

Abstract
Heparan sulfate acetyl-CoA:α-glucosaminide N-acetyltransferase (HGSNAT) catalyzes the transmembrane acetylation of heparan sulfate in lysosomes required for its further catabolism. Inherited deficiency of HGSNAT in humans results in lysosomal storage of heparan sulfate and causes severe neurodegenerative disease, mucopolysaccharidosis III type C (MPS IIIC). MPS IIIC patients can potentially benefit from a therapeutic approach based on active site-specific inhibitors of HGSNAT used as pharmacological chaperons to modify the folding of the mutant protein in the patient’s cells. This research however was hampered by the absence of the assay suitable for high-throughput screening of drug libraries for HGSNAT inhibitors. The existing method utilizing 4-methylumbelliferyl-β-d-glucosaminide (MU-βGlcN) requires the sequential action of two enzymes, HGSNAT and β-hexosaminidase, whereas the radioactive assay with [C14]-AcCoA is complicated and expensive. We describe a novel direct method to assay HGSNAT enzymatic activity using fluorescent BODIPY-glucosamine as a substrate. The specificity of the assay was tested using cultured fibroblasts of MPS IIIC patients, which showed a profound deficiency of HGSNAT activity as compared to normal controls as well as to MPS IIIA and D patients known to have normal HGSNAT activity. Known competitive HGSNAT inhibitor, glucosamine, had similar inhibition constants for MU-βGlcN and BODIPY-glucosamine acetylation reactions. Altogether our data show that novel HGSNAT assay is specific and potentially applicable for the biochemical diagnosis of MPS IIIC and high-throughput screening for HGSNAT inhibitors.
Electronic supplementary material
The online version of this chapter (doi:10.1007/8904_2015_501) contains supplementary material, which is available to authorized users.
Introduction
Mucopolysaccharidosis III type C (MPS IIIC) is a rare autosomal recessive disorder caused by deficiency of the lysosomal membrane enzyme, heparan sulfate acetyl-CoA:α-glucosaminide N-acetyltransferase (HGSNAT), which catalyzes transmembrane acetylation of the terminal glucosamine residues of heparan sulfate prior to their hydrolysis by lysosomal α-N-acetylglucosaminidase reviewed in Feldhammer et al. (2009a). MPS IIIC is clinically characterized by severe central nervous system degeneration and mild somatic abnormalities such as joint stiffness and coarse facies (reviewed in Neufeld and Muenzer 2001; Ruijter et al. 2008; Valstar et al. 2008). No treatment for MPS IIIC is currently available, but one of the potential therapeutic strategies that are currently being explored is the so-called pharmacological chaperone therapy based on partial refolding of mutant enzyme in the patient’s cells by small molecules (in most cases potent competitive inhibitors) specifically binding to the active cite (reviewed in Valenzano et al. 2011; Cox 2012). Previous data demonstrated that the treatment of cultured cell lines derived from MPS IIIC patients having a number of missense mutations with a competitive HGNSAT inhibitor, glucosamine, partially rescued deficient enzymatic activity (Feldhammer et al. 2009b). These data suggested that search for more potent pharmacological chaperones can lead to development of therapeutic options for this disease. Such compounds are routinely identified by screening the chemical libraries for the inhibitors, providing that the high-throughput assay is available. Unfortunately until now methods to assay HGSNAT enzymatic activity remained to be complex or lacked specificity required for high-throughput screening. HGSNAT was initially assayed using radiolabeled oligosaccharides derived from heparan sulfate (Klein et al. 1978). The assay simplified by using commercially available [14C]-labeled monosaccharide glucosamine or [14C]-labeled acetyl CoA as substrates was further applied for the biochemical diagnosis of MPS IIIC patients and heterozygous carriers using blood leukocytes or cultured skin fibroblasts (Klein et al. 1978; Hopwood and Elliott 1981; Pallini et al. 1984). Since the assay was still somewhat complicated and expensive as other radioactive assays, a fluorogenic derivative of a glucosamine, 4-methylumbelliferyl-β-d-glucosaminide (MU-βGlcN) was synthesized and used as a substrate for measuring enzymatic activity of HGSNAT (Voznyi et al. 1993). In this assay the liberation of the fluorochrome, 4-methylumbelliferyl from MU-βGlcN requires the sequential enzymatic actions of HGSNAT and β-hexosaminidase: β-hexosaminidase hydrolyses the reaction intermediate, 4-methylumbelliferyl-β-d-N-acetyl-glucosaminide formed by HGSNAT. Since no enzyme in mammalian cells is capable of hydrolyzing non-acetylated MU-βGlcN, the assay can be successfully used for biochemical diagnostics of MPS IIIC patients (Voznyi et al. 1993). Although the fluorescent assay is obviously more convenient than the radioactive assay, it is however unsuitable for identification of HGSNAT inhibitors since it involves two enzymatic steps. In particular, when we attempted to use MU-βGlcN-based assay to screen a library of 1,040 FDA-approved compounds (Huang et al. 2011), multiple inhibitors of β-hexosaminidase have been falsely identified as HGSNAT inhibitors (Buhas D. unpublished).
Here we describe a novel direct fluorometric HGSNAT assay utilizing glucosamine labeled at position C1 with BODIPY as a substrate and potentially suitable for both biochemical testing of MPS IIIC and high-throughput screening. In this method, HGSNAT converts positively charged substrate into the neutral product, BODIPY-N-acetyl-glucosamine, which is further separated from the substrate using cation exchange chromatography and quantified using a standard fluorometer plate reader.
Materials and Methods
Synthesis of BODIPY-Glucosamine
2-Azido-2-deoxy-d-glucose was from Carbosynth Ltd (Compton, Berkshire, UK), and succinimidyl ester of 4.4-difluoro-5,7-dimethyl-4-bora-3a,4a-diaza-s-indacene-3-propionic acid (BODIPY® FL, SE) was from Invitrogen (USA). Reactions were performed with the use of commercial reagents (Acros, Aldrich, and Fluka); anhydrous solvents were purified according to the standard procedures. Column chromatography was performed on silica gel 60 (particle size 0.040–0.063 mm, Merck), and gel filtration was carried out on Sephadex LH-20 (Pharmacia) columns. Solvents were removed by evaporation in vacuum at 30–40°С. Thin-layer chromatography (TLC) was performed on silica gel 60 F254 aluminum-backed plates (Merck). Compounds on TLC plates were visualized by spraying the plates with either 8% H3PO4 in H2O (followed by heating) or 2% ninhydrin in ethanol. 1Н NMR spectra were recorded on a Bruker BioSpin GmbH (700 МHz) spectrometer at 30°С, chemical shifts (δ, ppm) were referred to the peak of internal D2O (δ 4.750 ppm), and coupling constants (J) were measured in Hz. MALDI-TOF mass spectra were recorded on Bruker Daltonics Ultraflex MALDI-TOF/TOF mass spectrometer (Germany) using DHB matrix.
To produce 2-deoxy-2-azido-Glcβ-NH(CO)CH2NH2 (4), 2-deoxy-2-azido-Glc (1) (294 mg, 1.43 mmol) and powdered ammonium carbamate (447 mg, 5.73 mmol) were dissolved in a mixture of methanol (3.6 mL) and water (0.036 mL) at 40°C (Fig. 1). The solution was kept for 42 h at 40°C, cooled to room temperature, and then kept for 15 h at 4°C. The solution was diluted with water (1 mL), evaporated and dried in vacuum. The residue was dissolved in water (9 mL) and freeze-dried (at 0.002 mBar for 16 h). The mass of the residue corresponded to glycosylamine carbamate (2) (351 mg, 92%).
Fig. 1.
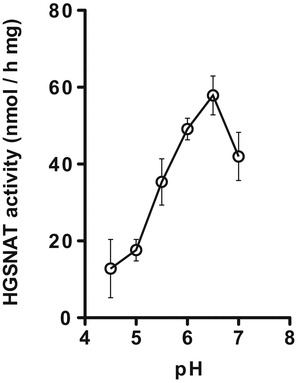
Dependence of the enzymatic activity of human recombinant HGSNAT measured with BODIPY-glucosamine on the pH of the reaction mixture. The reaction mixture contained 6 μL of McIlvain’s phosphate/citrate buffer (pH 4.5, 5.0, 5.5, 6.0, 6.5, and 7.0), 4 μL of 10 mM acetyl CoA in water, 4 μL of 40 μM BODIPY-glucosamine, and an aliquot of homogenate of COS-7 cells expressing human WT HGSNAT protein corresponding to 6 μg of protein. The data show means (±S.E.) of three independent measurements
To an intensively stirred solution of 2-deoxy-2-azido-Glc glycosylamine carbamate (2) (351 mg) in cooled (10°C) 1 M aqueous NaHCO3 (10.1 mL), a solution of chloroacetic acid anhydride (980 mg, 5.7 mmol) in toluene (5 mL) was added. The mixture was stirred for 30 min at room temperature, acidified with AcOH (164 μL), and dried.
The residue consisting of N-chloroacetamide (3) and sodium chloroacetate was dissolved in water (1 mL); 10 M aqueous NH3 (20 mL) was then added and the solution was kept for 24 h at room temperature. The solvent was further evaporated, the residue dissolved in water (3 mL) was acidified with AcOH (300 μL), and the solution was kept for 16 h at room temperature. Products were separated by ion-exchange chromatography on a Dowex 50X4-200 (H+) 40 mL column. The resin was washed with water (40 mL) and then with 1 M aqueous pyridine (70 mL) to remove all acidic and neutral components including glycine formed from excessive chloroacetate (Tuzikov et al. 2000). Further elution with 1 M aqueous NH3 (75 mL) yielded 243 mg (65%) of material consisting of target compounds 2-deoxy-2-azido-Glcβ-NH(CO)CH2NH2 (4) (90%), 2-deoxy-2-azido-Glcα-NH(CO)CH2NH2 (5%), and 2-deoxy-2-azido-Manβ-NH(CO)CH2NH2 (5%) according to 1H NMR data. Crystallization from MeOH/2-propanol (3 mL/2 mL (cr.1), m.1 from 1 mL/1 mL (cr.2), and cr.1+cr.2 from 1 mL/1 mL) yielded slightly yellow crystals of 2-deoxy-2-azido-Glcβ-NH(CO)CH2NH2. Gel-permeation chromatography on Sephadex G-10 (90 mL) in MeOH/0.1 M aqueous NH3 (1:3 v/v) and freeze-drying yielded 105 mg of pure colorless compound (4).
TLC: R f = 0.47 (2-propanol-MeOH-1 M aqueous Py•HOAc 2:3:1 v/v).
1H NMR of 2-deoxy-2-azido-Glcβ-NH(CO)CH2NH2 TFA salt (1) (700 MHz, D2O) δ 3.482 (~t, J = 9.6 Hz, 1H; H-2), 3.503 (~t, J = 9.5 Hz, 1H; H-4), 3.561 (ddd, J = 9.7, 5.3, 1.9 Hz, 1H; H-5), 3.672 (~t, J = 9.4 Hz, 1H; H-3), 3.753 (dd, J = 12.5, 5.3 Hz, 1H; H-6), 3.897 (dd, J = 12.5, 1.9 Hz, 1H; H-6`), 3.918 (d, J = 16.4 Hz, 1H; NCH), 3.948 (d, J = 16.4 Hz, 1H; NCH), and 5.073 (d, J = 9.7 Hz, 1H; H-1).
MALDI-TOF mass spectrum of 2-deoxy-2-azido-Glcβ-NH(CO)CH2NH2, m/z 236: [(M-N2+2H)+H], 262 [M+H], 284 [M+Na], and 300 [M+K].
To produce 2-deoxy-2-azido-Glcβ-NH(CO)CH2NH-BODIPY (6), N-glycylglycosylamine derivative of 2-azidoglucose (4) (6,7 mg, 0.026 mmol) was added to a solution of 10 mg (0.026 mmol) BODIPY® FL, SE (5) in 1 mL DMSO. The reaction mixture was kept for 5 min at 20°C, then 1h at −20°C. After the completion of reaction controlled by TLC in EtOH/BuOH/Py/H2O/AcOH (100:10:10:10:2 v/v), the solution was purified by passing through Sephadex LH-20 column (CH3CN/H2O, 1:1 v/v). Fractions containing BODIPY-derivative (6) were combined and concentrated by drying in vacuum. Column chromatography of the residue on silica gel (CH2Cl2/EtOAc, 8:1→6:1 v/v) yielded 10.5 mg (76%) of BODIPY-derivative (6). TLC: R f 0.4 (CH2Cl2-EtOAc 6:1 v/v). 1H NMR of (6) (700 MHz, D2O): δ 2.26 (s, 3H, CH3 BODIPY), 2.53 (s, 3H, CH3 BODIPY) 2.80 (t, 2H, J = 7.2, CH2 BODIPY), 3.22 (t, 2H, J = 7.2 COCH2 BODIPY), 3.33 (dd ≈t, 1H, J 1,2 ≈ J 2,3 ≈9.6, H-2), 3.37 (dd ≈ t, 1H, J 3,4 ≈ J 4,5 ≈ 9.5, H-4), 3.48 (m, 1H, H-5), 3.58 (dd ≈ t, 1H, J 2,3 ≈ J 3,4 ≈ 9.4, H-3), 3.67 (dd,1H, J 5,6a = 5.7, J 6a,6b = 12.4, H-6a), 3.85 (dd, 1H, J 5,6b = 1.6, J 6a,6b = 12.4, H-6b), 3.97 (m, 2H, COCH2NH), 4.97 (d, 1H, J 1,2 = 9.5, H-1), 6.32 (s, 1H, BODIPY), 6.43 (d, 1H, J = 3.8, BODIPY), 7.08 (d, 1H, J = 3.4, BODIPY), and 7.46 (s, 1H, BODIPY).
To produce 2-deoxy-2-amino-Glcβ-NH(CO)CH2NH-BODIPY (7), a solution of triphenylphosphine in THF/H2O (3:1,v/v) (10 mg/mL, 1.8 mL, 0.07 mmol) was added to a solution of 7.5 mg (0.014 mmol) of azido compound (6) in 1.5 mL THF/H2O (3:1,v/v). The reaction mixture was stirred overnight at 20°C. The solution was concentrated in vacuum, the residue was dissolved in 2 mL of H2O, and the excess of triphenylphosphine was removed by extraction with toluene (3 × 4 mL). Gel chromatography on Sephadex LH-20 (CH3CN/H2O, 1:1, 0.05% AcOH) gave 5.6 mg (80%) of amino compound (7) in the form of AcOH salt. TLC: R f 0.4 (CH2Cl2-EtOH-H20 6:5:1). 1H NMR (700 MHz, D2O): δ 2.26 (s, 3H, CH3 BODIPY), 2.53 (s, 3H, CH3 BODIPY), 2.81 (t, 2H, J = 7.2, CH2 BODIPY), 2.96 (dd ≈ t, 1H, J 1,2 ≈ J 2,3 ≈ 10.0, H-2), 3.22 (t, 2H, J = 7.4, COCH2 BODIPY), 3.41 (dd ≈ t, 1H, J 3,4 ≈ J 4,5 ≈ 9.3, H-4), 3.55 (m, 1H, H-5), 3.61 (dd ≈ t, 1H, J 2,3 ≈ J 3,4 ≈ 9.5, H-3), 3.71 (dd,1H, J 5,6a = 5.1, J 6a,6b = 12.4, H-6a), 3.88 (d, 1H, J 5,6b < 1, J 6a,6b = 11.9, H-6b), 4.01 (m, 2H, COCH2NH), 5.19 (d, 1H, J 1,2 = 9.5, H-1), 6.32 (s, 1H, BODIPY), 6.42 (d, 1H, J = 3.0, BODIPY), 7.07 (d, 1H, J = 2.8, BODIPY), and 7.45 (s, 1H, BODIPY). MALDI-TOF mass spectrum, m/z 510 [M+H], 532 [M+Na], and 548 [M+K].
Other researchers can get access to the BODIPY-glucosamine substrate by contacting the PI.
Production of Recombinant Human HGSNAT in COS-7 Cells
COS-7 cells were cultured in Eagle’s minimal essential medium (EMEM) supplemented with 10% (v/v) fetal bovine serum (Wisent Bio Products) and 1% Gibco’s penicillin/streptomycin antibiotics (Life Technologies). Transfections with pcTAP-HGSNAT plasmid encoding for human wild-type (WT) HGSNAT (Feldhammer et al. 2009b) were carried out in 10-cm petri dishes using 1 M polyethylenimine (PEI) as transfection reagent (Sigma-Aldrich). The plasmid (6–7 μg of DNA) and 24 μL of PEI were diluted in 500 μL of serum-free medium each and then mixed together. Cells were incubated with transfection mix for 24 h, which was then replaced with Eagle’s minimal essential medium (EMEM) supplemented with 1% dimethyl sulfoxide, serum, and antibiotics. Cells were harvested 24 h later in PBS with a rubber scraper, pelleted by 10 min centrifugation at 1,000 g, and homogenized in 1 mL of water by sonication.
Human Skin Fibroblasts Culture
Cultured skin fibroblasts of MPS IIIC patients and normal controls were obtained from cell depositories of Debrousse Hospital (France), Montreal Children’s Hospital (Canada), NIGMS Human Genetic Mutant Cell Repository and Department of Clinical Genetics, and Erasmus Medical Center (the Netherlands). Skin fibroblasts were grown to 100% confluency in 10-cm petri dishes in Dulbecco’s minimal essential medium supplemented with 10% fetal bovine serum and 1% Gibco’s penicillin/streptomycin. Cells were harvested in PBS with a rubber scraper, pelleted by 10 min centrifugation at 1,000 g, and homogenized in 500 μL of water by sonication. Protein concentration was measured according to the method of Bradford using Bio-Rad reagent.
Enzymatic HGSNAT Assay Using BODIPY-Glucosamine as a Substrate
The reaction mixture for measuring HGSNAT enzymatic activity in fibroblasts or COS-7 cells overexpressing human HGSNAT consisted of 6 μL of homogenate (~20 μg protein), 6 μL of McIlvain’s phosphate/citrate buffer pH 6.5, 4 μL of 10 mM acetyl CoA in water, and 4 μL of 40 μM BODIPY-glucosamine (1-[4,4-difluoro-5,7-dimethyl-4-bora-3a,4a-diaza-s-indacene-3-propionyl-glycylamino]-β-d-glucosamine). A blank sample contained all components except acetyl CoA. After incubation for 17 h at 37°C in 96-well PCR plates (BioScience Inc.), the reaction was terminated by the addition of 180 μL of 100 mM HCl. Twenty microliter aliquots of reaction mixtures were transferred to 96-well filter plate (Millipore, 40 μm nylon mesh) pre-embedded with 100 μL Toyopearl cation exchange media SP 650M (Tosoh) for each well. Prior to the assay, the resin was washed twice with 250 μL of water per well and the plates centrifuged at 50 g for 30 s to remove any excess water. The fluorescent neutral reaction product was eluted with four 90 μL aliquots of 1 M HCl by centrifugation of the plates at 50 g for 30 s. Combined eluent (360 μL) was transferred to 96-well ReaderBlack polystyrene plates (Life Science), and the amount of fluorescent product was measured using an EnVision 2104 Multilabel fluorimeter (PerkinElmer) at emission wavelength of 535 nm and excitation wavelength of 485 nm. Three independent duplicate measurements were performed for each experimental condition. Kinetic parameters of enzymatic reactions were analyzed by nonlinear regression using Prism GraphPad software.
In the experiments aimed on testing the effect of glucosamine on enzymatic activity of HGSNAT, the reaction mixture consisted of 6 μL (~6 μg of protein) of homogenate of COS-7 cells expressing recombinant human WT HGSNAT, 4 μL of 7.5 mM glucosamine in water, 6 μL of McIlvain’s phosphate/citrate buffer pH 6.5, 4 μL of 10 mM acetyl CoA in water, and 4 μL of 40 μM BODIPY-glucosamine. The reaction mixture was incubated for 3 h at 37°C, and after the reaction termination step, the assay followed the procedure for measuring HGSNAT activity in fibroblasts. Enzymatic activity of HGSNAT against MU-βGlcN substrate was measured as previously described (Feldhammer et al. 2009b).
Results and Discussion
The fluorescent derivative of BODIPY-glucosamine (1-[4,4-difluoro-5,7-dimethyl-4-bora-3a,4a-diaza-s-indacene-3-propionyl-glycylamino]-β-d-glucosamine) was synthesized as shown in Scheme 1. First, starting from 2-azido-2-deoxy-d-glucose (1) its N-glycylglycosylamine derivative (4) was synthesized through glycosylamine carbamate (2) and N-chloroacetamide (3) using the procedure previously described for GlcNAc (Likhosherstov et al. 2002). Conditions for reactions of N-chloroacetylation and ammonolysis (Manger et al. 1992) were modified as described in Materials and Methods. The target molecule, fluorescent BODIPY-derivative of 2-amino-2-deoxy-d-glucose (7) was synthesized from (4) condensation with succinimidyl ester of BODIPY (BODIPY® FL, SE) (5) followed by reduction of azido compound (6) with triphenylphosphine. The structures of synthesized compounds were confirmed by 1H NMR spectroscopy and mass spectrometry.
Scheme 1.
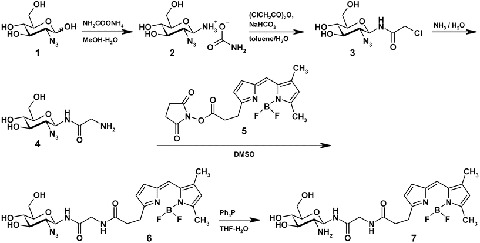
Synthesis of 2-deoxy-2-amino-d-Glcβ-NH(CO)CH2NH-BODIPY
The general scheme for the HGSNAT assay with BODIPY-glucosamine as a substrate is shown in Scheme 2 and the flow chart in the supplemental materials. HGSNAT converts the positively charged substrate, BODIPY-glucosamine, into the neutral product, BODIPY-N-acetyl-glucosamine; the reaction is terminated by addition of 100 mM HCl, and the neutral product is separated from the positively charged substrate using the Toyopearl cation exchange resin SP-650C. The product is further collected into the 96-well plate and its concentration is measured using fluorometer plate reader.
Scheme 2.
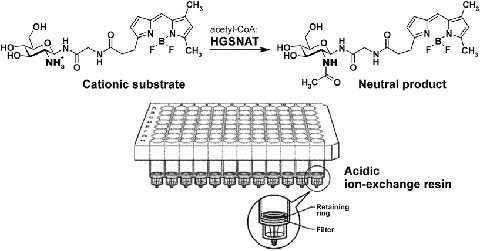
HGSNAT assay with 2-deoxy-2-amino-d-Glcβ-NH(CO)CH2NH-BODIPY substrate
The optimal conditions for the enzyme assay, including the amount of the cation exchange resin, protein concentration, incubation time, and pH of the reaction mixture, were determined experimentally. As shown in Supplementary Fig. 1, increasing the volume of cation exchange resin SP 650M in the filter plate from 25 to 100 μL reduced the amount of positively charged BODIPY-glucosamine substrate present in the product, yielding lower background level for enzyme activity. At the same time, further increase of the resin volume did not reduce the background but increased the amount of buffer necessary to elute all products. The optimal amount of cation exchange resin that had to be placed in each well of the filter plate was therefore estimated as 100 μL (Supplementary Fig. 1). After optimization of the procedure for separation of the product and the substrate, we determined the reaction conditions under which the amount of converted substrate is proportional to the enzyme concentration in the incubation mixture and the reaction time. As shown in Supplemental Fig. 2, the amount of acetylated substrate linearly increased with protein concentration. Also, the activity of HGSNAT increased nearly linearly with incubation time (Supplementary Fig. 3).
We further tested if enzymatic activity of HGSNAT toward BODIPY-glucosamine was dependent on pH and compared the pH optimum of the reaction to that of two other assays that utilized radioactive [14C] glucosamine or MU-βGlcN as substrates. The radioactive assay showed a broad pH optimum for HGSNAT with a plateau between pH 5.5 and 7.5 (Pallini et al. 1984), whereas fluorogenic assay with MU-βGlcN substrate showed that the pH optimum for HGSNAT from leukocytes and fibroblasts was 5.7 (Voznyi et al. 1993). According to our data (Fig. 1), the pH optimum of the human recombinant HGSNAT produced in COS-7 cells was pH 6.5. This pH optimum is slightly higher than that of the fluorogenic assay and is within the optimal pH range of the radioactive assay. It is important to notice, however, that the pH optimum of the HGSNAT assay with MU-βGlcN substrate may represent a combination of the pH optima of 2 enzymatic reactions, catalyzed by HGSNAT and beta-hexosaminidase, respectively. Since human beta-hexosaminidase has acidic pH optimum of 4.2, we suggest that it may reduce the pH optimum as determined by the fluorescent assay, whereas the radioactive assay and the assay with BODIPY-glucosamine reveal the true pH optimum of HGSNAT.
Specificity of the assay was further tested with known competitive HGSNAT inhibitor, glucosamine, and the cultured cells of the MPS IIIC patients having mutations in the HGSNAT gene (Supplementary Table 1). We first studied whether glucosamine would compete for the enzyme binding with BODIPY-glucosamine and decrease the rate of N-acetylation. As shown in Fig. 2, the inhibition profile of HGSNAT by glucosamine was similar when the activity was measured with the fluorescent MU-βGlcN substrate or with BODIPY-glucosamine (Ki values determined using BODIPY-glucosamine and MU-βGlcN assays were 0.26 ± 0.05 mM and 0.28 ± 0.02 mM, respectively). We further determined HGSNAT activity in total homogenates of cultured skin fibroblasts from MPS IIIC patients (n = 5). As shown in Fig. 3, all lines from MPS IIIC patients had a profound deficiency of HGSNAT enzymatic activity as compared to normal controls (n = 9) or to cells from MPS IIIA (n = 3) and D (n = 3) patients which have normal levels of HGSNAT activity.
Fig. 2.
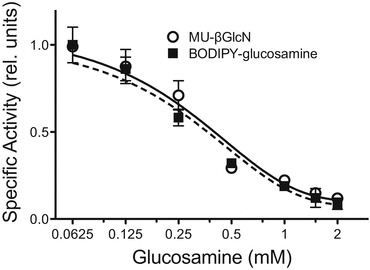
Effect of glucosamine on enzymatic activity of HGSNAT. The specific activity of recombinant HGSNAT expressed in COS-7 cells was measured in the pH optimum using 8 μM BODIPY-glucosamine (squares and solid line) or 37.5 μM MU-βGlcN (open circles and dashed line) as substrate and 2 mM acetyl CoA and glucosamine in the final concentration between 0 and 2 mM. The data (means ± S.D.) of two independent experiments are shown. The activity is presented as the fraction of the activity measured in the absence of glucosamine
Fig. 3.
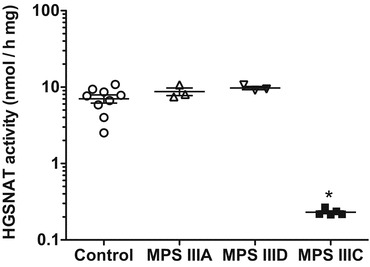
HGSNAT activity in fibroblasts from MPS IIIC patients (N = 5), MPS IIIA patients (N = 3), MPS IIID patients (N = 3), and normal healthy controls (N = 9). The data show means of two individual experiments. *p < 0.05 in unpaired two-tailed t-test
In conclusion, we have described novel method for HGSNAT enzymatic assay using the fluorescent BODIPY-glucosamine. It may be less convenient for the biochemical diagnosis of MPS IIIC than the widely used MU-βGlcN substrate since it includes an additional chromatographic step. However, in contrast to MU-βGlcN method requiring two enzymes to yield the fluorochrome, our method is direct and therefore suitable for high-throughput screening for HGSNAT effectors.
Electronic Supplementary Material
Below is the link to the electronic supplementary material.
Acknowledgments
The authors acknowledge Dr. Mila Ashmarina for the critical reading of the manuscript and helpful advice and Carmen Movila for the help in preparation of the manuscript. This work was supported in part by the operating grant (111068) from the Canadian Institutes of Health Research and JJB Foundation to A.V.P. Synthetic part of this work (ABT, TVO. NVB) was supported by the Russian Science Foundation (project No. 14-50-00131).
Compliance with Ethics Guidelines
Conflict of Interest
Yoo Choi, Alexander B. Tuzikov, Tatyana V. Ovchinnikova, Nicolai V. Bovin, and Alexey V. Pshezhetsky declare that they have no conflict of interest.
Informed Consent and Animal Rights
This article does not contain any studies with human or animal subjects performed by any of the authors.
Details of the Contributions of Individual Authors
Designed experiments: AP, NB; conducted experiments: YC, AT, TO; analyzed data: AP, YC, NB, AT, TO; wrote paper: AP, YC, NB, AT, TO.
Footnotes
Competing interests: None declared
References
- Cox TM (2012) Current treatments. In: Winchester AMAB (ed) Lysosomal storage disorders: a practical guide. Wiley, Oxford, pp 151–165
- Feldhammer M, Durand S, Mrazova L, et al. Sanfilippo syndrome type C: mutation spectrum in the heparan sulfate acetyl-CoA: alpha-glucosaminide N-acetyltransferase (HGSNAT) gene. Hum Mutat. 2009;30:918–925. doi: 10.1002/humu.20986. [DOI] [PubMed] [Google Scholar]
- Feldhammer M, Durand S, Pshezhetsky AV. Protein misfolding as an underlying molecular defect in mucopolysaccharidosis III type C. PloS One. 2009;4 doi: 10.1371/journal.pone.0007434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hopwood JJ, Elliott H (1981) The diagnosis of the Sanfilippo C syndrome, using monosaccharide and oligosaccharide substrates to assay acetyl-CoA: 2-amino-2-deoxy-alpha-glucoside N-acetyltransferase activity. Clin Chim Acta 112:67–75 [DOI] [PubMed]
- Huang R, Southall N, Wang Y et al (2011) The NCGC pharmaceutical collection: a comprehensive resource of clinically approved drugs enabling repurposing and chemical genomics. Sci Transl Med 3:80ps16 [DOI] [PMC free article] [PubMed]
- Klein U, Kresse H, von Figura K. Sanfilippo syndrome type C: deficiency of acetyl-CoA:alpha-glucosaminide N-acetyltransferase in skin fibroblasts. Proc Natl Acad Sci U S A. 1978;75:5185–5189. doi: 10.1073/pnas.75.10.5185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Likhosherstov LM, Novikova OS, Shibaev VN. New efficient synthesis of β-glucosylamines of mono- and disaccharides with the use of ammonium carbamate. Doklady Chem. 2002;383:89–92. doi: 10.1023/A:1015428720733. [DOI] [Google Scholar]
- Manger ID, Rademacher TW, Dwek RA. 1-N-glycyl beta-oligosaccharide derivatives as stable intermediates for the formation of glycoconjugate probes. Biochemistry. 1992;31:10724–10732. doi: 10.1021/bi00159a012. [DOI] [PubMed] [Google Scholar]
- Neufeld EF, Muenzer J. The Mucopolysaccharidoses. In: Scriver CR, Beaudet AL, Sly WS, Valle D, editors. The metabolic and molecular basis of inherited disease. New-York: McGraw-Hill; 2001. pp. 3421–3452. [Google Scholar]
- Pallini R, Leder IG, di Natale P. Sanfilippo type C diagnosis: assay of acetyl-CoA: alpha-glucosaminide N-acetyltransferase using [14C]glucosamine as substrate and leukocytes as enzyme source. Pediatric Res. 1984;18:543–545. doi: 10.1203/00006450-198406000-00013. [DOI] [PubMed] [Google Scholar]
- Ruijter GJ, Valstar MJ, van de Kamp JM, et al. Clinical and genetic spectrum of Sanfilippo type C (MPS IIIC) disease in The Netherlands. Mol Genet Metab. 2008;93:104–111. doi: 10.1016/j.ymgme.2007.09.011. [DOI] [PubMed] [Google Scholar]
- Tuzikov AB, Gambaryan AS, Juneja LR, Bovin NV. Conversion of complex Sialooligosaccharides into polymeric conjugates and their anti-influenza virus inhibitory potency. J Carbohydr Chem. 2000;19:1191–1200. doi: 10.1080/07328300008544143. [DOI] [Google Scholar]
- Valenzano KJ, Khanna R, Powe AC, et al. Identification and characterization of pharmacological chaperones to correct enzyme deficiencies in lysosomal storage disorders. Assay Drug Dev Technol. 2011;9:213–235. doi: 10.1089/adt.2011.0370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Valstar MJ, Ruijter GJ, van Diggelen OP, Poorthuis BJ, Wijburg FA. Sanfilippo syndrome: a mini-review. J Inherit Metab Dis. 2008;31:240–252. doi: 10.1007/s10545-008-0838-5. [DOI] [PubMed] [Google Scholar]
- Voznyi Ya V, Karpova EA, Dudukina TV, et al. A fluorimetric enzyme assay for the diagnosis of Sanfilippo disease C (MPS III C) J Inherit Metab Dis. 1993;16:465–472. doi: 10.1007/BF00710299. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.