

Summary
Across plants, leaves exhibit profound diversity in shape. As a single leaf expands, its shape is in constant flux. Plants may also produce leaves with different shapes at successive nodes. In addition, leaf shape varies among individuals, populations and species as a result of evolutionary processes and environmental influences.
Because leaf shape can vary in many different ways, theoretically, the effects of distinct developmental and evolutionary processes are separable, even within the shape of a single leaf. Here, we measured the shapes of > 3200 leaves representing > 270 vines from wild relatives of domesticated grape (Vitis spp.) to determine whether leaf shapes attributable to genetics and development are separable from each other.
We isolated latent shapes (multivariate signatures that vary independently from each other) embedded within the overall shape of leaves. These latent shapes can predict developmental stages independent from species identity and vice versa. Shapes predictive of development were then used to stage leaves from 1200 varieties of domesticated grape (Vitis vinifera), revealing that changes in timing underlie leaf shape diversity.
Our results indicate that distinct latent shapes combine to produce a composite morphology in leaves, and that developmental and evolutionary contributions to shape vary independently from each other.
Keywords: development, grape (Vitis vinifera), leaf morphology, leaf shape, phenotype
Introduction
Leaf morphology represents a beautiful and tangible example of the infinite phenotypic possibilities in nature. Underlying leaf shape diversity is a quantitative genetic (Langlade et al., 2005; Kimura et al., 2008; Tian et al., 2011; Chitwood et al., 2013) and developmental genetic (Bharathan et al., 2002; Kim et al., 2003; Blein et al., 2008) framework. It is possible that aspects of leaf shape are functionally neutral and reflect developmental constraint (Chitwood et al., 2012a,b), but numerous hypotheses about the function of different leaf shapes exist, including how shape impacts thermal regulation, hydraulic constraints, light interception, biomechanics and herbivory (Parkhurst & Loucks, 1972; Nicotra et al., 2011; Ogburn & Edwards, 2013). Fossil leaf size and dissection are correlated with the paleoclimate (Bailey & Sinnott, 1915; Wolfe, 1971; Greenwood, 1992; Wilf et al., 1998), a relationship that persists in extant taxa (Peppe et al., 2011), and with implications for the chemical, structural and physiological economics of leaves (Wright et al., 2004). Correspondingly, functional traits related to leaf shape display phylogenetic signal in some clades (Cornwell et al., 2014). An understanding of the spatial and temporal patterns of leaf shape variation is a central theme in studies focusing on plant biodiversity, the impacts of global climate change and agricultural efficiency.
Leaf shape varies not only across evolutionary timescales and within a functional ecological context, but during development as well. Two distinct temporal processes regulate leaf shape during development. First, the shape of individual leaves is in constant flux as local regions within the leaf expand at different rates. This phenomenon, allometric expansion, was explored as early as Hales’ Vegetable Staticks (1727). Using a grid of pins, regularly spaced puncture points in fig leaves were tracked to determine whether their relative spacing changed during development. The same experiment can be microscopically studied using fluorescent particles today (Remmler & Rolland‐Lagan, 2012; Rolland‐Lagan et al., 2014). Second, the leaves that emerge at successive nodes differ in their shape, as the shoot apical meristem from which they derive transitions from a juvenile to adult stage of development. This process, heteroblasty, can affect other features of leaves in addition to shape, such as cuticle and trichome patterning (Goebel, 1900; Ashby, 1948; Poethig, 1990, 2010; Kerstetter & Poethig, 1998).
The developmental stage of a leaf and the position of the node from which it arises (leaf number) are distinct temporal factors affecting leaf shape. Genetic changes in the timing of either process (e.g. protracted development of individual leaves or precociously adult leaf morphology in early nodes) between species can lead to evolutionary differences in leaf shape, a process known as heterochrony (Cartolano et al., 2015). The timing of these processes can be changed nongenetically as well, through responses to environmental changes during the lifetime of a plant, known as plasticity (Allsopp, 1954; Diggle, 2002). Distinguishing the effects of developmental stage from leaf number provides mechanistic insights into how genetic changes during evolution or plastic changes in response to the environment are achieved (Jones, 1993, 1995). For example, recent work has linked molecular pathways regulating the timing of shape changes throughout the shoot (miR156/172 and their targets) with leaf morphology (CUP‐SHAPED COTYLEDON (CUC)‐induced serrations) through a mediator (TCPs, TEOSINTE BRANCHED1/CYCLOIDEA/PROLIFERATING CELL FACTORs) (Rubio‐Somoza et al., 2014).
If distinct molecular pathways affect different traits within the infinite features defining the architecture of a leaf, the outline and venation topology of leaves should theoretically be decomposable into latent (hidden) shapes; that is, multivariate morphological signatures that vary independently from each other (Chitwood & Topp, 2015). For example, shape differences defining species, as well as shape differences defining developmental stage, may be detectable within each leaf and subsequently isolated from each other. Shape differences that define species, regardless of developmental context, may vary distinctly from those that define developmental context, regardless of species. These shapes are latent because, although they are present in each leaf, they manifest within the context of other factors that affect shape as well. Latent shapes globally affect leaf shape in different ways, and each of the features comprising the latent shapes alone do not necessarily discriminate the effects of genetics or development. From this perspective, the shape of a given leaf – from any species, from any time point during development or any position in the plant – would result from the confluence of latent shapes regulated by these processes. The single organ that we call a leaf would actually be a composite of latent features that vary by genetic vs developmental effects, independently from each other.
Grapevine (Vitis spp.) leaves exhibit a breathtaking range of variation in leaf shape, making this genus ideally suited to explore latent shapes resulting from evolutionary and developmental processes (Fig. 1). Taxonomists studying Vitis have used variation in leaf lobing and leaf margins to delimit the nearly 60 species in the genus (Moore, 1991; Ren & Wen, 2007). Leaf shape is also important in the assessment of the intraspecific variation in the European grapevine (Vitis vinifera ssp. vinifera), which is grown around the world for wine‐making and table grapes. Unique among crops, variation in grape leaf shape (together with other vine features) is used by viticulturists to quantitatively classify grape varieties, a field known as ampelography (αμπελος, ‘vine’ and γραφος, ‘writing’) (Galet, 1952). Grape leaves have several homologous points amenable to landmark‐based analyses (Galet, 1979; Chitwood et al., 2014), increasing the biological interpretation of morphometric data compared with species with stochastic venation topologies or limited homology (such as Arabidopsis, tomato, Antirrhinum, etc.). Like Hales’ grid of pins (1727), naturally homologous points in grape leaves allow developmental stage and leaf number effects to be quantitatively tracked, potentially revealing separable latent processes contributing to the composite morphology known as a leaf.
Figure 1.
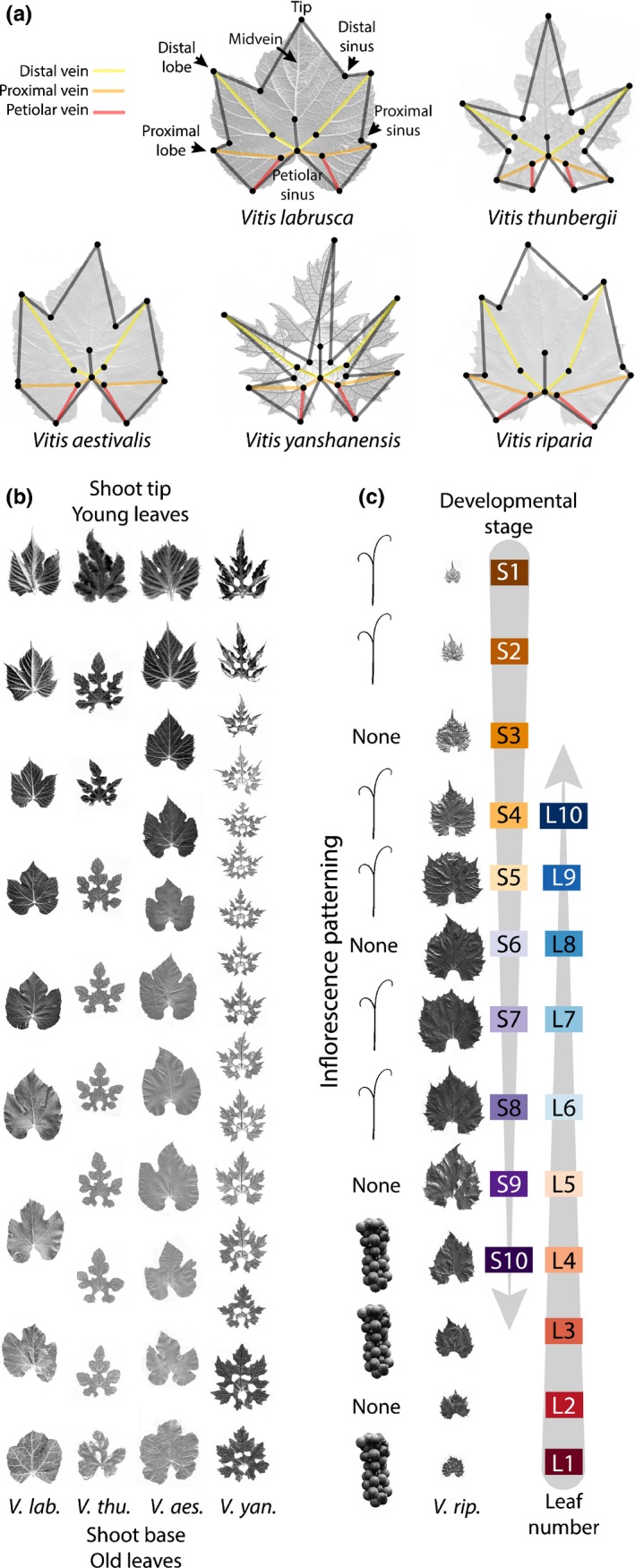
Morphological features defining the temporal development of Vitis leaves. (a) All Vitis leaves possess distal (yellow), proximal (orange) and petiolar (red) veins, as well as distal and proximal lobes and sinuses, and a petiolar sinus. Seventeen homologous landmarks (black circles) were used in this study. Morphologically diverse species are shown. (b) Leaf series, scaled by the length between distal lobe tips, showing developmental stage and leaf number shape variance. (c) Unscaled V. riparia leaves showing changes in size at the shoot tip and base. Developmental stage (Sn) is measured starting from the shoot tip and leaf number (Ln) is measured from the shoot base. Inflorescences, opposite leaves, skipping every third node, transform from clusters into tendrils from the shoot base to tip.
Materials and Methods
Germplasm, sample collection and scanning
Over 270 vines in the US Department of Agriculture (USDA) Vitis germplasm collection in Geneva, NY, USA were sampled in June 2013. Using an automatic label maker, vine identification number and the identity of organs (or lack thereof) at each node, beginning with the first sampled leaf at the tip, were recorded in the field. Beginning with the first leaf at the tip that could be flattened and scanned (c. 1 cm in length), leaves were collected in order, shoot tip to base, as a stack, and placed into a Ziploc bag to which the label was affixed. A single shoot was collected per vine, and bags were placed into a cooler until scanning. A description of the number of vines (genotypes) collected for each species and hybrid, as well as the number of leaves representing different developmental stages (Sn) and leaf numbers (Ln) can be found in Supporting Information Fig. S1.
Leaves were arranged on a scanner (Mustek A3 1200S; Mustek Systems, Hsinchu, Taiwan) in the collected order from the shoot, and next to each leaf was placed a small label indicating nodes (as measured by developmental stage) and organ identity opposite the node (C, cluster; T, tendril; N, no organ). The abaxial side of the leaves was imaged. The file name of the image indicates the vine ID, and the appended letter indicates which image in the series the file represents for each vine. The raw scans are publically available at the following link: https://dataverse.harvard.edu/dataverse/vitis_leaves.
Landmarking
For both wild Vitis species and reanalyzed V. vinifera ssp. vinifera data, 17 landmarks were placed, in order, for each leaf using the ImageJ (Abramoff et al., 2004) point tool. Landmarks and their order were as follows: (1) petiolar junction, (2) midvein tip, (3) left distal sinus, (4) right distal sinus, (5) left distal lobe tip, (6) right distal lobe tip, (7) left proximal sinus, (8) right proximal sinus, (9) left proximal lobe tip, (10) right proximal lobe tip, (11) left terminus petiolar vein, (12) right terminus petiolar vein, (13) branch point midvein, (14) branch point left distal vein, (15) branch point right distal vein, (16) branch point left proximal vein, (17) branch point right proximal vein. Using ggplot2 (Wickham, 2009) in R (R Core Team, 2014), graphs for landmarks from each image were visually checked for errors. If errors were detected, the landmarking was redone for those particular samples.
Morphometric analysis and visualization
Once a quality landmarked dataset had been created, a generalized Procrustes analysis (GPA) was undertaken using the R package shapes (Dryden, 2013). For 17 landmarks in two dimensions (x, y coordinates) for 3292 leaves of wild Vitis species and 9548 V. vinifera ssp. vinifera leaves, GPA was performed using the procGPA function, reflect = TRUE. Separate Procrustes analyses were performed for all wild Vitis species’ leaves at all shoot positions, and for V. vinifera ssp. vinifera and ssp. sylvestris leaves and wild Vitis species’ leaves selected for the corresponding shoot position to the domesticated grape dataset. Eigenleaves were visualized using the shapepca function and principal component (PC) scores, percentage variance explained by each PC and Procrustes‐adjusted coordinates were obtained from procGPA object values. To test for correlation between PCs and developmental stage or leaf number, Spearman's rho was calculated, whereas, for variability of PC values across species, a Kruskal–Wallis test was used.
Linear discriminant analysis (LDA) on Procrustes‐adjusted coordinates was performed using the lda function from the MASS package (Venables & Ripley, 2002). Species, developmental stage and leaf number were all analyzed independent of each other. The predict function (statistics package) and table function (base package) were employed (dependent on MASS) to reallocate leaves (whether by species, developmental stage or leaf number) using the linear discriminants. When predicting V. vinifera ssp. vinifera developmental stage and leaf number, the wild Vitis species data were used as a training set to predict values for domesticated grape leaves. For the wild Vitis species’ data, for each leaf, there are actual vs apparent species, developmental stage and leaf number identities. Relative developmental stage and relative leaf number are calculated as the apparent value – actual value. The mean relative developmental stage and relative leaf number values for each vine were employed to determine the significant deviation of the relative values of species from zero using a one‐sample, two‐tailed t‐test. Correlation between relative values and first tendril node was analyzed using Spearman's rho.
As described previously (Chitwood et al., 2014), Germplasm Resources Information Network (GRIN) trait values, averaged on a per accession basis, for V. vinifera ssp. vinifera vines were correlated with each other and with morphometric and predicted temporal data using the rcorr function from Hmisc (Harrell, 2013) employing Spearman's rho and a false discovery rate controlled using the Benjamini–Hochberg procedure (Benjamini & Hochberg, 1995). Hierarchical clustering for V. vinifera ssp. vinifera traits, and for averaged Procrustes‐adjusted coordinates for species throughout the genus Vitis, was carried out using the hclust function on a distance matrix calculated from correlation performed on complete, pairwise observations and visualized using the as.phylo function from the package ape (Paradis et al., 2004).
Visualization was performed in ggplot2 (Wickham, 2009) using geom_bar, geom_boxplot, geom_point, geom_segment, geom_tile and stat_smooth functions, among others, and color schemes derived from http://colorbrewer2.org.
Results
Developmental stage vs leaf number
To determine the effects of developmental stage, leaf number and evolutionary lineage (taxonomic identity) on leaf shape, we scanned > 3200 leaves from wild relatives of domesticated grape held by the USDA germplasm repository in Geneva, NY, USA. Like many living collections of perennial crops, the Geneva repository houses multiple genotypes of different Vitis species in common conditions. This extensive collection includes wild‐collected accessions of at least 19 North American and Asian Vitis species, as well as assorted V. vinifera hybrids. From the Geneva repository, we sampled > 270 vines representing 12 Vitis species, four V. vinifera hybrids and three species from the related genus Ampelopsis. Each sampled vine represents a unique genetic accession. The number of vines sampled for each species varied widely (Fig. S1), but, in this study we focused on 10 species for which five or more accessions were sampled: Vitis riparia (71 vines), V. labrusca (40 vines), V. cinerea (37 vines), V. rupestris (28 vines), V. acerifolia (16 vines), V. amurensis (16 vines), V. vulpina (13 vines), V. aestivalis (eight vines), V. coignetiae (five vines) and Vitis palmata (five vines). All Vitis leaves possess a midvein, distal and proximal veins, a petiolar vein, as well as proximal and distal lobes and sinuses, and wide variation in the width of the petiolar sinus (Fig. 1a). We leverage these homologous points and others to measure 17 landmarks in all leaves (Table S1). Later, we compare the leaves from the wild relatives of grape described earlier with previously published data on 1200 varieties of domesticated grape (Chitwood et al., 2014), which are described in subsequent sections.
For each vine accession, a representative shoot was selected and the shoot position of each leaf was recorded (Fig. 1b,c). Developmental stage was measured by counting from the youngest, first measureable leaf at the shoot tip. The time between successively initiated leaves is a plastochron, and the youngest initiated leaf primordium at the shoot apical meristem is denoted P1 (plastochron 1) to indicate this. However, because we begin not with P1 (which is micrometers in size), but with the first measureable leaf (c. 1 cm in size), we use S1 (for ‘stage’) to denote the youngest measured leaf at the shoot tip, counting numerically upwards (S2…Sn) towards the shoot base. Contrastingly, leaf number begins with the first initiated leaf (L1) found at the shoot base and counts numerically upwards (L2…Ln) towards the shoot tip (Fig. 1c). These two metrics are used to differentiate the effects of Sn from Ln (Fig. 1c).
The effects of developmental stage are expected to be strongest in young leaves at the shoot tip (with low S numbers and high L numbers; Fig. 1c), as their shape is in flux during their expansion. Older leaves at the shoot base (with low L numbers and high S numbers) are more strongly influenced by leaf number, because they reflect changes in the mature leaf shape at successive nodes. The vast majority of the vines sampled here possess leaves corresponding to S1–S10 and L1–L10 (86% represent developmental stages up to S10 and 88% represent leaf numbers up to L10, Fig. S1), and we restrict our analyses to these positions (Fig. 1c).
It is important to note that, because Sn and Ln are counted from opposite ends of the same shoots, it is anticipated that effects associated with each will generally be inversely related. Indeed, as described subsequently, this is often the case. However, the effects of each do not perfectly mirror each other, allowing the partial discernment of Sn‐ and Ln‐specific effects. The reason for this is that the total number of leaves per shoot varies, such that S1 does not always correspond to the same leaf number between shoots (Fig. S2a) and, likewise, L1 does not always correspond to the same developmental stage between shoots (Fig. S2b). Importantly, the total number of leaves per shoot is relatively constant between species (Fig. S1c), so that the overall distribution of Sn and Ln between species is not confounded by species identity. Intraspecies variability in the number of total leaves per shoot partially resolves the confounding between developmental stage and leaf number, allowing for insights into the trends affecting leaf ontogeny and heteroblastic shape progression. Short of tracking the developmental progression of each leaf in the shoot of each of hundreds of vines (something that is impossible at this time), the complementary indexing using Sn and Ln allows distinct ontogenetic and heteroblastic trends in leaf shape to be described.
Morphospace of leaves in the genus Vitis
Landmarks were aligned (accounting for translation, rotation and scaling) using a GPA. A principal component analysis (PCA) was then performed to visualize the major sources of shape variance among leaves, including all species and node positions. The first four PCs explain 73.2% of shape variance among leaves from Vitis species, developmental stages and leaf numbers (Fig. 2). More than one‐half of the shape variance, represented by PC1 and PC2, is influenced by lobing and the width of the petiolar sinus. Low PC1 and high PC2 values readily distinguish highly dissected Vitis species and some members of the related genus Ampelopsis (Fig. 2b). A subset of species with diverse leaf shapes is projected onto the overall morphospace in Fig. 2(b) for clarity, and to better discern the shape effects by genotype of different PCs (note: all node positions are included in this visualization). In Fig. S3, we project leaves found throughout all nodes in 10 Vitis spp. with a replication of five vines or more (Fig. S1b; V. amurensis, V. coignetiae, V. palmata, V. labrusca, V. aestivalis, V. vulpina, V. cinerea, V. rupestris, V. riparia and V. acerifolia). These 10 species exhibit unique shape differences, but are more closely related in shape than the extremely lobed species (A. brevipedunculata, A. acontifolia, V. thunbergii and V. piasezkii) depicted in Fig. 2(b).
Figure 2.
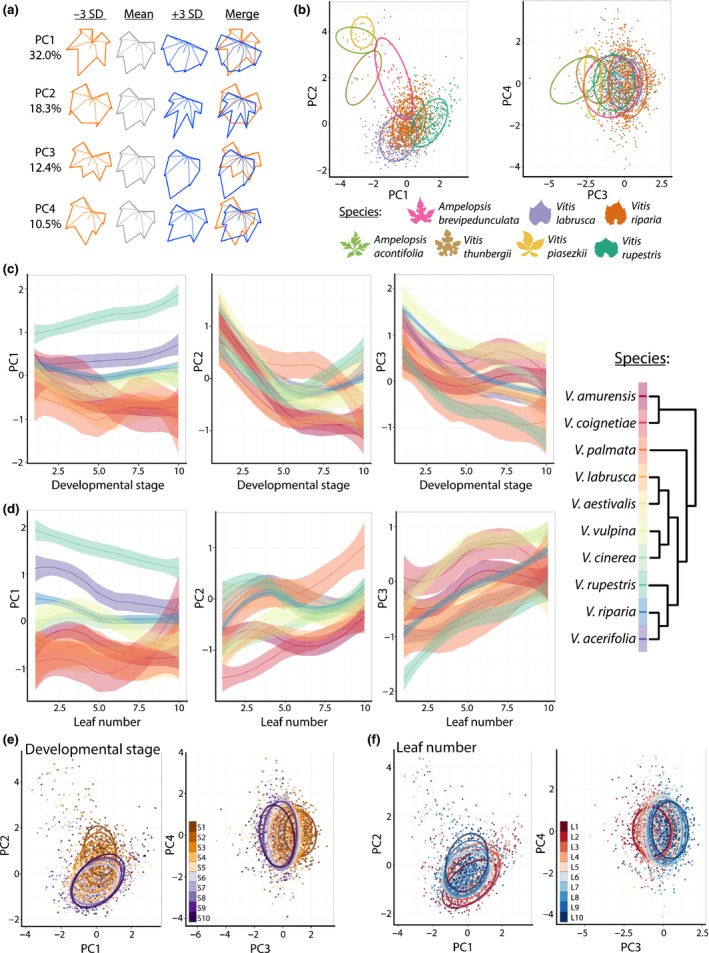
Morphospace of leaves in the genus Vitis. (a) ‘Eigenleaves’ showing leaf morphs represented by principal components (PCs) at ± 3SD and shape variance explained by each. The principal component analysis (PCA) morphospace is calculated for all species and shoot positions. (b) Select species (indicated by color) representing morphological diversity in the dataset projected onto the morphospace. Projected data for each species include all shoot positions. Confidence ellipses (95%) are drawn. Other species are projected onto the morphospace in Supporting Information Fig. S3. (c, d) Locally weighted scatterplot smoothing (LOWESS) showing the relationship between PCs 1–3 and (c) developmental stage and (d) leaf number. Phylogenetic relationships between species are indicated by color. Plots of developmental stage vs leaf number are found in Fig. S4. (e) Developmental stage and (f) leaf number projected onto the morphospace representing leaves from all species.
In order to examine the variability of developmental stage and leaf number in morphospace, we visualize each PC as a locally weighted scatterplot smoothing (LOWESS) curve plotted against stage and leaf number for the 10 Vitis spp. for which five or more vines were collected (Fig. S1b), allowing us to statistically estimate developmental shape effects by species. Developmental stage and leaf number vary less by PC1 than by PC2 and PC3 (i.e. PC1 values are relatively developmentally invariant compared with PC2 and PC3 values) (Fig. 2c,d). PC2 and PC3 values are largely indistinguishable between species across developmental stages (Fig. 2c) and leaf number (Fig. 2d), suggesting strongly conserved morphological features in developing leaves across species for these shape attributes, particularly for early developmental stages (Fig. 2c). Because PCs are orthogonal (i.e. uncorrelated), the fact that species, developmental stage and leaf number correlate differentially with PCs is suggestive that each might be represented by independent shape attributes present within leaves. Revisualization of the morphospace by developmental stage (Fig. 2e) and leaf number (Fig. 2f) demonstrates that these factors mostly vary by PC2 and PC3, whereas species shape differences traverse along morphospace paths defined by PC1, PC2 and PC3 (Fig. 2b). Although the trajectories of shape changes in each species across Sn and Ln are generally inversely related, this is not completely the case. In Fig. S4, Sn and Ln are plotted against each other in the same plot, demonstrating their partial independence from each other.
To help to qualitatively understand the different ways in which grape leaves differ among species and developmental contexts, we compared average shapes (Fig. 3). Related members of Moore's Series Ripariae (V. acerifolia, V. riparia and V. rupestris) (Moore, 1991; Miller et al., 2013) are defined by a shallow petiolar sinus, especially V. rupestris (Fig. 3a). Outside of the Ripariae, V. vulpina and V. cinerea also exhibit shallow petiolar sinuses, but to a lesser extent, whereas the petiolar sinus of the remaining species (V. aestivalis, V. labrusca, V. palmata, V. coignetiae, V. amurensis) is more acute. Vitis palmata exhibits especially deep distal lobing relative to other species. Each species’ leaves also vary by developmental stage (Fig. 3b) and leaf number (Fig. 3c), usually by the length of the leaf tip and the shallowness of the petiolar sinus. In the next section, we determine the extent to which shape attributes varying by species, developmental stage and leaf number are separable from each other.
Figure 3.
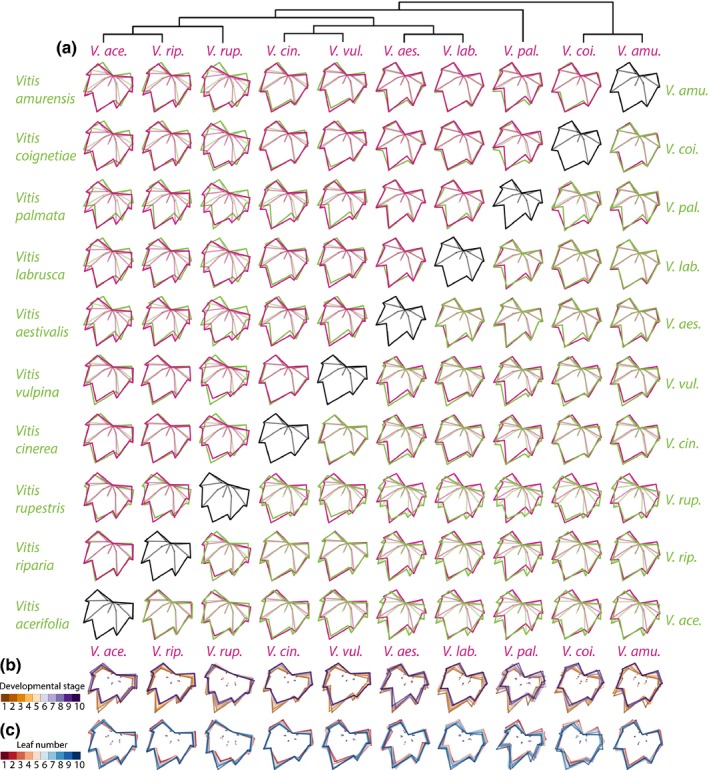
Shapes represented among Vitis species, developmental stage and leaf number. (a) Outlines representing the average shape of each pairwise comparison of species, indicated by magenta and green. Phylogenetic relationships are indicated. (b, c) Comparison of average shapes across (b) developmental stages (S1, dark orange to S10, dark purple) and (c) leaf number (L1, dark red to L10, dark blue) for each species indicated by color.
Latent shapes independently predict species, developmental stage and leaf number
We employed an LDA to maximize the separation of leaf attributes (whether species, developmental stage or leaf number) from each other using all measured shape information. The resulting linear discriminants can be employed to predict the apparent class of a leaf and are useful for comparing actual vs apparent leaf identities as confusion matrices (Fig. 4).
Figure 4.
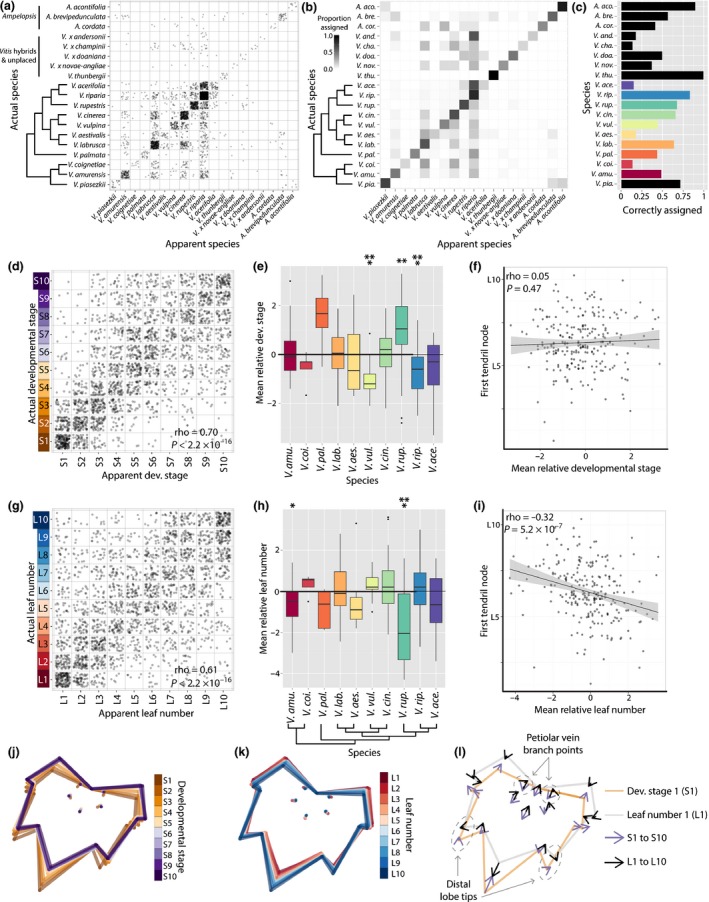
Species, developmental stage and leaf number can be predicted independently of each other. (a) Confusion matrix resulting from the prediction of species identity using a linear discriminant analysis (LDA). The panel should be read from left to right: for each species indicated on the left, leaves appeared to be derived from the apparent species indicated on the bottom. Developmental stage and leaf number were not considered in the prediction. (b) Same as (a), except the apparent species assigned to each actual species has been converted into a proportion. Proportion totals equal one adding from left to right. (c) The proportion of correctly assigned leaves for each species. (d) Similar to (a), actual developmental stage (left) and apparent developmental stage (bottom), predicted without species or leaf number information. (e) Relative developmental stage values (apparent – actual developmental stage) averaged for vines from each species. Significant deviations from zero are indicated: *, P < 0.05; **, P < 0.01; ***, P < 0.001. Median and first and third quartiles represented by the box; whiskers extend to the most extreme value within 1.5 times the interquartile range. (f) Correlation between mean relative developmental stage of vines and first tendril node. The fitted linear model is drawn. (g–i) Same as (d–f), but for leaf number. Note the significant correlation between the first tendril node and relative leaf number (i), not seen for developmental stage (f). (j, k) Latent shape attributes defining developmental stage (j) and leaf number (k) indicated with average leaf shapes. (l) Comparison of changes in latent shape attributes across developmental stage (orange outline, S1; purple arrow, S1 to S10) and leaf number (gray outline, L1; black arrow, L1 to L10). Note the non‐inverse relationship between developmental stage and leaf number changes in the distal lobe tips and petiolar vein branch points (indicated).
Linear discriminants separating species, without regard to developmental stage or leaf number, can be used to predict the identity of a species (Fig. 4a–c). For most taxa, the largest proportion of predicted leaves corresponds to the taxonomic identity assigned to that species (Fig. 4b; the diagonal indicates the proportion of correctly assigned leaves). Incorrectly assigned leaves are most often confused with those of other species that are phylogenetically related to the assigned species. For example, the majority of V. acerifolia leaves are confused with those of its close relative V. riparia; similarly, the largest group of V. aestivalis leaves are confused with those of its relative V. labrusca. By contrast, the Asian V. coignetiae leaves are confused with those of more distant relatives native to North America, including V. labrusca and V. cinerea, and sometimes even V. riparia, perhaps indicating convergent evolution, germplasm misidentification or segregating leaf shape differences among these accessions. Interestingly, V. × andersonii, described as a hybrid of V. coignetiae and either V. vulpina or V. riparia, is strongly mistaken for V. riparia, providing circumstantial evidence of parentage. As has been suggested by ampelographers and taxonomists (Galet, 1952; Moore, 1991), our results demonstrate that leaf shape is often sufficient to identify species, even in the absence of developmental information. Instances in which leaves of one species are assigned to another offer a valuable opportunity to develop hypotheses about phylogenetic history, hybridization and convergent evolution that can be tested with additional evolutionary and ecological analyses.
Reciprocally, we wondered whether developmental stage and leaf number, regardless of species identity, might be similarly informative about developmental context. Indeed, linear discriminants trained on developmental stage or leaf number can predict the developmental context of a leaf for either measure (Fig. 4d–i). Developmental stage can be predicted with a Spearman's correlation coefficient of rho = 0.70 (Fig. 4d), and leaf number with rho = 0.61 (Fig. 4g). The prediction of both developmental stage and leaf number is more accurate at the beginning of their respective series, where the effects of each are anticipated to be the strongest (Fig. 1c). Based on these observations, we conclude that developmental context, measured by either developmental stage or leaf number, can be predicted independently of genotypic information. These results suggest that the allometric changes in leaf shape during ontogeny and the changes in shape as a result of heteroblastic development are broadly conserved across the genus Vitis.
The ability to predict developmental context separate from species identity provides a method to quantify differences between species attributable to changes in developmental timing, also known as heterochrony. For each leaf, we calculated the relative developmental stage and relative leaf number as the apparent value minus the actual value (e.g. an S5 leaf predicted to be S7 would have a relative stage value of +S2, and an L4 leaf predicted to be L2 would have a relative value of −L2). Relative values indicate, for a given leaf, how many nodes ahead or behind (developmentally speaking) that leaf appears to be. Averaging the relative developmental stage and leaf number values for each vine, we can detect species that are precociously ahead or lagging behind the expected developmental stage (Fig. 4e) and leaf number values (Fig. 4h). Relative developmental stage and relative leaf number are inversely related to each other (compare Fig. 4e with 4h), but not completely so, consistent with the Sn and Ln numbers showing inversely related but unique trajectories through morphospace (Fig. S4). Relative developmental stage and leaf number provide contrasting insights into the temporal development of leaves. For example, the mean relative developmental stage of V. rupestris vines is c. +S1 ahead of their actual developmental stage (Fig. 4e) and c. –L2 behind their actual leaf number (Fig. 4h). This accelerated development (Sn) and protracted display of juvenile leaf types typically found at the base of the shoot (Ln) contribute to the unique shape of V. rupestris leaves compared with other Vitis spp. (Fig. 3). Vitis rupestris leaves (Fig. 3a), older leaves (high Sn values, Fig. 3b) and juvenile leaves (low Ln values, Fig. 3c) all share a characteristic wideness conferred by a diminished leaf tip.
Within Vitis, the identity of inflorescences, which appear opposite leaves, transform from clusters at the shoot base to tendrils at the shoot tip (Srinivasan & Mullins, 1981; Gerrath, 1988, 1993; Boss & Thomas, 2002) (Fig. 1c). Both changes in leaf shape at successive nodes and the transformation of inflorescence identity indicate temporal changes in the development of the shoot apical meristem, known as heteroblasty. This transition is not the same as flowering time, as both clusters and tendrils are inflorescences, and, moreover, these organ primordia are patterned the previous year (Carmona et al., 2008). Therefore, the cluster to tendril transformation serves as a discrete, binary indication of the heteroblastic transition, that is, the temporal development of the meristem. If heteroblasty regulates both the latent shapes predicting Ln and the cluster to tendril transition, we would assume that they would be correlated. Further, such a correlation should not be observed between Sn and the cluster to tendril transition, as leaf ontogeny (developmental stage) is not related to heteroblasty (cluster to tendril transition).
Indeed, there is a significant negative correlation between relative leaf number and the first node in which a tendril is observed: that is, in vines with precocious, adult leaves (i.e. higher relative leaf numbers, typical of leaves found closer to the shoot tip), tendrils appear at earlier nodes closer to the base of the shoot, linking two different measures of premature heteroblastic change (Fig. 4i). Moreover, this correlation is not observed for relative developmental stage (Fig. 4f). Taken together, these results indicate that latent shapes predictive of developmental stage and leaf number are functionally distinct, reflecting the developmental progression of leaves (Sn) and their heteroblastic transitions across successive nodes of the shoot (Ln) separately.
Average leaf shapes indicate that developmental stage and leaf number vary by the prominence of the leaf tip and the shallowness of the petiolar sinus (Fig. 4j–l). The shape changes for these two factors are inversely related, but obviously distinguishable from each other, given their differential correlation with tendril position, that is, heteroblasty (Fig. 4f,i). On closer inspection, the distal lobe tips and the branch point of the petiolar vein distinguish shape changes attributable to developmental stage and leaf number (Fig. 4l), isolating the shape attributes unique to these functionally distinct processes.
Morphological differences between domesticated grape and wild relatives
Previously, we measured > 9500 leaves from V. vinifera ssp. vinifera (domesticated grape) and its wild progenitor V. vinifera ssp. sylvestris. These leaves, collected from the USDA germplasm repository in Winters, CA, USA, represent over 2300 vines and 1200 varieties (Chitwood et al., 2014). These leaves were sampled to measure purely genetic effects and to minimize the influence of development by collecting four successive leaves from the midpoint of the shoot. That species identity can be predicted independently from developmental context in wild Vitis spp. (Fig. 4a–c) with disparate leaf shapes lends credence to the assumption that developmental context can be ignored, which is almost always invoked when mapping traits for quantitative genetic purposes or taxonomically defining species.
We wanted to compare leaves from domesticated grape and its wild progenitor with those from the other Vitis species described earlier. To do so, we restricted our analysis in silico to leaves at the same shoot positions as those from which we had originally collected leaves in domesticated grape (the four leaves closest to the midpoint of the shoot). In the combined morphospace, PC1 and PC2 describe nearly 60% of shape variance (Fig. 5). Similar to the wild Vitis species‐only morphospace (Fig. 2a), lobing and the petiolar sinus define the first PCs. Petiolar veins in domesticated grape varieties can be angled so that they cross each other and, because of this, the closure of the petiolar sinus represented by high PC1 values is particularly strong (Fig. 5a). Leaf dissection defined by high PC1 and high PC2 values clearly delineates less lobed wild Vitis species from more acutely lobed domesticated grape varieties (Fig. 5b). Highly dissected species, such as Ampelopsis acontifolia, V. piasezkii, V. thunbergii and V. vinifera var. Ciotat, form a distinct morphological grouping.
Figure 5.
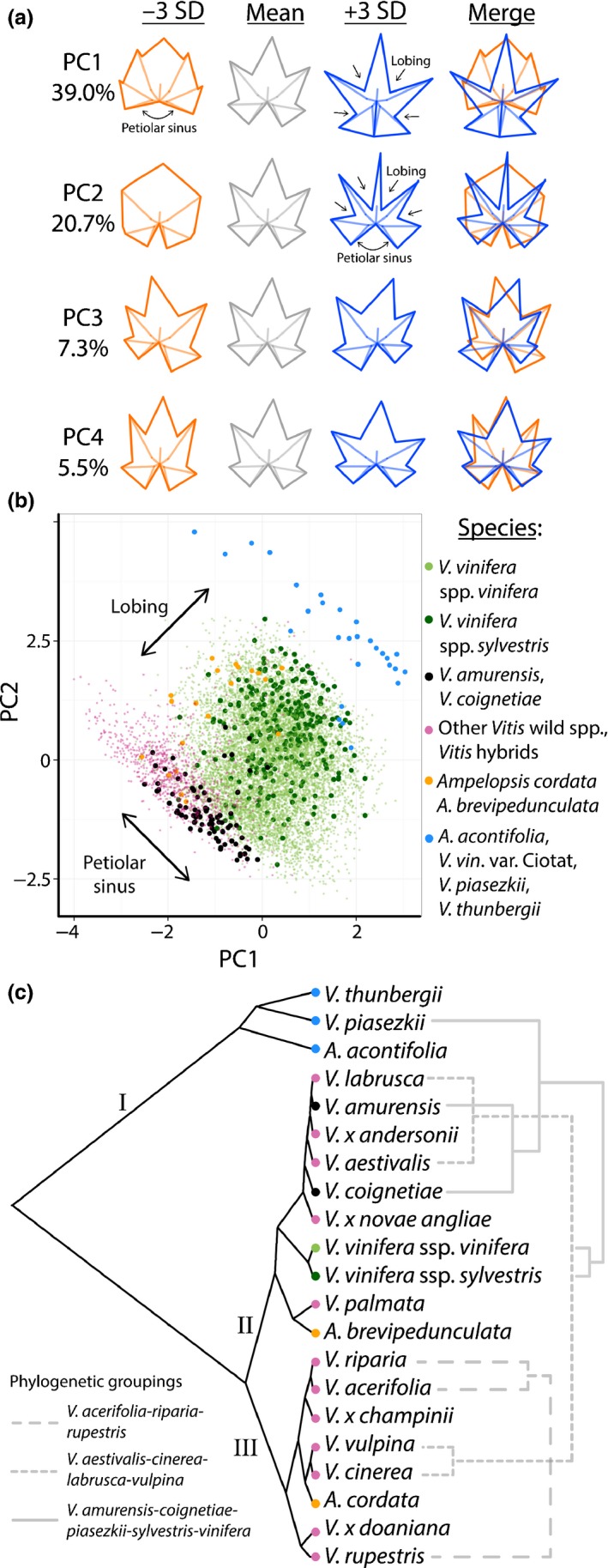
Morphospace of wild Vitis species and domesticated grape. (a) ‘Eigenleaves’ showing leaf morphs represented by principal components (PCs) at ± 3SD and shape variance explained by each. Lobing and petiolar sinus variation are indicated. (b) Comparison of wild Vitis species leaf shape (red, black) with V. vinifera ssp. vinifera and V. vinifera ssp. sylvestris (green, dark green, respectively) and highly lobed species (blue). Strong shape variation in lobing and petiolar sinus depth indicated. (c) Hierarchical clustering based on morphology. Phylogenetic groupings indicated in light gray. Groups mentioned in the Results section indicated with Roman numerals.
Although there are clear patterns in leaf shape related to development and species identity, the correspondence between the morphospace of wild and domesticated Vitis species and the evolutionary relationships among Vitis species are complex. Some, but not all, well‐known phylogenetic relationships are reflected in leaf morphology (Fig. 5c) (Zecca et al., 2012; Miller et al., 2013). Notably, clustering the averaged shapes from different genotypes reveals that V. vinifera ssp. vinifera is morphologically sister to V. vinifera ssp. sylvestris, the wild progenitor of domesticated grape (Fig. 5c, Group II) (Myles et al., 2011; Zecca et al., 2012). Also within Group II are two Asian species, V. amurensis and V. coignetiae, which, together with V. vinifera ssp. vinifera and V. vinifera ssp. sylvestris, represent members of a Eurasian clade. Within Group III are found members of Series Ripariae (V. acerifolia, V. riparia, V. rupestris). The leaf shapes of Vitis hybrids cluster on the basis of parental lineages, with V. × champinii and V. × doaniana clustering in Group III, presumably because of their V. rupestris and V. acerifolia heritages, respectively, and V. × andersonii and V. × novae‐angliae clustering closer to their V. coignetiae and V. labrusca parents, respectively. These observations suggest that similarities in leaf shape may reflect recent evolutionary events, domestication or contemporary interspecific gene flow.
At broader levels of phylogenetic scale, however, patterns of leaf shape similarity do not appear to correspond with known evolutionary relationships. For example, together with V. vinifera ssp. vinifera, V. vinifera ssp. sylvestris and two Asian species found in Group II (discussed earlier), there are other North American species (V. labrusca, V. palmata), several hybrids and Ampelopsis brevipedunculata, taxa that are not known to be closely related to one another. A previously identified clade based on molecular data (V. aestivalis, V. cinerea, V. labrusca and V. vulpina) is divided here between Groups II and III. Although Vitis is well known to be a monophyletic genus (Wen et al., 2007, 2013), on the basis of leaf shape, three species of Ampelopsis cluster in three different groups, together with Vitis species. These data suggest that, although leaf shape may bear signatures of recent evolutionary events, leaf shape does not appear to track with phylogeny at larger scales; distantly related species, even from distinct genera, resemble each other in leaf shape through evolutionary convergence.
Changes in developmental timing underlie morphological diversity in domesticated grape leaves
Although leaves from equivalent positions in the shoot clearly separate species and varieties by genetic effects (Fig. 5), such effects may still be developmental in nature, an example of heterochrony. If changes in the relative timing of Sn or Ln have occurred, they would contribute to morphological differences between species, as described earlier for wild Vitis spp. (Fig. 4e,h). The conserved latent shapes defining developmental stage and leaf number in morphologically disparate Vitis species can be used predictively to detect such changes in timing.
We used wild species’ leaves, from S1 to S10 and L1 to L10 (where developmental stage and leaf number effects, respectively, are strongest), as a training set to predict the Sn and Ln values of domesticated grape leaves (Fig. 6). A caveat of this approach is that the two datasets are collected from different environments and years. When using the Geneva, NY, USA, 2013 dataset to predict developmental timing in the Winters, CA, USA, 2011 dataset, we apply the assumption that environment affects leaf morphology independently from developmental timing. In addition, the wild species training set is influenced by strong morphological trends at the tip and base of the shoot, whereas the domesticated dataset is collected from the middle of the shoot, where these effects are weaker. Nonetheless, developmental timing can be predicted independently from genotype in the middle of the shoot as well, even though the effects are weaker than at the ends of the shoot (see Fig. 4d,g).
Figure 6.
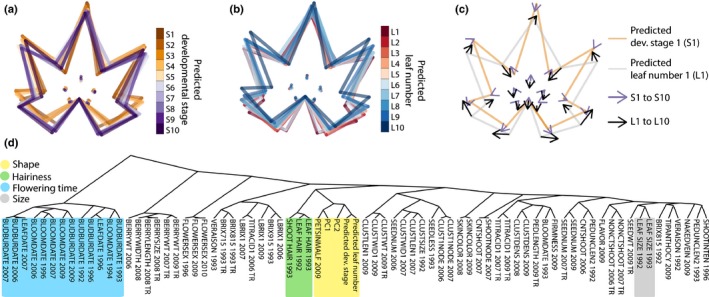
Evolution in developmental timing underlies leaf shape diversity in domesticated grape. (a, b) Latent shape attributes defining developmental stage (a) and leaf number (b) in domesticated grape indicated with average leaf shapes. (c) Comparison of changes in latent shape attributes across predicted developmental stage (orange outline, S1; purple arrow, S1 to S10) and predicted leaf number (gray outline, L1; black arrow, L1 to L10). (d) Hierarchical clustering of measured traits in grape. Morphological and predicted temporal traits (yellow), hirsuteness (green), reproductive transitions (blue) and overall leaf size (gray) are indicated.
When comparing leaf averages across predicted developmental stages (Fig. 6a) and leaf numbers (Fig. 6b), it is apparent that variation in petiolar sinus depth is the major predictor of developmental timing in domesticated grape leaves (Fig. 6c). Variation in the petiolar sinus is a recurrent theme in morphology associated with both developmental stage and leaf number (Figs 3b,c and 4j–l).
Previously, numerous other traits, from the timing of bud burst and flowering to berry ripening and titratable acids and sugar content, have been measured on the domesticated grapevines described above (Chitwood et al., 2014). We were curious about the correlational context of overall leaf shape (represented by PCs) and developmental timing (predicted Sn and Ln) to these other traits. The predicted Sn and Ln of domesticated grape leaves are significantly correlated with PC1 and PC2 (Table S2; Fig. 6d), which together explain near 60% of the total shape variance for measured Vitis genotypes (Fig. 5a). From this, we conclude that latent shape attributes modulating developmental stage and leaf number potentially explain large amounts of shape variance in domesticated grape, indicating that changes in developmental timing (heterochrony) may underlie the morphological evolution of domesticated grape varieties. In addition, one trait, ‘PETSINMALF 2009’, is a qualitative, 0–8 rating of petiolar sinus depth, and is tightly correlated with PC1, PC2, and predicted developmental stage and leaf number, confirming that our quantitative measures reflect intuitive perceptions of leaf shape (Fig. 6d; Table S2).
Leaf morphology traits, including predicted developmental timing, are largely separate from bud burst and bloom dates (‘BUDBURST DATE’, ‘BLOOMDATE’, ‘LEAFDATE’), indicating that these timing features are separate from the overall transition to reproductive development in grapevine (Fig. 6d). Further, leaf shape is distinct from the overall size of leaves (‘LEAF SIZE’), demonstrating that grape leaf shape is not merely a developmental constraint correlated with overall size (i.e. allometry) (Fig. 6d). One trait significantly correlated and clustering with leaf shape is hirsuteness, in both the leaves and shoot (‘LEAF HAIR’, ‘SHOOT HAIR’), which we observed previously in domesticated grape (Chitwood et al., 2014). Leaf hirsuteness varies by both developmental stage, as trichomes become less dense as the leaf expands, and by leaf number. It is interesting to imagine how thermoregulation, disease resistance or other trichome‐mediated functionality may have varied as a developmental constraint as developmental timing has changed over the evolution and domestication of grapevines.
Discussion
Many descriptions of complex traits, from faces to voices, and even the evolution of cultural products, such as violins (Zhang et al., 1997; Kuhn et al., 2000; Chitwood, 2014; Claes et al., 2014),, potentially stand to benefit from the isolation of features uniquely regulated by distinct pathways. Like a face, leaf shape is modulated in complicated ways by genetics, development and the environment. The latent shapes discussed here, describing developmental stage and leaf number, embedded within the overall shape of a leaf, and conserved across the genus Vitis, fit the definition of a ‘cryptotype’. Recently, we have suggested the term ‘cryptotype’, borrowed from linguistics (which uses both ‘phenotype’ and ‘cryptotype’ parallel to the biological definitions) (Whorf, 1945), to describe latent, combinatorial features (e.g. 17 homologous landmarks) within a shared multivariate space that vary independently from each other (Chitwood & Topp, 2015). Cryptotypes are not unreal or even abstract; they are simply a set of features, that, when combined, best discriminate one biological process (e.g. evolutionary history) from others (e.g. developmental stage). The shapes that arise through this combination of features are latent only because they are not immediately recognizable when each of their component landmarks is considered alone; the effects of genetics and development affect the entirety of the leaf. The multivariate cryptotypes of genotype, developmental stage and leaf number stand in contrast with the traditional univariate phenotypes, such as height, biomass, or leaf length, width or area.
Defining such combinative, latent shapes is critical to an understanding of the mechanisms by which morphological diversity manifests in plants. The unequal expa nsion of leaves (allometry) and the different types of leaves displayed at nodes (resulting from heteroblasty) have been studied for centuries (Hales, 1727; Goethe, 1817, 1952; Goebel, 1900), but the inability to distinguish the effects at a morphological level, within the shared morphology of single leaves in which these processes act, has led to confusion. Early hypotheses that shade prolonged juvenility through a heteroblastic mechanism were later refuted by careful study of the morphology of leaf primordia (Jones, 1995). Similarly, the degree to which changes in the development of leaves or the heteroblastic series influences evolutionary changes – and plasticity – remains an open question.
Recent studies have uncovered a transcriptomic basis underlying leaf ontogeny (Efroni et al., 2008). Similarly, dramatic transitions to reproductive fates, reflecting the temporal development of meristems, have been described (Park et al., 2012). An understanding of molecular clocks, regulating phase change through small RNAs (Chuck et al., 2007; Wu et al., 2009), and their signals, such as sugar (Yang et al., 2013; Yu et al., 2013), has come into focus. Vegetative, heteroblastic changes in leaf shape have an intimate relationship with reproductive transitions, as demonstrated by recent work linking natural variation in a floral repressor, FLOWERING LOCUS C (FLC), with changes in leaf morphology between species (Cartolano et al., 2015). Of the many gene regulatory networks regulating leaf shape, specific pathways have been identified mediating heteroblastic shape changes (Rubio‐Somoza et al., 2014). Similar molecular descriptions of leaf development exist (Ichihashi et al., 2014) and the crosstalk between these two processes is large. Strangely, although molecular correlates of leaf development and heteroblasty have been uncovered, a phenotypic basis for the morphological effects of temporal patterning remains obscure, except in a qualitative sense.
The decomposition of complex morphologies into latent shapes regulated by distinct pathways is required to precisely describe the mechanisms which, together, produce the composite shape of a single leaf. By isolating latent shapes that are regulated by evolution and development, we can focus on those attributes of the leaf most relevant for the production of the diversity of leaf shapes observed in nature, and consider the functional contributions of leaf morphology to assessments of biodiversity, plant responses to global climate change and the genetic improvement of crop species.
Author contributions
D.H.C., L.L.K., A.J.M. and J.P.L. designed the research; D.H.C. contributed new analytic/computational (etc.) tools; D.H.C., L.L.K., R.O.H., S.C., M.G. and C.K. analyzed the data; D.H.C., L.L.K., A.J.M. and J.P.L. wrote the paper.
Supporting information
Please note: Wiley Blackwell are not responsible for the content or functionality of any supporting information supplied by the authors. Any queries (other than missing material) should be directed to the New Phytologist Central Office.
Fig. S1 Numbers of species, vines, leaves and shoot positions sampled.
Fig. S2 Developmental stage and leaf number are partially confounded.
Fig. S3 Projection of species with five vines or more onto the morphospace.
Fig. S4 Comparison of leaf shape changes caused by developmental stage (Sn) and leaf number (Ln) for different species.
Table S1 Procrustes‐aligned coordinates
Table S2 Correlation between traits
Acknowledgements
This work was funded by research stipends awarded to D.H.C., L.L.K. and J.P.L. through the Grape Research Coordination Network, DBI0741876.
References
- Abramoff MD, Magalhaes PJ, Ram SJ. 2004. Image processing with ImageJ. Biophotonics International 11: 36–43. [Google Scholar]
- Allsopp A. 1954. Juvenile stages of plants and the nutritional status of the shoot apex. Nature 173: 1032–1035. [Google Scholar]
- Ashby E. 1948. Studies in the morphogenesis of leaves. I. An essay on leaf shape. New Phytologist 47: 152–176. [Google Scholar]
- Bailey IW, Sinnott EW. 1915. A botanical index of Cretaceous and Tertiary climates. Science 41: 831–834. [DOI] [PubMed] [Google Scholar]
- Benjamini Y, Hochberg Y. 1995. Controlling the false discovery rate: a practical and powerful approach to multiple testing. Journal of the Royal Statistical Society. Series B (Methodological) 57: 289–300. [Google Scholar]
- Bharathan G, Goliber TE, Moore C, Kessler S, Pham T, Sinha NR. 2002. Homologies in leaf form inferred from KNOXI gene expression during development. Science 296: 1858–1860. [DOI] [PubMed] [Google Scholar]
- Blein T, Pulido A, Vialette‐Guiraud A, Nikovics K, Morin H, Hay A, Johansen IE, Tsiantis M, Laufs P. 2008. A conserved molecular framework for compound leaf development. Science 322: 1835–1839. [DOI] [PubMed] [Google Scholar]
- Boss PK, Thomas MR. 2002. Association of dwarfism and floral induction with a grape ‘green revolution’ mutation. Nature 416: 847–850. [DOI] [PubMed] [Google Scholar]
- Carmona MJ, Chaib J, Martinez‐Zapater JM. 2008. A molecular genetic perspective of reproductive development in grapevine. Journal of Experimental Botany 59: 1579–2596. [DOI] [PubMed] [Google Scholar]
- Cartolano M, Pieper B, Lempe J, Tattersall A, Huijser P, Tresch A, Darrah PR, Hay A, Tsiantis M. 2015. Heterochrony underpins natural variation in Cardamine hirsuta leaf form. Proceedings of the National Academy of Sciences, USA 112: 10539–10544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chitwood DH. 2014. Imitation, genetic lineages, and time influenced the morphological evolution of the violin. PLoS ONE 9: e109229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chitwood DH, Headland LR, Ranjan A, Martinez CC, Braybrook SA, Koenig DP, Kuhlemeier C, Smith RS, Sinha NR. 2012a. Leaf asymmetry as a developmental constraint imposed by auxin‐dependent phyllotactic patterning. Plant Cell 24: 2318–2327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chitwood DH, Kumar R, Headland LR, Ranjan A, Covington MF, Ichihashi Y, Fulop D, Jimenez‐Gomez JM, Peng J, Maloof JN et al 2013. A quantitative genetic basis for leaf morphology in a set of precisely defined tomato introgression lines. Plant Cell 25: 2465–2481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chitwood DH, Naylor DT, Thammapichai P, Weeger AC, Headland LR, Sinha NR. 2012b. Conflict between intrinsic leaf asymmetry and phyllotaxis in the resupinate leaves of Alstroemeria psittacina . Frontiers in Plant Science 3: 182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chitwood DH, Ranjan A, Martinez CC, Headland LR, Thiem T, Kumar R, Convington MF, Hatcher T, Naylor DT, Zimmerman S et al 2014. A modern ampelography: a genetic basis for leaf shape and venation patterning in grape. Plant Physiology 164: 259–272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chitwood DH, Topp CN. 2015. Revealing plant cryptotypes: defining meaningful phenotypes among infinite traits. Current Opinion in Plant Biology 24C: 54–60. [DOI] [PubMed] [Google Scholar]
- Chuck G, Cigan AM, Saeteum K, Hake S. 2007. The heteroblastic maize mutant Corngrass1 results from overexpression of a tandem microRNA. Nature Genetics 39: 544–549. [DOI] [PubMed] [Google Scholar]
- Claes P, Liberon DK, Daniels K, Rosana KM, Quillen EE, Pearson LN, McEvoy B, Bauchet M, Zaidi AA, Yao W et al 2014. Modeling 3D facial shape from DNA. PLoS Genetics 10: e1004224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cornwell WK, Westoby M, Falster DS, FitzJohn RG, O'Meara BC, Pennell MW, McGlinn DJ, Eastman JM, Moles AT, Reich PB et al 2014. Functional distinctiveness of major plant lineages. Journal of Ecology 102: 345–356. [Google Scholar]
- Diggle PK. 2002. A developmental morphologist's perspective on plasticity. Evolutionary Ecology 16: 267–283. [Google Scholar]
- Dryden IL. 2013. shapes: statistical shape analysis. R package version 1.1‐9. [WWW document] URL http://CRAN.R-project.org/package=shapes [accessed 1 December 2013].
- Efroni I, Blum E, Goldshmidt A, Eshed Y. 2008. A protracted and dynamic maturation schedule underlies Arabidopsis leaf development. Plant Cell 20: 2293–2306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Galet P. 1952. Précis d'ampélographie pratique. Imprimerie P. Déhan. Montpellier, France: Lavoisier. [Google Scholar]
- Galet P. 1979. A practical ampelography: grapevine identification. Translated by L. Morton. Ithaca, NY, USA: Cornell University Press. [Google Scholar]
- Gerrath JM. 1988. Morphological and anatomical development in the Vitaceae. I. Vegetative development in Vitis riparia . Canadian Journal of Botany 66: 209–224. [Google Scholar]
- Gerrath JM. 1993. Developmental morphology and anatomy of grape flowers. Horticultural Reviews 13: 315–337. [Google Scholar]
- Goebel K. 1900. Organography of plants. I. General organography. New York, NY, USA: Hafner Publishing Co. [Google Scholar]
- Goethe JW. 1817. Italienishe Reise. Goethe's Werk. Stuttgart, Germany: Dreizehnter Band. [Google Scholar]
- Goethe JW. 1952. Botanical writings. Translation, B. Mueller. Honolulu, HI, USA: University of Hawaii Press. [Google Scholar]
- Greenwood DR. 1992. Taphonomic constraints on foliar physiognomic interpretations of Late Cretaceous and Tertiary palaeoclimates. Review of Palaeobotany and Palynology 71: 149–190. [Google Scholar]
- Hales S. 1727. Vegetable staticks or, an account of some statistical experiments on the sap in vegetables. London, UK: W. and J. Innys. [Google Scholar]
- Harrell FE. 2013. Hmisc: Harrell miscellaneous. R package version 3.10‐1.1. [WWW document] URL http://CRAN.R-project.org/package=Hmisc [accessed 1 December 2013].
- Ichihashi Y, Aguilar‐Martinez JA, Farhi M, Chitwood DH, Kumar R, Milon LV, Peng J, Maloof JN, Sinha NR. 2014. Evolutionary developmental transcriptomics reveals a gene network module regulating interspecific diversity in plant leaf shape. Proceedings of the National Academy of Sciences, USA 111: E2616–E2621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones CS. 1993. Heterochrony and heteroblastic leaf development in two subspecies of Curbita argyrosperma (Cucurbitaceae). American Journal of Botany 80: 778–795. [Google Scholar]
- Jones CS. 1995. Does shade prolong juvenile development? A morphological analysis of leaf shape changes in Cucurbita argyrosperma subsp. Sororia (Cucurbitaceae). American Journal of Botany 82: 346–359. [Google Scholar]
- Kerstetter RA, Poethig RS. 1998. The specification of leaf identity during shoot development. Annual Review of Cell and Developmental Biology 14: 373–398. [DOI] [PubMed] [Google Scholar]
- Kim M, McCormick S, Timmermans M, Sinha N. 2003. The expression domain of PHANTASTICA determines leaflet placement in compound leaves. Nature 424: 438–443. [DOI] [PubMed] [Google Scholar]
- Kimura S, Koenig D, Kang J, Yoong FY, Sinha N. 2008. Natural variation in leaf morphology results from mutation of a novel KNOX gene. Current Biology 18: 672–677. [DOI] [PubMed] [Google Scholar]
- Kuhn R, Junqua CJ, Nguyen P, Niedzielski N. 2000. Rapid speaker adaptation in eigenvoice space. IEEE Transactions on Speech and Audio Processing 8: 695–707. [Google Scholar]
- Langlade NB, Feng X, Dransfield T, Copsey L, Hanna AI, Thebaud C, Bangham A, Hudson A, Coen E. 2005. Evolution through genetically controlled allometry space. Proceedings of the National Academy of Sciences, USA 102: 10221–10226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller AJ, Matasci N, Schwaninger H, Aradhya MK, Prins B, Zhong G‐Y, Simon C, Buckler ES, Myles S. 2013. Vitis phylogenomics: hybridization intensities from a SNP array outperform genotype calls. PLoS ONE 8: e78680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moore M. 1991. Classification and systematics of eastern North American Vitis L. (Vitaceae) north of Mexico. SIDA, Contributions to Botany 14: 339–367. [Google Scholar]
- Myles S, Boyko AR, Owens CL, Brown PJ, Grassi F, Aradhya MK, Prins B, Reynolds A, Chia J‐M, Ware D et al 2011. Genetic structure and domestication history of the grape. Proceedings of the National Academy of Sciences, USA 108: 3530–3535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nicotra AB, Leigh A, Boyce CK, Jones CS, Niklas KJ, Royer DL, Tsukaya H. 2011. The evolution and functional significance of leaf shape in the angiosperms. Functional Plant Biology 38: 535–552. [DOI] [PubMed] [Google Scholar]
- Ogburn RM, Edwards EJ. 2013. Repeated origin of three‐dimensional leaf venation releases constraints on the evolution of succulence in plants. Current Biology 23: 722–726. [DOI] [PubMed] [Google Scholar]
- Paradis E, Claude J, Strimmer K. 2004. APE: analyses of phylogenetics and evolution in R languages. Bioinformatics 20: 289–290. [DOI] [PubMed] [Google Scholar]
- Park SJ, Jiang K, Schatz MC, Lippman ZB. 2012. Rate of meristem maturation determines inflorescence architecture in tomato. Proceedings of the National Academy of Sciences, USA 109: 639–644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parkhurst DF, Loucks OL. 1972. Optimal leaf size in relation to environment. Journal of Ecology 60: 505–537. [Google Scholar]
- Peppe DJ, Royer DL, Cariglino B, Oliver SY, Newman S, Leight E, Enikolopov G, Fernandez‐Burgos M, Herrera F, Adamas JM et al 2011. Sensitivity of leaf size and shape to climate: global patterns and paleoclimatic applications. New Phytologist 190: 724–739. [DOI] [PubMed] [Google Scholar]
- Poethig RS. 1990. Phase change and the regulation of shoot morphogenesis in plants. Science 250: 923–930. [DOI] [PubMed] [Google Scholar]
- Poethig RS. 2010. The past, present, and future of vegetative phase change. Plant Physiology 154: 541–544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- R Core Team . 2014. R: a language and environment for statistical computing. R version 3.0.3 ‘Warm Puppy’. Vienna, Austria: R Foundation for Statistical Computing. [Google Scholar]
- Remmler L, Rolland‐Lagan AG. 2012. Computational method for quantifying growth patterns at the adaxial leaf surface in three dimensions. Plant Physiology 159: 27–39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ren H, Wen J. 2007. Vitis. Flora of China 12: 210–222. [Google Scholar]
- Rolland‐Lagan AG, Remmler L, Girard‐Bock C. 2014. Quantifying shape changes and tissue deformation in leaf development. Plant Physiology 165: 496–505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rubio‐Somoza I, Zhou CM, Confraria A, Martinho C, von Born P, Baena‐Gonzalez E, Wang JW, Weigel D. 2014. Temporal control of leaf complexity by miRNA‐regulated licensing of protein complexes. Current Biology 24: 2714–2719. [DOI] [PubMed] [Google Scholar]
- Srinivasan C, Mullins MG. 1981. Physiology of flowering in the grapevine – a review. American Journal of Enology and Viticulture 32: 47–63. [Google Scholar]
- Tian F, Bradbury PJ, Brown PJ, Hung H, Sun Q, Flint‐Garcia S, Rocheford TR, McMullen MD, Holland JB, Buckler ES. 2011. Genome‐wide association study of leaf architecture in the maize nested association mapping population. Nature Genetics 43: 159–162. [DOI] [PubMed] [Google Scholar]
- Venables WN, Ripley BD. 2002. Modern applied statistics with S, 4th edn New York, NY, USA: Springer. [Google Scholar]
- Wen J, Nie Z‐L, Soejima A, Meng Y. 2007. Phylogeny of Vitaceae based on the nuclear GAI1 gene sequences. Canadian Journal of Botany 85: 731–745. [Google Scholar]
- Wen J, Xiong Z, Nie Z‐L, Mao L, Zhu Y, Kan X‐Z, Ickert‐Bond SM, Gerrath J, Zimmer EA, Fang X‐D. 2013. Transcriptome sequences resolve deep relationships of the grape family. PLoS ONE 8: e74394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whorf BJ. 1945. Grammatical categories. Linguistic Society of America 21: 1–11. [Google Scholar]
- Wickham H. 2009. ggplot2: elegant graphics for data analysis. Dordrecht, the Netherlands: Springer Science & Business Media. [Google Scholar]
- Wilf P, Wing SL, Greenwood DR, Greenwood CL. 1998. Using fossil leaves as paleoprecipitation indicators: an Eocene example. Geology 26: 203–206. [Google Scholar]
- Wolfe JA. 1971. Tertiary climate fluctuations and methods of analysis of Tertiary floras. Palaeogeography, Palaeoclimatology, Palaeoecology 9: 27–57. [Google Scholar]
- Wright IJ, Reich PB, Westoby M, Ackerly DD, Baruch Z, Bongers F, Cavender‐Bares J, Chapin T, Cornelissen JHC, Diemer M et al 2004. The worldwide leaf economics spectrum. Nature 428: 821–827. [DOI] [PubMed] [Google Scholar]
- Wu G, Park MY, Conway SR, Wang JW, Weigel D, Poethig RS. 2009. The sequential action of miR156 and miR172 regulates developmental timing in Arabidopsis. Cell 138: 750–759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang L, Xu M, Koo Y, He J, Poethig RS. 2013. Sugar promotes vegetative phase change in Arabidopsis thaliana by repressing the expression of MIR156A and MIR156C. eLIFE 2: e00260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu S, Cao L, Zhou C‐M, Zhang T‐Q, Lian H, Sun Y, Wu J, Huang J, Wang G, Wang J‐W. 2013. Sugar is an endogenous cue for juvenile‐to‐adult phase transition in plants. eLIFE 2: e00269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zecca G, Abbott JR, Sun WB, Spada A, Sala F, Grassi F. 2012. The timing and the mode of evolution of wild grapes (Vitis). Molecular Phylogenetics and Evolution 62: 736–747. [DOI] [PubMed] [Google Scholar]
- Zhang J, Yan Y, Lades M. 1997. Face recognition: eigenface, elastic matching, and neural nets. Proceedings of the IEEE 85: 1423–1435. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Please note: Wiley Blackwell are not responsible for the content or functionality of any supporting information supplied by the authors. Any queries (other than missing material) should be directed to the New Phytologist Central Office.
Fig. S1 Numbers of species, vines, leaves and shoot positions sampled.
Fig. S2 Developmental stage and leaf number are partially confounded.
Fig. S3 Projection of species with five vines or more onto the morphospace.
Fig. S4 Comparison of leaf shape changes caused by developmental stage (Sn) and leaf number (Ln) for different species.
Table S1 Procrustes‐aligned coordinates
Table S2 Correlation between traits