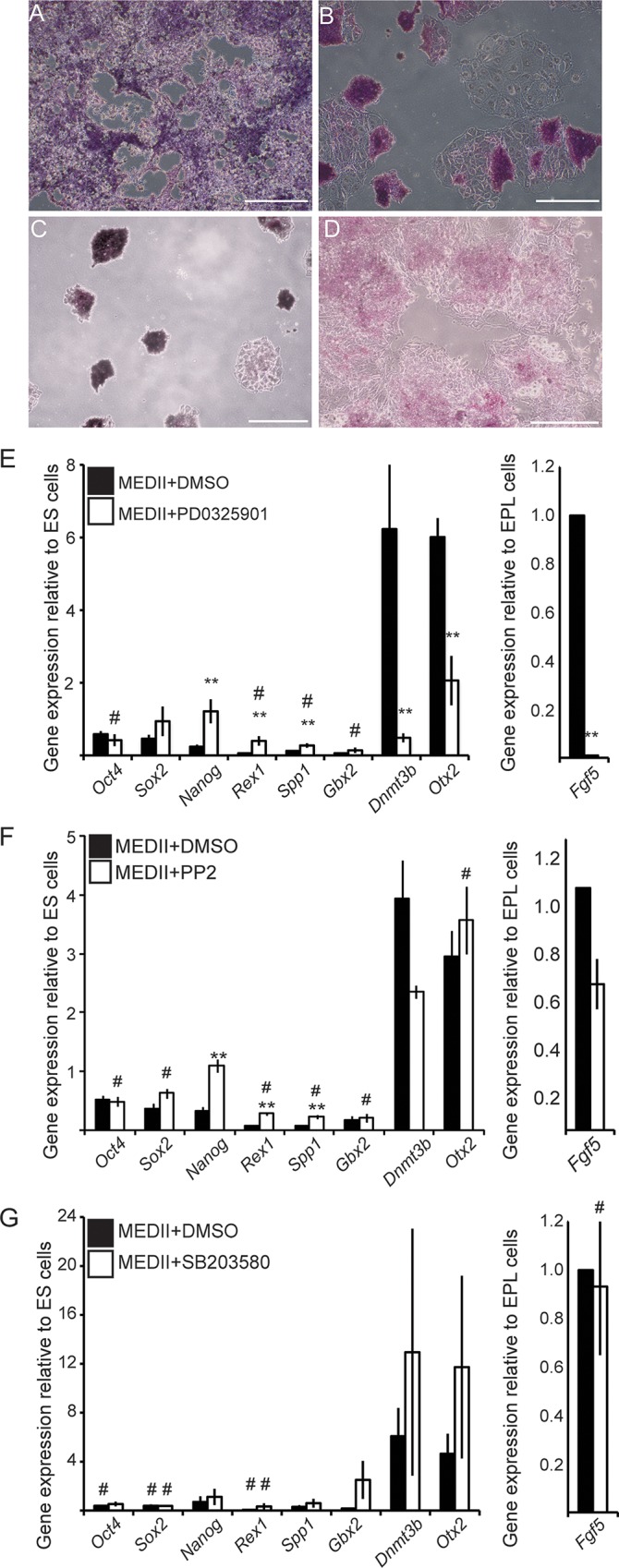
A-D. Photomicrographs of mES cells cultured in MEDII for 2 days and subsequently in MEDII containing medium supplemented with DMSO (A), 1 μM PD0325901 (B) 10 μM PP2 (C) and 10 μM SB203580 (D). Cells were stained for alkaline phosphatase activity (red/purple stain). Scale bar = 200 μm. E, F. mES cells were cultured in MEDII for 2 days and subsequently in MEDII containing medium supplemented with DMSO (■), DMSO with 1 μM PD0325901 (E, □), DMSO with 10 μM PP2 (F, □) or DMSO with 10 μM SB203580 (G, □). RNA from these cells was analyzed for expression of Oct4, Sox2, Nanog, Rex1, Spp1, Gbx2, Dnmt3b and Otx2 by real-time PCR. Expression was normalized to β-actin. Error bars represent SEM; n = 3. EPL cells cultured in MEDII with inhibitor were compared to EPL cells in MEDII with DMSO, ** p ≤ 0.05, or mES cells, # p≤ 0.05.
ROS-signalling and EPL cell formation
Several lines of evidence suggest that c-SRC can be activated by increased intracellular ROS [16, 51]. Likewise, others have implicated ROS in the activation of p38 MAPK (for example [52–54]). Metabolism of l-proline by proline dehydrogenase within the cell can result in increased intracellular ROS levels [55], providing a potential link between l-proline uptake and signaling pathway activation. mES cells with and without l-proline were assayed for steady state levels of intracellular ROS qualitatively and quantitatively (Fig 5). Qualitatively, a population of ES cells comprised a mix of ROS-bright and ROS-dull cells (Fig 5A); this was reflected in a basal level of biochemically detected ROS within the population (Fig 5D). The addition of l-proline resulted in an increase in the number of ROS-bright cells in the population and a concomitant increase in the levels of ROS detected (Fig 5B and 5D). The PRODH inhibitor 3,4-dehydro-L-proline (DHP) reduced the levels of ROS in l-proline-treated cells to basal levels, but had no impact on ROS levels in untreated mES cells. Addition of an antioxidant, ascorbic acid (Fig 5C) or glutathione (GSH; Fig 5D), abolished ROS-bright cells in the population and reduced the levels of ROS to below basal (mES cell) levels. Finally, inhibiting l-proline metabolism with DHP reduced the expression of Dnmt3b and Otx2 in l-proline-treated cells (Fig 5E), suggesting a requirement for metabolism in the regulation of EPL cell gene expression.
Fig 5. The addition of l-proline to ES cells increases ROS.

A-C. MitoSox Red staining of mES incubated with 200 μM l-proline (B) or l-proline and ascorbic acid (C) and compared to untreated cells (A). Hoechst staining of the same fields of view are shown in Ai, Bi and Ci. D. ROS levels were measured as fluorescence using ROS-Glo™ H2O2 Assay (Promega) in cells that had been incubated with l-proline, 150 μM DHP or 1 mM GSH, as denoted. Results are shown as arbitrary fluorescent units. n = 4, *p≤0.05 when compared with mES cells. # p≤0.05 when compared with mES cells incubated with l-proline. E. The expression of Dnmt3b and Otx2 was analysed in mES cells cultured in medium supplemented 200 μM l-proline or 200 μM l-proline and 150 μM DHP. Expression was normalized to β-actin and expressed relative to mES cells. Error bars represent SEM; n = 4. Comparisons were made to mES cells (*p ≤ 0.05) or mES cells cultured with 200 μM l-proline (# p ≤ 0.05).
Discussion
EPL cell inductive factors act on a primed ES cell population
mES cells can be cultured in the naïve or the primed state. The naive pluripotent cell state is achieved by preventing signaling through ERK1/2 in combination with a GSK3ß inhibitor, and thereby maintaining cells in an intrinsically self-maintaining state regulated by Nanog and shielded from differentiation signals [9, 11]. With increasing ERK signaling mES cells become primed, or able to respond to differentiation signals [31], while deletion of Erk2-/- biases cells towards self-renewal [56]. Inhibition of ERK2 signaling prevented the formation of EPL cells in response to l-proline or MEDII, suggesting that active ERK2, and the concomitant primed mES cell state, was required for EPL cell formation. Inhibition of Src activity can replace the inhibition of ERK signaling in the maintenance of naïve mES cells [57, 58], and c-Src inhibition has been shown to promote the formation of naïve human pluripotent cells [59]. The ability of PP2 to inhibit the formation of EPL cells from ES cells in response to l-proline or MEDII is consistent with a requirement for a primed mES cell substrate in primitive ectoderm formation.
Acquisition and maintenance of the EPL cell state requires the activity of multiple intracellular pathways
Src family kinase signaling was required for the maintenance of the EPL cell state in culture, and suggests a role for this pathway beyond facilitating mES cell priming. Several reports have noted the functional similarity between human (h)ES cells and primitive ectoderm-like cell lines from mouse, EPL cells and EpiSCs [26, 60, 61]. Pluripotency in hES cells is maintained, in part, by fibroblast growth factor-2 (FGF-2). Analysis of the pathways activated by FGF signalling in hES cells demonstrated tyrosine phosphorylation of Src family kinases and Src substrates [62]. This was interpreted as indicating a role for Src kinase activity in the maintenance of hES cell pluripotency by FGF-2, and is consistent with the role of Src family kinase activity in maintaining the primitive ectoderm-like identity of EPL cells in culture.
The formation of EPL cells from mES cells, like the formation of primitive ectoderm from pluripotent cells of the blastocyst, is accompanied by epithelialisation of the pluripotent cells. PP2 prevented this change in colony morphology and promoted growth in compact colonies; others have seen this effect [63]. This effect has also been seen in non-pluripotent cells, in which PP2 inhibited cell migration, and also, surprisingly, in fibroblasts deficient in c-Src, Yes and Fyn, suggesting that migration in these cells depended on a non-Src family kinase mechanism that is affected by PP2 [63]. Consistent with this, the Src family inhibitor SU6656 did not inhibit the epithelialisation of mES cells cultured in MEDII (Data not shown). It is yet to be determined if the mechanisms that regulate colony morphology changes seen in pluripotent cell lineage progression are equivalent to those that regulate migration in somatic cells.
Treatment of mES cells with l-proline resulted in increased p38 MAPK phosphorylation. Inhibition of p38 MAPK activity during EPL cell formation impacted the change in colony morphology, prevented the up regulation of primitive ectoderm markers Dnmt3b and Fgf5 and blocked the change in differentiation kinetics that accompanies EPL cell formation, suggesting a role for p38 MAPK in the establishment of EPL cells. Inhibiting signaling through p38 MAPK did not, however, restore the expression of pluripotent / ES cell markers. The deletion of p38α resulted reduced basal expression of primitive ectoderm markers when compared to WT cells; expression of these markers increased with the addition of MEDII but there was no accompanying alteration in differentiation kinetics (data not shown). Previous reports showed that knockdown of p38δ, the second highest p38 MAPK isoform found in ES cells, in p38α-/- ES cells did not affect ES cell differentiation [64]. Although the loss of p38α did prevent aspects of EPL cell formation, p38α-/- mES cells do not phenocopy mES cells cultured in a p38 MAPK inhibitor, suggesting that in the absence of other p38α factors, including p38β, may regulate EPL cell marker gene expression.
Multiple pathways regulate pluripotent lineage progression
MES cells in culture do not grow as a homogenous population but exist in a metastable state. Heterogeneity has been revealed by the non-uniform expression of Zfp42, Dppa3, Nanog, Pecam1 and Otx2 in cells expressing Oct4 [65–70]. These genes mark interchangeable pluripotent cell states corresponding to an ICM-like, Otx2-low state and a later pluripotent cell state, marked by higher Otx2 expression, that coexist and ensure the self-renewal and perpetuation of pluripotency and a susceptibility to differentiation factors [10]. Otx2 has been shown to be intrinsic to the metastable state; overexpression of Otx2 resulted in a population of cells representative of late epiblast of the post-implantation embryo and EpiSC, while Otx2-/- mES cells are highly enriched for Nanog-expressing cells and predisposed to self-renewal [69]. Otx2 has also been shown to be required for the transition of mES cells from the ICM-like state and later, primed, pluripotent cell states [71]. Otx2 has been used here as a marker of EPL cells. Inhibition of neither Src family kinases, p38 MAPK, nor MEK1/ERK2 affected the regulation of Otx2 expression during the formation of EPL cells, although the loss of ERK1/2 signalling in formed EPL cells did reduce the expression of Otx2 (S4 Fig). The inability to prevent Otx2 expression when mES cells are exposed to l-proline or MEDII may underlie the failure of these inhibitors to prevent all aspects of differentiation and to maintain an mES cell in culture. Several transcriptional modules have been identified in the regulation of mES cells pluripotency, each potentially regulating different gene cohorts and regulated by different upstream triggers [72–76]. It is likely that Src family kinases, p38 MAPK and ERK2 signaling regulate some, but not all, of these transcriptional modules, resulting in the incomplete inhibition of EPL cell formation and maintenance of Otx2 expression.
Multiple pluripotent cell states in culture
The regulation of pluripotent lineage progression is not well understood, but these findings, coupled with the findings of others, can be integrated into a model of the process (Fig 6). In culture, mES cells in the naïve state are hypothesized to represent the earlier pluripotent cells of the lineage [12, 77]. In culture, these cells are maintained by supressing GSK3β signaling in combination with the inhibition of ERK1/2 signaling (through the inhibition of MEK1), Src family kinases or calcineurin [9, 11, 57, 58]. It is thought that calcineurin signaling collaborates with ERK signaling to activate Src, which in turn drives the priming of the cells for differentiation [58]; inhibition of any of these signalling components will prevent Src activation. In primed mES cells ERK activity is present. The absence of calcineurin signaling in these cells [58] will likely prevent the up regulation and activation of c-Src in response to ERK1/2 signaling.
Fig 6. The regulation of progression of the pluripotent lineage in culture.

The cell states represented in vitro, naïve mES cells, primed mES cells and EPL cells have been aligned with the expression of Nanog and Otx2 and with their deduced intracellular signaling activity. Inducers of lineage progression are shown in orange; Calcineurin exerts its effects through dephosphorylation of NFAT and promotes NFAT translocation to the nucleus.
The transition of mES cells to EPL cells involves the activation of Src family kinases and p38 MAPK and occurs in response to l-proline. The addition of l-proline to primed mES cells increases ROS within the cell population. The enzyme central to proline metabolism in the cell, proline dehydrogenase (PRODH or POX), converts l-proline to Δ1-pyrroline-5-carboxylate (P5C), with the concomitant transfer of electrons to oxygen and generation of ROS [78, 79]. ROS increases in mES cells were prevented by inhibition of PRODH, and EPL cell morphology could be prevented by ROS scavengers and PRODH inhibition [21], suggesting a role for proline metabolism in mES cell differentiation. c-Src and p38 MAPK can be activated by increased intracellular ROS [51, 53, 54, 80], providing a pathway for the control of differentiation by l-proline. The action of MEDII and l-proline, through ROS induction, does not appear to be able to overcome the suppression of Src activation that can be achieved when MEK1 is inhibited.
The question arises to the relevance of calcineurin or l-proline in the regulation of Src activity within ES cells. The combination of ERK and calcineurin resulted in differentiation of mES cells to cells of the extraembryonic lineages, primarily trophectoderm and endoderm [58], whereas the combination of ERK and l-proline results in the formation of EPL cells and subsequently cells of the germ lineages [15, 24, 25, 81]. These likely represent different pathways to activate c-Src that achieve alternate cellular outcomes, potentially through the differential regulation of p38 MAPK, and place Src family kinase signaling as pivotal in ES cell differentiation.
Materials and Methods
Cell culture
D3 mouse mES cell line [82] was maintained as described previously in Rathjen and Rathjen, 2003 [83]. EPL cells were formed by culturing mES cells in medium supplemented with 50% MEDII, or medium supplemented with LIF and 200 μM l-proline (Sigma-Aldrich) [20, 83]. Chemical inhibitors (PP2 (Calbiochem), SB203580 (Tocris Bioscience), SU6656 (Sigma Aldrich) and PD0325901 (Selleck Chemicals)) were added every 24 hours or as described in the text. Inhibition of SRC family kinase phosphorylation by PP2 was still apparent at 24 hours (Fig 1G), suggesting this inhibitor was stable in these culture conditions. Inhibition p38 MAPK by SB203580 was clear after 60 minutes (Fig 3A) but was less predictable at 24 hours (data not shown), suggesting activity remained after extended culture but some potency had been lost. Inhibition of ERK2 phosphorylation by PD0325901 was obvious after 60 minutes (Fig 5A) but was not detected at 24 hours (data not shown), suggesting this inhibitor was not stable in culture over this time. MES cells were seeded at a density of 2.5 x 104 cells / cm2 onto tissue culture-treated plasticware (NuncTM), pretreated with 0.2% fetal porcine gelatin (Sigma-Aldrich). Cells were cultured for 3 days (in 50% MEDII) or 4 days (with 200 μM l-proline) with daily medium replenishment. Chemical inhibitors were suspended in dimethyl sulfoxide (DMSO) (Sigma-Aldrich) and added to cells as described in the text. For EPL cell maintenance experiments mES cells were cultured in medium supplemented with 50% MEDII for 2 days, followed by the addition of chemical inhibitors to the medium for a further 2 days, as described in text. Alkaline phosphatase activity was performed according to manufacturer’s instructions using an alkaline phosphatase detection kit (Sigma-Aldrich; catalogue # 86C) with modifications to allow staining in tissue culture plasticware [13]. Images were taken on an Olympus IX51 with an F-view II digital camera (Olympus) or UC-30 digital camera. Embryoid bodies (EBs) were formed from mES cells that had been cultured for 3 days (in 50% MEDII) or 4 days (with 200 μM l-proline) as described previously [83]; chemical inhibitors were added as described in text.
Gene expression analysis
Reverse Transcriptase-PCR (RT-PCR): Total RNA was isolated from cells using TRIzol® reagent (InvitrogenTM) as per the manufacturer’s instructions and DNaseI treated (Ambion). cDNA synthesis was performed using M-MLV Reverse Transcriptase (Promega). PCR reactions, containing GoTaq® Green Master Mix (Promega), 200 nM each of forward and reverse primers and 1 μl of cDNA were initially heated to 95°C for 2 minutes and then cycled through 95°C for 30 seconds, 60°C for 30 seconds and 72°C for 30 seconds on a MJ Research or Bio-rad thermocycler. Primer sequences, the number of cycles used for each primer pair and the size of the expected amplicon are listed in S1 Table.
Quantitative Real-time PCR (qPCR)
RNA was isolated, DNaseI treated and cDNA was synthesized as for RT-PCR. Reactions, consisting of cDNA, HOT FirePol EvaGreen Mix (Integrated Sciences) with 200 nM each of forward and reverse primers, were amplified using MJ research thermocycler with a Chromo4 Continuous Fluorescence Detection System (MJ Research) or ViaTM 7 real time PCR system. The samples were initially heated to 95°C for 15 minutes followed by 40 cycles of 95°C for 15 s, 60°C for 15 s and 72°C for 30 s. The cycle threshold (Ct) values for β-actin were normalized across samples and the raw Ct values were analyzed using Q-Gene software package [84]. Primer sequences are listed in S1 Table.
Western Blot
Total cellular protein was extracted from mES cells with 200 μl Laemmli sample buffer (Bio-rad) containing β-mercaptoethanol (Sigma-Aldrich), protease inhibitor (#P-8350; Sigma-Aldrich) and phosphatase inhibitor, as per manufacturer’s instructions. Protein extract was incubated at 100°C for 10 minutes and separated using a 10% SDS polyacrylamide gel. Standard immunoblot procedures were followed and proteins were visualized using the Immun-StarTM WesternCTM Chemiluminescence Kit (Bio-Rad) and a Bio-Rad ChemiDocTM XRS as per the manufacturer’s instructions. Antibodies, antibody suppliers and dilutions used are stated in S1 Table. Band intensities were estimated using Quantity One software (Bio-Rad).
Kinome array
mES cells were transferred to DMEM + 0.1% FCS (Life technologies) for 4 hours prior to setting up the assay. At time 0, cells were mechanically removed from the plastic and aliquots of approximately 1 x 107 cells were treated with LIF (5 minutes), 200 μM l-proline (5 and 15 minutes) or maintained in DMEM + 0.1% FCS (15 minutes). Cells were lysed and lysates applied to a Proteome ProfilerTM Human Phospo-MAPK Array (R&D Systems, catalogue # ARY002B) as per the manufacturer’s instructions. Signals were developed using enhanced chemiluminescence (GE Lifescience) and quantified using QuantityOne (Bio-Rad).
BrdU Assay
mES cells were cultured with 5’ Bromo 2’ deoxyuridine (BrdU) (10 μM; Sigma Aldrich) for 2 hours before cells were washed and reduced to a single cell suspension. Cells were fixed in ice-cold 70% ethanol, permeabilised in 1 M HCl, and BrdU incorporation detected using an α-BrdU monoclonal antibody (Bioclone Australia) in combination with a rabbit α-mouse IgG Alexa Fluor 488 secondary antibody (Invitrogen). Cells that had incorporated BrdU were quantified by flow cytommetry.
ROS analysis
Live cell imaging
mES cells were seeded at 2.5 x 104 cells per well into 96 well black imaging plates (BD Falcon, 31053) and cultured for 4 days. L-proline (200 μM) was added on day 3, and ascorbic acid (200 μM) was added 4 hours prior to imaging. 30 minutes prior to imaging, MitoSOX™ Red mitochondrial superoxide indicator (5 μM, Invitrogen, M36008) and Hoechst (Life Technologies, H3570) were added as per the manufacturer’s specifications. Cells were washed in live cell imaging buffer (DMEM without Phenol Red (Gibco, 31053), 10% Foetal calf serum (Life Technologies), 0.1mM β-mercaptoethanol (Sigma)) and imaged using a Leica DMIRB inverted fluorescent microscope (Leica Microsystems), connected to a Hamamatsu ORCA-ER digital camera. NIS Elements D software (version 4.0, Nikon®) was used to view the images and control exposure times.
Biochemical analysis of ROS
ROS production was quantified using the ROS-Glo™ H202 Assay (Promega, G8820). MES cells were seeded at 1 x 106 cells per well into a 96 well white imaging plates (BD Falcon, 351130) and allowed to adhere overnight. 200 μM l-proline was added at the time of seeding as denoted in the text. 4 hours prior to performing the assay, DHP (150 μM; Sigma Aldrich) or 1 mM GSH (Sigma-Aldrich, PHR1359) were. The H2O2 substrate solution was added 1 hour after treatments were added and the plate was incubated at 37°C for an additional 3 hours. Cells were washed with PBS, the ROS-Glo detection solution was added to with PBS to a total volume of 200 μl and the cells were incubated at room temperature for 20 minutes. Luminescence readings were taken using the GloMax® -96 Microplate luminometer (Promega).
Statistical Analysis
Experiments were analyzed using one-way ANOVA followed by Tukey’s multiple comparison post-hoc test and significance was achieved at p = 0.05 or less with the use of Graphpad Prism.
Supporting Information
A. ES cells were cultured in ESCM and ESCM + 200 μM L-proline with or without 10 μM PP2, as indicated, for 4 days. Scale bar = 200 μm. B. ES cells were cultured in MEDII + DMSO and MEDII + 5, 10 or 20 μM PP2 for 3 days. Scale bar = 200 μm. Although a clear change in the appearance of colonies could be seen with 5 μM PP2 when compared to MEDII + DMSO, full suppression of EPL cell morphology was seen at 10 and 20 μM. C. ES cells were cultured with 5 and 20 μM PP2 for 5 (■) or 6 (□) days. Cells in S-phase were identified by immunofluorescence for incorporated BrdU followed by flow cytometry. The number of cells incorporating BrdU is shown relative to ES cells. Error bars represent SEM; n = 3. Comparisons were made to untreated ES cells, ** p ≤ 0.01. D. ES cells were cultured in ESCM, MEDII, MEDII + DMSO and MEDII + 10 μM PP2, as indicated, for 3 days and formed into EBs. EBs were collected on days 2, 3 and 4. RNA was isolated and analyzed for expression of Fgf8, Wnt3, Tgfβ1, Bmp4 and Gapdh by RT-PCR; n = 3, a representative image is shown.
(TIF)
A, B. ES cells were cultured in medium supplemented with 200 μM l-proline and DMSO (■) or l-proline and 10 μM PP2 (□) (A) or 10 μM SB203580 (□)(B), for 4 days. RNA from these cells was analyzed for transcripts of Oct4, Sox2, Nanog, Rex1, Spp1, Gbx2, Dnmt3b, Otx2 and Fgf5 by real-time PCR. Expression was normalized to β-actin and expressed relative to ES cells. Error bars represent SEM; n = 3. ES cells in Proline + PP2 were compared to cells cultured in Proline + DMSO (** p ≤ 0.05) or ES cells (# p ≤ 0.05). C, D. ES cells were cultured with l-proline + DMSO, l-proline +10 μM PP2 (C) and L-proline +10 μM SB203580 (D) for 4 days before being formed into embryoid bodies (EBs). EBs were analysed as for the expression of T by RT-PCR. n = 3.
(TIF)
A. ES cells were treated with LIF or 100 μM of l-proline for 5 or 20 minutes, as indicated. Cells were lysed and protein bound to the kinome array and binding quantified. Binding to antibodies specific for pp38α, pp38β, pHsbp2 and MSK2 is shown; n = 2, values have been averaged. B. WT ES cell and p38α KO ES cells were cultured in ESCM. RNA was analyzed by qPCR for the expression of Nanog, Rex1, Spp1, Gbx2, Fgf5, Dnmt3b and Otx2. Expression was normalized to Oct4 and expressed relative to WT ES cells. Error bars represent SEM; n = 3. **p ≤ 0.01 when compared to WT ES cells. Loss of p38α decreased expression of primitive ectoderm markers in the ES cell population. C. p38α KO ES cells were cultured in ESCM and MEDII for 3 days to form EPL cells. RNA was analyzed by qPCR for the expression of Nanog, Rex1, Spp1, Gbx2, Fgf5, Dnmt3b and Otx2 by real-time PCR. Expression was normalized to Oct4 and expressed relative to p38α KO ES cells. Error bars represent SEM; n = 3. **p ≤ 0.01, *p ≤ 0.05 when compared to KO ES cells.
(TIF)
ES cells were cultured in MEDII + DMSO and MEDII + 1, 2 or 4 μM SU6656 for 3 days. Scale bar = 200 μm.
(TIF)
(DOCX)
Acknowledgments
The authors would like to thank members of the Rathjen laboratory for insightful discussions of the project. This work was supported by the University of Melbourne, the Australian Stem Cell Centre, Stem Cells Australia and the David Hay Postgraduate Writing up Award. BSNT was supported by an Endeavour International Postgraduate Research Scholarship and a Melbourne International Research Scholarship. CY was supported by an Australian Postgraduate Awards with additional support from the Australian Stem Cell Centre.
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This work was funded by the University of Melbourne, the Australian Stem Cell Centre (funding agreements P019 and P084), Stem Cells Australia, the David Hay Postgraduate Writing Up award and the University of Tasmania. BSNT was supported by an Endeavour International Postgraduate Research Scholarship and a Melbourne International Research Scholarship. CY was supported by an Australian Postgraduate Awards with additional support from the Australian Stem Cell Centre. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Chazaud C, Yamanaka Y, Pawson T, Rossant J. Early lineage segregation between epiblast and primitive endoderm in mouse blastocysts through the Grb2-MAPK pathway. Dev Cell. 2006;10(5):615–24. 10.1016/j.devcel.2006.02.020 . [DOI] [PubMed] [Google Scholar]
- 2.Pera MF, Tam PP. Extrinsic regulation of pluripotent stem cells. Nature. 2010;465(7299):713–20. 10.1038/nature09228 . [DOI] [PubMed] [Google Scholar]
- 3.Rathjen J. The States of Pluripotency: Pluripotent Lineage Development in the Embryo and in the Dish. ISRN Stem Cells. 2014;2014 Article ID 208067. 10.1155/2014/208067 [DOI] [Google Scholar]
- 4.Evans MJ, Kaufman MH. Establishment in culture of pluripotential cells from mouse embryos. Nature. 1981;292(5819):154–6. 10.1038/292154a0 . [DOI] [PubMed] [Google Scholar]
- 5.Martin GR. Isolation of a pluripotent cell line from early mouse embryos cultured in medium conditioned by teratocarcinoma stem cells. Proc Natl Acad Sci U S A. 1981;78(12):7634–8. 10.1073/pnas.78.12.7634 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Brons IG, Smithers LE, Trotter MW, Rugg-Gunn P, Sun B, Chuva de Sousa Lopes SM, et al. Derivation of pluripotent epiblast stem cells from mammalian embryos. Nature. 2007;448(7150):191–5. 10.1038/nature05950 . [DOI] [PubMed] [Google Scholar]
- 7.Tesar PJ, Chenoweth JG, Brook FA, Davies TJ, Evans EP, Mack DL, et al. New cell lines from mouse epiblast share defining features with human embryonic stem cells. Nature. 2007;448(7150):196–9. 10.1038/nature05972 . [DOI] [PubMed] [Google Scholar]
- 8.Kojima Y, Kaufman-Francis K, Studdert JB, Steiner KA, Power MD, Loebel DA, et al. The transcriptional and functional properties of mouse epiblast stem cells resemble the anterior primitive streak. Cell Stem Cell. 2014;14(1):107–20. 10.1016/j.stem.2013.09.014 . [DOI] [PubMed] [Google Scholar]
- 9.Ying QL, Wray J, Nichols J, Batlle-Morera L, Doble B, Woodgett J, et al. The ground state of embryonic stem cell self-renewal. Nature. 2008;453(7194):519–23. 10.1038/nature06968 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Silva J, Barrandon O, Nichols J, Kawaguchi J, Theunissen TW, Smith A. Promotion of reprogramming to ground state pluripotency by signal inhibition. PLoS Biol. 2008;6(10):e253 10.1371/journal.pbio.0060253 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Silva J, Nichols J, Theunissen TW, Guo G, van Oosten AL, Barrandon O, et al. Nanog is the gateway to the pluripotent ground state. Cell. 2009;138(4):722–37. 10.1016/j.cell.2009.07.039 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Boroviak T, Loos R, Bertone P, Smith A, Nichols J. The ability of inner-cell-mass cells to self-renew as embryonic stem cells is acquired following epiblast specification. Nat Cell Biol. 2014;16(6):516–28. 10.1038/ncb2965 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Rathjen J, Lake JA, Bettess MD, Washington JM, Chapman G, Rathjen PD. Formation of a primitive ectoderm like cell population, EPL cells, from ES cells in response to biologically derived factors. J Cell Sci. 1999;112 (Pt 5):601–12. . [DOI] [PubMed] [Google Scholar]
- 14.Pelton TA, Sharma S, Schulz TC, Rathjen J, Rathjen PD. Transient pluripotent cell populations during primitive ectoderm formation: correlation of in vivo and in vitro pluripotent cell development. J Cell Sci. 2002;115(Pt 2):329–39. . [DOI] [PubMed] [Google Scholar]
- 15.Harvey NT, Hughes JN, Lonic A, Yap C, Long C, Rathjen PD, et al. Response to BMP4 signalling during ES cell differentiation defines intermediates of the ectoderm lineage. J Cell Sci. 2010;123(Pt 10):1796–804. 10.1242/jcs.047530 . [DOI] [PubMed] [Google Scholar]
- 16.Zhang Y, Li TS, Lee ST, Wawrowsky KA, Cheng K, Galang G, et al. Dedifferentiation and proliferation of mammalian cardiomyocytes. PloS one. 2010;5(9):e12559 10.1371/journal.pone.0012559 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Guo G, Yang J, Nichols J, Hall JS, Eyres I, Mansfield W, et al. Klf4 reverts developmentally programmed restriction of ground state pluripotency. Development. 2009;136(7):1063–9. 10.1242/dev.030957 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Hanna J, Markoulaki S, Mitalipova M, Cheng AW, Cassady JP, Staerk J, et al. Metastable pluripotent states in NOD-mouse-derived ESCs. Cell Stem Cell. 2009;4(6):513–24. 10.1016/j.stem.2009.04.015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Tan BS, Lonic A, Morris MB, Rathjen PD, Rathjen J. The amino acid transporter SNAT2 mediates L-proline-induced differentiation of ES cells. Am J Physiol Cell Physiol. 2011;300(6):C1270–9. 10.1152/ajpcell.00235.2010 . [DOI] [PubMed] [Google Scholar]
- 20.Washington JM, Rathjen J, Felquer F, Lonic A, Bettess MD, Hamra N, et al. L-Proline induces differentiation of ES cells: a novel role for an amino acid in the regulation of pluripotent cells in culture. Am J Physiol Cell Physiol. 2010;298(5):C982–92. 10.1152/ajpcell.00498.2009 . [DOI] [PubMed] [Google Scholar]
- 21.Casalino L, Comes S, Lambazzi G, De Stefano B, Filosa S, De Falco S, et al. Control of embryonic stem cell metastability by L-proline catabolism. J Mol Cell Biol. 2011;3(2):108–22. 10.1093/jmcb/mjr001 . [DOI] [PubMed] [Google Scholar]
- 22.Comes S, Gagliardi M, Laprano N, Fico A, Cimmino A, Palamidessi A, et al. L-Proline Induces a Mesenchymal-like Invasive Program in Embryonic Stem Cells by Remodeling H3K9 and H3K36 Methylation. Stem Cell Reports. 2013;1(4):307–21. 10.1016/j.stemcr.2013.09.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Rathjen J, Rathjen PD. Formation of neural precursor cell populations by differentiation of embryonic stem cells in vitro. ScientificWorldJournal. 2002;2:690–700. 10.1100/tsw.2002.134 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Lake J, Rathjen J, Remiszewski J, Rathjen PD. Reversible programming of pluripotent cell differentiation. J Cell Sci. 2000;113 (Pt 3):555–66. . [DOI] [PubMed] [Google Scholar]
- 25.Vassilieva S, Goh HN, Lau KX, Hughes JN, Familari M, Rathjen PD, et al. A system to enrich for primitive streak-derivatives, definitive endoderm and mesoderm, from pluripotent cells in culture. PloS one. 2012;7(6):e38645 10.1371/journal.pone.0038645 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Hughes JN, Dodge N, Rathjen PD, Rathjen J. A novel role for gamma-secretase in the formation of primitive streak-like intermediates from ES cells in culture. Stem Cells. 2009;27(12):2941–51. 10.1002/stem.218 . [DOI] [PubMed] [Google Scholar]
- 27.Rodda SJ, Kavanagh SJ, Rathjen J, Rathjen PD. Embryonic stem cell differentiation and the analysis of mammalian development. Int J Dev Biol. 2002;46(4):449–58. . [PubMed] [Google Scholar]
- 28.Yap C, Goh HN, Familari M, Rathjen PD, Rathjen J. The formation of proximal and distal definitive endoderm populations in culture requires p38 MAPK activity. J Cell Sci. 2014. 10.1242/jcs.134502 . [DOI] [PubMed] [Google Scholar]
- 29.Hughes JN, Washington JM, Zheng Z, Lau XK, Yap C, Rathjen PD, et al. Manipulation of cell:cell contacts and mesoderm suppressing activity direct lineage choice from pluripotent primitive ectoderm-like cells in culture. PloS one. 2009;4(5):e5579 10.1371/journal.pone.0005579 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Ernst M, Gearing DP, Dunn AR. Functional and biochemical association of Hck with the LIF/IL-6 receptor signal transducing subunit gp130 in embryonic stem cells. EMBO J. 1994;13(7):1574–84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Kunath T, Saba-El-Leil MK, Almousailleakh M, Wray J, Meloche S, Smith A. FGF stimulation of the Erk1/2 signalling cascade triggers transition of pluripotent embryonic stem cells from self-renewal to lineage commitment. Development. 2007;134(16):2895–902. 10.1242/dev.02880 . [DOI] [PubMed] [Google Scholar]
- 32.Stavridis MP, Lunn JS, Collins BJ, Storey KG. A discrete period of FGF-induced Erk1/2 signalling is required for vertebrate neural specification. Development. 2007;134(16):2889–94. 10.1242/dev.02858 . [DOI] [PubMed] [Google Scholar]
- 33.Burdon T, Stracey C, Chambers I, Nichols J, Smith A. Suppression of SHP-2 and ERK signalling promotes self-renewal of mouse embryonic stem cells. Dev Biol. 1999;210(1):30–43. 10.1006/dbio.1999.9265 . [DOI] [PubMed] [Google Scholar]
- 34.Bain J, Plater L, Elliott M, Shpiro N, Hastie CJ, McLauchlan H, et al. The selectivity of protein kinase inhibitors: a further update. Biochem J. 2007;408(3):297–315. 10.1042/BJ20070797 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Kong L, Deng Z, Shen H, Zhang Y. Src family kinase inhibitor PP2 efficiently inhibits cervical cancer cell proliferation through down-regulating phospho-Src-Y416 and phospho-EGFR-Y1173. Mol Cell Biochem. 2011;348(1–2):11–9. 10.1007/s11010-010-0632-1 . [DOI] [PubMed] [Google Scholar]
- 36.Hanke JH, Gardner JP, Dow RL, Changelian PS, Brissette WH, Weringer EJ, et al. Discovery of a novel, potent, and Src family-selective tyrosine kinase inhibitor. Study of Lck- and FynT-dependent T cell activation. J Biol Chem. 1996;271(2):695–701. 10.1074/jbc.271.2.695 . [DOI] [PubMed] [Google Scholar]
- 37.Wu JX, Adamson ED. Kinase-negative mutant epidermal growth factor receptor (EGFR) expression during embryonal stem cell differentiation favours EGFR-independent lineages. Development. 1996;122(10):3331–42. . [DOI] [PubMed] [Google Scholar]
- 38.Meyn MA 3rd, Schreiner SJ, Dumitrescu TP, Nau GJ, Smithgall TE. SRC family kinase activity is required for murine embryonic stem cell growth and differentiation. Mol Pharmacol. 2005;68(5):1320–30. 10.1124/mol.104.010231 . [DOI] [PubMed] [Google Scholar]
- 39.Meyn MA 3rd, Smithgall TE. Chemical genetics identifies c-Src as an activator of primitive ectoderm formation in murine embryonic stem cells. Sci Signal. 2009;2(92):ra64 10.1126/scisignal.2000311 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Aouadi M, Bost F, Caron L, Laurent K, Le Marchand Brustel Y, Binetruy B. p38 mitogen-activated protein kinase activity commits embryonic stem cells to either neurogenesis or cardiomyogenesis. Stem Cells. 2006;24(5):1399–406. 10.1634/stemcells.2005-0398 . [DOI] [PubMed] [Google Scholar]
- 41.Chen S, Do JT, Zhang Q, Yao S, Yan F, Peters EC, et al. Self-renewal of embryonic stem cells by a small molecule. Proc Natl Acad Sci U S A. 2006;103(46):17266–71. 10.1073/pnas.0608156103 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Wu J, Kubota J, Hirayama J, Nagai Y, Nishina S, Yokoi T, et al. p38 Mitogen-activated protein kinase controls a switch between cardiomyocyte and neuronal commitment of murine embryonic stem cells by activating myocyte enhancer factor 2C-dependent bone morphogenetic protein 2 transcription. Stem Cells Dev. 2010;19(11):1723–34. 10.1089/scd.2010.0066 . [DOI] [PubMed] [Google Scholar]
- 43.Barruet E, Hadadeh O, Peiretti F, Renault VM, Hadjal Y, Bernot D, et al. p38 mitogen activated protein kinase controls two successive-steps during the early mesodermal commitment of embryonic stem cells. Stem Cells Dev. 2011;20(7):1233–46. 10.1089/scd.2010.0213 . [DOI] [PubMed] [Google Scholar]
- 44.Ying QL, Stavridis M, Griffiths D, Li M, Smith A. Conversion of embryonic stem cells into neuroectodermal precursors in adherent monoculture. Nat Biotechnol. 2003;21(2):183–6. 10.1038/nbt780 . [DOI] [PubMed] [Google Scholar]
- 45.Blake RA, Broome MA, Liu X, Wu J, Gishizky M, Sun L, et al. SU6656, a selective src family kinase inhibitor, used to probe growth factor signaling. Mol Cell Biol. 2000;20(23):9018–27. 10.1128/mcb.20.23.9018-9027.2000 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Williams MR, Arthur JS, Balendran A, van der Kaay J, Poli V, Cohen P, et al. The role of 3-phosphoinositide-dependent protein kinase 1 in activating AGC kinases defined in embryonic stem cells. Curr Biol. 2000;10(8):439–48. 10.1016/s0960-9822(00)00441-3 . [DOI] [PubMed] [Google Scholar]
- 47.Anneren C, Cowan CA, Melton DA. The Src family of tyrosine kinases is important for embryonic stem cell self-renewal. J Biol Chem. 2004;279(30):31590–8. 10.1074/jbc.M403547200 . [DOI] [PubMed] [Google Scholar]
- 48.Rouse J, Cohen P, Trigon S, Morange M, Alonso-Llamazares A, Zamanillo D, et al. A novel kinase cascade triggered by stress and heat shock that stimulates MAPKAP kinase-2 and phosphorylation of the small heat shock proteins. Cell. 1994;78(6):1027–37. 10.1016/0092-8674(94)90277-1 . [DOI] [PubMed] [Google Scholar]
- 49.Darragh J, Soloaga A, Beardmore VA, Wingate AD, Wiggin GR, Peggie M, et al. MSKs are required for the transcription of the nuclear orphan receptors Nur77, Nurr1 and Nor1 downstream of MAPK signalling. Biochem J. 2005;390(Pt 3):749–59. 10.1042/BJ20050196 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Allen M, Svensson L, Roach M, Hambor J, McNeish J, Gabel CA. Deficiency of the stress kinase p38alpha results in embryonic lethality: characterization of the kinase dependence of stress responses of enzyme-deficient embryonic stem cells. J Exp Med. 2000;191(5):859–70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Giannoni E, Buricchi F, Raugei G, Ramponi G, Chiarugi P. Intracellular reactive oxygen species activate Src tyrosine kinase during cell adhesion and anchorage-dependent cell growth. Mol Cell Biol. 2005;25(15):6391–403. 10.1128/MCB.25.15.6391-6403.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Cuadrado A, Nebreda AR. Mechanisms and functions of p38 MAPK signalling. Biochem J. 2010;429(3):403–17. 10.1042/BJ20100323 . [DOI] [PubMed] [Google Scholar]
- 53.Ray PD, Huang BW, Tsuji Y. Reactive oxygen species (ROS) homeostasis and redox regulation in cellular signaling. Cell Signal. 2012;24(5):981–90. 10.1016/j.cellsig.2012.01.008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Choi TG, Lee J, Ha J, Kim SS. Apoptosis signal-regulating kinase 1 is an intracellular inducer of p38 MAPK-mediated myogenic signalling in cardiac myoblasts. Biochim Biophys Acta. 2011;1813(8):1412–21. 10.1016/j.bbamcr.2011.04.001 . [DOI] [PubMed] [Google Scholar]
- 55.Liu W, Phang JM. Proline dehydrogenase (oxidase) in cancer. Biofactors. 2012;38(6):398–406. 10.1002/biof.1036 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Hamilton WB, Kaji K, Kunath T. ERK2 suppresses self-renewal capacity of embryonic stem cells, but is not required for multi-lineage commitment. PloS one. 2013;8(4):e60907 10.1371/journal.pone.0060907 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Shimizu T, Ueda J, Ho JC, Iwasaki K, Poellinger L, Harada I, et al. Dual inhibition of Src and GSK3 maintains mouse embryonic stem cells, whose differentiation is mechanically regulated by Src signaling. Stem Cells. 2012;30(7):1394–404. 10.1002/stem.1119 . [DOI] [PubMed] [Google Scholar]
- 58.Li X, Zhu L, Yang A, Lin J, Tang F, Jin S, et al. Calcineurin-NFAT signaling critically regulates early lineage specification in mouse embryonic stem cells and embryos. Cell Stem Cell. 2011;8(1):46–58. 10.1016/j.stem.2010.11.027 . [DOI] [PubMed] [Google Scholar]
- 59.Theunissen TW, Powell BE, Wang H, Mitalipova M, Faddah DA, Reddy J, et al. Systematic Identification of Culture Conditions for Induction and Maintenance of Naive Human Pluripotency. Cell Stem Cell. 2014. 10.1016/j.stem.2014.07.002 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Vallier L, Mendjan S, Brown S, Chng Z, Teo A, Smithers LE, et al. Activin/Nodal signalling maintains pluripotency by controlling Nanog expression. Development. 2009;136(8):1339–49. 10.1242/dev.033951 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Ware CB, Wang L, Mecham BH, Shen L, Nelson AM, Bar M, et al. Histone deacetylase inhibition elicits an evolutionarily conserved self-renewal program in embryonic stem cells. Cell Stem Cell. 2009;4(4):359–69. 10.1016/j.stem.2009.03.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Ding VM, Boersema PJ, Foong LY, Preisinger C, Koh G, Natarajan S, et al. Tyrosine phosphorylation profiling in FGF-2 stimulated human embryonic stem cells. PloS one. 2011;6(3):e17538 10.1371/journal.pone.0017538 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Tamm C, Galito SP, Anneren C. Differential effects on cell motility, embryonic stem cell self-renewal and senescence by diverse Src kinase family inhibitors. Exp Cell Res. 2012;318(4):336–49. 10.1016/j.yexcr.2011.12.008 . [DOI] [PubMed] [Google Scholar]
- 64.Chakraborty S, Kang B, Huang F, Guo YL. Mouse embryonic stem cells lacking p38alpha and p38delta can differentiate to endothelial cells, smooth muscle cells, and epithelial cells. Differentiation. 2009;78(2–3):143–50. 10.1016/j.diff.2009.05.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Toyooka Y, Shimosato D, Murakami K, Takahashi K, Niwa H. Identification and characterization of subpopulations in undifferentiated ES cell culture. Development. 2008;135(5):909–18. 10.1242/dev.017400 . [DOI] [PubMed] [Google Scholar]
- 66.Mitsui K, Tokuzawa Y, Itoh H, Segawa K, Murakami M, Takahashi K, et al. The homeoprotein Nanog is required for maintenance of pluripotency in mouse epiblast and ES cells. Cell. 2003;113(5):631–42. 10.1016/s0092-8674(03)00393-3 . [DOI] [PubMed] [Google Scholar]
- 67.Hayashi K, Lopes SM, Tang F, Surani MA. Dynamic equilibrium and heterogeneity of mouse pluripotent stem cells with distinct functional and epigenetic states. Cell Stem Cell. 2008;3(4):391–401. 10.1016/j.stem.2008.07.027 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Chambers I, Silva J, Colby D, Nichols J, Nijmeijer B, Robertson M, et al. Nanog safeguards pluripotency and mediates germline development. Nature. 2007;450(7173):1230–4. 10.1038/nature06403 . [DOI] [PubMed] [Google Scholar]
- 69.Acampora D, Di Giovannantonio LG, Simeone A. Otx2 is an intrinsic determinant of the embryonic stem cell state and is required for transition to a stable epiblast stem cell condition. Development. 2013;140(1):43–55. 10.1242/dev.085290 . [DOI] [PubMed] [Google Scholar]
- 70.Furusawa T, Ohkoshi K, Honda C, Takahashi S, Tokunaga T. Embryonic stem cells expressing both platelet endothelial cell adhesion molecule-1 and stage-specific embryonic antigen-1 differentiate predominantly into epiblast cells in a chimeric embryo. Biol Reprod. 2004;70(5):1452–7. 10.1095/biolreprod.103.024190 . [DOI] [PubMed] [Google Scholar]
- 71.Yang SH, Kalkan T, Morissroe C, Marks H, Stunnenberg H, Smith A, et al. Otx2 and Oct4 Drive Early Enhancer Activation during Embryonic Stem Cell Transition from Naive Pluripotency. Cell Rep. 2014;7(6):1968–81. 10.1016/j.celrep.2014.05.037 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Hu G, Kim J, Xu Q, Leng Y, Orkin SH, Elledge SJ. A genome-wide RNAi screen identifies a new transcriptional module required for self-renewal. Genes Dev. 2009;23(7):837–48. 10.1101/gad.1769609 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Westerman BA, Braat AK, Taub N, Potman M, Vissers JH, Blom M, et al. A genome-wide RNAi screen in mouse embryonic stem cells identifies Mp1 as a key mediator of differentiation. J Exp Med. 2011;208(13):2675–89. 10.1084/jem.20102037 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Wong DJ, Liu H, Ridky TW, Cassarino D, Segal E, Chang HY. Module map of stem cell genes guides creation of epithelial cancer stem cells. Cell Stem Cell. 2008;2(4):333–44. 10.1016/j.stem.2008.02.009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Kim J, Chu J, Shen X, Wang J, Orkin SH. An extended transcriptional network for pluripotency of embryonic stem cells. Cell. 2008;132(6):1049–61. 10.1016/j.cell.2008.02.039 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Kim J, Woo AJ, Chu J, Snow JW, Fujiwara Y, Kim CG, et al. A Myc network accounts for similarities between embryonic stem and cancer cell transcription programs. Cell. 2010;143(2):313–24. 10.1016/j.cell.2010.09.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Nichols J, Silva J, Roode M, Smith A. Suppression of Erk signalling promotes ground state pluripotency in the mouse embryo. Development. 2009;136(19):3215–22. 10.1242/dev.038893 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Donald SP, Sun XY, Hu CA, Yu J, Mei JM, Valle D, et al. Proline oxidase, encoded by p53-induced gene-6, catalyzes the generation of proline-dependent reactive oxygen species. Cancer Res. 2001;61(5):1810–5. . [PubMed] [Google Scholar]
- 79.Liu Y, Borchert GL, Surazynski A, Hu CA, Phang JM. Proline oxidase activates both intrinsic and extrinsic pathways for apoptosis: the role of ROS/superoxides, NFAT and MEK/ERK signaling. Oncogene. 2006;25(41):5640–7. 10.1038/sj.onc.1209564 . [DOI] [PubMed] [Google Scholar]
- 80.Lee SH, Lee YJ, Park SW, Kim HS, Han HJ. Caveolin-1 and integrin beta1 regulate embryonic stem cell proliferation via p38 MAPK and FAK in high glucose. J Cell Physiol. 2011;226(7):1850–9. 10.1002/jcp.22510 . [DOI] [PubMed] [Google Scholar]
- 81.Rathjen J, Haines BP, Hudson KM, Nesci A, Dunn S, Rathjen PD. Directed differentiation of pluripotent cells to neural lineages: homogeneous formation and differentiation of a neurectoderm population. Development. 2002;129(11):2649–61. . [DOI] [PubMed] [Google Scholar]
- 82.Doetschman TC, Eistetter H, Katz M, Schmidt W, Kemler R. The in vitro development of blastocyst-derived embryonic stem cell lines: formation of visceral yolk sac, blood islands and myocardium. J Embryol Exp Morphol. 1985;87:27–45. . [PubMed] [Google Scholar]
- 83.Rathjen J, Rathjen PD. Lineage specific differentiation of mouse ES cells: formation and differentiation of early primitive ectoderm-like (EPL) cells. Methods Enzymol. 2003;365:3–25. 10.1016/S0076-6879(03)65001-9 . [DOI] [PubMed] [Google Scholar]
- 84.Simon P. Q-Gene: processing quantitative real-time RT-PCR data. Bioinformatics. 2003;19(11):1439–40. 10.1093/bioinformatics/btg157 . [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
A. ES cells were cultured in ESCM and ESCM + 200 μM L-proline with or without 10 μM PP2, as indicated, for 4 days. Scale bar = 200 μm. B. ES cells were cultured in MEDII + DMSO and MEDII + 5, 10 or 20 μM PP2 for 3 days. Scale bar = 200 μm. Although a clear change in the appearance of colonies could be seen with 5 μM PP2 when compared to MEDII + DMSO, full suppression of EPL cell morphology was seen at 10 and 20 μM. C. ES cells were cultured with 5 and 20 μM PP2 for 5 (■) or 6 (□) days. Cells in S-phase were identified by immunofluorescence for incorporated BrdU followed by flow cytometry. The number of cells incorporating BrdU is shown relative to ES cells. Error bars represent SEM; n = 3. Comparisons were made to untreated ES cells, ** p ≤ 0.01. D. ES cells were cultured in ESCM, MEDII, MEDII + DMSO and MEDII + 10 μM PP2, as indicated, for 3 days and formed into EBs. EBs were collected on days 2, 3 and 4. RNA was isolated and analyzed for expression of Fgf8, Wnt3, Tgfβ1, Bmp4 and Gapdh by RT-PCR; n = 3, a representative image is shown.
(TIF)
A, B. ES cells were cultured in medium supplemented with 200 μM l-proline and DMSO (■) or l-proline and 10 μM PP2 (□) (A) or 10 μM SB203580 (□)(B), for 4 days. RNA from these cells was analyzed for transcripts of Oct4, Sox2, Nanog, Rex1, Spp1, Gbx2, Dnmt3b, Otx2 and Fgf5 by real-time PCR. Expression was normalized to β-actin and expressed relative to ES cells. Error bars represent SEM; n = 3. ES cells in Proline + PP2 were compared to cells cultured in Proline + DMSO (** p ≤ 0.05) or ES cells (# p ≤ 0.05). C, D. ES cells were cultured with l-proline + DMSO, l-proline +10 μM PP2 (C) and L-proline +10 μM SB203580 (D) for 4 days before being formed into embryoid bodies (EBs). EBs were analysed as for the expression of T by RT-PCR. n = 3.
(TIF)
A. ES cells were treated with LIF or 100 μM of l-proline for 5 or 20 minutes, as indicated. Cells were lysed and protein bound to the kinome array and binding quantified. Binding to antibodies specific for pp38α, pp38β, pHsbp2 and MSK2 is shown; n = 2, values have been averaged. B. WT ES cell and p38α KO ES cells were cultured in ESCM. RNA was analyzed by qPCR for the expression of Nanog, Rex1, Spp1, Gbx2, Fgf5, Dnmt3b and Otx2. Expression was normalized to Oct4 and expressed relative to WT ES cells. Error bars represent SEM; n = 3. **p ≤ 0.01 when compared to WT ES cells. Loss of p38α decreased expression of primitive ectoderm markers in the ES cell population. C. p38α KO ES cells were cultured in ESCM and MEDII for 3 days to form EPL cells. RNA was analyzed by qPCR for the expression of Nanog, Rex1, Spp1, Gbx2, Fgf5, Dnmt3b and Otx2 by real-time PCR. Expression was normalized to Oct4 and expressed relative to p38α KO ES cells. Error bars represent SEM; n = 3. **p ≤ 0.01, *p ≤ 0.05 when compared to KO ES cells.
(TIF)
ES cells were cultured in MEDII + DMSO and MEDII + 1, 2 or 4 μM SU6656 for 3 days. Scale bar = 200 μm.
(TIF)
(DOCX)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.