

Abstract
The major histocompatibility complex (MHC) is a key component of adaptive immunity in all jawed vertebrates, and understanding the evolutionary mechanisms that have shaped these genes in amphibians, one of the earliest terrestrial tetrapods, is important. We characterised MHC class I variation in three common Japanese Rana species (Rana japonica, Rana ornativentris and Rana tagoi tagoi) and identified a total of 60 variants from 21 individuals. We also found evolutionary signatures of gene duplication, recombination and balancing selection (including trans-species polymorphism), all of which drive increased MHC diversity. A unique feature of MHC class I from these three Ranidae species includes low synonymous differences per site (d S) within species, which we attribute to a more recent diversification of these sequences or recent gene duplication. The resulting higher d N/d S ratio relative to other anurans studied could be related to stronger selection pressure at peptide binding sites. This is one of the first studies to investigate MHC in Japanese amphibians and permits further exploration of the polygenetic factors associated with resistance to infectious diseases.
Electronic supplementary material
The online version of this article (doi:10.1007/s00251-016-0934-x) contains supplementary material, which is available to authorized users.
Keywords: Major histocompatibility complex, Selection, Rana, Anura
Introduction
Major histocompatibility complex (MHC) genes code for membrane-bound glycoproteins that recognise, bind and present specific antigens to T lymphocytes and, thus, are important components of adaptive immunity in jawed vertebrates. MHC class I molecules bind endogenous antigenic peptides, presenting them to cytotoxic T cells, and comprised of an α heavy chain and β2m microglobulin chain. The α chain of MHC class I consists of three extracellular domains (α1, α2 and α3) encoded by exons 2, 3 and 4, respectively. The anchor residues within the peptide binding groove, sometimes referred to as peptide binding sites, are found in the α1 and α2 domains (Hughes and Yeager 1998). There are two groups of class I genes based on functional differences: class Ia (classical) genes which are highly polymorphic and expressed in all nucleated cells and class Ib (non-classical) that are less polymorphic and have variable expression across tissues (Goyos et al. 2011; Hughes and Yeager 1998).
Classical MHC genes are one of the most polymorphic genes of vertebrate genomes (Hedrick 1994), and they evolve under a birth and death process (Nei and Rooney 2005), resulting in variation in MHC loci number between different species. MHC variation is primarily maintained by pathogen-mediated balancing selection (Bernatchez and Landry 2003; Hughes and Yeager 1998). One of the characteristics of balancing selection is trans-species polymorphism, whereby alleles in different species are more closely related than conspecific alleles (Klein et al. 1998), and this has been seen in MHC genes (Wei et al. 2010; Jaratlerdsiri et al. 2014). As the MHC is important for adaptive immunity, several studies in vertebrates have found associations with MHC variation and infectious diseases (summarised by Sommer 2005). In particular, recent studies have identified MHC–disease associations in a number of amphibian species (Bataille et al. 2015; Savage and Zamudio 2011; Teacher et al. 2009).
Anuran amphibians are ectothermic tetrapods with a metamorphic life cycle, comprising a developing pre-metamorphic stage (tadpoles) inhabiting aqueous environments and a post-metamorphic stage occupying terrestrial and/or aquatic habitats. A change in morphology and environment during the life cycle presents the potential for differential exposure to different pathogens during development. There are reports that certain pathogens may have stronger impact on either pre- (Chambouvet et al. 2015; Haislip et al. 2011) or post-metamorphic life stages (Blaustein et al. 2005), supporting that developmental and environmental changes could influence amphibian host immunity. Many amphibian diseases caused by pathogens, including bacteria, viruses, fungi and parasites, have been reported (summarised by Densmore and Green 2007); one of the most commonly documented amphibian infectious diseases in recent times is chytridiomycosis.
Chytridiomycosis is a serious disease in amphibians caused by the chytrid fungus Batrachochytridium dendrobatidis, which was identified by Berger et al (1998) and characterised by Longcore et al (1999). Many studies have attributed chytridiomycosis, which causes epidermal disruption, to the decrease in amphibian populations around the world (Daszak et al. 2003; Longcore et al. 2007; Skerratt et al. 2007). In Japan, several strains of B. dendrobatidis were characterised as either non-native or apparently endemic strains, although low incidence (4.1 %) was found in free-ranging amphibians (Goka et al. 2009). Some amphibian species in Japan where the fungus was detected include exotic species from the pet trade and the naturalised Lithobates (Rana) catesbeianus, a key carrier species attributed with the spread of this global disease (Garner et al. 2006). It has been suggested that B. dendrobatidis is endemic to Asia (Goka et al. 2009; Bataille et al. 2013), implying the coexistence between Japanese (or Asian) amphibians and B. dendrobatidis for a long time, which may have allowed the amphibians to evolve an effective resistance against chytrid infection. This could explain the tolerance of endemic amphibians to the fungi and the resulting low incidence in Japan (or Asia). Therefore, initiating the examination of immune genes of Japanese amphibian species will contribute to further understanding amphibian host–disease dynamics.
Within the family Ranidae, MHC class I has been studied in L. catesbeianus (American bullfrog), Lithobates clamitans (green frog), Lithobates yavapaiensis (lowland leopard frog), Lithobates pipiens (Northern leopard frog) and Rana temporaria (common frog), and signatures of balancing selection have been detected (Flajnik et al. 1999; Kiemnec-Tyburczy et al. 2012; Teacher et al. 2009). Apart from balancing selection, recombination and gene duplication also play a role in driving MHC diversity (Carrington 1999; Yeager and Hughes 1999). Recombination between exons at a single locus has been identified in Xenopus laevis (African clawed frog) and two Rhacophoridae species (Bos and Waldman 2006; Zhao et al. 2013). Whilst some model Pipidae and Ranidae species express a single classical MHC class I locus (Flajnik et al. 1999; Goyos et al. 2011; Ohta et al. 2006; Teacher et al. 2009), copy number variation was revealed following research across more expansive families and additional Ranidae species (Kiemnec-Tyburczy et al. 2012; Lillie et al. 2014; Zhao et al. 2013). The aim of this study was to characterise MHC class I genes in three Japanese frog species from the Rana genus. In addition, we wanted to investigate whether selective mechanisms and gene duplication have shaped MHC variation. This will contribute to further understanding about the evolution of amphibian MHC class I.
Methods
Sample collection and nucleic acid isolation
We selected three Rana frog species that are commonly found across Japan: the Japanese brown frog (Rana japonica), the montane brown frog (Rana ornativentris) and Tago’s brown frog (Rana tagoi tagoi). For genetic material, adult individuals (n = 7 per species) originating from separate locations within Hiroshima prefecture, Japan, were used: R. japonica raised in captivity and originating from multiple egg clusters collected from an island population in Etajima (34°16′14″N, 132°28′37″E); R. ornativentris raised in captivity from tadpoles collected from Yoshiwa (34°25′04″N, 132°05′15″E); and R. t. tagoi from adult frogs collected from Shobara (34°05′04″N, 132°49′43″E). Adult frogs were euthanized by immersion in tricaine methanesulfonate (MS222, 0.5–3 g/L water) and spleen was collected. Total RNA was extracted using ISOGEN (Nippon Gene, Tokyo, Japan) following the manufacturer’s protocol, and first-strand complementary DNA was synthesised using PrimeScriptTM RT reagent kit (Takara Bio Inc., Otsu, Japan).
MHC class I PCR
Primers were designed to amplify partial coding sequence of MHC class I, spanning from exons 1 to 4, based on transcriptome data (T. Igawa, unpublished data) from skin samples of R. japonica as well as several Ranidae species from southern Japanese islands: Odorrana narina, Odorrana amamiensis, Odorrana supranarina, Odorrana ishikawae, Odorrana splendida, Babina holsti and Babina subaspera. Based on conserved regions in aligned sequences, we designed primers using Primer3 (Rozen and Skaletsky 1998): forward 5′-GTGTCAGGRGTGKAKTGTG-3′ and reverse 5′-GAAYMTMCTCCAGACTGCTGT-3′, the latter being similar to that of Kiemnec-Tyburczy et al (2012).
Polymerase chain reaction (PCR) amplification was performed in Applied Biosystems® Veriti® thermal cycler in 10 μl reactions containing 0.2 U KOD FX Neo DNA polymerase (Toyobo, Osaka, Japan), 1× PCR buffer, 0.4 mM each dNTP and 0.5 μM each primer. General cycle conditions included initial Taq activation at 95 °C for 1 min, followed by 30 cycles of 30-s denaturation at 95 °C, 30-s annealing at 56 °C and 30-s extension at 72 °C, then a final extension of 72 °C for 3 min. 3′-dA overhangs were then added to PCR products using 10× A-attachment mix (Toyobo), and then the products were ligated into T-Vector pMD20 (Takara Bio Inc.) using DNA Ligation Kit 2.1 (Takara Bio Inc.) and incubated for 16 °C for 30 min. For cloning, ligation reactions were transformed into One Shot® TOP10 competent cells (Invitrogen, Tokyo, Japan). Positive clones (8–16 per individual) were amplified with M13 primers in similar PCR conditions, purified with ExoSAP-IT® (Affymetrix Inc., Santa Clara, USA) and sequenced with BigDye® Terminator Cycle Sequencing Kit (Applied Biosystems, Foster City, USA) and an ABI 3130xl automated sequencer.
Sequence and phylogenetic analyses
All ABI trace files were edited and analysed using CodonCode Aligner 5.1.5 (CodonCode Co., Centerville, USA). To avoid sequences resulting from PCR or cloning artefacts, we used the following criteria for inclusion of sequences: (1) amplified from more than one clone and (2) differ from other sequences by at least three nucleotides. We allowed for two exceptions where pairs of sequences (Raja-UA*06 and 07, and Raor-UA*19 and 25) differed by only a single non-synonymous substitution at exon 3 (α2 domain) as they were confirmed by repeat PCR and cloning reactions. For each species, we used MEGA7 (Kumar et al. 2016) to calculate measures of overall mean nucleotide and amino acid distances (p distance) across the entire sequence and each domain independently. Using ClustalW, the derived class I amino acid sequences were aligned with those of previously studied amphibian species, including Ambystoma mexicanum, X. laevis, Rhacophorus omeimontis, Polypedates megacephalus, Agalychnis callidryas, Espadarana prosoblepon, Smilisca phaeota, L. catesbeianus, L. clamitans and L. yavapaiensis (Flajnik et al. 1999; Kiemnec-Tyburczy et al. 2012; Sammut et al. 1997; Zhao et al. 2013). For phylogenetic analyses, neighbour-joining trees (p distance) were constructed from amino acid alignments in MEGA7 using 1000 bootstrap replicates. Xenopus non-classical Ib and Gallus gallus class I sequences were also included, with the latter used as an outgroup. Trees were constructed separately for α1, α2 and α3 domains. Maximum-likelihood trees (JTT model + G + I as model of best fit, 500 bootstrap replicates) were also constructed in MEGA7 to confirm inferred phylogenetic relationships.
Tests for recombination and selection
Following Zhao et al (2013), we used two approaches to check for the presence of recombination, briefly: (1) GARD (genetic algorithm recombination detection; Kosakovsky Pond et al. 2006) executed in the online Datamonkey website (http://www.datamonkey.org) and (2) RDP4 (Recombination Detection Program, version 4) which implements multiple methods (Martin et al. 2015). Within RDP4, we only considered breakpoints that were identified by at least four of the seven methods tested (RDP, BOOTSCAN, GENECONV, MAXCHI, CHIMAERA, SISCAN and 3SEQ).
Non-synonymous substitutions per non-synonymous site (d N) and synonymous substitutions per synonymous site (d S) were calculated in MEGA7 with the Nei–Gojobori method (proportion). We used codon-based Z tests in MEGA7 (10,000 bootstrap replicates, partial deletion) to test for global neutral (d N = d S), positive (d N > d S) and purifying (d N < d S) selections in MHC class I sequences independently for each of the three species. In addition, sequence-wide positive selection was also tested using PARRIS, which is implemented in the Datamonkey website. To test for signatures of positive selection on specific codon sites, we used omegaMap version 5.0 (McVean and Wilson 2006) as well as three methods hosted by the Datamonkey website: fixed effects likelihood (FEL), random effects likelihood (REL) and mixed effects model of evolution (MEME) models. These tests were conducted independently in each of the species as species-specific selection pressures are expected and omegaMap makes the assumption of random mating within all individuals. We used omegaMap to perform Bayesian inference on independent alignments for each species and inferred positively selected codon sites (ω or d N/d S > 1) even in the presence of recombination. For each species, we conducted two independent omegaMap runs following the same conditions as Lau et al (2015), but with 1 × 106 MCMC iterations and 100,000 burn-in iterations. We used conservative cutoff probabilities for identification of selected sites; for omegaMap, ω > 1 with posterior probability >0.95, FEL and MEME p values <0.05 and REL Bayes factor >200. We considered a codon site as being under selection (or positively selected site, PSS) if it could be identified in at least two of the four methods used.
Results and discussion
MHC sequence characterisation and copy number variation
We identified a total of 10, 28 and 22 MHC class I sequences (referred to here as variants) from R. japonica, R. ornativentris and R. t. tagoi, respectively (Table 1); these variants are designated Raja-UA*01–10, Raor-UA*01–28 and Rata-UA*01–22 (Genbank accession nos. KX100486–KX100545). These newly characterised variants were all 780 bp (260 codons) in length (Fig. 1), with the exception of Raja-UA*02 (259 codons) and Rata-UA*21 and 22 (262 codons). All variants likely belong to classical class I loci as they were amplified from RNA, clustered phylogenetically with other classical class I sequences (Fig. 2) and had no premature stop codons or aberrant indels. There were no MHC class I variants shared between the species studied. Within R. japonica, many variants were shared across individuals, especially Raja-UA*01 (amplified from six of seven individuals) and Raja-UA*03 and Raja-UA*04 (amplified from four of seven individuals each); this could be attributed to the lower effective population size of this island population studied (T. Igawa, unpublished data). In contrast, there were no shared MHC class I variants between the seven R. t. tagoi individuals studied and only one shared variant (Raor-UA*06) between two R. ornativentris individuals (Supplementary material 1A). Our initial characterisation of MHC class I sequences in these three Japanese frog species provides the framework for future studies of genetic diversity of these abundant species across their broad habitat range across Japan.
Table 1.
Summary of MHC class I variants, genetic divergence and codon-based Z tests for selection in the three Japanese Rana species
| Species | No. of variants (total) | No. of variants (per individual) | Total no. of segregating sitesa | Range of divergence within individual | Mean distance (nucleotide, amino acid) | Z tests for selection for α1 and α2 domains combined (Z statistic) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Nucleotide | Amino acid | Nucleotide | Amino acid | α1 | α2 | α3 | Neutrality | Purifying | Positive | |||
| R. japonica | 10 | 2–5 | 136 | 78 | 0.012–0.115 | 0.031–0.212 | 0.064, 0.135 | 0.080, 0.155 | 0.024, 0.045 | 4.199* | −4.204 n.s. | 4.219** |
| R. ornativentris | 28 | 3–5 | 192 | 94 | 0.063–0.104 | 0.108–0.173 | 0.067, 0.158 | 0.067, 0.122 | 0.035, 0.048 | 3.446* | −3.422 n.s. | 3.372* |
| R. t. tagoi | 22 | 2–5 | 167 | 83 | 0.050–0.101 | 0.100–0.169 | 0.081, 0.149 | 0.081, 0.152 | 0.036, 0.060 | 3.808** | −3.808 n.s. | 3.920** |
*p < 0.01, **p < 0.0001, n.s.: p > 0.05
aExcludes insertions or deletions
Fig. 1.

Amino acid alignment of selected MHC class I variants from R. japonica (Raja), R. ornativentris (Raor) and R. t. tagoi (Rata). Alignment is divided into the three domains studied. Codon sites shaded in grey are detected as under positive selection independently in each species by at least two of four methods used (omegaMap, FEL, REL and MEME); referred to as positively selected sites (PSS). Putative PBR sites inferred from humans (Bjorkman et al. 1987; Saper et al. 1991) are indicated by black bars above the leading sequence. Amino acid sequence for a well-characterised HLA allele, HLA-B*35, is also included (HLA-B*35:01:01:01, IMGT/HLA acc. no. HLA00237)
Fig. 2.
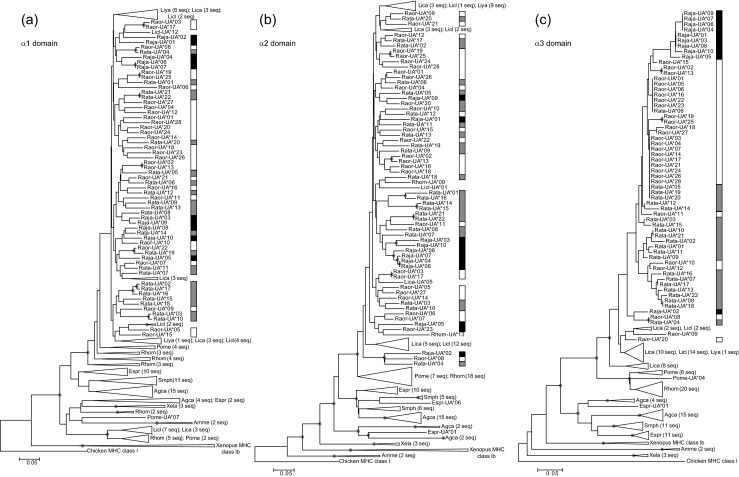
Phylogenetic relationships between MHC I variants identified in R. japonica (Raja, black bar), R. ornativentris (Raor, white bar) and R. t. tagoi (Rata, grey bar) and other amphibians (neighbour-joining method). We constructed phylogenies independently for the α1 domain (a), α2 domain (b) and α3 domain (c). Shaded circles indicate nodes with bootstrap support >70 %. Accession numbers for sequences from other species include Gallus gallus MHC class I (L28958.2); Xenopus MHC class Ib (NM_001247995, FJ589643, L20726); MHC class Ia: X. laevis Xela (AF185580, AF185582, AF185583), Ambystoma mexicanum Amme (U83137, U83138), Rhacophorus omeimontis Rhom and Polypedates megacephalus Pome (KC261637–KC261663), Agalychnis callidryas Agca, Espadarana prosoblepon Espr, Smilisca phaeota Smph, Lithobates catesbeianus Lica, L. clamitans Licl and L. yavapaiensis Liya (JQ679312–JQ679390)
Two to five sequences were characterised from each individual (Supplementary material 1A); it is likely we have underestimated the total number variants per individual due to conservative approaches for confirming MHC variants, similar to Zhao et al (2013), as well as the limited number of sequenced clones. Future studies of additional individuals and populations will likely uncover more MHC class I variation in these expansive and unthreatened species. The sequence divergence within individuals at the nucleotide and amino acid levels (intra-individual variation) was similar amongst the three study species (Table 1 and Supplementary material 1A). The variation is also comparable to the six frog species (n = 5 individuals per species) studied by Kiemnec-Tyburczy et al (2012), although higher amino acid divergence was detected in some A. callidryas (0.451) and E. prosoblepon (0.336) individuals. This could be related to the unusually low levels of per-site differences at MHC class I in the three Japanese Rana species, especially the average synonymous differences per synonymous site (d S; Fig. 3). Subsequently, the resulting d N/d S ratio (or ω) of MHC class I from R. japonica (ω = 1.67), R. ornativentris (ω = 1.07) and R. t. tagoi (ω = 1.26) is higher than that of previously studied frogs (ω = 0.64–0.76), with the exception of L. yavapaiensis (ω = 1.02).
Fig. 3.

Average number of non-synonymous differences per non-synonymous site (d N, grey bars) or synonymous differences per synonymous site (d S, white bars) in MHC class I of various frog species using the Nei–Gojobori model (proportion) with 1000 bootstrap replicates. Error bars represent standard error estimates; circles represent maximum d N or d S from a pair of variants. Sequences from other anuran species were characterised by Kiemnec-Tyburczy et al (2012) and Zhao et al (2013)
Since up to five variants were found in each individual, up to three loci were amplified; this aligns with recent anuran studies that have found MHC class I copy number variation, including up to three loci in some Ranidae species (Kiemnec-Tyburczy et al. 2012; Zhao et al. 2013). As with MHC studies in other non-model species, it remains difficult to completely map out the total number of MHC loci in these species without the availability of whole genome sequences. Comparison of maximum synonymous divergence between assumed paralogous sequences (that is, the most divergent sequence pairs) showed lower maximum d S in the three focal species (Fig. 3), indicative of a more recent gene duplication relative to other frogs studied.
The total number of variants in the studied cohort of R. japonica was lower than that of the other two focal species, likely due to the population history or kin relationships. Despite the lower total number of variants and segregating sites in R. japonica, this cohort had a higher maximum intra-individual sequence variation and comparable overall mean sequence distance relative to the other two species (Table 1). This suggests that the sufficient sequence diversity could offset the low variant diversity in the R. japonica Etajima cohort. Further studies of more extant populations of R. japonica may reveal whether there is additional sequence variation in this species. The α1 and α2 domains (exons 2 and 3, respectively) in all species had higher mean genetic distances than the α3 domain (exon 4). This is consistent with the theory that the α3 domain is more conserved and functionally constrained whilst the α1 and α2 domains have high genetic variation for diverse peptide recognition capacity (Colombani 1990).
Phylogenetic relationships and trans-species polymorphism
We constructed phylogenetic trees of each domain of MHC class I from the three Japanese Rana species along with previously studied amphibians. Within the three focal species, we found no species-specific phylogenetic clustering at the α1 and α2 domains, whereas there were possible monophyletic groups in the α3 domain, especially for R. japonica (Fig. 2 and Supplementary material 1B). Whilst there was generally weak bootstrap support across most branches, which restricts us from making strong conclusions regarding trans-species polymorphism, this polyphyletic pattern is consistent with previous frog MHC class I studied by Kiemnec-Tyburczy et al (2012) and Zhao et al (2013). For α1 and α2 domains, we observed that some variants from other Ranidae species clustered together with Japanese Rana (Fig. 2a, b); in addition, within the α2 domain, there was clustering with sequences from more extant Rhacophoridae species. Despite the low branch support, there were three variants, one from each of the focal species (Raja-UA*02, Raor-UA*08 and Rata-UA*04), that formed a distinct basal polyphyletic group in the α2 and α3 domains (Fig. 2 and Supplementary material 1B). Although these three variants were only identified in one individual per species, they could be derived from a more ancient MHC locus that has been maintained since the three Japanese species diverged, adding support for trans-species polymorphism.
Evidence of recombination
Recombination is one of the main mechanisms proposed as evolutionary drivers of high diversity in MHC and generation of novel alleles (Carrington 1999). We tested the full-length MHC class I sequences of each of the three focal species (covering exons 2–4) and found multiple signals of recombination. Using GARD, three to five recombination breakpoints were identified in each species (Table 2), some of which were located near exon boundaries located at nucleotide positions 261 and 546. This supports recombination through the mechanism of exon shuffling, which is commonly found in lower vertebrates such as fishes and amphibians (Bos and Waldman 2006; Wang et al. 2010; Zhao et al. 2013). The RDP programme was more conservative in detecting recombination, identifying only one and three breakpoints in R. ornativentris and R. t. tagoi, respectively (Table 2); many of these breakpoints were concordant (within a 20-bp window) with those detected by GARD (Table 2). We have found evidence of recombination acting on MHC class I sequences of the three focal species, and such recombination was accounted for in subsequent tests for detecting codon-based selection.
Table 2.
Identification of recombination breakpoints in MHC class I variants within each of the three Rana species studied
| Species | GARD: nucleotide breakpoints | RDP | ||
|---|---|---|---|---|
| Nucleotide breakpoint | Recombinant variant(s) | Potential major/minor parent variants | ||
| R. japonica | 92, 329, 493 | None | – | – |
| R. ornativentris | 82, 362, 516 | 510 | Raor-UA*01 and 04 | Unknown/Raor-UA*05 |
| R. t. tagoi | 127, 248, 393, 501, 641 | 194 and 406 | Rata-UA*19 | Rata-UA*20/Rata-UA*04 |
| 510 | Rata-UA*17 and 18 | Rata-UA*06/Rata-UA*08 | ||
Selection acting on MHC sequences
At the amino acid level, balancing selection can maintain alleles and variation for evolutionarily long periods and can even retain similar or identical alleles between diverged species (Klein et al. 1998), which we observed in the phylogenetic relationships (Fig. 2). Additional support for selection in MHC class I genes of the three focal Rana species came from identification of (1) specific codon sites under positive selection and (2) overall selection at the sequence level.
The total number of sites identified to be under positive selection varied between the four codon-based methods used and the three study species (Supplementary material 1C). Therefore, we identified PSS under the criterion of detection by at least two of the four methods and found a total of 18, 30 and 28 PSS in R. japonica, R. ornativentris and R. t. tagoi, respectively (Table 3 and Fig. 1; Supplementary material 1C). Overall, across the three focal species, 18 PSS was defined in the α1 domain and 21 PSS defined in the α2 domain. Of these 39 PSS from Japanese Rana, a majority were identical sites to those defined in other frogs (27 sites) by Kiemnec-Tyburczy et al (2012) and Zhao et al (2013), as well as PBR sites in human leukocyte antigen (HLA; 28 sites) determined by Bjorkman et al (1987) and Saper et al (1991) (Table 3). The previous anuran studies combined more than one species to identify PSS, not considering differences in selection between species, whilst we found just as many PSS independently in the three Japanese Rana species; this includes some selected sites in the α2 domain that may be species-specific (codon sites 124, 125, 133 and 177; Fig. 1 and Table 3). Nonetheless, the many PSS that are in agreement with other anurans and humans are likely to be polymorphic sites that are functionally important for antigen binding and maintained across vertebrate evolutionary history. In the α3 domain, only two PSS were identified across the three study species, re-emphasising that this domain has been conserved at the sequence level and has not been under the impact of diversifying selection.
Table 3.
Codon sites predicted to be under positive selection in the three Japanese Rana species, referred to as positively selected sites
| Species | Codon sites predicted to be under positive selection | ||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| α1 domain | α2 domain | α3 domain | |||||||||||||||||||||||||||||||||||||||
| 5 | 9 | 11 | 22 | 24 | 31 | 34 | 45 | 62 | 63 | 66 | 67 | 69 | 70 | 73 | 74 | 77 | 81 | 94 | 95 | 97 | 99 | 114 | 116 | 117 | 124 | 125 | 133 | 135 | 145 | 149 | 154 | 156 | 160 | 164 | 167 | 171 | 175 | 177 | 255 | 258 | |
| R. japonica | ˄ | ˄ | ˅ | ˅ | ˅ | ˄ | ˄ | ˄ | ˅ | ˄ | ˅ | ˄ | ˄ | ˄ | ˄ | ˄ | ˄ | ˄ | |||||||||||||||||||||||
| R. ornativentris | ˅ | X | X | ˄ | ˄ | ˅ | X | X | X | ˅ | X | X | X | ˄ | X | X | X | ˅ | ˅ | X | ˅ | ˅ | ˅ | ˄ | X | X | ˅ | ˄ | ˄ | ˅ | |||||||||||
| R. t. tagoi | ˄ | X | X | ˅ | ˄ | ˅ | ˄ | X | ˄ | X | X | X | ˄ | ˄ | ˅ | ˄ | ˅ | X | ˄ | ˄ | X | X | ˄ | ˅ | ˄ | ˄ | X | ||||||||||||||
| Humana | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y | – | – | |||||||||||
| Other frogsb | – | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y | ||||||||||||
Sites are considered to be under positive selection if detected by at least two (˄), three (˅) or all (X) of the four methods used (omegaMap, FEL, REL and MEME). Many PSS were also deduced or predicted to be selected sites (Y) in humans or other frogs. Codon sites are based on the alignment in Fig. 1. ‘–’ indicates the site was not tested
aDeduced PBR sites from HLA based on Bjorkman et al (1987) and Saper et al (1991)
bPredicted selected sites in other frog species based on Kiemnec-Tyburczy et al (2012) and Zhao et al (2013)
Selection tests over the entire sequence showed evidence of positive selection (Z = 1.950–4.024, p < 0.05) and rejection of neutrality and purifying selection (Supplementary material 1D). We further examined selection at each domain and found that the α1 and α2 domains showed evidence supporting positive selection (p < 0.01) and against neutrality and purifying selection (Table 1 and Supplementary material 1D). We also identified similar support for positive selection when using PARRIS (Supplementary material 1D). Meanwhile, the α3 domain showed no sequence-wide positive selection (Z = −2.373–0.0431, p > 0.40), and for the particular case for R. ornativentris, there was even evidence of purifying selection at this domain (Z = 2.335, p = 0.011; Supplementary material 1D). These sequence-wide selection tests contribute to supporting the functional roles of the α1 and α2 domains and the conserved nature of the α3 domain.
Interestingly, Kiemnec-Tyburczy et al (2012) used one-tailed Z tests and found no evidence of selection in any domain in any of the six anuran species that were studied, whilst PARRIS could detect positive selection. We confirmed this pattern in those sequences as well as sequences isolated from Rhacophoridae species by Zhao et al (2013), supporting that the positively selected sites in MHC class I are masked by the large number of neutrally evolving sites. The inability to detect overall neutrality at MHC class I in the three focal study species here could be attributed to either lower per-site synonymous differences and/or stronger selection pressure acting on the molecule. The small extent of synonymous differences (d S) of MHC class I within Japanese Rana species (Fig. 3) could be explained by both within- and between-loci divergence. Low per-site differences in variants (within-loci) could be attributed to severe bottlenecks associated with hypothesised shrinkage of ancestral frog populations during migration from the Asian continent to the Japanese archipelago, perhaps restricted through land bridges (Igawa et al. 2006; Komaki et al. 2015). For between-loci divergence, lower maximum synonymous differences (Fig. 3) in Japanese Rana species support a more recent duplication of loci and subsequent diversification of paralogous variants. The low d S results in a positive and inflated average ω (d N/d S) relative to other anurans, which suggests that the selection pressure acting on MHC class I in Japanese Rana species may be stronger than in other frog species studied. More specifically, the strong positive selection pressure could be targeted at peptide binding sites since we found a similar number of PSS in the three focal species compared to other frogs.
Conclusion
Here, we have characterised the MHC class I classical variation in three Japanese true Rana frog species, with high diversity found in these common unthreatened species. We have identified evidence supporting three major mechanisms that drive high MHC diversity. Signatures of recombination, balancing selection and recent gene duplication found in Japanese Rana MHC class I are similar to those of previous anuran studies. This study is the springboard for investigating genetic variation at other adaptive loci in Japanese amphibians, which will help to better understand the polygenetic basis of resistance to infectious diseases such as chytridiomycosis.
Electronic supplementary material
Below is the link to the electronic supplementary material.
(PDF 534 kb)
Acknowledgments
We thank Naoyuki Takahata for checking the manuscript and the staff at Hiroshima University Institute for Amphibian Biology. This project and QL is financially supported by the Japan Society for the Promotion of Science (JSPS).
Compliance with ethical standards
Research funded by the Japan Society for the Promotion of Science (JSPS). All sample collection is approved by Hiroshima University Animal Research Committee, approval number G14-2.
Conflict of interest
The authors declare that they have no potential conflict of interest.
References
- Bataille A, Fong JJ, Cha M, Wogan GOU, Baek HJ, Lee H, Min MS, Waldman B. Genetic evidence for a high diversity and wide distribution of endemic strains of the pathogenic chytrid fungus Batrachochytrium dendrobatidis in wild Asian amphibians. Mol Ecol. 2013;22:4196–4209. doi: 10.1111/mec.12385. [DOI] [PubMed] [Google Scholar]
- Bataille A, Cashins SD, Grogan L, Skerratt LF, Hunter D, McFadden M, Scheele B, Brannelly LA, Macris A, Harlow PS, Bell S, Berger L, Waldman B. Susceptibility of amphibians to chytridiomycosis is associated with MHC class II conformation. Proc R Soc B Biol Sci. 2015;282:20143127–20143127. doi: 10.1098/rspb.2014.3127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berger L, Speare R, Daszak P, Green DE, Cunningham AA, Goggin CL, Slocombe R, Ragan MA, Hyatt AD, McDonald KR, Hines HB, Lips KR, Marantelli G, Parkes H. Chytridiomycosis causes amphibian mortality associated with population declines in the rain forests of Australia and Central America. Proc Natl Acad Sci U S A. 1998;95:9031–9036. doi: 10.1073/pnas.95.15.9031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bernatchez L, Landry C. MHC studies in nonmodel vertebrates: what have we learned about natural selection in 15 years? J Evol Biol. 2003;16:363–377. doi: 10.1046/j.1420-9101.2003.00531.x. [DOI] [PubMed] [Google Scholar]
- Bjorkman PJ, Saper MA, Samraoui B, Bennett WS, Strominger JL, Wiley DC. The foreign antigen binding site and T cell recognition regions of class I histocompatibility antigens. Nature. 1987;329:512–518. doi: 10.1038/329512a0. [DOI] [PubMed] [Google Scholar]
- Blaustein AR, Romansic JM, Scheessele EA, Han BA, Pessier AP, Longcore JE. Interspecific variation in susceptibility of frog tadpoles to the pathogenic fungus Batrachochytrium dendrobatidis. Conserv Biol. 2005;19:1460–1468. doi: 10.1111/j.1523-1739.2005.00195.x. [DOI] [Google Scholar]
- Bos DH, Waldman B. Evolution by recombination and transspecies polymorphism in the MHC class I gene of Xenopus laevis. Mol Biol Evol. 2006;23:137–143. doi: 10.1093/molbev/msj016. [DOI] [PubMed] [Google Scholar]
- Carrington M. Recombination within the human MHC. Immunol Rev. 1999;167:245–256. doi: 10.1111/j.1600-065X.1999.tb01397.x. [DOI] [PubMed] [Google Scholar]
- Chambouvet A, Gower DJ, Jirků M, Yabsley MJ, Davis AK, Leonard G, Maguire F, Doherty-Bone TM, Bittencourt-Silva GB, Wilkinson M, Richards TA. Cryptic infection of a broad taxonomic and geographic diversity of tadpoles by Perkinsea protists. Proc Natl Acad Sci U S A. 2015;112:E4743–E4751. doi: 10.1073/pnas.1500163112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Colombani J. Conserved and variable structures in HLA class I molecules: a review. Tissue Antigens. 1990;35:103–113. doi: 10.1111/j.1399-0039.1990.tb01765.x. [DOI] [PubMed] [Google Scholar]
- Daszak P, Cunningham AA, Hyatt AD. Infectious disease and amphibian population declines. Divers Distrib. 2003;9:141–150. doi: 10.1046/j.1472-4642.2003.00016.x. [DOI] [Google Scholar]
- Densmore CL, Green DE. Diseases of amphibians. ILAR J. 2007;48:235–254. doi: 10.1093/ilar.48.3.235. [DOI] [PubMed] [Google Scholar]
- Flajnik MF, Ohta Y, Greenberg AS, Salter-Cid L, Carrizosa A, Du Pasquier L, Kasahara M. Two ancient allelic lineages at the single classical class I locus in the Xenopus MHC. J Immunol. 1999;163:3826–3833. [PubMed] [Google Scholar]
- Garner TWJ, Perkins MW, Govindarajulu P, Seglie D, Walker S, Cunningham AA, Fisher MC. The emerging amphibian pathogen Batrachochytrium dendrobatidis globally infects introduced populations of the North American bullfrog, Rana catesbeiana. Biol Lett. 2006;2:455–459. doi: 10.1098/rsbl.2006.0494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goka K, Yokoyama J, Une Y, Kuroki T, Suzuki K, Nakahara M, Kobayashi A, Inaba S, Mizutani T, Hyatt AD. Amphibian chytridiomycosis in Japan: distribution, haplotypes and possible route of entry into Japan. Mol Ecol. 2009;18:4757–4774. doi: 10.1111/j.1365-294X.2009.04384.x. [DOI] [PubMed] [Google Scholar]
- Goyos A, Sowa J, Ohta Y, Robert J. Remarkable conservation of distinct nonclassical MHC class I lineages in divergent amphibian species. J Immunol. 2011;186:372–381. doi: 10.4049/jimmunol.1001467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haislip NA, Gray MJ, Hoverman JT, Miller DL. Development and disease: how susceptibility to an emerging pathogen changes through anuran development. PLoS ONE. 2011;6(7):e22307. doi: 10.1371/journal.pone.0022307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hedrick PW. Evolutionary genetics of the major histocompatibility complex. Am Nat. 1994;143:945–964. doi: 10.1086/285643. [DOI] [Google Scholar]
- Hughes AL, Yeager M. Natural selection at major histocompatibility complex loci of vertebrates. Annu Rev Genet. 1998;32:415–435. doi: 10.1146/annurev.genet.32.1.415. [DOI] [PubMed] [Google Scholar]
- Igawa T, Kurabayashi A, Nishioka M, Sumida M. Molecular phylogenetic relationship of toads distributed in the Far East and Europe inferred from the nucleotide sequences of mitochondrial DNA genes. Mol Phylogenet Evol. 2006;38:250–260. doi: 10.1016/j.ympev.2005.09.003. [DOI] [PubMed] [Google Scholar]
- Jaratlerdsiri W, Isberg SR, Higgins DP, Miles LG, Gongora J (2014) Selection and trans-species polymorphism of major histocompatibility complex class II genes in the order Crocodylia. PLoS ONE 9(2):e87534. doi:10.1371/journal.pone.0087534 [DOI] [PMC free article] [PubMed]
- Kiemnec-Tyburczy KM, Richmond JQ, Savage AE, Lips KR, Zamudio KR. Genetic diversity of MHC class I loci in six non-model frogs is shaped by positive selection and gene duplication. Heredity (Edinb) 2012;109:146–155. doi: 10.1038/hdy.2012.22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klein J, Sato A, Nagl S, O’hUigín C. Molecular trans-species polymorphism. Annu Rev Ecol Syst. 1998;29:1–21. doi: 10.1146/annurev.ecolsys.29.1.1. [DOI] [Google Scholar]
- Komaki S, Igawa T, Lin SM, Tojo K, Min MS, Sumida M. Robust molecular phylogeny and palaeodistribution modelling resolve a complex evolutionary history: glacial cycling drove recurrent mtDNA introgression among Pelophylax frogs in East Asia. J Biogeogr. 2015;42:2159–2171. doi: 10.1111/jbi.12584. [DOI] [Google Scholar]
- Kosakovsky Pond SL, Posada D, Gravenor MB, Woelk CH, Frost SDW. GARD: a genetic algorithm for recombination detection. Bioinformatics. 2006;22:3096–3098. doi: 10.1093/bioinformatics/btl474. [DOI] [PubMed] [Google Scholar]
- Kumar S, Stecher G, Tamura K (2016) MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol Biol Evol 33:1870–1874. doi:10.1093/molbev/msw054 [DOI] [PMC free article] [PubMed]
- Lau Q, Chow N, Gray R, Gongora J, Higgins DP. Diversity of MHC DQB and DRB genes in the endangered Australian sea lion (Neophoca cinerea) J Hered. 2015;106:395–402. doi: 10.1093/jhered/esv022. [DOI] [PubMed] [Google Scholar]
- Lillie M, Shine R, Belov K (2014) Characterisation of major histocompatibility complex class I in the Australian cane toad, Rhinella marina. PLoS ONE 9(8):e102824. doi:10.1371/journal.pone.0102824 [DOI] [PMC free article] [PubMed]
- Longcore JE, Pessier AP, Nichols DK. Batrachochytrium dendrobatidis gen. et sp. nov., a chytrid pathogenic to amphibians. Mycologia. 1999;91:219–227. doi: 10.2307/3761366. [DOI] [Google Scholar]
- Longcore JRE, Pessier AP, Halteman WA. Chytridiomycosis widespread in anurans of northeastern United States. J Wildl Manage. 2007;71:435–444. doi: 10.2193/2006-345. [DOI] [Google Scholar]
- Martin DP, Murrell B, Golden M, Khoosal A, Muhire B. RDP4: detection and analysis of recombination patterns in virus genomes. Virus Evol. 2015;1:1–5. doi: 10.1093/ve/vev003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nei M, Rooney AP. Concerted and birth-and-death evolution of multigene families. Annu Rev Genet. 2005;39:121–152. doi: 10.1146/annurev.genet.39.073003.112240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ohta Y, Goetz W, Hossain MZ, Nonaka M, Flajnik MF. Ancestral organization of the MHC revealed in the amphibian Xenopus. J Immunol. 2006;176:3674–3685. doi: 10.4049/jimmunol.176.6.3674. [DOI] [PubMed] [Google Scholar]
- Rozen S, Skaletsky HJ. Primer3. 1998. [Google Scholar]
- Sammut B, Laurens V, Tournefier A. Isolation of MHC class I cDNAs from the axolotl Ambystoma mexicanum. Immunogenetics. 1997;45:285–294. doi: 10.1007/s002510050207. [DOI] [PubMed] [Google Scholar]
- Saper MA, Bjorkman PJ, Wiley DC. Refined structure of the human histocompatibility antigen HLA-A2 at 2.6 Å resolution. J Mol Biol. 1991;219:277–319. doi: 10.1016/0022-2836(91)90567-P. [DOI] [PubMed] [Google Scholar]
- Savage AE, Zamudio KR. MHC genotypes associate with resistance to a frog-killing fungus. Proc Natl Acad Sci U S A. 2011;108:16705–16710. doi: 10.1073/pnas.1106893108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Skerratt LF, Berger L, Speare R, Cashins S, McDonald KR, Phillott AD, Hines HB, Kenyon N. Spread of chytridiomycosis has caused the rapid global decline and extinction of frogs. Ecohealth. 2007;4:125–134. doi: 10.1007/s10393-007-0093-5. [DOI] [Google Scholar]
- Sommer S. The importance of immune gene variability (MHC) in evolutionary ecology and conservation. Front Zool. 2005;2:16. doi: 10.1186/1742-9994-2-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Teacher AGF, Garner TWJ, Nichols RA (2009) Evidence for directional selection at a novel major histocompatibility class I marker in wild common frogs (Rana temporaria) exposed to a viral pathogen (Ranavirus). PLoS ONE 4(2):e4616. doi:10.1371/journal.pone.0004616 [DOI] [PMC free article] [PubMed]
- Wang D, Zhong L, Wei Q, Gan X, He S. Evolution of MHC class I genes in two ancient fish, paddlefish (Polyodon spathula) and Chinese sturgeon (Acipenser sinensis) FEBS Lett. 2010;584:3331–3339. doi: 10.1016/j.febslet.2010.05.065. [DOI] [PubMed] [Google Scholar]
- Wei K, Zhang Z, Wang X, Zhang W, Xu X, Shen F, Yue B. Lineage pattern, trans-species polymorphism, and selection pressure among the major lineages of feline MHC-DRB peptide-binding region. Immunogenetics. 2010;62:307–317. doi: 10.1007/s00251-010-0440-5. [DOI] [PubMed] [Google Scholar]
- Wilson DJ, McVean G. Estimating diversifying selection and functional constraint in the presence of recombination. Genetics. 2006;172:1411–1425. doi: 10.1534/genetics.105.044917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yeager M, Hughes AL. Evolution of the mammalian MHC: natural selection, recombination, and convergent evolution. Immunol Rev. 1999;167:45–58. doi: 10.1111/j.1600-065X.1999.tb01381.x. [DOI] [PubMed] [Google Scholar]
- Zhao M, Wang Y, Shen H, Li C, Chen C, Luo Z, Wu H. Evolution by selection, recombination, and gene duplication in MHC class I genes of two Rhacophoridae species. BMC Evol Biol. 2013;13:113. doi: 10.1186/1471-2148-13-113. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(PDF 534 kb)