

Abstract
In this review, we analyze the effects of growth hormone on a number of tissues and organs and its putative role in the longitudinal growth of an organism. We conclude that the hormone plays a very important role in maintaining the homogeneity of tissues and organs during the normal development of the human body or after an injury. Its effects on growth do not seem to take place during the fetal period or during the early infancy and are mediated by insulin-like growth factor I (IGF-I) during childhood and puberty. In turn, IGF-I transcription is dependent on an adequate GH secretion, and in many tissues, it occurs independent of GH. We propose that GH may be a prohormone, rather than a hormone, since in many tissues and organs, it is proteolytically cleaved in a tissue-specific manner giving origin to shorter GH forms whose activity is still unknown.
Keywords: growth hormone, growth, nervous system, gonads, liver, cardiovascular system
Introduction
The possible existence of a factor responsible for the longitudinal growth of an organism was first postulated in 1921, when Evans and Long1 reported that treating rats with extracts obtained from bovine anterior pituitary glands led to an increased growth of these animals. However, the lack of the necessary technology delayed for more than 30 years the identification of the factor putatively responsible of such effect on growth, despite the clinical findings indicating that human dwarfs were able to grow after administering human pituitary extracts. In 1956, a protein present in the anterior pituitary gland could be extracted from human pituitaries and then administered successfully to human dwarfs, a novel treatment first communicated by Raben2 for a clinical situation that until then had no solution. However, at that time, the existing technical methods were not sufficient for obtaining a pure and safe product for therapeutic purposes; the pituitary extracts had to be obtained from human cadavers, and some of them were contaminated with nervous tissue from the posterior pituitary lobe. This led to the occurrence of a number of cases of iatrogenic Creutzfeldt-Jakob disease in children who received pituitary extracts in which pathogenic prions were present in the nervous tissue of the posterior pituitary lobe.3,4
In 1978, Sairam et al5 reported the isolation of growth hormone (GH) from human pituitary glands in an active form well suited for clinical use. This had been preceded, since 1944, by numerous studies by Li and Dixon6 in different animal species, including human beings, that culminated in the identification of the primary sequence of human GH in 1971. After that, the advances in the field of genetical engineering allowed to produce, since 1981, practically unlimited quantities of the hormone, pure and safe, by DNA recombinant technology, first in prokaryotes (Escherichia coli) and later in eukaryotes (murine fibroblasts), to be used in GH-deficient (GHD) children. The only difference between the hormone produced in prokaryotes and eukaryotes is the presence of the 20 kDa GH variant in the preparations obtained from eukaryotes, since alternative splicing cannot take place in prokaryotes; however, no differences have been described in terms of growth effectivity or any other effects between the different recombinant GHs in the market. From 1985, the spectrum of therapeutic applications of the recombinant GH was expanded, especially in the pediatric population, such as children born small for gestational age, children with idiopathic short stature, and children with growth delay due to chronic renal insufficiency, Turner syndrome, Prader–Willi syndrome, SHOX gene deficiency, and Noonan syndrome, and also in adults with GH deficiency (in Spain, whenever they have any other pituitary hormone deficiency [excepting prolactin]) and in patients with acquired immunodeficiency syndrome (AIDS) wasting.
While it is clear that the longitudinal growth of a GHD child is defective unless he/she receives the adequate daily dose of the hormone, a number of clinical and experimental findings lead us to ask ourselves if GH is in fact the real GH.
A classic summary of GH effects would describe GH as: “a metabolic hormone that induces hyperglycemia (as a counterregulatory hormone), lipolysis, and protein anabolism. In addition, GH induces the expression of many other growth factors (mainly insulin-like growth factor I [IGF-I]) and has direct effects on cellular proliferation and differentiation”. The GH effects on growth probably occur as a consequence of the metabolic induction of expression of other growth factors and the proliferative actions during a short period of time along the life of an organism.
According to a recent review by Murray and Clayton,7 human growth is a process that starts after conception and finalizes in adolescence at the time of growth plate fusion. They divide growth into the following four phases: (1) fetal, (2) infancy, (3) childhood, and (4) puberty. These different growth stages have been previously described and widely analyzed by Karlberg et al.8–11 For instance, they first described that “linear growth during the first three years of life can be represented mathematically using a combination of a quickly decelerating Infancy component with the addition of a slowly decelerating Childhood component that acts from the second half of the first year of life”. The appearance of this second component leads to an abrupt increase in growth velocity during the second half of the first year of life. Since a similar increase is observed in GHD children when GH therapy begins, they conclude that the Childhood component defines the age at which GH begins to play a significant physiological influence on linear growth.8
Before discussing the involvement of GH in each of these phases, some important concepts need to be analyzed.
The GH gene family
Clasically, it was thought that proteic hormones exert only their biological effects after interacting with specific membrane receptors. Then, the consequent activation of these receptors would initiate a cascade of intracellular signaling pathways, leading to the appearance of hormone-specific biological effects. That is, the proteic hormones would not enter into cells for inducing, or modifying, any kind of activity, different to what happens with steroids, for instance, which have to enter into cells because their receptors are located in the cytoplasm or in the nucleus of the cell. However, this concept was challenged with the report that insulin and epidermal growth factor could be internalized in living cells,12 followed by the description of the mechanism by which polypeptide hormones may be internalized and the significance of this finding.13 Waters et al14–16 first demonstrated that GH, after interacting with its membrane receptor, is internalized together with its receptor (GHR) via the endosomal pathway and that this mechanism allows the translocation of both GH and GHR to the nucleus where they promote the transcription of a number of genes. On this basis, the detection of GHR in the nucleus of a cell indicates that there has been a previous interaction GH–GHR at the cell membrane level. But apart from this nuclear translocation and its effects on gene transcription, we demonstrated that the internalized GH also suffers a tissue-specific proteolytic processing, influenced by sex and age, which gives origin to a number of different molecular forms, whose actions are largely unknown.17 The knowledge of these concepts is important for a better understanding of the diverse effects of GH.
Equally important is to know that GH is a pleiotropic hormone, expressed not only at the pituitary level but also in a number of cells and tissues, which plays a number of different roles (endocrine, autocrine/paracrine) far beyond those classically described.17–20 The hGH/hCS locus contains two GH genes and three CS (chorionic somatomammotropin, now known as placental lactogens) genes spanning 48 kb of DNA in the chromosome 17 in the order: 5′-(hGH-1/hCS-5/hCS-1/hGH-2/hCS-2)-3′.21 Among them, GH-1 (now known as GH-N) is expressed in the pituitary gland and peripheral tissues, while the other four seem to be expressed only in the placenta, although N-glycosylated GH-related peptides have been found in human pituitary extracts, suggesting that the hGH-2 gene (now known as GH-variant [GH-V]), or other unknown GH-related genes, could be expressed too at the pituitary level, and perhaps in other tissues, since the hGH-N gene lacks the consensus sequence for N-glycosylation observed in some nonplacental GH-related products.22,23
Figure 1 shows the hGH gene family and the main products of expression of its genes.
Figure 1.

The products of expression of the hGH gene family share 95% of homology among them. The main GH form corresponds to the 22 kDa GH-N, expressed not only at the pituitary level but also in many different peripheral cells and tissues. About 10% of pituitary GH-N transcripts suffer a physiological postranscriptional modification, occurred by alternative splicing of exon III, giving origin to a 20 kDa variant. Among placental GH genes, the most important seems to be the GH-V, which differs from 22 kDa GH-N in only eight amino acids.
However, this is only a schematic description of the hGH gene family, but pituitary and tissue GH heterogeneity is really high.
For instance, in human pituitary, many GH isoforms may be found. Apart from the principal GH products, 22 and the 20 kDa GH form, the later resulting from the alternative mRNA splicing of the GH-N gene, there are a number of posttranslationally modified GH forms (N-acylated, deamidated and O-glycosilated monomeric GH forms), as well as both noncovalent and disulfide-linked oligomers up to pentameric GH.24,25 Moreover, in human pituitary extracts, some significant amounts of shorter molecular weight GH variants (17 and 5 kDa) can be found. These are originated by selective cleavage of the bond between amino acids 43 and 44, leading to the production of fragments 1–43 and 44–191 in the own somatotroph cells, and they have also been found in plasma and tissues from human beings and some different species.17,25–30 Given that these forms can be originated by specific cleavage of the main 22 kDa GH form, it is likely that they appear in tissues as a result of posttranslational modification of the hormone. Interestingly, while human GH 22 kDa has both insulin-like and diabetogenic effects, these fragments proteolytically generated show opposite effects: the short GH 5 kDa potentiates the effects of the insulin, while the 17 kDa form shows diabetogenic activity.26 Moreover, since these two isoforms can be generated under acidic conditions, it has been postulated that they can play a significant physiological role because the hormone in the cells is exposed to an acidic environment.26 Similar conclusions regarding the effects of these 17 and 5 kDa GH variants had been reached by Lewis et al at the end of the last century.31,32 According to their data, these GH fragments, obtained by recombinant DNA technology, have potent in vivo effects on glucose homeostasis in rodents, but cannot stimulate body growth.
Once established these important concepts, we will now analyze how growth occurs and what is the role that GH plays in the different steps of this physiological process.
GH and IGF relationships
In 1957, Salmon and Daughaday33 reported the presence of a factor hormonally controlled in serum, which stimulated sulfate incorporation by cartilage in vitro. Soon this factor was isolated and was found to be a small peptide very similar to insulin, both in its structure and mechanisms of action. Therefore, the initial name of somatomedin C was substituted by IGF-I. The purpose of this review is not to analyze how IGF-I acts, but some descriptions about it are needed.
Initially it was thought that IGF-I was produced only in the liver and that its hepatic expression was fully dependent on GH. However, we know today, that while liver is the main source of circulating IGF-I, this peptide is produced, as it occurs with GH, practically in all cells and tissues, where it plays an auto/paracrine role. Even IGF-I is expressed within the pituitary gland, where it may act as a paracrine regulator of GH secretion.34 The original somatomedin hypothesis proposed that GH stimulates hepatic production of IGF-I, which in turn promotes growth directly at the epiphyseal plate.35 Posteriorly, a dual somatomedin hypothesis was proposed,36 suggesting that local IGF-I promotes chondrocyte maturation and longitudinal growth in an auto/paracrine manner,37 while the relationships between GH and IGF-I mainly consist on GH inducing liver IGF-I transcription which, in turns, negatively controls pituitary GH secretion.38 (For a detailed description about how pituitary secretion of GH is controlled, see Refs. 39 and 40.) However, there are direct actions of GH on bone: GH induces unilateral epiphyseal growth when injected,41 or infused locally,42 and promotes chondrocyte proliferation in vitro.43 Moreover, IGF-I antibodies block the proliferative effects of GH, indicating a local IGF-I action dependent on GH stimulation of local IGF-I production.44 Taken together, these findings indicate that GH acts directly on chondrocyte stem cells and also promotes chondrocyte proliferation via local IGF-I production. IGF-I then would stimulate clonal expansion of proliferating cells in an auto/paracrine manner.45
A direct evidence of the role of IGF-I in the longitudinal growth of the human body proceeds from the first observation by Laron and Kauli, almost 60 years ago, communicating that some short-height children who resembled patients with isolated GHD had high serum levels of GH but a defect in liver GHR that impeded GH signaling for hepatic IGF-I expression.46,47 Since that first report, a number of publications demonstrated that defective liver GHR led to dwarfism because of the liver’s inability to synthesize IGF-I. This situation reverted when IGF-I produced by recombinant DNA technology became available and children with Laron syndrome could be successfully treated. The same occurs in GHR null mice, in which a severe postnatal growth retardation exists, and while plasma IGF-I levels are markedly reduced, plasma GH levels are elevated.48,49
According to the concepts described above, IGF-I would really be the GH, while GH would merely be an inducer of its hepatic and chondrocyte expression. However, some clinical evidences indicate that the role of GH as an inducer of hepatic IGF-I expression is dependent on the nutritional status of the organism. In fact, growth velocity in obese children is normal or even higher than that in their age-related normal body weight children, despite that GH secretion is decreased, both physiologically and in response to any provocative stimulation, or even absent in obese children, plasma IGF-I values are elevated.50,51 Multiple deconvolution studies have defined deficiencies in both basal and pulsatile GH secretion with increasing body mass index and, consistently, a reduction in both GH pulse amplitude and orderliness of secretion. Reduced frequency of pulses and alterations in the half-life of GH has been variably reported as contributing to the overall reduction in GH secretion.52
The opposite situation can be seen in anorexia nervosa patients. In spite of the increased GH secretion, plasma IGF-I values are drastically reduced.53,54 Looking at the ends of the spectrum of body weight and analyzing how GH and IGF-I behave in these extremes, one might conclude that there is a need for an efficient hepatic metabolism of glucose to GH may induce hepatic expression of IGF-I. Even more, it has been demonstrated, in cultured hepatocytes from perinatal rats, in a serum-free condition, and in the presence of nonstimulatory dose of insulin, that glucose was able to induce a clear increase of IGF-I and IGF-II mRNA. However, administering 2-deoxyglucose, that the liver cannot metabolize, blocks IGF-I gene expression. This agrees with the clinical data described above and suggests that IGFs belong to a family of genes that are positively regulated by glucose.55 Moreover, we found a high prevalence of low plasma IGF-I values and a clearly delayed growth in children with spastic cerebral palsy.56 It is feasible to assume that spasticity increases the muscular uptake of glucose, therefore decreasing the liver uptake with the subsequent effects on IGF-I production and longitudinal growth.
Figure 2 shows a schematic overview of how the hepatic expression of IGF-I may be regulated.
Figure 2.

Possible modulation of the hepatic expression of IGF-I gene. GH and insulin may directly induce the transcription of IGF-I gene, but for that, glucose uptake and metabolization by the liver is needed. Hepatic glucose metabolism may directly induce the transcription of IGF-I gene. In addition, liver uptake of some amino acids, induced by GH or insulin, facilitates IGF-I expression, because these amino acids stabilize the IGF-I mRNA.
Given that the longitudinal growth of the organism occurs at the epiphyseal growth plate level as a combination of chondrogenesis and ossification, we will analyze briefly how this process occurs (for a more detailed review, see Ref. 57).
Growth plate physiology
In mammals, the growth plate is a cartilaginous tissue with specific functions since growth begins until epiphysis is closed after puberty. It is composed of three different zones: the resting zone, the proliferative zone, and the hypertrophic zone. In the resting zone, there are stem cells that slowly replicate and generate new clones of proliferative chondrocytes.58 These replicate at a high rate and align themselves in columns oriented parallel to the long axis of the bone.59 As they are progressively separated from the epiphysis, replication is decreasing until it stops; they then enlarge and alter the extracellular matrix to form the hypertrophic zone. This hypertrophic cartilage attracts the blood vessels, osteoclasts, and differentiating osteoblasts, which remodel the newly formed cartilage into bone tissue.60
Progressively, after puberty, the growth plate experiences functional and structural changes, leading to what is known as growth plate senescence. As a consequence, the longitudinal growth of bone slows until ceasing and determining the length or height of the adult body. This process has been postulated to be due to a mechanism intrinsic to the growth plate.61
As indicated above, a number of evidences indicate that the GH/IGF-I system regulates longitudinal bone growth at the growth plate, either by GH-induced liver production of IGF-I, which then acts as an endocrine factor to stimulate longitudinal bone growth (this would explain the results obtained when treating with recombinant IGF-I children with Laron syndrome), or by a local action of GH on bone,41 an effect that may be mediated in part by stimulation of local IGF-I production.42,43 It has been proposed that GH would act mainly on the resting zone of the growth plate, whereas IGF-I would act on the proliferative zone.62 However, this has not been confirmed by other studies.63,64
In an elegant study, using manual microdissection and quantitative real-time PCR, Parker et al57 found that IGF-II, and not IGF-I, is the predominantly expressed IGF in growth plate chondrocytes of young rapidly growing rats. Moreover, they found that in one-week-old rats, GH receptor (GHR) and type 1 IGF receptor (IGFR1) are expressed in all zones of the growth plate, while the expression of IGF-binding proteins (IGFBPs) is very low as compared with that found in other rat tissues. Since growth velocity in rats decreases with age, IGF-II expression drastically decreases while the expression of many of the IGFBPs and the type 2 IGFR increases. Given that the availability of IGFs depends on the amount of IGFBPs, these data indicate that the abundance of free bioactive IGFs in growth plate cartilage decreases with age. Their findings also indicate that IGF-I is expressed at very low levels in growth plates, suggesting that local production of IGF-I by growth plate chondrocytes does not seem to be biologically important. They suggested that IGF-I protein in the growth plate is likely a product that mainly arrives by diffusion from local sources in perichondrium and/or adjacent bone and from liver via the circulation. Contrary to IGF-I, the same study found that IGF-II is highly expressed in the growth plate cartilage,57 confirming data from previous studies using in situ hybridization.65,66 Moreover, in their study, Parker et al57 found that expression of IGF-II mRNA is higher in the resting and the proliferative zones than in the hypertrophic zone; this led them to suggest that IGF-II may regulate cell proliferation in the former two zones of the growth plate. With regard to GH, its receptor seems to be expressed at similar levels in all zones of the growth plate, whereas IGFR1 was expressed at threefold higher levels in the proliferative zone than in the hypertrophic zone. These data indicate that GH acts on the three zones of the growth plate, and since the type 1 IGFR is also expressed throughout the growth plate, it can be deduced that IGFs (I and II) regulate all zones of the growth plate. According to the data of the study by Parker et al,57 the possibility exists that GH may act on growth plate chondrocytes by an IGF-independent mechanism, but the hormone may also exert effects by stimulating IGF-I production in the perichondrium. Interestingly, in their study, they found that mRNA expression of IGFBPs increases with age. IGFBP-1, IGFBP-2, IGFBP-3, and IGFBP-4 increased in perichondrium, and IGFBP-3 and IGFBP-4 increased in metaphyseal bone. This pattern of increased expression of many of the IGFBPs with age in the growth plate, perichondrium, and bone further supports the idea that growth plate senescence may be in part due to decreased availability of IGFs. Interestingly, the most consistent increase in IGFBP expression is seen in the perichondrium where IGF-I mRNA expression is the highest, suggesting an inhibitory effect on IGF-I action while aging.57
The study by Parker et al57 relates GH, IGFs, and IGFBPs to the longitudinal growth of the organism, and their results in rats partially agree with clinical findings such as those described in children with Laron syndrome. As stated before, this syndrome is a cause of short stature due to an abnormality of GHR that is traduced in a complete insensitivity to GH. This leads not only to a very short stature but also to an early hypoglycemia. The lack of GH responsiveness, and also the hypoglycemia, leads to the absence of production of IGFs. Therefore, treatment with recombinant IGF-I is the only therapeutic option that improves the prognosis and the height in these patients. In this case, however, it seems to be impossible that IGF-II could be expressed in the growth plate. The only possible explanation to the improvement in growth velocity and final height in children with Laron syndrome obtained during treatment with IGF-I is that, while its expression in the resting and proliferative zones of the growth plate is physiologically low, the exogenous IGF-I administration may compensate this low expression. Moreover, as described above, type 1 IGFR is expressed throughout the three growth plate zones.
Recently, it has been published that, in mice, GH stimulates growth plate chondrogenesis and longitudinal bone growth directly at the growth plate, even when the local effects of IGF-I and IGF-II are prevented.67
In summary, and according to the studies described here, the effects of GH, IGF-I, and IGF-II on growth remain to be fully elucidated; even more, a recent study challenged the importance of IGF-I for endochondral growth, demonstrating, in mice, the critical importance of suppressor of cytokine signaling-2 (SOCS2) in regulating GH ability to promote bone growth via a mechanism that is independent of IGF-I.68 To better clarify this concept, this study was performed in juvenile mice SOCS2 (−/−). In these animals, but not in wild-type animals, IGF-II is elevated, and the IGF receptor is required to be expressed. Therefore, the possibility exists that in these conditions, GH promotes bone growth via IGF-II acting on type 1 IGFR.
Further studies are needed to clarify the exact role of GH and IGFs on the longitudinal growth of the organism, and also how aging may change the role played by each of these growth factors at bone level.
We will now analyze how plasma GH changes across the life and how growth differentially depends on nutritional and/or growth factors. Before that, we must recapitulate about what are GH effects: the hormone promotes stem cell activation, cell proliferation, differentiation, and survival, either directly or through the induction of IGF-1 and/or many other trophic factors. GH acts via its cell membrane receptor to initiate a cascade of signal transduction pathways downstream of the GHR. The transcription factor STAT5b is regarded as the major signaling pathway by which GH achieves its physiological effects, including the liver expression of IGF-I,69 and also regulates the expression of a host of other genes, some of which are important growth regulators. In addition to signaling from the cell membrane, the GHR translocates to the nucleus in a GH-dependent manner, and once in the nucleus it regulates the expression of other cell growth-related genes and sensitizes the cell to the proliferative action of GH.70 Initially, we will analyze the effects of GH on gonads and gametogenesis (for a more detailed review, see Ref. 71).
GH and gonads
GH and gonadal function in males
The testis seems to behave as a small hypophysis, since different to what happens at the nervous system, circulating GH cannot easily access testicular cells within the blood–testis barrier. Therefore, the ligands for the GHRs on these cells are thus likely to be produced within the testis. Supporting this hypothesis, GH gene expression has been detected within the rat, human, and chicken testis.72 Curiously, the GH-variant (GH-V) gene products, previously thought to be pregnancy specific, are the most abundant GH mRNA isoform in the human testis.73
GHR immunoreactivity has been detected in the human testis, and it is concentrated in Leydig cells.74 Potential GH regulators are similarly expressed in the testis. In rats and human beings, for instance, testicular Growth Hormone Releasing Hormone (GHRH) closely resembles placental GHRH and is capable of stimulating pituitary GH release and Sertoli cell adenylate cyclase activity.72 GHRH receptors have a wide distribution in human beings, including Leydig cells, Sertoli cells, germ cells, and the prostate gland, suggesting that GHRH may exert testicular actions distinct from GH.75
More recently, ghrelin and putative ghrelin receptors have been localized in the testis, and ghrelin alters testosterone synthesis and other testicular parameters.76,77 In addition, negative regulators of GH secretion and actions, such as somatostatin and its receptors (SSTR 1–5), have been detected in mice Sertoli cells;78 somatostatin treatment significantly promotes apoptosis of these cells and decreases IGF-I expression together with a dose-dependent suppression in the mRNA level of kitl gene, which is important in the regulation of spermatogenesis. These findings suggest that somatostatin and its receptors (mainly SSTR2 and SSTR5) play an important role in the regulation and development of Sertoli cells.78
In all, these data indicate that all the components of the hypothalamic–somatotropic axis exist in the testes, although how it acts and its relationship with the similar endocrine axis are not well known.
However, endocrine GH promotes testicular growth and development and stimulates gametogenesis and steroidogenesis in the adult testes. These actions seem to be mediated by IGF-I, since it can recover testicular differentiation in fetal mice treated with GH antibodies and testicular growth in children with Laron syndrome.79,80 In addition, GH is a potent steroidogenic factor in vitro that stimulates androgen and estradiol production by Leydig cells in a number of species including human beings. In vivo, GH treatment has been seen to improve the production of testosterone induced by chorionic gonadotropin in fertile GHD subjects.81,82 However, GH treatment in hypopituitary or moderately obese men decreases the concentrations of total serum testosterone,83,84 most likely an effect due to a stimulatory effect on aromatase activity and the resulting conversion of testosterone to estradiol.85 The effects of GH on testicular steroidogenesis may be due to enhancing testicular LH sensitivity and promoting development of Leydig cells.
The relevance of testicular IGF-I to gonadal function is well established;86 however, gonadotropins, rather than GH, may be its primary regulator.87,88 Testicular IGF-I in rats, for instance, responds poorly to changes in the systemic GH concentration.89 GH does not stimulate IGF-I production in Leydig cells isolated from horse,90 and in chickens, testicular IGF-I production appears to be entirely GH independent, since it is elevated in GH-resistant dwarf chickens.91
In summary, from these data, it is likely that, in a normal man, endocrine GH synergizes with gonadotropins, potentiating the effects of these hormones on testicular cells while the role of the testicular GH axis and its relationships with endocrine GH remain unknown.
GH and gonadal function in females
Today it is clear that GH is a necessary hormone for female fertility. Fertility is decreased in GHD women,92 and GH replacement allows successful pregnancies in previously infertile GHD women.93
Normal fertility requires two processes, namely, oogenesis and folliculogenesis, that depend on a complicated system of intracrine, juxtacrine, autocrine, paracrine, and endocrine signals. GH, both of pituitary and ovarian origin, may be a modulatory signal in this complex interplay.
In order to produce a normal viable embryo, steroidogenesis, folliculogenesis, and oocyte maturation have to be significantly linked and optimized.
Pituitary gonadotropins are the primary regulators of ovarian steroidogenesis, but in vitro evidence suggests that GH also modulates progesterone and estradiol release. GH stimulates progesterone and estradiol production from bovine granulosa cells94,95 and human luteinized granulosa cells.96,97 GH effects vary throughout the ovarian cycle, since GH stimulates basal progesterone production in porcine corpora lutea, but not follicles,98 and enhances leptin-induced progesterone production in follicles.99
In the presence of FSH, GH promotes early reactions in the steroidogenic pathway via increased local IGF-I, thereby enhancing progesterone synthesis. Conversely, GH inhibits FSH-induced aromatase activity and thus estradiol synthesis, by an IGF-I independent pathway, since IGF-I alone stimulates aromatase activity.
In vivo studies of GH-induced steroidogenesis have produced contradictory results, perhaps because the effects of exogenous GH administration may be modified by other factors, for instance ovarian GH production. However, the fact that GHR-deficient cattle shows a partial progesterone deficiency suggests that GH is physiologically relevant to ovarian steroidogenesis.100 Many different studies indicate that GH is also important for the development and maintenance of ovarian follicles. Its administration increases follicular size and number,101 effects related to the proliferative and antiapoptotic actions of the hormone. In fact, mice with GHR deletion have more primordial follicles and fewer primary, secondary, preantral, and antral follicles, as well as increased follicular atresia.102,103 These result in lower ovulation and implantation rates, fewer corpora lutea, and smaller litter sizes.102 The follicular actions of GH may be due to interactions with the bone morphogenetic protein (BMP) system (one of the factors secreted by oocyte), since GH downregulates BMP receptors and upregulates inhibitors of BMR signaling (such as Smad 6/7).104 In turn, BMP signaling inhibits the formation of GHRs, IGF-I, and IGF-IRs.104 GH has been shown to promote nuclear maturation of denuded oocytes from mice,105 and human beings,106 suggesting that the hormone also acts directly at the oocyte. Moreover, GHR mRNA is readily detectable in oocytes from many species, including human beings,107 and in mature human ovaries, GHR immunoreactivity has also been detected in the oocyte nucleus.108 Since, as described before, the translocation of GHR to the nucleus depends on a previous interaction GH–GHR, this finding is consistent with a direct effect of GH on oocytes. Therefore, GH may improve nuclear maturation and thus oocyte quality. This is important for the use of the hormone in in vitro fertilization protocols.
At the earlier stages of pregnancy, the maintenance of the corpus luteum, secreting progesterone in the needed amounts for allowing implantation and avoiding abortion, is a key process. During these stages, until the placenta begins to produce progesterone, it seems that GH plays a role in luteal cells, proliferative and antiapoptotic, since GHR mRNA and GHR immunoreactivity have been detected in these cells in a number of species including human beings.109 While these effects of GH on the ovary are beyond any doubt, the question is to know whether they are produced by endocrine or exogenously administered GH or they depend on the production of GH by their own ovary.
It has been suggested that plasma GH modulates ovarian function;110 however, granulosa cells and oocytes are avascular, and they are separated from the systemic circulation by the basal lamina.111 Hence, the ovarian actions of GH are most likely due to the local production of the hormone.
In fact, GH mRNA and immunoreactivity are detectable in ovarian stromal and follicular tissue from numerous species, including human beings.112 Ovarian GH production is greater in the inner, avascular follicular compartments, since GH mRNA is detectable in granulosa cells and oocytes, but is absent in cumulus cells and is less abundant in or absent in thecal cells, at least in bovines and chicken.113,114
GH gene expression is initiated very early in follicular development in human beings, since GH mRNA and immunoreactivity were detected in the oocyte cytoplasm and occasionally the granulosa cells of fetal primordial follicles.108
It has been proposed that the GH gene expression increases during follicular development, since GH transcripts have been detected in mature (but not immature) bovine follicles.113 The temporal and spatial patterns of follicular GH expression are parallel to those of GHR expression in human follicles,108 supporting the possibility of auto/paracrine ovarian GH actions.
It is unclear how ovarian GH synthesis is regulated, although GHRH mRNA and GHRH receptor immunoreactivity are present in human beings and rats.115,116 Most likely, ghrelin may act as the primary ovarian GH secretagogue, since ghrelin increases GH secretion but not synthesis in cultured porcine follicles,117 and both ghrelin and its receptor have been found in the ovaries of pig118 and hen.119 However, the role of ghrelin in human ovaries is not known.
As described before, GH-induced hepatic IGF-I is the mediator of many of the GH effects in different tissues. However, the fact that GHR mRNA has been detected in ovarian follicles in many species suggest that follicular GH actions are independent of hepatic IGF-I,101 but does not exclude the possibility that a local expression of IGF-I could mediate GH actions on ovaries. In fact, GH stimulates IGF-I expression in porcine120 and rat granulosa cells.104 In addition, IGF-I antibodies block GH-induced oocyte maturation in rat follicles,121 and IGF-I administration improves follicular maturation in GHR knockout mice102 and the maturation of mouse oocytes nonsynergistically with GH.105 However, other data suggest that GH and IGF-I act independently, at least in part. Follicular IGF-I levels are normal in GHR knockout mice; despite delayed follicular maturation,102,103 this may possibly reflect that other hormones (for instance, hCG and estradiol) are responsible for activating IGF-I production.122 The possibility exists that local IGF-I mediates GH effects on ovaries in some species but not in others, ie, a species-specific manner. Supporting this concept, it has been observed that IGF-I and GH have synergistic effects (hence independent) on the development of ovine preantral follicles.123 Finally, GH stimulates IGF-II production in cultured human granulosa cells,124 suggesting that IGF-II, instead of or in addition to IGF-I, may be an important follicular mediator of GH action; the hormone would sensitize the granulosa cells to the IGF-II produced by the cells themselves, which acts through the IGF-I receptor.
In this section, we analyze the multiple effects of GH on gonads. Many aspects of GH effects at this level remain to be well established, and it is likely that some controversial data depend on species specificity. However, it seems to be clear that GH is a key hormone for a normal gonadal function, in both sexes. Whether its effects depend on endocrine GH or on the hormone produced locally is not well known, since in both testes and ovaries there is a GH axis, similar to the hypothalamic–somatotropic axis. Therefore, it is likely that, at least for some effects, there is an association between systemic GH and the hormone produced in the gonads, as we will analyze later in the central nervous system (CNS).
We will now review how growth occurs along the four stages above described: (1) fetal, (2) infancy, (3) childhood, and (4) puberty.
GH and growth
Growth during the fetal period
The relationship between GH and gonads has been widely analyzed, and it is clear that GH/IGFs play regulatory roles in reproduction. Supporting these data, many women with GH deficiency suffer gonadal deficiencies or insufficiencies.92,125 Clinically, hypogonadotropic hypogonadism women, who responded poorly to high-dose gonadotropin treatment, achieved more oocytes, higher fertilization, and pregnancy rates after being co-stimulated with GH.126 It was reported that the use of GH reduced the human menopausal gonadotropin dose and duration required for ovulation induction and improved success rates.127,128
Interestingly, coinciding with the progressive increase of ovarian production of estradiol during the late follicular phase of a menstrual cycle, usually ending with ovulation, both the amplitude and frequency of plasma GH peaks reach very high values.40 While their origin is related to the effect of estradiol on pituitary GH secretion, it is likely that the objective of this increased GH release is related to the upregulation of the GHR in the oocyte. The GHR has been found in the nucleus of the zygote, therefore indicating that a GH–GHR membrane interaction had to occur after fertilization, as shown in Figure 3.
Figure 3.

Preimplantation period. (1) After interacting with GH, most likely secreted from the oviduct, the GHR is translocated to the nucleus of the zygote, promoting the transcription of genes involved in cells proliferation. GH induces the expression of the glucose transporter Glut 1, which leads to glucose uptake by the zygote therefore providing the substrate needed for the production of the energy necessary for beginning proliferation processes shown in (2) It is unclear if IGF-I is secreted from the oviduct in 1 or 2. (3) While the newly formed embryo is progressing along the oviduct to the uterus, there is a clear secretion of GH and IGF-I from the oviduct walls; IGF-I receptor appears now together with GHR in the membrane of cells constituting the morula, facilitating cell proliferation. (4) In a further step, the blastocyst is formed and their cells begin to express GH, GHR, IGF-I, and IGF-I R. This precedes the implantation in the uterus.
GHRs are expressed in the two-cell embryo, and GH in the blastocyst.129 GH-cultured blastocysts contain more blastomeres.130 GH mRNA is expressed consistently between the two-cell and blastocyst stages;131 hence, in these stages, local GH may be acting in an autocrine or a paracrine manner. Paracrine GH has been implicated in the enhanced growth of two-day embryos, since GH antibodies retard embryonic growth.130 Of the two major cell populations in the blastocyst, GH appears to directly target the trophoblasts rather than the inner cell mass,132 despite that GHRs are present in both cell types.133 In turn, IGF-I most likely acts on the inner cell mass, independent of GH.132
For a successful embryonic implantation and placental formation, there is the need of maternal modifications, as it occurs in the embryo. In this regard, it has been shown that two-cell embryos incubated to the blastocyst stage in the presence of GH show higher levels of in vivo implantation.130 This is because the early blastocyst needs to escape from the zona pellucida that surrounds it, a protective matrix composed of glycoproteins and carbohydrates, before it can implant into the endometrium. GH improves the rate of this escaping in bovine,134 and murine,135 blastocyst populations. Therefore, GH facilitates implantation, perhaps by increasing the production of matrix metalloproteinases and also by selectively stimulating trophoblast cell proliferation, since these cells participate in blastocyst cavity formation and invasion of the maternal endometrium.132 Moreover, placental GH stimulates the invasive activity of these trophoblasts, and GHR expression is correlated with the degree of invasiveness. An autocrine/paracrine interaction is possible, since invasive extravillous cytotrophoblast cells express GH-V mRNA and secrete placental GH.136
Together with its effects on the zygote and blastocyst, GH also facilitates the development of the most appropriate maternal environment. This has to be initiated very early in gestation.
GH and GHR are expressed in pregnant and nonpregnant uteri, and GHR is differentially regulated during pregnancy and the menstrual cycle. GHR is expressed in glandular cells of the human endometrium and decidua (but not stromal cells) during the mid and late luteal phases but not during the proliferative or early luteal phases.137
Both endometrial gland development and secretory capacity are stimulated by the hormone which, furthermore, increases the amniotic glucose concentration and endometrial protein synthesis. This suggests a positive effect on nutrient transfer to the embryo. In addition, GH promotes the growth of the uterus, according to the fact that pregnancy maintenance requires significant uterine hypertrophy. In fact, GHD women have smaller uteruses than non-GHD women, most likely reflecting that a normal GH axis may be necessary for estrogen-induced uterine hypertrophy, since uterine GHR mRNA abundance is strongly correlated with estrogen-induced uterine growth.138 In summary, although most of the GH effects described in this section proceed from studies carried out in bovine and murine species, it is likely that similar effects occur in human beings and GH actions already begin in the zygote and continue in preimplantation embryos by stimulating glucose uptake and glycogen utilization. This would provide the needed production of energy for the high rate of cellular proliferation in this stage.129,139
On this basis, it would be feasible to affirm that during the preimplantation period, growth is controlled, directly or indirectly, by GH and IGF-I. It is not known whether IGF-I expression during this period is dependent on GH, but in uterus it is not.
Once implantation occurred and the placenta commences to develop, placental GH genes begin to be expressed. Among them, giving its similitude with the pituitary GH-N, GH-V protein seems to play the most important role. Placental GH appears to be secreted selectively into the maternal compartment.140 Therefore, a progressive increase in maternal plasma levels of placental GH starts from 8 to 10 weeks of pregnancy,141 leading to a clear increase in maternal plasma levels of IGFs. The negative feedback that GH exerts on its own pituitary secretion (via simulating hypothalamic somatostatin release),39 added to the inhibitory effect that IGF-I also exerts on pituitary GH release, both directly at the pituitary level and by stimulating somatostatin,39 leads to a progressive decrease in maternal plasma levels of GH-N, parallel to the increase in GH-V concentrations. Moreover, placental GH-V, different to GH-N secretion, is not pulsatile;142 thus, the continuous negative feedback exerted by GH-V will completely block maternal GH-N release, but this does not seem to affect fetal growth. For a better understanding of these concepts, see Figure 4.
Figure 4.
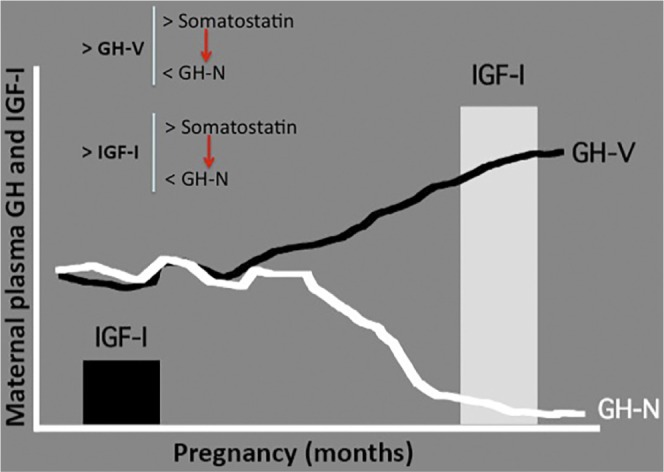
Evolution of maternal plasma levels of GH (N and V) and IGF-I during pregnancy. As pregnancy progresses, placental GH-V increases in the maternal blood. This leads to increased IGF-I levels and decreased GH-N levels. The decrease of plasma GH-N is due to the negative feedback that GH-V and IGF-I directly exert on pituitary GH-N secretion and also due to the fact that both GH-V and IGF-I stimulate hypothalamic somatostatin secretion that, in turn, negatively controls pituitary GH-N release. The maternal increase in plasma levels of IGF-II is not shown in the figure. Red arrows indicate negative control.
Fetal growth is not affected if pituitary maternal GHD exists;143 however, intrauterine growth retardation appears when maternal plasma levels of IGF-I are low or placental GHs production is deficient.144 In the fetal pituitary, GH is detected at week 10 of gestation, and the hormone begins to be released to fetal plasma two weeks later,145 peaking during weeks 20–24 and then declining. GHR expression in fetal liver and other tissues parallels the pattern of GH secretion. However, fetal GH production does not seem to have a significant impact on fetal growth, since, for instance, the uterine growth of anencephalic fetuses is not different from that in normal fetuses, despite the absence of the hypothalamo–hypophyseal axis.146 Currently, it is thought that the fetal GH accomplishes an important developmental programming role in virtually all tissues and organs,147 but does not act as a fetal “growth hormone”.
In fact, three mouse strains (Ames, Jackson, and Snell dwarf mice), all with spontaneously occurring autosomal recessive mutations, have a normal size at birth, despite all having pituitary hypoplasia, and GH synthesis cannot be detected in their pituitary glands.148
Going back to the role that GH-V may play in the maternal blood, it is important to know that its somatogenic properties are stronger than that of GH-N. It would explain its effects on maternal IGFs which, in turn, would play a critical role for the supply of nutrients to the fetus, particularly glucose and amino acids. Increased fetal glycemia would lead to increased release of insulin from the fetal pancreas. Then, insulin, directly or through an IGFs-mediated action would be the hormone responsible for the fetal growth, that is, insulin would be the real “growth hormone”. In support to this concept gestational diabetes usually leads to the birth of macrosomic children, as it occurs when nesidioblastosis (fetal hyperinsulinism due to hyperplasia in pancreatic beta cells) exists.
These concepts are schematically depicted in Figure 5.
Figure 5.

Fetal growth. (1) Placental GHs, particularly GH-V, are progressively increasing in maternal blood inducing an increase in plasma levels of glucose. This, and presumably (2) the effect of GH-V on liver, increases the hepatic production of IGFs, which optimizes the supply of nutrients to the fetus (3), particularly glucose. The resulting increase in fetal glycemia (4) induces increased release of fetal pancreatic insulin, and the hepatic production of IGFs, which also can be induced by insulin. Despite pituitary fetal GH-N secretion, it is unlikely (4?) that it contributes to fetal hyperglycemia. For its part, the placenta significantly contributes to the increase in fetal plasma concentrations of IGFs by producing IGF-II. Fetal GH most likely plays a role on the developmental programming of virtually all tissues and organs. (5) Insulin, mainly, and IGFs are the hormones that the fetus needs for growing.
Abbreviation: +, stimulates.
Despite the known role of IGF-I during this stage of development, IGF-II is the most abundant fetal IGF. It acts as a paracrine growth regulator synthesized by multiple fetal tissues, independent of GH-N.149 Effects of IGFs on fetal growth are additive, as deletion of the receptor for IGF-I, through which both IGFs act, reduces birth weight to a greater extent than deletion of either IGF-I or IGF-II genes alone.150 IGF-II is thought to provide the stimulus for fetoplacental growth, while, as described before, IGF-I appears to regulate fetal growth in response to nutrient availability. IGF-II increases in maternal plasma during pregnancy and declines toward term. In all species studied, the placenta synthesizes IGF-II, at least in early pregnancy, being produced by the syncytiotrophoblast layer in direct contact with the maternal blood.151,152 Curiously, in pregnant women, IGF-II is secreted as a biologically inactive pro-peptide, which undergoes proteolytic cleavage to become active; this proteolytic activation increases during pregnancy and correlates with maternal IGF-II plasma concentrations.153,154 In addition to IGF-I and insulin receptors (InsR), the placenta expresses an hybrid receptor IGF1R–InsR; in women, these receptors are localized to the syncytiotrophoblast layer bathed in maternal blood;152 this permits endocrine regulation of placental growth and function by maternal IGFs.
In summary, IGFs in the maternal circulation are important regulators of fetal growth via their actions on both the mother and the placenta. They influence maternal tissue growth and metabolism and, thereby, modulate nutrient availability for conceptus growth. They also regulate placental morphogenesis, substrate transport, and hormone secretion, which influence fetal growth either via indirect effects on maternal substrate availability, or through direct impacts on fetal nutrient supply and its endocrine environment.
Maternal IGF-I and IGF-II may have complementary, but overlapping, roles in optimizing fetal nutrient acquisition for growth and survival. Maternal IGF-I appears to act predominantly on maternal tissues to influence substrate availability, whereas maternal IGF-II acts on the placenta to influence substrate delivery to the fetus. In turn, this increased supply of nutrients, particularly glucose, positively regulates expression of IGFs in fetal liver.55
According to these extensive data, growth before birth depends on a number of hormonal and nutritional factors. Among them, GH seems to play a key role in the early stages of development, since the zygote is formed until the blastocyst is implanted in the uterus. Then, when the placenta begins to develop and placental GHs are secreted, they act mainly on maternal increase of IGFs; these contribute to the further development of the placenta and also act on maternal tissues for improving the supply of nutrients to the fetus. Therefore, plasma glucose increases in the fetus leading to increased insulin release that, in addition to glucose, stimulates the liver production of IGFs. Insulin and IGFs would then be the main regulators of fetal growth as pregnancy progresses.
Growth during the early infancy
Nutrition plays a key role for growing during the first months of postnatal growth, even when pituitary GH secretion is normal. After birth, growth continues at the same rate as is seen in utero, despite the onset of pulsatile release of GH occurs soon after birth. This high rate of growth occurs until 6–10 months of age, after which growth slows. Therefore, while growth velocity in utero follows an exponential curve, it changes to a double exponential curve during the first year of life (Fig. 6). This is indicative that two kinds of different factors influence growth velocity during this first year of life: nutritional factors (again, the glucose–IGF-I–insulin axis seem to play the role of a “growth hormone”) and GH–IGF-I. Therefore, during this first period of life, GH does not seem to play a significant role on the longitudinal growth of the organism. Thus, when a GHD exists in neonates, growth failure is usually detected by 6–10 months of age. However, there are also data indicating that congenital GHD may present with typical clinical signs and symptoms (for instance, hypoglycemia) very early in postnatal life, among them is decreased growth velocity.155,156 It would be interesting to know if the decreased growth velocity observed in these congenital GHD children depends on the absence of GH secretion, or on the lack of induction by glucose of the liver production of IGF-I.
Figure 6.
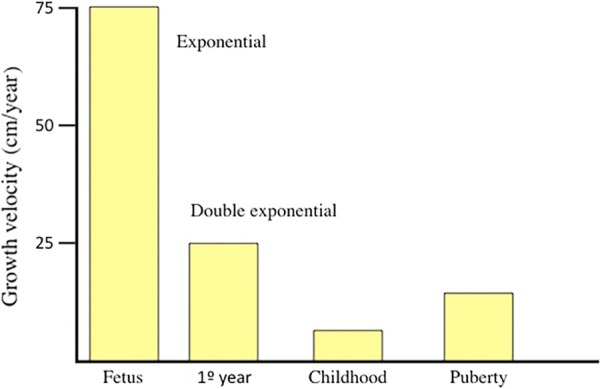
Growth velocity along life. The maximum growth velocity is observed during the fetal life, reaching 75 cm/year and following an exponential curve. After birth, growth velocity follows a double exponential curve; the consequence is that the high growth velocity observed after birth progressively slows since 6–10 months of age, indicating that two dfferent kinds of factors are acting in this period. During childhood, growth velocity (lineal now) decreases to 5 cm/year, and when puberty begins, a number of factors (mainly sex steroids) acting on pituitary GH synthesis and release again increase growth velocity until 10–12 cm/year.
Growth during childhood
The impact of GH on growth increases over the first years, and approximately from the age of 3 years until puberty, GH and thyroxine predominate as the major influences on growth, leading to normal growth velocities of 5 cm/year (Fig. 6). Thus, GH is the growth hormone during this stage of life, but as described before, its effects on the longitudinal growth are dependent on hepatic or local (chondrocytes) IGF-I production, although the possibility exists that GH may act on growth plate chondrocytes by an IGF-independent mechanism.41,57
Vertebral growth mainly depends on sex steroids.
Growth at puberty
As puberty approaches, growth velocity increases, until reaching values of 10–12 cm/year (Fig. 6). This is a consequence of increased GH secretion and the associated IGF-I increase. The reasons for these changes are due to the effects of sex steroids on the hypothalamic–pituitary axis; sex steroids act on catecholamine pathways that negatively act on somatostatin secretion and facilitate hypothalamic GHRH release.39 However, this increased growth velocity lasts for a short time, because the own sex steroids (estradiol acting directly on growth plates and producing its ossification, while testosterone is locally transformed in estradiol by the action of an aromatase) lead to the epiphyseal fusion once puberty concludes. Longitudinal growth then ends.
However, and despite that no longitudinal growth already exists, GH secretion continues, although an exponential decline in 24-hour plasma GH concentrations can be seen in both sexes, starting from 18 to 30 years of age. It is a curious event, since sex steroids secretion continues, but GH release progressively declines until almost undetectable values while the subject is aging (from 60 years old, more or less). The fact that GH secretion is not brusquely interrupted, once puberty ends, indicates that the hormone plays other important roles in the organism, basically related, but not only, to its metabolic actions, and suggesting that GH is not “the growth hormone”, although in some periods of the life, as described, its role on growth is important.
Other effects of GH
Besides its actions on metabolism, GH as a pleiotropic hormone plays a number of endo-, auto-, and paracrine actions on practically any tissue or organ in the organism. We will try to summarize most of these actions with a particular emphasis on its actions to nervous level (central and peripheral).
GH and liver
The liver is an important target of the actions of GH. Mice with liver-specific knockouts for critical GH signaling proteins, GH receptor (GHR), Janus kinase (JAK) 2, and Stat5 share a common phenotype of hepatic steatosis,157–159 demonstrating that GH plays an important physiological role in hepatic triglyceride secretion. Steatosis leads to hepatic degeneration, which may be corrected by GH administration. Even more, a high prevalence of liver dysfunction has been reported recently in adult GHD patients.160 GH replacement therapy significantly reduced serum liver enzyme concentrations in these patients and improved the histological changes in the liver.160 However, the liver produces its own factor of regeneration: hepatocyte growth factor (HGF) was first identified in the sera of 70% hepatectomized rats, as a mitogen of adult rat hepatocytes.161,162 Indeed, during hepatic injuries, blood and liver HGF levels markedly increase, via both extrahepatic and intrahepatic pathways.163–165 Animal studies using either anti-HGF antibody or c-Met gene destruction techniques revealed that both the endocrine and paracrine effects of HGF are involved in liver growth after 70% hepatectomy, and for recovery from hepatitis, respectively.163–166 HGF has a positive regenerative and protective effect in numerous organs and diseases.167,168 In spite of its liver production and its strong liver regenerative properties, it was found that in hypophysectomized rats treated with GH, HGF mRNA levels were increased three hours after partial hepatectomy and reached peak levels after five hours. In rats with intact pituitaries and in hypophysectomized rats not given GH treatment, HGF mRNA levels in liver were unchanged during the first 5 hours following hepatectomy and reached peak levels after 10–18 hours. DNA synthesis in the liver of GH-treated rats increased from low levels, 10 hours after hepatectomy, to peak levels, after 18 hours. In rats without GH treatment, the synthesis of DNA was still low, 18 hours after hepatectomy, and was increased, after 26 hours. HGF mRNA levels were constantly lower after sham hepatectomy than after partial hepatectomy. In summary, in hypophysectomized rats, the responses of hepatic HGF gene expression and DNA synthesis to partial hepatectomy were accelerated by treatment with GH.169 Whether GH stimulates the transcription of HGF or facilitates it is not known, but we found that GH is expressed in the liver of hypophysectomized rats subjected to partial hepatectomy (Fig. 7) and that this GH promotes the hepatic regeneration, directly or via HGF induction. Moreover, the analysis of the products obtained with the enzyme of restriction RsaI demonstrated that the hepatic GH gives origin to two bands in the expected molecular weight position (238 and 90 bp), identical to the bands obtained from pituitary rat GH (Fig. 7).19 From these data, it is clear that there is a hepatic expression of GH that contributes to, or determines, the high degree of regenerating ability of the liver, apart from playing important metabolic functions in this organ.
Figure 7.

Hepatic expression of GH. GH mRNA from the pituitary and liver (partially hepatectomized rats) was retrotranscripted with specific primers and the resultant cDNAs were resolved in 2% agarose and stained with ethidium bromide, before and after using the enzyme of restriction RsaI. As the figure shows pituitary (pit) and liver (liv), GH was detected with the expected molecular weight: 328 bp, because the primers used flanked a region situated between exons 4 and 5 of rat GH gene. The bands obtained after cutting the main GH amplified with RsaI led to the appearance of two bands in the molecular weight expected (238 and 90 bp), both in pituitary and liver GH. Black arrows indicate the main GH product. Blue arrows indicate the products obtained after cutting with RsaI. Reprinted with permission of the publisher. Source: J Devesa, Devesa P, Reimunde P. Growth hormone: actions and preventive and therapeutic applications. Med Clin (Barc) 2010; 135 (14): 665-670. Copyright © 2009 Elsevier Spain, S.L. All rights reserved.
Abbreviations: MWM, molecular weight markers; bp, basepairs.
GH and adrenal glands
It has been shown that GH and IGF-I enhance steroidogenesis responsiveness to ACTH in cultured adrenal cells and that adrenal steroid responsiveness to ACTH increases in Turner syndrome after long-term treatment with high GH doses.170 GH is an important modulator of the activity of 11β-hydroxysteroid dehydrogenase type 1 enzyme in the adrenal gland,171 as indicated by the fact that plasma DHEAS levels are significantly lower in GHD patients (even in the patients with normal ACTH secretion) than in age-matched controls. GH replacement therapy in these GHD patients significantly increases DHEAS plasma levels. This suggests that, in the permissive presence of ACTH, GH stimulates adrenal androgen secretion.
GHR is strongly expressed in the ovine fetal adrenal gland.172 However, in normal subjects or laboratory animals, the stimulation of adrenal steroidogenesis by GH seems to be restricted to the fetal period.173
In a previous study, in rats, we demonstrated that the compensatory adrenal hypertrophy that follows a unilateral adrenalectomy seems to be mediated by adrenal GH expression. In that study, one adrenal gland was surgically removed and weighed, and 24 hours later, the contralateral gland was removed and weighed, and mRNA GH was extracted from both glands for cDNA RTPCR. Our results show that the contralateral gland had increased its weight and GH expression was detected in 2% agarose gel stained with ethidium bromide, while basal GH expression in the control gland was very low, as shown in Figure 8.19
Figure 8.

Adrenal expression of GH in rats. GH mRNA from an adrenal removed (control) and from the contralateral adrenal gland (24 hours after unilateral adrenalectomy) was retrotranscripted with specific primers, and the resultant cDNAs were resolved in 2% agarose and stained with ethidium bromide. GH products were detected in the expected molecular weight: 328 bp. Adrenalectomy markedly increased the expression of GH in the contralateral gland 24 hours after the adrenalectomy (Ax 24 h). This may indicate that, apart from ACTH action, GH is overexpressed in the adrenal gland for mediating the compensatory hypertrophy. Reprinted with permission of the publisher. Source: J Devesa, Devesa P, Reimunde P. Growth hormone: actions and preventive and therapeutic applications. Med Clin (Barc) 2010; 135 (14): 665-670. Copyright © 2009 Elsevier Spain, S.L. All rights reserved.
Abbreviation: MWM, molecular weight markers.
These results indicate that apart from the putative effects of GH on adrenal steroidogenesis, the hormone may also play a trophic regenerative role (as in liver) on the adrenal glands.
GH and the cardiovascular system
GH acts directly on myocardial growth and heart function during the fetal development.174 GH (or myocardial GH-induced IGF-I) induces mRNA expression for specific contractile proteins and cardiomyocyte hypertrophy. In addition, and perhaps even most important, GH increases the force of contraction and leads to myosin phenoconversion toward the low ATPase activity V3 isoform.174 The prevalence of V3 isoform increases the number of actin–myosin cross-bridges and their attachment time, enhances protein calcium sensitivity and calcium availability, and allows the myocardium to function at lower energy cost. In these conditions, the fetal heart is able to beat at high frequency without spending too much energy. After birth, this changes, and a hypertrophy and myocardial remodelation occurs; the V1 myosin is then expressed, which implies a higher ATPase activity. However, if an excess of GH exists, such as that which occurs in acromegaly, the fetal genetic pattern reappears because the V3 isoform is then mostly expressed again; this could lead to a myocardial insufficiency.
The GH–IGF-1 axis may also regulate cardiac metabolism, by increasing amino acid uptake, protein synthesis, cardiomyocyte size, and muscle-specific gene expression. GH increases the collagen deposition rate in the heart,175–179 while GH-induced IGF-1 influences the trophic status of myocardium by reducing apoptosis of cardiomyocytes, thus preventing myocyte loss.175–181
The important role that GH plays at the myocardial level can easily be deducted by analyzing what happens in untreated GHD children. They show cardiac atrophy with a significant reduction in the left ventricle mass, relative wall thickness, and cavity dimensions, compared with age-, sex-, and height-matched controls.182–186 Moreover, patients have a low ejection fraction, low cardiac output, and high peripheral vascular resistance.182,184–187 These alterations are more pronounced during physical exercise; they reduce exercise intensity, and duration is reduced in these children. In turn, adult-onset GHD does not produce a reduction in cardiac mass, but cardiac performance and exercise capacity are impaired.188–190
GH replacement trials show an increase in left ventricular mass and improvement in cardiac performance, diastolic filling, and systolic function after GH treatment is given to these children or adults.182–184,187,190 As a consequence of its positive effects on heart, it may be speculated that GH treatment might be useful in patients with heart failure, mainly if they are GHDs or GH secretion is insufficient.
Apart from its direct or IGF-I-mediated GH effects on heart, the hormone also acts at the vascular level.
In vascular system, the GH–IGF-1 axis activates the nitric oxide (NO) system and regulates nonendothelial-dependent actions.191–196 NO production relaxes arterial smooth muscle cells, thereby reducing vascular tone. Furthermore, NO inhibits proliferation and migration of smooth muscle cells, reduces platelet adhesion, and decreases lipoxygenase activity and oxidized LDL-cholesterol.191–196
Another possible mechanism for the GH/IGF-I influence on vascular tone involves the regulation of gene expression of the vascular smooth muscle KATP channel.197
Some vasoactive effects of GH may have central origin. In fact, GHD patients have markedly increased muscle sympathetic nerve activity.198 Instead, one year of substitution therapy with GH had a significant effect on decreasing sympathetic nerve activity to the muscle vascular bed.199 This suggests that GH may regulate the central sympathetic outflow, affecting peripheral resistance.
Many evidences indicate that GH plays a significant role on angiogenesis, contributing to regulate vascular growth and function. GHR has been found in many different blood vessels,200–203 and in cultured endothelial cells,204,205 where GH stimulates endothelial cell proliferation206,207 and tube formation.208,209 Treatment with GH increases the number of cerebral cortical arterioles in aging rats,210 augments VEGF expression and angiogenesis in the rat myocardium after infarction,211,212 stimulates wound angiogenesis in diabetic rats213 and mice,214 and enhances vascularization by mobilizing bone marrow-derived endothelial progenitor cells into the blood.215,216 In agreement with these findings, the skin of adult GHD patients shows reduced capillary density and permeability that improves after treatment with GH,217 and GHD adults and children have reduced retinal vascularization.218,219 As in other territories, IGF-I may mediate the proangiogenic actions of GH, as IGF-I receptors are widely expressed in endothelial cells, and IGF-I has been shown to stimulate angiogenesis in vivo and in vitro (for review, see Ref. 220). However, some actions of GH on endothelial cells may be independent of IGF-I, since GH is unable to increase the transcription of IGF-I in endothelial cells. In addition, systemic221 or local infusions of GH222 acutely increase forearm blood flow and NO release in healthy human beings without significantly raising plasma IGF-I levels or muscle IGF-I mRNA expression. While these observations argue in favor of an autonomous GH action mediated by NO, IGF-I is also vasoactive due to its activation of eNOS.
In addition, GH may have context-dependent vascular actions influenced by other angiogenic agents, including IGF-I, NO, and VEGF, and GH itself. GH is produced by endothelial cells, and endothelium-derived GH stimulates the proliferation, migration, survival, and capillary formation of endothelial cells in an autocrine manner.208 Therefore, the lack of action on angiogenesis, sometimes observed, of exogenous GH may relate to endothelial cell GHRs being already occupied by the endogenous hormone. Other important factors affecting GH actions on angiogenesis include its local proteolysis to vasoinhibins, which inhibit blood vessel dilation, permeability, growth of new vessels, and survival of newly formed. Finally, human GH has the ability to activate PRL receptors, which can also mediate proangiogenic signals.
Despite these proangiogenic effects, increased GH levels are not always related to angiogenesis. The best example are GHD patients (children or adults) receiving GH treatment for long term: the risk of retinopathy is not increased in them.219
For a more detailed comprehension of GH effects on the cardiovascular system, see Refs. 223–225.
GH and the adipose tissue
A typical finding in untreated GHD children and adults is the increase in fat mass,226–227 preferentially visceral fat, which is mainly due to the fact that GH stimulates lipolysis by increasing lipase activity,228 although changes in the secretion of certain adipokines, such as adiponectin, have also been suggested as mediators of the increased adiposity in GHD states.229 The administration of GH replacement therapy reverts this increased adiposity,226,227 therefore confirming the relationship between GHD and increased fat mass. Among other factors, since GH secretion is decreasing from puberty to elderly, it is likely to speculate that the progressive increase in fat stores usually seen along aging might be due to the deficitary or insufficient GH secretion.
Apart from this metabolic effect, GH plays a role in pre-adipocyte proliferation, differentiation, and senescence.230
GH and skeletal muscle
The GH–IGF-1 axis constitutes an important physiological regulatory mechanism for coordinating postnatal skeletal muscle expansion and hypertrophy. Administration of GH to both animals and GHD human beings improves muscle strength and reduces body fat.231–233 A number of studies have shown that mice globally deficient in GHR have reduced muscle mass with defective myofiber specification and growth.234 This seems to be logic, given the anabolic effects of GH; however, during a time it has been discussed if this GH effect would be restricted to inducing enhanced uptake of amino acids by muscle, while the effects on muscle protein synthesis would be dependent on GH-induced IGF-I expression, mediated by STAT5b. In fact, recent in vitro studies show that treatment of primary myoblasts with GH quickly increases IGF-I mRNA, while administration of IGF-I leads to a significant increase in primary myoblasts proliferation.235 Therefore, the role of GH on muscle would be dependent on its induction of production of IGF-I by myoblasts, and IGF-I would then be responsible for stimulating myoblasts proliferation in an autocrine manner. However, GH affects myofiber development, in a mechanism independent of IGF-I, by stimulating accumulation of additional myonuclei into nascent myotubes; however, the increased myoblast fusion is an IGF-I effect, although mediated by IGF-I expression induced by GH.235 The disruption of GHR in skeletal muscle and the consequent histomorphometric changes in myofiber type and size and myonuclei number result in functionally impaired skeletal muscle. In agreement with these effects, the histology of muscles of untreated GHD patients is strongly altered, and glucose and triglyceride uptake and metabolism in skeletal muscle of GHR mutant mice are affected.
A recent study demonstrates that a single bolus of GH induces gene expression of regulators of substrate metabolism and cellular growth in human skeletal muscle in vivo. While some of these genes, for instance GISH gene, seem to be directly induced by GH, others, like ANGPTL4 gene,236 seem to be expressed in relation to the subsequent increase in free fatty acid levels induced by GH-dependent lipolysis. These results agree with the role that GH plays on lipid metabolism.
With regard to the putative effects of GH on muscle strength, GH use has been speculated to improve physical capacity in subjects without GHD through stimulation of collagen synthesis in the tendon and skeletal muscle, which leads to better exercise training and increased muscle strength. In this context, the use of GH in healthy elderly should be an option for increasing muscle strength. However, a clinical trial showed that after six months of therapy, muscle strength in the bench press responsive muscles did not increase in groups treated with GH (no GHD) or placebo and showed a statistically significant increase in the leg press responsive muscles in the GH group. The study demonstrated an increase in muscle strength only in the lower body part (quadriceps, for instance) after GH therapy in healthy men.237 Therefore, GH administration does not provide significant improvements in increasing muscle power, except when GHD exists.
Of interest, sarcopenia appears while aging or after a prolonged immobilization. Although most likely this is a multifactorial process, a predominant role is played by myostatin, a muscular hormone that inhibits cell cycle progression and reduces levels of myogenic regulatory factors, thereby controlling myoblastic proliferation and differentiation during developmental myogenesis, as we and others demonstrated.238–240 GH-induced muscular expression of the IGF-I-Akt-mTOR (mammalian target of rapamycin) pathway, which mediates both differentiation in myoblasts and hypertrophy in myotubes, has been shown to inhibit myostatin-dependent signaling. Blockade of the Akt-mTOR pathway, using siRNA to RAPTOR, a component of TORC1 (TOR signaling complex 1), facilitates myostatin’s inhibition of muscle differentiation because of an increase in Smad2 phosphorylation.241 Therefore, GH administration in these conditions of muscle wasting may be useful for recovering muscle mass at expenses of inhibiting myostatin signaling.
GH and bone
The actions of the GH–IGF-I axis at the growth plate to promote longitudinal growth have already been described. However, this axis also regulates skeletal development and mineral acquisition.242 Mouse models with disruptions of GH–IGF-I axis show a clear deterioration in parameters of bone health, dependent on GH-induced IGF-I expression, which increases bone mineral density.243–245 Apart from GH, other GH-independent mechanisms regulate bone IGF-I expression, for instance parathormone (PTH).246 Experimental mouse models reveal that osteoblast-derived IGF-I is a key determinant of bone mineralization. Targeted osteoblast-specific overexpression of Igf1 via the osteocalcin promoter produced a phenotype of increased bone mineral density and trabecular bone volume,247 whereas knockout of the gene in bone (and muscle) but not liver via Cre recombinase expressed by the collagen type 1α2 promoter included a phenotype of reduced bone accretion.248
GH and kidney
GH affects renal function and kidney growth. GH, acting via IGF-I, increases glomerular filtration rate (GFR) and renal plasma flow (RPF) in GHD patients as well as in normal adults. Furthermore, GFR and RPF are low in hypopituitarism and elevated in acromegaly. IGF-I is implicated in compensatory renal hypertrophy after unilateral nephrectomy or ischemic renal degeneration.249 Disordered regulation of the IGF system has been implicated in a number of kidney diseases. IGF activity is enhanced in early diabetic nephropathy and polycystic kidneys, whereas IGF resistance is found in chronic kidney failure. IGFs have a potential role in enhancing stem cell repair after a kidney injury.250 Importantly, children with chronic kidney disease show growth failure that can be safely treated with GH, as a recent clinical trial demonstrated.251 Regarding other renal effects, GH increases sodium retention and increases plasma bicarbonate concentration; therefore, it seems to be useful for the treatment of metabolic acidosis.252
GH and the hematopoietic system
Several reports suggest a role of GH in the regulation of the hematopoietic system: normal differentiation and function of blood cells. An intriguing study shows that after one year of GH treatment in non-GHD children, erythropoietin (Epo) plasma levels significantly decreased and granulocyte-colony stimulating factor (G-CSF) levels increased from basal to 12 months of therapy, whereas in GHD children, they did not change significantly. Circulating levels of G-CSF are significantly lower in GHD than in non-GHD children. In non-GHD children, the number of red blood cells, hemoglobin (Hb), and hematocrit values significantly increased after one year of GH treatment. GH therapy influences Epo and G-CSF levels in short non-GHD children, while it shows no effects in GHD children.253 These findings need further clarification, since GH increases plasma Epo levels and Hb in adult GHD patients,254 and increases plasma G-CSF levels and neutrophil counts in adult GHD patients.255 Another study carried out in GHD patients treated with GH showed that the replacement therapy exerted a beneficial effect leading to a significant increase of erythrocyte parameters and recovery from anemia (typical of GHD patients during childhood), without affecting the number of leukocytes or platelets.256 In all, these data indicate that GH exerts a positive role on the hematopoietic system, similar to that played by G-CSF.257
In recent years, it has been proposed that GH has a strong influence on the immune system. The production and action of immune cell-derived GH is now well known, although its important role in immunity is still emerging. A number of data reveal the production of GH, GHRH, and IGF-I and its receptor on cells of the immune system and their influence via endocrine/autocrine/paracrine and intracrine pathways on immune function.258 The intracellular mechanisms of action of immune cell-derived GH are not well known, but, for instance, GH promotes the maturation and activation of dendritic cells that, as antigen-presenting cells, participate in the immune response of the organism.259
Throughout many years, Weigent et al published data indicating that GH produced in lymphocytes plays a role for lymphocyte growth, survival, and production of cytokines.260–269 Lymphocyte GH appears to stimulate IFNγ production with a small positive effect on IL-10 production.269 Both norepinephrine and cortisol inhibit lymphocyte GH production, and the studies suggest that lymphocyte GH may be an important mediator of cellular immune function mediated by the TH-1 pathway.269 Treatment of rat lymphocytes with a specific GH antisense oligodeoxynucleotide decreased the amount of lymphocyte GH synthesized and, at the same time, reduced lymphocyte proliferation.260 By using neutralizing antibodies to GH, the number of cells positive for IGF-I decreased twofold, supporting an important role for endogenously produced GH in the induction of lymphocyte-derived IGF-I.261 The small amounts of GH synthesized and secreted by cells of the immune system,270 together with data showing that the same cells that synthesize GH also produce IGF-I,271 suggest that GH may be classified as an intracrine hormone, acting primarily intracellularly.272 More recently, it has been shown that overexpression of GH in a lymphoid cell line, devoid of the GHR, decreases the production of superoxide and increases the production of nitric oxide and the expression of IGF-I and IGF-IR, resulting in protection from apoptosis by a mechanism most likely involving an increase in the production of BcL-2.262–265
These results, together with other data, suggest that a complex intracrine/autocrine regulatory circuit may be important for the production and function of leukocyte-derived GH and IGF-I within the immune system. Therefore, this circuit could fulfill local tissue needs for these hormones independent of the pituitary or liver without disrupting homeostasis of other organ systems. For example, cells of the immune system would recognize the association of bacteria, virus, and tumors as an oxidative stress event and signal the release and transport of a lower molecular weight isoform of GH into the nucleus. Once in the nucleus, GH would be free to influence transcriptional responses to the stress event and to defend the cell against oxidative damage. The results provided by Weigent273 support the concept that changes in the cellular redox status influence the intracellular levels of lymphocyte GH, which may exert effects on elements mediating the oxidative stress response.
Interestingly, in his study,273 Weigent provided evidence that high molecular weight forms of GH (100 K, 65 K, and 48 kDa isoforms) are present in cells of the immune system. However, the main 22 kDa GH was not detected. Under normal culture conditions, lymphocyte GH exists in the cytoplasm primarily as an isoform of approximately 100 kDa and in the nucleus as an isoform of approximately 48 kDa (EL4 cells) or 65 kDa (primary spleen cells). That is, GH protein isoforms appear to reside in different compartments; however, the actual site(s) at which it exerts its function and the physiological significance of these isoforms remain unknown. However, it is likely that they represent an important regulatory function in mediating intracellular GH activity. This may be supported by the fact that higher levels of the cytoplasmic 100 kDa isoform and particularly the nuclear 65 kDa isoforms of GH were found in B-cells compared to T-cells. Moreover, data from this and other studies suggest the possibility that the GHR and/or the GH-binding protein (GHBP) may serve particular roles in the intracrine actions of lymphocyte-derived GH in cells of the immune system.
GH and the nervous system
Since the pioneer discovery of the presence of the GHR in cerebral areas known to be actively involved in neurogenesis within the juvenile brain, including the subgranular zone (SGZ) of the dentate gyrus and subventricular zone (SVZ), by Lobie et al,274 a number of studies during the last years revealed that GH exerts many different and positive effects on the central and peripheral nervous system. GH immunoreactivity has been detected in the first stages of neural development,275 in accordance with the key roles that both, GH and IGF-I, play in the early brain development, maturation, and function.276 Despite GH-binding sites have been found in the choroid plexus, indicating that they may act as carriers for introducing plasma GH into the CNS,277 they have also been found in a number of brain structures, such as hippocampus, putamen, and thalamus.277 This is consistent with the finding that GH is also synthesized in the CNS and is regulated differentially to its hypophyseal counterpart. IGF-I is synthesized in the CNS, and in the early postnatal period it is regulated by peripherally secreted GH.278 However, IGF-I expression has also been detected in neural stem cells (NSCs) obtained from fetal human forebrains induced by exogenously administered GH.279 Both GH and IGF-I alter the size and morphology of the CNS during development and affect differentiated cell function in the CNS, with consequent modulation of cognitive function. Differential utilization of the same signal transduction molecules indicates that GH and IGF-I possess distinct overlapping roles in CNS function.278 Apart from its role during fetal development, GH plays a significant modulatory function on brain and seems to play a key role for brain repair after an injury.
The hypothesis that GH and IGF-I play a role on brain repair after an injury has been postulated years ago.280 This role may be played by inducing neurogenesis in more brain areas than previously established (SGZ and SVZ). As described, GHR is expressed in regions of the brain in which neurogenesis occurs during embryonic brain development281,282 and in neurogenic regions of the postnatal rat brain.274 GH itself is also found in cells of the ventricular zone during embryonic neurogenesis282 and is produced endogenously within the postnatal hippocampus (Fig. 9).283–286
Figure 9.

GH and GHR are expressed in NSCs from mice. NSCs obtained from nine-day-old mice cultured in proliferation media, free of GH, form neurospheres in which confocal microscopy immunoreactivity for SOX2 in the nuclei indicate that these cells are in an early phase of self-renewal. These cells show high immunoreactivity (green labeling) for GH (left) and its receptor (right), indicating that both GH and GHR are expressed in NSCs. Magnification: 40×. © 2012 Devesa J, Devesa P, Reimunde P, Arce V. Published in Growth Hormone and Kynesitherapy for Brain Injury Recovery343 under CC BY 3.0 license. Available from: http://dx.doi.org/10.5772/26998.
Abbreviation: NSCs, neural stem cells.
GH gene expression within the hippocampus is increased by some factors known to increase neurogenesis,287 including learning.283 Studies of the effects of GH on embryonic rat cerebral cortical,288 and hippocampal neuronal cultures of aged mice,289–290 found that it induces the proliferation and differentiation of these cells.289–291 The fact that the blockade of GHR impedes the proliferation and survival of these NSCs indicates that the effect of GH is highly specific.292
It has been demonstrated that exogenously applied GH and PRL promote the proliferation and migration of NSCs derived from fetal human forebrains in the absence of epidermal growth factor (EGF) or basic fibroblast growth factor (bFGF).279 This agrees with previous preclinical data derived by us and others, demonstrating that exogenous GH administration promotes the proliferation of hippocampal neural precursors after brain injury induced by kainate administration,291 and in a number of zones in the intact adult rat brain.293
According to these evidences, it is likely that GH may facilitate the proliferation, differentiation, survival, and migration of new neurons in response to brain injury. However, to date, only few studies in human beings explore such a possibility. While these studies indicate a positive effect for GH treatment together with specific neurorehabilitation, both in children with cerebral palsy294,295 and in TBI patients,296,297 or in a patient suffering from a neurogenic dysphagia after oncological brain surgery,298 all patients in these studies had GHD most likely occurring as a consequence of their brain damage. However, we recently demonstrated that GH administration, together with specific rehabilitation, after an important brain injury, is able to recover laboratory animals299 and patients without GHD.300,301 Similar results have been obtained after a stroke. In rats, delayed and chronic treatment of stroke with central GH may accelerate some aspects of functional recovery and improve spatial memory in the long term,302 and a pilot study in human patients who suffered a stroke showed improvements after administering GH.303 In line with these, the expression of both GH and GHR is strongly upregulated after brain injury and specifically associated with stressed neurons and glia.280,304,305 From these and other studies, it is now clear that GH plays a key role in both physiologic and reparative neurogenesis,306 being its effect specially marked on cognitive functions,307 most likely throughout the interaction of the hormone with GHRs expressed in the hippocampus and frontal cortex,308,309 brain areas especially related to memory and cognition, respectively.
It is already well known that GHD adults have impaired psychological well-being, including energy, motivation, emotion, memory, and cognition.310–312 However, all these abnormalities improved during GH replacement therapy,310,312–314 which leads to marked improvements in the quality of life.315 The same occurs in GHD after being treated with GH.316 Attention, perception, and cognitive capacity improve in them.317 Cognitive impairments and mood disturbances are common findings in patients with GH deficiency.318,319 GH treatment significantly improves memory and cognitive functions in these patients, as shown by functional MRI studies.319,320 As described before, similar results have been obtained recently by us and others, both in adult GHD patients296,297 and in patients with normal GH secretion.300,301
These effects may be due to the fact that GH may be an important regulator of hippocampus-dependent spatial learning and memory, as it is able to counteract memory deficits related to alterations in cholinergic neurons and an imbalance in hippocampal glutamatergic and GABAergic synapses.321
Several studies have indicated that GH affects cognitive functioning by enhancing excitatory synaptic transmission through NMDA receptors.308,322–324 Furthermore, the hormone might potentially affect the levels of the various receptor subunits in the NMDA receptor complex.
Several studies indicate that the neurobiological consequences of the decline in GH/IGF-I, that occurs physiologically during aging, include decreased neurogenesis in the dentate gyrus of the hippocampus,325,326 where IGF-I appears to affect primarily the survival of newborn neurons and may also influence the maturation and differentiation of newborn cells.327,328
Despite that neurogenesis is dramatically reduced during senescence, newborn granule cells in the aged dentate gyrus retain the capacity for participation in functional hippocampal networks;329 this is in agreement with the aforementioned hypothesis, suggesting that it is the age-associated lack of neurotrophic factors (GH/IGF-I?), the cause of decreased neurogenesis. In fact, GH prevents neuronal loss in the aged rat hippocampus.330
The effects of the GH–IGF-I axis on cell turnover in the adult brain probably are not limited to neuronal progenitors, since IGF-I can promote proliferation of oligodendrocyte progenitor cells and differentiation and survival of oligodendrocytes,331,332 an effect also induced by EGF,333 whose expression and that of its receptor may also be induced by GH.334 Thus, it is likely that the aging-related decline in GH/IGF-I and dependent changes in oligodendrocyte genesis and/or maturation may contribute to impaired remyelination in the CNS of aged individuals335 and to a decline in normal cognitive function.
Moreover, apart from IGF-I and EGF and its receptor, GH induces a number of neurotrophic factors that may act as neuroprotective and neuroregenerative. Among them, EPO, VEGF, FGF, BDNF, and several cytokines, as described before, facilitate the gonadal production of testosterone and estradiol.
In addition, GH affects most of the major neurotransmitters differently in several brain regions, including monoamines (serotonin, noradrenaline), the dopaminergic system, the glutamatergic system (at the receptor level), the opioid system, and the cholinergic system.
For instance, it has been shown that GH affects the monoamine levels within 15 minutes after intraperitoneal administration in rats, and its effects are different in normal and hypophysectomized rats.336 In normal rats, GH decreases the levels of both 5-hydroxyindoleacetic acid (5-HIAA) and norepinephrine in the diencephalon and brainstem, without affecting telencephalic concentrations. In hypophysectomized rats, however, GH produces significant elevations of norepinephrine and 5-HIAA levels in all brain regions.336 Similarly, bGH transgenic mice evidence increased tissue levels of serotonin and 5-HIAA acid in several brain regions.337 These two different experimental conditions show similarities with respect to the serotonin system.
GH also has been shown to affect the dopaminergic system. GH reduces dopamine (DA) release in the rat sub-ependymal layer and median eminence after intravenous administration.338 Perhaps this is a mechanism for increasing somatostatin release and therefore for reducing pituitary GH secretion.39 In line with this, bGH transgenic mice show decreased DA levels in the brain stem and decreased levels of the DA metabolite 3,4-dihydroxyphenylacetic acid in the mesencephalon and diencephalon.336 With regard to the glutamatergic receptor system, it is present throughout the brain in both neurons and astrocytes. Three major types of receptors are known, one of these being the NMDA receptor. The NMDA receptor subunits NR1 and NR2B are thought to be involved in memory formation in the hippocampus, and interestingly, GH appears to affect these transcripts. GH treatment in young adult hypophysectomized rats decreases the hippocampal mRNA expression of NR1, but increases the NR2B subunit,323 whereas in elderly rats, GH increases both the NR1 and NR2A transcripts. Similar results have been obtained with IGF-I therapy.339 The results are particularly interesting as it appears that the ratio of NR2B to NR2A mirrors the potential for synaptic plasticity.
Finally, it appears that the cholinergic system has functional links with GH and IGF-I. Functional stimulation of muscarinic transmission by the cholinesterase inhibitor pyridostigmine may enhance GH release from the pituitary under certain conditions.340,341 Moreover, IGF-I affects potassium-evoked acetylcholine (ACh) release in tissue slices of the adult hippocampus and cerebral cortex. In slices from the adult hippocampus, IGF-I decreases the release of ACh.342
In all, there are many evidences that GH and IGF-I affect several of the major neurotransmitter systems. Many of these effects act directly via neurons and can be observed after only a short exposure. However, some of these effects also interact with the other main cell types of the brain, the glial cells.
The effects of GH also take place on central and peripheral nerves. Delayed conduction from the retina to the occipital cortex is a common finding in children with cerebral palsy, but it can be corrected with GH treatment and specific visual stimulation.343 This indicates that the hormone increases the number of fibers in the optical nerve and also promotes myelination. The same happens in peripheral nerves. We and others have been able to regenerate the sciatic nerve, after its section in rats, by using GH and rehabilitation, promoting muscle reinnervation and inducing a highly proliferative state of Schwann cells responsible for remyelination.344,345 Therefore, GH is also a promising therapy for peripheral nerve injuries.
Conclusions
We have shown a number of actions of GH exerted on multiple tissues and organs far beyond the classic effects of the hormone on the intermediate metabolism and growth. The high diversity of actions of GH can be explained only by the fact that the hormone plays many different roles by activating a high number of proteins involved in cell signaling and displaying different mechanisms of action, as recently demonstrated by Ray et al346 and Carter-Su et al.347 The possibility exists that, rather than a hormone, GH is a prohormone that depending on the tissue may be proteolytically cleaved giving origin to different and shorter GH derivatives with tissue-specific properties. In addition, GH may activate the proliferation of tissue-specific stem cells that then would act in tissue repair after an injury. In this sense, GH is safe if administered in the appropriate doses and frequency. A classical GH-dependent adverse effect such as hyperglycemia is not important if the hormone is administered before physical exercise. Moreover, recently, it has been reported that GH is able to induce the rescue of pancreatic β-cell and function in streptozotocin-treated mice and, therefore, may be of interest in the treatment of type 1 diabetes.348
Footnotes
ACADEMIC EDITOR: Nigel Irwin, Editor in Chief
PEER REVIEW: Two peer reviewers contributed to the peer review report. Reviewers’ reports totaled 1150 words, excluding any confidential comments to the academic editor.
FUNDING: This review and some of the studies presented here have been supported by Foundation Foltra (Teo, Spain). The authors confirm that the funder had no influence over the study design, content of the article, or selection of this journal.
COMPETING INTERESTS: Authors disclose no potential conflicts of interest.
Provenance: the authors were invited to submit this paper.
Paper subject to independent expert single-blind peer review. All editorial decisions made by independent academic editor. Upon submission manuscript was subject to anti-plagiarism scanning. Prior to publication all authors have given signed confirmation of agreement to article publication and compliance with all applicable ethical and legal requirements, including the accuracy of author and contributor information, disclosure of competing interests and funding sources, compliance with ethical requirements relating to human and animal study participants, and compliance with any copyright requirements of third parties. This journal is a member of the Committee on Publication Ethics (COPE).
Author Contributions
Conceived and designed the experiments: JD. Analyzed the data: PD, CA and JD. Wrote the first draft: JD. Contributed to the writing: PD and CA. Agree with results and conclusions: JD, PD, CA. Jointly developed the structure and arguments: JD, PD, CA. Made critical revisions and approved final version: JD, CA and PD. All authors reviewed and approved of the final manuscript.
REFERENCES
- 1.Evans HM, Long JA. The effect of the anterior lobe administered intraperitoneally upon growth maturation, and oestrus cycles of the rat. Anatomical Record. 1921;21:62–63. [Google Scholar]
- 2.Raben MS. Treatment of a pituitary dwarf with human growth hormone. J Clin Endocrinol Metab. 1958;18:901–903. doi: 10.1210/jcem-18-8-901. [DOI] [PubMed] [Google Scholar]
- 3.Keohane C. Prion diseases in men. Arch Anat Cytol Pathol. 1994;42:69–75. [PubMed] [Google Scholar]
- 4.Stockdale T. Contaminated material caused Creutzfeld-Jakob disease (CJD) in some undersized children who were treated with growth hormone (GH) Nutr Health. 2000;14:141–142. doi: 10.1177/026010600001400207. [DOI] [PubMed] [Google Scholar]
- 5.Sairam MR, Chrétien M, Li CH. On the isolation of human pituitary hormones. J Clin Endocrinol Metab. 1978;47:1002–1008. doi: 10.1210/jcem-47-5-1002. [DOI] [PubMed] [Google Scholar]
- 6.Li CH, Dixon JS. Human pituitary growth hormone. 32. The primary structure of the hormone: revision. Arch Biochem Biophys. 1971;146:233–236. doi: 10.1016/s0003-9861(71)80060-7. [DOI] [PubMed] [Google Scholar]
- 7.Murray PG, Clayton PE. Endocrine control of growth. Am J Med Genet C Semin Med Genet. 2013;163C:76–85. doi: 10.1002/ajmg.c.31357. [DOI] [PubMed] [Google Scholar]
- 8.Karlberg J, Albertsson-Wikland K. Infancy growth pattern related to growth hormone deficiency. Acta Paediatr Scand. 1988;77:385–391. doi: 10.1111/j.1651-2227.1988.tb10665.x. [DOI] [PubMed] [Google Scholar]
- 9.Karlberg J. The infancy-childhood growth spurt. Acta Paediatr Scand Suppl. 1990;367:111–118. [PubMed] [Google Scholar]
- 10.Albertsson-Wikland K, Boguszewski M, Karlberg J. Children born small-for-gestational age: postnatal growth and hormonal status. Horm Res. 1998;49(Suppl 2):7–13. [PubMed] [Google Scholar]
- 11.Low LC, Tam SY, Kwan EY, Tsang AM, Karlberg J. Onset of significant GH dependence of serum IGF-I and IGF-binding protein 3 concentrations in early life. Pediatr Res. 2001;50:737–742. doi: 10.1203/00006450-200112000-00018. [DOI] [PubMed] [Google Scholar]
- 12.Schlessinger J, Shechter Y, Willingham MC, Pastan I. Direct visualization of binding, aggregation, and internalization of insulin and epidermal growth factor on living fibroblastic cells. Proc Natl Acad Sci U S A. 1978;75:2659–2663. doi: 10.1073/pnas.75.6.2659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Gorden P, Carpentier JL, Freychet PO, Orci L. Internalization of polypeptide hormones: mechanism, intracellular localization and significance. Diabetologia. 1980;18:263–274. doi: 10.1007/BF00251003. [DOI] [PubMed] [Google Scholar]
- 14.Waters MJ, Rowlinson SW, Clarkson RW, et al. Signal transduction by the growth hormone receptor. Proc Soc Exp Biol Med. 1994;206:216–220. doi: 10.3181/00379727-206-43745. [DOI] [PubMed] [Google Scholar]
- 15.Lobie PE, Mertani H, Morel G, Morales-Bustos O, Norstedt G, Waters MJ. Receptor-mediated nuclear translocation of growth hormone. J Biol Chem. 1994;269:21330–21339. [PubMed] [Google Scholar]
- 16.Lobie PE, Wood TJ, Chen CM, Waters MJ, Norstedt G. Nuclear translocation and anchorage of the growth hormone receptor. J Biol Chem. 1994;269:31735–31746. [PubMed] [Google Scholar]
- 17.García-Barros M, Devesa J, Arce VM. Proteolytic processing of human growth hormone (GH) by rat tissues in vitro: influence of sex and age. J Endocrinol Invest. 2000;23:748–754. doi: 10.1007/BF03345065. [DOI] [PubMed] [Google Scholar]
- 18.Devesa J, Díaz MJ, Odriozola A, Arce V, Lima L. Neurorregulación de la secreción de hormona de crecimiento (GH) y expresión del gen de esta hormona en pro- y eucariotas. Endocrinología. 1991;38:33–41. [Google Scholar]
- 19.Devesa J, Devesa P, Reimunde P. Growth hormone revisited. Med Clin (Barc) 2010;135:665–670. doi: 10.1016/j.medcli.2009.10.017. [DOI] [PubMed] [Google Scholar]
- 20.Arce VM, Devesa P, Devesa J. Role of growth hormone (GH) in the treatment on neural diseases: from neuroprotection to neural repair. Neurosci Res. 2013;76:179–186. doi: 10.1016/j.neures.2013.03.014. [DOI] [PubMed] [Google Scholar]
- 21.Hirt H, Kimelman J, Birnbaum MJ, et al. The human growth hormone gene locus: structure, evolution, and allelic variations. DNA. 1987;6:59–70. doi: 10.1089/dna.1987.6.59. [DOI] [PubMed] [Google Scholar]
- 22.Diaz MJ, Dominguez F, Haro LS, Ling N, Devesa J. A 12-kilodalton N-glycosylated growth hormone-related peptide is present in human pituitary extracts. J Clin Endocrinol Metab. 1993;77:134–138. doi: 10.1210/jcem.77.1.8325936. [DOI] [PubMed] [Google Scholar]
- 23.García-Barros M, Costoya JA, Ríos R, Arce V, Devesa J. N-glycosylated variants of growth hormone in human pituitary extracts. Horm Res. 2000;53:40–45. doi: 10.1159/000023512. [DOI] [PubMed] [Google Scholar]
- 24.Baumann G. Growth hormone heterogeneity: genes, isohormones, variants and binding proteins. Endocr Rev. 1991;121:424–449. doi: 10.1210/edrv-12-4-424. [DOI] [PubMed] [Google Scholar]
- 25.Baumann GP. Growth hormone isoforms. Growth Horm IGF Res. 2009;19:333–340. doi: 10.1016/j.ghir.2009.04.011. [DOI] [PubMed] [Google Scholar]
- 26.Spolaore B, Polverino de Laureto P, Zambonin M, Fontana A. Limited proteolysis of human growth hormone at low pH: isolation, characterization, and complementation of the two biologically relevant fragments 1–44 and 45–191. Biochemistry. 2004;43:6576–6586. doi: 10.1021/bi049491g. [DOI] [PubMed] [Google Scholar]
- 27.Such-Sanmartín G, Bosch J, Segura J, Gutiérrez-Gallego R. Generation of 5 and 17 kDa human growth hormone fragments through limited proteolysis. Growth Factors. 2009;27:255–264. doi: 10.1080/08977190903110121. [DOI] [PubMed] [Google Scholar]
- 28.Luna M, Martínez-Moreno CG, Ahumada-Solórzano MS, Harvey S, Carranza M, Arámburo C. Extrapituitary growth hormone in the chicken reproductive system. Gen Comp Endocrinol. 2014;203:60–68. doi: 10.1016/j.ygcen.2014.02.021. [DOI] [PubMed] [Google Scholar]
- 29.Lewis UJ, Ling N, Haro LS. Variants forms of human growth hormone in serum. Acta Paediatr. 1994;399(Suppl):29–31. doi: 10.1111/j.1651-2227.1994.tb13282.x. [DOI] [PubMed] [Google Scholar]
- 30.Sinha YN, Jacobsen BP. Human growth hormone (hGH)-(44–191) a reportedly diabetogenic fragment of hGH, circulates in human blood: measurement by radioimmunoassay. J Clin Endocrinol Metab. 1994;78:1411–1418. doi: 10.1210/jcem.78.6.8200944. [DOI] [PubMed] [Google Scholar]
- 31.Lewis UJ, Lewis MJ, Salem MAM, Staten NR, Galosy SS, Krivi GG. A recombinant-DNA-derived modification of human growth hormone (hGH 44–191) with enhanced diabetogenic activity. Mol Cell Endocrinol. 1991;78:45–54. doi: 10.1016/0303-7207(91)90184-t. [DOI] [PubMed] [Google Scholar]
- 32.Rowlinson SW, Waters MJ, Lewis UJ, Barnard R. Human growth hormone fragments 1–43 and 44–191: in vitro somatogenic activity and receptor binding characteristics in human and nonprimate systems. Endocrinology. 1996;137:90–95. doi: 10.1210/endo.137.1.8536647. [DOI] [PubMed] [Google Scholar]
- 33.Salmon W, Daughaday W. A hormonally controlled serum factor which stimulates sulfate incorporation by cartilage in vitro. J Lab Clin Med. 1957;49:825–836. [PubMed] [Google Scholar]
- 34.Fagin JA, Fernandez-Mejia C, Melmed S. Pituitary insulin-like growth factor-I gene expression: Regulation by triiodothyronine and growth hormone. Endocrinology. 1989;125:2385–2391. doi: 10.1210/endo-125-5-2385. [DOI] [PubMed] [Google Scholar]
- 35.Daughaday WH, Hall K, Raben MS, Salmon WD, Jr, van den Brande JL, van Wyk JJ. Somatomedin: proposed designation for sulphation factor. Nature. 1972;235:107. doi: 10.1038/235107a0. [DOI] [PubMed] [Google Scholar]
- 36.Green H, Morikawa M, Nixon T. A dual effector theory of growth-hormone action. Differentiation. 1985;29:195–198. doi: 10.1111/j.1432-0436.1985.tb00316.x. [DOI] [PubMed] [Google Scholar]
- 37.Reinecke M, Schmid AC, Heyberger-Meyer B, Hunziker EB, Zapf J. Effect of growth hormone and insulin-like growth factor I (IGF-I) on the expression of IGF-I messenger ribonucleic acid and peptide in rat tibial growth plate and articular chondrocytes in vivo. Endocrinology. 2000;141:2847–2853. doi: 10.1210/endo.141.8.7624. [DOI] [PubMed] [Google Scholar]
- 38.Ohlsson C, Sjögren K, Jansson JO, Isaksson OG. The relative importance of endocrine versus autocrine/paracrine insulin-like growth factor-I in the regulation of body growth. Pediatr Nephrol. 2000;14:541–543. doi: 10.1007/s004670000348. [DOI] [PubMed] [Google Scholar]
- 39.Devesa J, Lima L, Tresguerres JA. Neuroendocrine control of growth hormone secretion in humans. Trends Endocrinol Metab. 1992;3:175–183. doi: 10.1016/1043-2760(92)90168-z. [DOI] [PubMed] [Google Scholar]
- 40.Steyn FJ, Tolle V, Chen C, Epelbaum J. Neuroendocrine Regulation of Growth Hormone Secretion. Compr Physiol. 2016;6:687–735. doi: 10.1002/cphy.c150002. [DOI] [PubMed] [Google Scholar]
- 41.Isaksson OG, Jansson JO, Gause IA. Growth hormone stimulates longitudinal bone growth directly. Science. 1982;216:1237–1239. doi: 10.1126/science.7079756. [DOI] [PubMed] [Google Scholar]
- 42.Schlechter NL, Russell SM, Spencer EM, Nicoll CS. Evidence suggesting that the direct growth-promoting effect of growth hormone on cartilage in vivo is mediated by local production of somatomedin. Proc Natl Acad Sci U S A. 1986;83:7932–7934. doi: 10.1073/pnas.83.20.7932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Lindahl A, Isgaard J, Nilsson A, Isaksson OG. Growth hormone potentiates colony formation of epiphyseal chondrocytes in suspension culture. Endocrinology. 1986;118:1843–1848. doi: 10.1210/endo-118-5-1843. [DOI] [PubMed] [Google Scholar]
- 44.Jux C, Leiber K, Hügel U. Dexamethasone impairs growth hormone (GH)-stimulated growth by suppression of local insulin-like growth factor (IGF)-I production and expression of GH- and IGF-I-receptor in cultured rat chondrocytes. Endocrinology. 1998;139:3296–3305. doi: 10.1210/endo.139.7.6099. [DOI] [PubMed] [Google Scholar]
- 45.Ohlsson C, Bengtsson BA, Isaksson OG, Andreassen TT, Slootweg MC. Growth hormone and bone. Endocr Rev. 1998;19:55–79. doi: 10.1210/edrv.19.1.0324. [DOI] [PubMed] [Google Scholar]
- 46.Laron Z, Pertzelan A, Mannheimer S. Genetic pituitary dwarfism with high serum concentation of growth hormone—a new inborn error of metabolism? Isr J Med Sci. 1966;2:152–155. [PubMed] [Google Scholar]
- 47.Laron Z, Kauli R2. Fifty seven years of follow-up of the Israeli cohort of Laron Syndrome patients-From discovery to treatment. Growth Horm IGF Res. 2016;28:53–56. doi: 10.1016/j.ghir.2015.08.004. [DOI] [PubMed] [Google Scholar]
- 48.Zhou Y, Xu BC, Maheshwari HG, et al. A mammalian model for Laron syndrome produced by targeted disruption of the mouse growth hormone receptor/binding protein gene (the Laron mouse) Proc Natl Acad Sci U S A. 1997;94:13215–13220. doi: 10.1073/pnas.94.24.13215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Sjögren K, Bohlooly-Y M, Olsson B, et al. Disproportional skeletal growth and markedly decreased bone mineral content in growth hormone receptor –/– mice. Biochem Biophys Res Commun. 2000;267:603–608. doi: 10.1006/bbrc.1999.1986. [DOI] [PubMed] [Google Scholar]
- 50.Marcovecchio ML, Chiarelli F. Obesity and growth during childhood and puberty. World Rev Nutr Diet. 2013;106:135–141. doi: 10.1159/000342545. [DOI] [PubMed] [Google Scholar]
- 51.De Leonibus C, Marcovecchio ML, Chiarelli F. Update on statural growth and pubertal development in obese children. Pediatr Rep. 2012;4:e35. doi: 10.4081/pr.2012.e35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Pijl H, Langendonk JG, Burggraaf J, et al. Altered neuroregulation of GH secretion in viscerally obese premenopausal women. J Clin Endocrinol Metab. 2001;86:5509–5515. doi: 10.1210/jcem.86.11.8061. [DOI] [PubMed] [Google Scholar]
- 53.Ghione S, Maestro S, Dati E, Silvestri V, Baroncelli GI, Bertelloni S. Growth impairment and growth hormone-IGF-1 axis in infantile anorexia nervosa. J Pediatr Endocrinol Metab. 2015;28:999–1001. doi: 10.1515/jpem-2013-0400. [DOI] [PubMed] [Google Scholar]
- 54.Golden NH, Kreitzer P, Jacobson MS, et al. Disturbances in growth hormone secretion and action in adolescents with anorexia nervosa. J Pediatr. 1994;125:655–660. doi: 10.1016/s0022-3476(94)70030-3. [DOI] [PubMed] [Google Scholar]
- 55.Goya L, de la Puente A, Ramos S, Martín MA, Escrivá F, Pascual-Leone AM. Regulation of insulin-like growth factor-I and -II by glucose in primary cultures of fetal rat hepatocytes. J Biol Chem. 1999;274:24633–24640. doi: 10.1074/jbc.274.35.24633. [DOI] [PubMed] [Google Scholar]
- 56.Devesa J, Casteleiro N, Rodicio C, López N, Reimunde P. Growth hormone deficiency and cerebral palsy. Ther Clin Risk Manag. 2010;6:413–418. doi: 10.2147/tcrm.s12312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Parker EA, Hegde A, Buckley M, Barnes KM, Baron J, O Nilsson O. Spatial and temporal regulation of GH–IGF-related gene expression in growth plate cartilage. J Endocrinology. 2007;194:31–40. doi: 10.1677/JOE-07-0012. [DOI] [PubMed] [Google Scholar]
- 58.Abad V, Meyers JL, Weise M, et al. The role of resting zone in growth plate chondrogenesis. Endocrinology. 2002;143:1851–1857. doi: 10.1210/endo.143.5.8776. [DOI] [PubMed] [Google Scholar]
- 59.Aszodi A, Hunziker EB, Brakebusch C, Fässler R. β1 Integrins regulate chondrocyte rotation, G1 progression, and cytokinesis. Genes and Development. 2003;17:2465–2479. doi: 10.1101/gad.277003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Gerber H, Vu TH, Ryan AM, Kowalski J, Werb Z, Ferrara N. VEGF couples hypertrophic cartilage remodeling, ossification and angiogenesis during endochondral bone formation. Nature Medicine. 1999;5:623–628. doi: 10.1038/9467. [DOI] [PubMed] [Google Scholar]
- 61.Nilsson O, Baron J. Fundamental limits on longitudinal bone growth: growth plate senescence and epiphyseal fusion. Trends Endocrinol Metab. 2004;15:370–374. doi: 10.1016/j.tem.2004.08.004. [DOI] [PubMed] [Google Scholar]
- 62.Gevers EF, Milne J, Robinson IC, Loveridge N. Single cell enzyme activity and proliferation in the growth plate: effects of growth hormone. J Bone Miner Res. 1996;11:1103–1111. doi: 10.1002/jbmr.5650110809. [DOI] [PubMed] [Google Scholar]
- 63.Heinrichs C, Yanovski JA, Roth AH, et al. Dexamethasone increases growth hormone receptor messenger ribonucleic acid levels in liver and growth plate. Endocrinology. 1994;135:1113–1118. doi: 10.1210/endo.135.3.8070354. [DOI] [PubMed] [Google Scholar]
- 64.Gevers EF, van der Eerden BC, Karperien M, Raap AK, Robinson IC, Wit JM. Localization and regulation of the growth hormone receptor and growth hormone-binding protein in the rat growth plate. J Bone Miner Res. 2002;17:1408–1419. doi: 10.1359/jbmr.2002.17.8.1408. [DOI] [PubMed] [Google Scholar]
- 65.Shinar DM, Endo N, Halperin D, Rodan GA, Weinreb M. Differential expression of insulin-like growth factor-I (IGF-I) and IGF-II messenger ribonucleic acid in growing rat bone. Endocrinology. 1993;132:1158–1167. doi: 10.1210/endo.132.3.8440176. [DOI] [PubMed] [Google Scholar]
- 66.Wang E, Wang J, Chin E, Zhou J, Bondy CA. Cellular patterns of insulin-like growth factor system gene expression in murine chondrogenesis and osteogenesis. Endocrinology. 1995;136:2741–2751. doi: 10.1210/endo.136.6.7750499. [DOI] [PubMed] [Google Scholar]
- 67.Wu S, Yang W, De Luca F. Insulin-Like Growth Factor-Independent Effects of Growth Hormone on Growth Plate Chondrogenesis and Longitudinal Bone Growth. Endocrinology. 2015;156:2541–2551. doi: 10.1210/en.2014-1983. [DOI] [PubMed] [Google Scholar]
- 68.Dobie R, Ahmed SF, Staines KA, et al. Increased linear bone growth by GH in the absence of SOCS2 is independent of IGF-1. J Cell Physiol. 2015;230:2796–2806. doi: 10.1002/jcp.25006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Chia DJ. Minireview: Mechanisms of Growth Hormone-Mediated Gene Regulation. Mol Endocrinol. 2014;28:1012–1025. doi: 10.1210/me.2014-1099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Waters MJ, Brooks AJ. Growth hormone and cell growth. Endocr Dev. 2012;23:86–95. doi: 10.1159/000341761. [DOI] [PubMed] [Google Scholar]
- 71.Hull KL, Harvey S. Growth Hormone and Reproduction: A Review of Endocrine and Autocrine/Paracrine Interactions. Int J Endocrinol. 2014;2014:234014. doi: 10.1155/2014/234014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Hull KL, Harvey S. Growth hormone: roles in male reproduction. Endocrine. 2000;13:243–250. doi: 10.1385/ENDO:13:3:243. [DOI] [PubMed] [Google Scholar]
- 73.Untergasser G, Hermann M, Rumpold H, Berger P. Complex alternative splicing of the GH-V gene in the human testis. European J Endocrinology. 1998;139(4):424–427. doi: 10.1530/eje.0.1390424. [DOI] [PubMed] [Google Scholar]
- 74.Berensztein EB, Baquedano MS, Pepe CM, et al. Role of IGFs and insulin in the human testis during postnatal activation: differentiation of steroidogenic cells. Pediatr Res. 2008;63:662–666. doi: 10.1203/PDR.0b013e31816c8ffc. [DOI] [PubMed] [Google Scholar]
- 75.Gallego Gómez R, Pintos E, et al. Cellular distribution of growth hormone-releasing hormone receptor in human reproductive system and breast and prostate cancers. Histol Histopathol. 2005;20:697–706. doi: 10.14670/HH-20.697. [DOI] [PubMed] [Google Scholar]
- 76.Tena-Sempere M. Exploring the role of ghrelin as novel regulator of gonadal function. Growth Horm IGF Res. 2005;15:83–88. doi: 10.1016/j.ghir.2005.02.001. [DOI] [PubMed] [Google Scholar]
- 77.Barreiro ML, Tena-Sempere M. Ghrelin and reproduction: a novel signal linking energy status and fertility? Mol Cell Endocrinol. 2004;226:1–9. doi: 10.1016/j.mce.2004.07.015. [DOI] [PubMed] [Google Scholar]
- 78.Riaz H, Liang A, Khan MK, et al. Somatostatin and its receptors: functional regulation in the development of mice Sertoli cells. J Steroid Biochem Mol Biol. 2013;138:257–266. doi: 10.1016/j.jsbmb.2013.06.007. [DOI] [PubMed] [Google Scholar]
- 79.Nguyen AP, Chandorkar A, Gupta C. The role of growth hormone in fetal mouse reproductive tract differentiation. Endocrinology. 1996;137:3659–3666. doi: 10.1210/endo.137.9.8756530. [DOI] [PubMed] [Google Scholar]
- 80.Laron Z, Klinger B. Effect of insulin-like growth factor-I treatment on serum androgens and testicular and penile size in males with Laron syndrome (primary growth hormone resistance) European Journal of Endocrinology. 1998;138:176–180. doi: 10.1530/eje.0.1380176. [DOI] [PubMed] [Google Scholar]
- 81.Carani C, Granata ARM, De Rosa M, et al. The effect of chronic treatment with GH on gonadal function in men with isolated GH deficiency. European Journal of Endocrinology. 1999;140:224–230. doi: 10.1530/eje.0.1400224. [DOI] [PubMed] [Google Scholar]
- 82.Balducci R, Toscano V, Mangiantini A, et al. The effect of growth hormone administration on testicular response during gonadotropin therapy in subjects with combined gonadotropin and growth hormone deficiencies. Acta Endocrinol. 1993;128:19–23. doi: 10.1530/acta.0.1280019. [DOI] [PubMed] [Google Scholar]
- 83.Oscarsson J, Lindstedt G, Lundberg PA, Edén S. Continuous subcutaneous infusion of low dose growth hormone decreases serum sex-hormone binding globulin and testosterone concentrations in moderately obese middle-aged men. Clin Endocrinol (Oxf) 1996;44:23–29. doi: 10.1046/j.1365-2265.1996.635457.x. [DOI] [PubMed] [Google Scholar]
- 84.Giavoli C, Ferrante E, Ermetici F, et al. Effect of recombinant hGH (rhGH) replacement on gonadal function in male patients with organic adult-onset GH deficiency. Clin Endocrinology. 2006;65:717–721. doi: 10.1111/j.1365-2265.2006.02655.x. [DOI] [PubMed] [Google Scholar]
- 85.Andreassen M, Frystyk J, Faber J, Kristensen LØ, Juul A. Growth hormone (GH) activity is associated with increased serum oestradiol and reduced Anti-Müllerian Hormone in healthy male volunteers treated with GH and a GH antagonist. Andrology. 2013;1:595–601. doi: 10.1111/j.2047-2927.2013.00096.x. [DOI] [PubMed] [Google Scholar]
- 86.Jones JI, Clemmons DR. Insulin-like growth factors and their binding proteins: biological actions. Endocr Rev. 1995;16:3–34. doi: 10.1210/edrv-16-1-3. [DOI] [PubMed] [Google Scholar]
- 87.Cailleau J, Vermeire S, Verhoeven G. Independent control of the production of insulin-like growth factor I and its binding protein by cultured testicular cells. Mol Cell Endocrinol. 1990;69:79–89. doi: 10.1016/0303-7207(90)90091-l. [DOI] [PubMed] [Google Scholar]
- 88.Bartke A, Sun LY, Longo V. Somatotropic signaling: trade-offs between growth, reproductive development, and longevity. Physiol Rev. 2013;93:571–598. doi: 10.1152/physrev.00006.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Dombrowicz D, Hooghe-Peters EL, Gothot A, et al. Cellular localization of IGF-I and IGF-II mRNAs in immature hypophysectomized rat testis and epididymis after in vivo hormonal treatment. Arch Int Physiol Biochim Biophys. 1992;100:303–308. doi: 10.3109/13813459209000717. [DOI] [PubMed] [Google Scholar]
- 90.Roser JF. Regulation of testicular function in the stallion: an intricate network of endocrine, paracrine and autocrine systems. Anim Reprod Sci. 2008;107:179–196. doi: 10.1016/j.anireprosci.2008.05.004. [DOI] [PubMed] [Google Scholar]
- 91.Tanaka M, Hayashida Y, Sakaguchi K, et al. Growth hormone-independent expression of insulin-like growth factor I messenger ribonucleic acid in extrahepatic tissues of the chicken. Endocrinology. 1996;137:30–34. doi: 10.1210/endo.137.1.8536628. [DOI] [PubMed] [Google Scholar]
- 92.De Boer JAM, Schoemaker J, Van Der Veen EA. Impaired reproductive function in women treated for growth hormone deficiency during childhood. Clin Endocrinol (Oxf) 1997;46:681–689. doi: 10.1046/j.1365-2265.1997.1800999.x. [DOI] [PubMed] [Google Scholar]
- 93.Giampietro A, Milardi D, Bianchi A, et al. The effect of treatment with growth hormone on fertility outcome in eugonadal women with growth hormone deficiency: report of four cases and review of the literature. Fertil Steril. 2009;91:930.e7–930.11. doi: 10.1016/j.fertnstert.2008.09.065. [DOI] [PubMed] [Google Scholar]
- 94.Sirotkin AV, Makarevich AV, Hetényi L. Effect of growth hormone and inhibitors of protein kinase a on IGF-I, oxytocin and progesterone release by cultured bovine granulosa cells. Ann Endocrinol (Paris) 2000;61:154–158. [PubMed] [Google Scholar]
- 95.Sirotkin AV. Direct action of growth hormone on bovine ovarian cells: effects on estradiol, oxytocin, vasopressin release by granulosa cells and on oocyte maturation and cleavage in vitro. Ann Endocrinol (Paris) 1996;57:219–224. [PubMed] [Google Scholar]
- 96.Karamouti M, Kollia P, Kallitsaris A, Vamvakopoulos N, Kollios G, Messinis IE. Growth hormone, insulin-like growth factor I, and leptin interaction in human cultured lutein granulosa cells steroidogenesis. Fertil Steril. 2008;90:1444–14450. doi: 10.1016/j.fertnstert.2007.08.076. [DOI] [PubMed] [Google Scholar]
- 97.Doldi N, Bassan M, Bonzi V, Ferrari A. Effects of growth hormone and growth hormone-releasing hormone on steroid synthesis in cultured human luteinizing granulosa cells. Gynecol Endocrinol. 1996;10:101–108. doi: 10.3109/09513599609097899. [DOI] [PubMed] [Google Scholar]
- 98.Gregoraszczuk EL, Ptak A. In vitro effect of leptin on growth hormone (GH)-and insulin-like growth factor-I (IGF-I)-stimulated progesterone secretion and apoptosis in developing and mature corpora lutea of pig ovaries. J Reprod Dev. 2005;51:727–733. doi: 10.1262/jrd.17038. [DOI] [PubMed] [Google Scholar]
- 99.Gregoraszczuk EL, Ptak A, Wojtowicz AK, Gorska T, Nowak KW. Estrus cycle-dependent action of leptin on basal and GH or IGF-I stimulated steroid secretion by whole porcine follicles. Endocr Regul. 2004;38:15–21. [PubMed] [Google Scholar]
- 100.Chase CC, Jr, Kirby CJ, Hammond AC, Olson TA, Lucy MC. Patterns of ovarian growth and development in cattle with a growth hormone receptor deficiency. J Anim Sci. 1998;76:212–219. doi: 10.2527/1998.761212x. [DOI] [PubMed] [Google Scholar]
- 101.Hull KL, Harvey S. Growth hormone: roles in female reproduction. J Endocrinol. 2001;168:1–23. doi: 10.1677/joe.0.1680001. [DOI] [PubMed] [Google Scholar]
- 102.Bachelot A, Monget P, Imbert-Bolloré P, et al. Growth hormone is required for ovarian follicular growth. Endocrinology. 2002;143:4104–4112. doi: 10.1210/en.2002-220087. [DOI] [PubMed] [Google Scholar]
- 103.Slot KA, Kastelijn J, Bachelot A, Kelly PA, Binart N, Teerds KJ. Reduced recruitment and survival of primordial and growing follicles in GH receptor-deficient mice. Reproduction. 2006;131:525–532. doi: 10.1530/rep.1.00946. [DOI] [PubMed] [Google Scholar]
- 104.Nakamura E, Otsuka F, Inagaki K, et al. Mutual regulation of growth hormone and bone morphogenetic protein system in steroidogenesis by rat granulosa cells. Endocrinology. 2012;153:469–480. doi: 10.1210/en.2011-1646. [DOI] [PubMed] [Google Scholar]
- 105.Kiapekou E, Loutradis D, Drakakis P, Zapanti E, Mastorakos G, Antsaklis A. Effects of GH and IGF-I on the in vitro maturation of mouse oocytes. Hormones (Athens) 2005;4:155–160. doi: 10.14310/horm.2002.11153. [DOI] [PubMed] [Google Scholar]
- 106.Menezo YJR, Nicollet B, Rollet J, Hazout A. Pregnancy and delivery after in vitro maturation of naked ICSI-GV oocytes with GH and transfer of a frozen thawed blastocyst: case report. J Assist Reprod Genet. 2006;23:47–49. doi: 10.1007/s10815-005-9014-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Ménézo YJR, El Mouatassim S, Chavrier M, Servy EJ, Nicolet B. Human oocytes and preimplantation embryos express mRNA for growth hormone receptor. Zygote. 2003;11:293–297. doi: 10.1017/s096719940300234x. [DOI] [PubMed] [Google Scholar]
- 108.Abir R, Garor R, Felz C, Nitke S, Krissi H, Fisch B. Growth hormone and its receptor in human ovaries from fetuses and adults. Fertil Steril. 2008;90:1333–1339. doi: 10.1016/j.fertnstert.2007.08.011. [DOI] [PubMed] [Google Scholar]
- 109.Lobie PE, Breipohl W, Aragón JG, Waters MJ. Cellular localization of the growth hormone receptor/binding protein in the male and female reproductive systems. Endocrinology. 1990;126:2214–2221. doi: 10.1210/endo-126-4-2214. [DOI] [PubMed] [Google Scholar]
- 110.Silva JRV, Figueiredo JR, van den Hurk R. Involvement of growth hormone (GH) and insulin-like growth factor (IGF) system in ovarian folliculogenesis. Theriogenology. 2009;71:1193–1208. doi: 10.1016/j.theriogenology.2008.12.015. [DOI] [PubMed] [Google Scholar]
- 111.Lobie PE, García-Aragón J, Wang BS, Baumbach WR, Waters MJ. Cellular localization of the growth hormone binding protein in the rat. Endocrinology. 1992;130:3057–3065. doi: 10.1210/endo.130.5.1374020. [DOI] [PubMed] [Google Scholar]
- 112.Schwärzler P, Untergasser G, Hermann M, et al. Selective growth hormone/placental lactogen gene transcription and hormone production in pre- and postmenopausal human ovaries. J Clin Endocrinol Metab. 1997;82:3337–3341. doi: 10.1210/jcem.82.10.4316. [DOI] [PubMed] [Google Scholar]
- 113.Izadyar F, Zhao J, van Tol HT, et al. Messenger RNA expression and protein localization of growth hormone in bovine ovarian tissue and in cumulus oocyte complexes (COCs) during in vitro maturation. Mol Reprod Dev. 1999;53:398–406. doi: 10.1002/(SICI)1098-2795(199908)53:4<398::AID-MRD5>3.0.CO;2-I. [DOI] [PubMed] [Google Scholar]
- 114.Hrabia A, Paczoska-Eliasiewicz HE, Berghman LR, Harvey S, Rzaąsa J. Expression and localization of growth hormone and its receptors in the chicken ovary during sexual maturation. Cell Tissue Res. 2008;332:317–328. doi: 10.1007/s00441-008-0595-7. [DOI] [PubMed] [Google Scholar]
- 115.Moretti C, Bagnato A, Solan N, Frajese G, Catt KJ. Receptor-mediated actions of growth hormone releasing factor on granulosa cell differentiation. Endocrinology. 1990;127:2117–2126. doi: 10.1210/endo-127-5-2117. [DOI] [PubMed] [Google Scholar]
- 116.Bagnato A, Moretti C, Ohnishi J, Frajese G, Catt KJ. Expression of the growth hormone-releasing hormone gene and its peptide product in the rat ovary. Endocrinology. 1992;130:1097–1102. doi: 10.1210/endo.130.3.1537276. [DOI] [PubMed] [Google Scholar]
- 117.Rak A, Gregoraszczuk EŁ. Local feedback loop of ghrelin-GH in the pig ovary: action on estradiol secretion, aromatase activity and cell apoptosis. Growth Horm IGF Res. 2008;18:221–227. doi: 10.1016/j.ghir.2007.09.004. [DOI] [PubMed] [Google Scholar]
- 118.Rak A, Szczepankiewicz D, Gregoraszczuk EŁ. Expression of ghrelin receptor, GHSR-1a, and its functional role in the porcine ovarian follicles. Growth Horm IGF Res. 2009;19:68–76. doi: 10.1016/j.ghir.2008.08.006. [DOI] [PubMed] [Google Scholar]
- 119.Sirotkin AV, Grossmann R, María-Peon MT, Roa J, Tena-Sempere M, Klein S. Novel expression and functional role of ghrelin in chicken ovary. Mol Cell Endocrinol. 2006;257–258:15–25. doi: 10.1016/j.mce.2006.06.004. [DOI] [PubMed] [Google Scholar]
- 120.Ptak A, Kajta M, Gregoraszczuk EL. Effect of growth hormone and insulin-like growth factor-I on spontaneous apoptosis in cultured luteal cells collected from early, mature, and regressing porcine corpora lutea. Anim Reprod Sci. 2004;80:267–279. doi: 10.1016/j.anireprosci.2003.07.006. [DOI] [PubMed] [Google Scholar]
- 121.Apa R, Lanzone A, Miceli F, Caruso A, Mancuso S, Canipari R. Growth hormone induction of rat granulosa cell tissue-plasminogen activator expression and progesterone synthesis. Mol Cell Endocrinol. 1994;99:153–159. doi: 10.1016/0303-7207(94)90003-5. [DOI] [PubMed] [Google Scholar]
- 122.Hsu C-J, Hammond JM. Gonadotropins and estradiol stimulate immunoreactive insulin-like growth factor-I production by porcine granulosa cells in vitro. Endocrinology. 1987;120:198–207. doi: 10.1210/endo-120-1-198. [DOI] [PubMed] [Google Scholar]
- 123.Arunakumari G, Shanmugasundaram N, Rao VH. Development of morulae from the oocytes of cultured sheep preantral follicles. Theriogenology. 2010;74:884–894. doi: 10.1016/j.theriogenology.2010.04.013. [DOI] [PubMed] [Google Scholar]
- 124.Barreca A, Artini PG, Del Monte P, et al. In vivo and in vitro effect of growth hormone on estradiol secretion by human granulosa cells. J Clin Endocrinol Metab. 1993;77:61–67. doi: 10.1210/jcem.77.1.8325961. [DOI] [PubMed] [Google Scholar]
- 125.De Boer JA, van der Meer M, van der Veen EA, Schoemaker J. Growth hormone (GH) substitution in hypogonadotropic GH-deficient women decreases the follicle-stimulating hormone threshold for monofollicular growth. J Clin Endocrinol Metab. 1999;84:590–595. doi: 10.1210/jcem.84.2.5452. [DOI] [PubMed] [Google Scholar]
- 126.Homburg R, Ostergaard H. Clinical applications of growth hormone for ovarian stimulation. Hum Reprod Update. 1995;1:264–275. doi: 10.1093/humupd/1.3.264. [DOI] [PubMed] [Google Scholar]
- 127.Harper K, Proctor M, Huges E. Growth hormone for in vitro fertilization. Cochrane Database Syst Rev. 2003;(3):CD000099. doi: 10.1002/14651858.CD000099. [DOI] [PubMed] [Google Scholar]
- 128.Kucuk K, Kozinoglu H, Kaba A. Growth hormone co-treatment within a GnRH agonist long protocol in patients with poor ovarian response: a prospective, randomized, clinical trial. J Assist Reprod Genet. 2008;25:123–127. doi: 10.1007/s10815-008-9212-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Pantaleon M, Whiteside EJ, Harvey MB, et al. Functional growth hormone (GH) receptors and GH are expressed by preimplantation mouse embryos: a role for GH in early embryogenesis? Proc Nat Acad Sci U S A. 1997;94:5125–130. doi: 10.1073/pnas.94.10.5125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Fukaya T, Yamanaka T, Terada Y, Murakami T, Yajima A. Growth hormone improves mouse embryo development in vitro, and the effect is neutralized by growth hormone receptor antibody. Tohoku J Exp Med. 1998;184:113–122. doi: 10.1620/tjem.184.113. [DOI] [PubMed] [Google Scholar]
- 131.Joudrey EM, Lechniak D, Petrik J, King WA. Expression of growth hormone and its transcription factor, Pit-1, in early bovine development. Mol Reprod Dev. 2003;64:275–283. doi: 10.1002/mrd.10237. [DOI] [PubMed] [Google Scholar]
- 132.Markham KE, Kaye PL. Growth hormone, insulin-like growth factor I and cell proliferation in the mouse blastocyst. Reproduction. 2003;125:327–336. doi: 10.1530/rep.0.1250327. [DOI] [PubMed] [Google Scholar]
- 133.Izadyar F, van Tol HT, Hage WG, Bevers MM. Preimplantation bovine embryos express mRNA of growth hormone receptor and respond to growth hormone addition during in vitro development. Mol Reprod Dev. 2000;57:247–255. doi: 10.1002/1098-2795(200011)57:3<247::AID-MRD6>3.0.CO;2-Q. [DOI] [PubMed] [Google Scholar]
- 134.Pozzobon SE, Lagares MA, Brum DS, Leivas FG, Rubin MIB. Addition of recombinant human growth hormone to in vitro maturation medium of bovine oocytes. Reprod Domest Anim. 2005;40:19–22. doi: 10.1111/j.1439-0531.2004.00547.x. [DOI] [PubMed] [Google Scholar]
- 135.Drakakis P, Loutradis D, Milingos S, et al. A preliminary study of the effect of growth hormone on mouse preimplantation embryo development in vitro. Gynecol Obst Invest. 1995;40:222–226. doi: 10.1159/000292341. [DOI] [PubMed] [Google Scholar]
- 136.Lacroix M-C, Guibourdenche J, Fournier T, et al. Stimulation of human trophoblast invasion by placental growth hormone. Endocrinology. 2005;146:2434–2444. doi: 10.1210/en.2004-1550. [DOI] [PubMed] [Google Scholar]
- 137.Sbracia M, Scarpellini F, Poverini R, Alò PL, Rossi G, Di Tondo U. Immunohistochemical localization of the growth hormone in human endometrium and decidua. Am J Reprod Immunol. 2004;51:112–116. doi: 10.1046/j.8755-8920.2003.00127.x. [DOI] [PubMed] [Google Scholar]
- 138.Stygar D, Muravitskaya N, Eriksson B, Eriksson H, Sahlin L. Effects of SERM (selective estrogen receptor modulator) treatment on growth and proliferation in the rat uterus. Reprod Biol Endocrinol. 2003;1:40. doi: 10.1186/1477-7827-1-40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Kölle S, Stojkovic M, Reese S, Reichenbach H-D, Wolf E, Sinowatz F. Effects of growth hormone on the ultrastructure of bovine preimplantation embryos. Cell Tissue Res. 2004;317:101–108. doi: 10.1007/s00441-004-0898-2. [DOI] [PubMed] [Google Scholar]
- 140.Frankenne F, Closset J, Gomez F, Scippo ML, Smal J, Hennen G. The physiology of growth hormones (GHs) in pregnant women and partial characterization of the placental GH variant. J Clin Endocrinol Metab. 1988;66:1171–1180. doi: 10.1210/jcem-66-6-1171. [DOI] [PubMed] [Google Scholar]
- 141.Lonberg U, Damm P, Andersson AM, et al. Increase in maternal placental growth hormone during pregnancy and disappearance during parturition in normal and growth hormone-deficient pregnancies. Am J Obstet Gynecol. 2003;188:247–251. doi: 10.1067/mob.2003.82. [DOI] [PubMed] [Google Scholar]
- 142.Eriksson L, Frankenne F, Eden S, Hennen G, Von Schoultz B. Growth hormone 24-h serum profiles during pregnancy–lack of pulsatility for the secretion of the placental variant. Br J Obstet Gynaecol. 1989;96:949–953. doi: 10.1111/j.1471-0528.1989.tb03352.x. [DOI] [PubMed] [Google Scholar]
- 143.Curran AJ, Peacey SR, Shalet SM. Is maternal growth hormone essential for a normal pregnancy? Eur J Endocrinol. 1998;139:54–58. doi: 10.1530/eje.0.1390054. [DOI] [PubMed] [Google Scholar]
- 144.Mirlesse V, Frankenne F, Alsat E, Poncelet M, Hennen G, Evain-Brion D. Placental growth hormone levels in normal pregnancy and in pregnancies with intrauterine growth retardation. Pediatr Res. 1993;34:439–442. doi: 10.1203/00006450-199310000-00011. [DOI] [PubMed] [Google Scholar]
- 145.Belfort MB, Brown RS. Fetal and postnatal growth: Mechanisms, consequences and controversies. Curr Opin Endocrinol Diabetes Obes. 2007;14:1–2. doi: 10.1097/MED.0b013e328014506e. [DOI] [PubMed] [Google Scholar]
- 146.Hubinont C, Fisk NM, Nicolini U, Rodeck CH. Fetal hyperinsulinism in anencephaly. Am J Obstet Gynecol. 1989;161:306–307. doi: 10.1016/0002-9378(89)90505-x. [DOI] [PubMed] [Google Scholar]
- 147.Oberbauer AM. Developmental programming: the role of growth hormone. J Anim Sci Biotechnol. 2015;6:8. doi: 10.1186/s40104-015-0001-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Murray PG, Higham CE, Clayton PE. The hypothalamo-GH axis: the past 60 years. J Endocrinol. 2015;226:T123–T140. doi: 10.1530/JOE-15-0120. [DOI] [PubMed] [Google Scholar]
- 149.Sfezurri-Perri AN, Owens JA, Pringle KG, Roberts CT. The neglected role of insulin-like growth factors in the maternal circulation regulating fetal growth. J Physiol. 2011;589:7–20. doi: 10.1113/jphysiol.2010.198622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Louvi A, Accili D, Efstratiadis A. Growth-promoting interaction of IGF-II with the insulin receptor during mouse embryonic development. Dev Biol. 1997;189:33–48. doi: 10.1006/dbio.1997.8666. [DOI] [PubMed] [Google Scholar]
- 151.Han VK, Carter AM. Spatial and temporal patterns of expression of messenger RNA for insulin-like growth factors and their binding proteins in the placenta of man and laboratory animals. Placenta. 2000;21:289–305. doi: 10.1053/plac.1999.0498. [DOI] [PubMed] [Google Scholar]
- 152.Sferruzzi-Perri AN, Owens JA, Pringle KG, Robinson JS, Roberts CT. Insulin-like growth factors-I and -II act via different pathways to promote fetal growth. Endocrinology. 2006;147:3344–3355. doi: 10.1210/en.2005-1328. [DOI] [PubMed] [Google Scholar]
- 153.Qiu Q, Basak A, Mbikay M, Tsang BK, Gruslin A. Role of pro-IGF-II processing by proprotein convertase 4 in human placental development. Proc Natl Acad Sci U S A. 2005;102:11047–11052. doi: 10.1073/pnas.0502357102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Van Mieghem T, van Bree R, Van Herck E, Deprest J, Verhaeghe J. Insulin-like growth factor-II regulates maternal hemodynamic adaptation to pregnancy in rats. Am J Physiol Regul Integr Comp Physiol. 2009;297:R1615–R1621. doi: 10.1152/ajpregu.00463.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Nielsen J, Jensen RB, Juul A. Growth hormone deficiency in children. Ugeskr Laeger. 2015;177:1260–1263. [PubMed] [Google Scholar]
- 156.Murray PG, Clayton PE. Disorders of Growth Hormone in Childhood. In: De Groot LJ, Beck-Peccoz P, Chrousos G, Dungan K, Grossman A, Hershman JM, Koch C, McLachlan R, New M, Rebar R, Singer F, Vinik A, Weickert MO, editors. Endotext [Internet] South Dartmouth (MA): MDText.com, Inc.; 2000–2013. [Google Scholar]
- 157.Fan Y, Menon RK, Cohen P, et al. Liver-specific deletion of the growth hormone receptor reveals essential role of growth hormone signaling in hepatic lipid metabolism. J Biol Chem. 2009;284:19937–19944. doi: 10.1074/jbc.M109.014308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Sos BC, Harris C, Nordstrom SM, et al. Abrogation of growth hormone secretion rescues fatty liver in mice with hepatocyte-specific deletion of JAK2. J Clin Invest. 2011;121:1412–1423. doi: 10.1172/JCI42894. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Cui Y, Hosui A, Sun R, et al. Loss of signal transducer and activator of transcription 5 leads to hepatosteatosis and impaired liver regeneration. Hepatology. 2007;46:504–513. doi: 10.1002/hep.21713. [DOI] [PubMed] [Google Scholar]
- 160.Nishizawa H, Iguchi G, Murawaki A, et al. Nonalcoholic fatty liver disease in adult hypopituitary patients with GH deficiency and the impact of GH replacement therapy. Eur J Endocrinol. 2012;167:67–74. doi: 10.1530/EJE-12-0252. [DOI] [PubMed] [Google Scholar]
- 161.Nakamura T, Nawa K, Ichihara A. Partial purification and characterization of hepatocyte growth factor from serum of hepatectomized rats. Biochem Biophys Res Commun. 1984;122:1450–1459. doi: 10.1016/0006-291x(84)91253-1. [DOI] [PubMed] [Google Scholar]
- 162.Nakamura T. Structure and function of hepatocyte growth factor. Prog Growth Factor Res. 1991;3:67–85. doi: 10.1016/0955-2235(91)90014-u. [DOI] [PubMed] [Google Scholar]
- 163.Matsumoto K, Nakamura T. Hepatocyte growth factor: molecular structure, roles in liver regeneration, and other biological functions. Crit Rev Oncog. 1992;3:27–54. [PubMed] [Google Scholar]
- 164.Birchmeier C, Gherardi E. Developmental roles of HGF/SF and its receptor, the c-Met tyrosine kinase. Trends Cell Biol. 1998;8:404–410. doi: 10.1016/s0962-8924(98)01359-2. [DOI] [PubMed] [Google Scholar]
- 165.Fausto N, Campbell JS, Riehle KJ. Liver regeneration. Hepatology. 2006;43(Suppl 1):S45–S53. doi: 10.1002/hep.20969. [DOI] [PubMed] [Google Scholar]
- 166.Kinoshita T, Hirao S, Matsumoto K, Nakamura T. Possible endocrine control by hepatocyte growth factor of liver regeneration after partial hepatectomy. Biochem Biophys Res Commun. 1991;17:330–335. doi: 10.1016/0006-291x(91)91987-n. [DOI] [PubMed] [Google Scholar]
- 167.Mizuno S, Nakamura T. Hepatocyte growth factor: a regenerative drug for acute hepatitis and liver cirrhosis. Regen Med. 2007;2:161–170. doi: 10.2217/17460751.2.2.161. [DOI] [PubMed] [Google Scholar]
- 168.Nakamura T, Mizuno S. The discovery of Hepatocyte Growth Factor (HGF) and its significance for cell biology, life sciences and clinical medicine. Proc Jpn Acad Ser B Phys Biol Sci. 2010;86:588–610. doi: 10.2183/pjab.86.588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Ekberg S, Luther M, Nakamura T, Jansson JO. Growth hormone promotes early initiation of hepatocyte growth factor gene expression in the liver of hypophysectomized rats after partial hepatectomy. J Endocrinol. 1992;135:59–67. doi: 10.1677/joe.0.1350059. [DOI] [PubMed] [Google Scholar]
- 170.Balducci R, Toscano V, Larizza D, et al. Effects of long-term growth hormone therapy on adrenal steroidogenesis in Turner syndrome. Horm Res. 1998;49:210–215. doi: 10.1159/000023173. [DOI] [PubMed] [Google Scholar]
- 171.Isidori AM, Kaltsas GA, Perry L, Burrin JM, Besser GM, Monson JP. The effect of growth hormone replacement therapy on adrenal androgen secretion in adult onset hypopituitarism. Clin Endocrinol (Oxf) 2003;58:601–611. doi: 10.1046/j.1365-2265.2003.01759.x. [DOI] [PubMed] [Google Scholar]
- 172.McFarlane AC, Edmondson SR, Wintour EM, Werther GA. GH-receptor distribution in the ovine foetal adrenal gland: ontogenic and functional studies. J Endocrinol. 1999;162:197–205. doi: 10.1677/joe.0.1620197. [DOI] [PubMed] [Google Scholar]
- 173.Devaskar UP, Devaskar SU, Voina S, Velayo N, Sperling MA. Growth hormone stimulates adrenal steroidogenesis in the fetus. Nature. 1981;290:404–405. doi: 10.1038/290404a0. [DOI] [PubMed] [Google Scholar]
- 174.Saccà L, Cittadini A, Fazio S. Growth hormone and the heart. Endocr Rev. 1994;15:555–573. doi: 10.1210/edrv-15-5-555. [DOI] [PubMed] [Google Scholar]
- 175.Li Q, Li B, Wang X, et al. Overexpression of insulin-like growth factor-I in mice protects from myocyte death after infarction, attenuating ventricular dilation, wall stress and cardiac hypertrophy. J Clin Invest. 1997;100:1991–1999. doi: 10.1172/JCI119730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Timsit J, Riou B, Bertherat J, et al. Effects of chronic growth hormone hypersecretion on intrinsic contractility, energetics, isomyosin pattern, and myosin adenosine triphosphatase activity of rat left ventricle. J Clin Invest. 1990;86:507–515. doi: 10.1172/JCI114737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Cittadini A, Ishiguro Y, Stromer H, et al. Insulin-like growth factor-1 but not growth hormone augments mammalian myocardial contractility by sensitizing the myofilament to Ca2+ through a wortmannin-sensitive pathway: studies in rat and ferret isolated muscles. Circ Res. 1998;83:50–59. doi: 10.1161/01.res.83.1.50. [DOI] [PubMed] [Google Scholar]
- 178.Mayoux E, Ventura-Clapier R, Timsit J, Behar-Cohen F, Hoffmann C, Mercadier JJ. Mechanical properties of rat cardiac skinned fibers are altered by chronic growth hormone hypersecretion. Circ Res. 1993;72:57–64. doi: 10.1161/01.res.72.1.57. [DOI] [PubMed] [Google Scholar]
- 179.Bruel A. Oxlund Biosynthetic growth hormone increase the collagen deposition rate in rat aorta and heart. Eur J Endocrinol. 1995;132:195–199. doi: 10.1530/eje.0.1320195. [DOI] [PubMed] [Google Scholar]
- 180.Buerke M, Murohara T, Skurk C, Nuss C, Tomaselli K, Lefer AM. Cardioprotective effect of insulin-like growth factor I in myocardial ischemia followed by reperfusion. Proc Natl Acad Sci U S A. 1995;92:8031–8035. doi: 10.1073/pnas.92.17.8031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Ren J. Short-term administration of insulin-like growth factor (IGF-1) does not induce myocardial IGF-1 resistance. Growth Horm IGF Res. 2002;12:162–168. doi: 10.1016/s1096-6374(02)00020-5. [DOI] [PubMed] [Google Scholar]
- 182.Longobardi S, Cuocolo A, Merola B, et al. Left ventricular function in young adults with childhood and adulthood onset growth hormone deficiency. Clin Endocrinol. 1998;48:137–143. doi: 10.1046/j.1365-2265.1998.00281.x. [DOI] [PubMed] [Google Scholar]
- 183.Thuesen L, Jorgensen JOL, Muller JR, et al. Short and long-term cardiovascular effects of growth hormone deficient adults. Clin Endocrinol. 1994;41:615–650. doi: 10.1111/j.1365-2265.1994.tb01827.x. [DOI] [PubMed] [Google Scholar]
- 184.Valcavi R, Gaddi O, Zini M, Iavicoli M, Mellino U, Portioli I. Cardiac performance and mass in adult with hypopituitarism: effects of one year of growth treatment. J Clin Endocrinol Metab. 1995;80:659–666. doi: 10.1210/jcem.80.2.7852533. [DOI] [PubMed] [Google Scholar]
- 185.Johannsson G, Bengtsson BA, Andersson B, Isgaard J, Caidahl K. Long-term cardiovascular effects of growth hormone treatment in GH-deficient adults: preliminary data in a small group of patients. Clin Endocrinol. 1996;45:305–314. doi: 10.1046/j.1365-2265.1996.00820.x. [DOI] [PubMed] [Google Scholar]
- 186.Cuneo RC, Salomon F, Wiles CM, Hesp R, Sonksen PH. Growth hormone treatment in growth hormone deficient adults. I. Effects on muscle mass and strength. J Appl Physiol. 1991;70:688–694. doi: 10.1152/jappl.1991.70.2.688. [DOI] [PubMed] [Google Scholar]
- 187.Cuneo RC, Salomon F, Wiles CM, Hesp R, Sonksen PH. Growth hormone treatment in growth hormone deficient adults. II. Effects on exercise performance. J Appl Physiol. 1991;70:695–670. doi: 10.1152/jappl.1991.70.2.695. [DOI] [PubMed] [Google Scholar]
- 188.Colao A, Cuocolo A, Di Somma C, et al. Impaired cardiac performance in elderly patients with growth hormone deficiency. J Clin Endocrinol Metab. 1999;85:3950–3955. doi: 10.1210/jcem.84.11.6112. [DOI] [PubMed] [Google Scholar]
- 189.Colao A, Di Somma C, Cuocolo A, et al. The severity of GH deficiency (GHD) correlates with the severity of cardiac impairment in 100 adult patients with hypopituitarism: an observational, case-control study. J Clin Endocrinol Metab. 2004;89:5998–6004. doi: 10.1210/jc.2004-1042. [DOI] [PubMed] [Google Scholar]
- 190.Maison P, Chanson P. Cardiac effects of growth hormone in adults with growth hormone deficiency. Circulation. 2003;108:2648–2652. doi: 10.1161/01.CIR.0000100720.01867.1D. [DOI] [PubMed] [Google Scholar]
- 191.Walsh MF, Barazi M, Pete G, Muniyappa R, Dunbar JC, Sowers JR. Insulin-like growth factor I diminishes in vivo and in vitro vascular contractility: role of vascular nitric oxide. Endocrinology. 1996;137:1798–1803. doi: 10.1210/endo.137.5.8612517. [DOI] [PubMed] [Google Scholar]
- 192.Tsukahara H, Gordienko DV, Tonshoff B, Gelato MC, Goligorsky MS. Direct demonstration of insulin-like growth factor-I-induced nitric oxide production by endothelial cells. Kidney Int. 1994;45:598–604. doi: 10.1038/ki.1994.78. [DOI] [PubMed] [Google Scholar]
- 193.Muniyappa R, Walsh MF, Rangi JS, et al. Insulin like growth factor 1 increases vascular smooth muscle nitric oxide production. Life Sci. 1997;61:925–931. doi: 10.1016/s0024-3205(97)00594-8. [DOI] [PubMed] [Google Scholar]
- 194.Haylor J, Singh I, Nahas AM. Nitric oxide synthesis inhibitor prevents vasodilatation by insulin-like growth factor I. Kidney Int. 1991;39:333–335. doi: 10.1038/ki.1991.42. [DOI] [PubMed] [Google Scholar]
- 195.Capaldo B, Guardasole V, Pardo F, et al. Abnormal vascular reactivity in growth hormone deficiency. Circulation. 2001;103:520–524. doi: 10.1161/01.cir.103.4.520. [DOI] [PubMed] [Google Scholar]
- 196.Napoli R, Guardasole V, Matarazzo M, et al. Growth hormone corrects vascular dysfunction in patients with chronic heart failure. J Am Coll Cardiol. 2002;39:90–95. doi: 10.1016/s0735-1097(01)01707-7. [DOI] [PubMed] [Google Scholar]
- 197.Tivesten A, Barlind A, Caidahl K, et al. Growth hormone-induced blood pressure decrease is associated with increased mRNA levels of the vascular smooth muscle KATP channel. J Endocrinol. 2004;183:195–202. doi: 10.1677/joe.1.05726. [DOI] [PubMed] [Google Scholar]
- 198.Sverrisdóttir Y, Elam M, Bengtsson B-Å, Johannsson G. Intense sympathetic nerve activity in adults with hypopituitarism and untreated growth hormone deficiency. J Clin Endocrinol Metab. 1998;83:1881–1885. doi: 10.1210/jcem.83.6.4895. [DOI] [PubMed] [Google Scholar]
- 199.Sverrisdottir Y, Elam M, Caidahl K, Söderling AS, Herlitz H, Johannsson G. The effect of growth hormone (GH) replacement therapy on sympathetic nerve hyperactivity in hypopituitary adults: a double-blind, placebo-controlled, crossover, short-term trial followed by long-term open GH replacement in hypopituitary adults. J Hypertens. 2003;10:1905–1914. doi: 10.1097/01.hjh.0000084757.37215.55. [DOI] [PubMed] [Google Scholar]
- 200.Lobie PE, Breipohl W, Aragon JG, Waters MJ. Cellular localization of the growth hormone receptor/binding protein in the male and female reproductive systems. Endocrinology. 1990;126:2214–2221. doi: 10.1210/endo-126-4-2214. [DOI] [PubMed] [Google Scholar]
- 201.Sharara FI, Nieman LK. Identification and cellular localization of growth hormone receptor gene expression in the human ovary. J Clin Endocrinol Metab. 1994;79:670–672. doi: 10.1210/jcem.79.2.7519196. [DOI] [PubMed] [Google Scholar]
- 202.Temmim L, Kolle S, Baker H, Sinowatz F. Expression of growth hormone receptor in human liposarcomas and lipomas. Oncol Rep. 2000;7:757–760. doi: 10.3892/or.7.4.757. [DOI] [PubMed] [Google Scholar]
- 203.Werther GA, Haynes K, Waters MJ. Growth hormone (GH) receptors are expressed on human fetal mesenchymal tissues—identification of messenger ribonucleic acid and GH-binding protein. J Clin Endocrinol Metab. 1993;76:1638–1646. doi: 10.1210/jcem.76.6.7684746. [DOI] [PubMed] [Google Scholar]
- 204.Li G, Del Rincon JP, Jahn LA, et al. Growth hormone exerts acute vascular effects independent of systemic or muscle insulin-like growth factor I. J Clin Endocrinol Metab. 2008;93:1379–1385. doi: 10.1210/jc.2007-2021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 205.Thum T, Tsikas D, Frolich JC, Borlak J. Growth hormone induces eNOS expression and nitric oxide release in a cultured human endothelial cell line. FEBS Lett. 2003;555:567–571. doi: 10.1016/s0014-5793(03)01356-5. [DOI] [PubMed] [Google Scholar]
- 206.Rymaszewski Z, Cohen RM, Chomczynski P. Human growth hormone stimulates proliferation of human retinal microvascular endothelial cells in vitro. Proc Natl Acad Sci U S A. 1991;88:617–621. doi: 10.1073/pnas.88.2.617. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 207.Struman I, Bentzien F, Lee H, et al. Opposing actions of intact and N-terminal fragments of the human prolactin/growth hormone family members on angiogenesis: an efficient mechanism for the regulation of angiogenesis. Proc Natl Acad Sci U S A. 1999;96:1246–1251. doi: 10.1073/pnas.96.4.1246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 208.Brunet-Dunand SE, Vouyovitch C, Araneda S, et al. Autocrine human growth hormone promotes tumor angiogenesis in mammary carcinoma. Endocrinology. 2009;150:1341–1352. doi: 10.1210/en.2008-0608. [DOI] [PubMed] [Google Scholar]
- 209.Frerich B, Kurtz-Hoffmann J, Lindemann N. Influence of growth hormone on maintenance of capillary-like structures in an in vitro model of stromal vascular tissue: results from morphometric analysis. Artif Organs. 2005;29:338–341. doi: 10.1111/j.1525-1594.2005.29057.x. [DOI] [PubMed] [Google Scholar]
- 210.Sonntag WE, Lynch CD, Cooney PT, Hutchins PM. Decreases in cerebral microvasculature with age are associated with the decline in growth hormone and insulin-like growth factor 1. Endocrinology. 1997;138:3515–3520. doi: 10.1210/endo.138.8.5330. [DOI] [PubMed] [Google Scholar]
- 211.Kusano K, Tsutsumi Y, Dean J, et al. Long-term stable expression of human growth hormone by rAAV promotes myocardial protection post-myocardial infarction. J Mol Cell Cardiol. 2007;42:390–399. doi: 10.1016/j.yjmcc.2006.10.016. [DOI] [PubMed] [Google Scholar]
- 212.Rong SL, Lu YX, Liao YH, et al. Effects of transplanted myoblasts transfected with human growth hormone gene on improvement of ventricular function of rats. Chin Med J. 2008;121:347–354. [PubMed] [Google Scholar]
- 213.Garcia-Esteo F, Pascual G, Gallardo A, San-Roman J, Bujan J, Bellon JM. A biodegradable copolymer for the slow release of growth hormone expedites scarring in diabetic rats. J Biomed Mater Res B Appl Biomater. 2007;81:291–304. doi: 10.1002/jbm.b.30665. [DOI] [PubMed] [Google Scholar]
- 214.Thorey IS, Hinz B, Hoeflich A, et al. Transgenic mice reveal novel activities of growth hormone in wound repair, angiogenesis, and myofibroblast differentiation. J Biol Chem. 2004;279:26674–26684. doi: 10.1074/jbc.M311467200. [DOI] [PubMed] [Google Scholar]
- 215.Devin JK, Vaughan DE, Blevins LS, Jr, et al. Low-dose growth hormone administration mobilizes endothelial progenitor cells in healthy adults. Growth Horm IGF Res. 2008;18:253–263. doi: 10.1016/j.ghir.2007.11.001. [DOI] [PubMed] [Google Scholar]
- 216.Thum T, Hoeber S, Froese S, et al. Age-dependent impairment of endothelial progenitor cells is corrected by growth-hormone-mediated increase of insulin-like growth-factor-1. Circ Res. 2007;100:434–443. doi: 10.1161/01.RES.0000257912.78915.af. [DOI] [PubMed] [Google Scholar]
- 217.Oomen PH, Beentjes JA, Bosma E, Smit AJ, Reitsma WD, Dullaart RP. Reduced capillary permeability and capillary density in the skin of GH-deficient adults: improvement after 12 months GH replacement. Clin Endocrinol. 2002;56:519–524. doi: 10.1046/j.1365-2265.2002.01517.x. [DOI] [PubMed] [Google Scholar]
- 218.Hellstrom A, Carlsson B, Niklasson A, et al. IGF-I is critical for normal vascularization of the human retina. J Clin Endocrinol Metab. 2002;87:3413–3416. doi: 10.1210/jcem.87.7.8629. [DOI] [PubMed] [Google Scholar]
- 219.Hellstrom A, Svensson E, Carlsson B, Niklasson A, Albertsson-Wikland K. Reduced retinal vascularization in children with growth hormone deficiency. J Clin Endocrinol Metab. 1999;84:795–798. doi: 10.1210/jcem.84.2.5484. [DOI] [PubMed] [Google Scholar]
- 220.Delafontaine P, Song YH, Li Y. Expression, regulation, and function of IGF-1, IGF-1R, and IGF-1 binding proteins in blood vessels. Arterioscler Thromb Vasc Biol. 2004;24:435–444. doi: 10.1161/01.ATV.0000105902.89459.09. [DOI] [PubMed] [Google Scholar]
- 221.Li G, Del Rincon JP, Jahn LA, et al. Growth hormone exerts acute vascular effects independent of systemic or muscle insulin-like growth factor I. J Clin Endocrinol Metab. 2008;93:1379–1385. doi: 10.1210/jc.2007-2021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 222.Napoli R, Guardasole V, Angelini V, et al. Acute effects of growth hormone on vascular function in human subjects. J Clin Endocrinol Metab. 2003;88:2817–2820. doi: 10.1210/jc.2003-030144. [DOI] [PubMed] [Google Scholar]
- 223.Castellano G, Affuso F, Di Conza P, Fazio S. The GH/IGF-I Axis and Heart Failure. Curr Cardiol Rev. 2009;5:203–215. doi: 10.2174/157340309788970306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 224.Isgaard J, Arcopinto M, Karason K, Cittadini A. GH and the cardiovascular system: an update on a topic at heart. Endocrine. 2015;48:25–35. doi: 10.1007/s12020-014-0327-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 225.Clapp C, Thebault S, Jeziorski MC, Martínez de la Escalera G. Peptide Hormone Regulation of Angiogenesis. Physiol Rev. 2009;89:1177–1215. doi: 10.1152/physrev.00024.2009. [DOI] [PubMed] [Google Scholar]
- 226.Møller N, Jørgensen JO. Effects of growth hormone on glucose, lipid, and protein metabolism in human subjects. Endocr Rev. 2009;30:152–177. doi: 10.1210/er.2008-0027. [DOI] [PubMed] [Google Scholar]
- 227.Beshyah SA, Freemantle C, Thomas E, et al. Abnormal body composition and reduced bone mass in growth hormone deficient hypopituitary adults. Clin Endocrinol (Oxf) 1995;42:179–189. doi: 10.1111/j.1365-2265.1995.tb01860.x. [DOI] [PubMed] [Google Scholar]
- 228.Johansen T, Richelsen B, Hansen HS, Din N, Malmlöf K. Growth hormone-mediated breakdown of body fat: effects of GH on lipases in adipose tissue and skeletal muscle of old rats fed different diets. Horm Metab Res. 2003;35:243–250. doi: 10.1055/s-2003-39481. [DOI] [PubMed] [Google Scholar]
- 229.List EO, Berryman DE, Funk K, et al. The role of GH in adipose tissue: lessons from adipose-specific GH receptor gene-disrupted mice. Mol Endocrinol. 2013;27:524–535. doi: 10.1210/me.2012-1330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 230.Berryman DE, Glad CA, List EO, Johannsson G. The GH/IGF-1 axis in obesity: pathophysiology and therapeutic considerations. Nat Rev Endocrinol. 2013;9:346–356. doi: 10.1038/nrendo.2013.64. [DOI] [PubMed] [Google Scholar]
- 231.Baum HB, Biller BM, Finkelstein JS, et al. Effects of physiologic growth hormone therapy on bone density and body composition in patients with adult-onset growth hormone deficiency. A randomized, placebo-controlled trial. Ann Intern Med. 1996;125:883–890. doi: 10.7326/0003-4819-125-11-199612010-00003. [DOI] [PubMed] [Google Scholar]
- 232.Ullman M, Oldfors A. Effects of growth hormone on skeletal muscle I. Studies on normal adult rats. Acta Physiol Scand. 1989;135:531–536. doi: 10.1111/j.1748-1716.1989.tb08612.x. [DOI] [PubMed] [Google Scholar]
- 233.Weber MM. Effects of growth hormone on skeletal muscle. Horm Res. 2002;58(suppl 3):43–48. doi: 10.1159/000066482. [DOI] [PubMed] [Google Scholar]
- 234.Sotiropoulos A, Ohanna M, Kedzia C, et al. Growth hormone promotes skeletal muscle cell fusion independent of insulin-like growth factor 1 up-regulation. Proc Natl Acad Sci U S A. 2006;103:7315–7320. doi: 10.1073/pnas.0510033103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 235.Mavalli MD, DiGirolamo DJ, Fan Y, et al. Distinct growth hormone receptor signaling modes regulate skeletal muscle development and insulin sensitivity in mice. J Clin Invest. 2010;120:4007–4020. doi: 10.1172/JCI42447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 236.Clasen BFF, Krusenstjerna-Hafstrøm T, Holm Vendelbo M, et al. Gene expression in skeletal muscle after an acute intravenous GH bolus in human subjects: identification of a mechanism regulating ANGPTL4. J Lipid Res. 2013;54:1988–1997. doi: 10.1194/jlr.P034520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 237.Tavares AB, Micmacher E, Biesek S, et al. Effects of Growth Hormone Administration on Muscle Strength in Men over 50 Years Old. Int J Endocrinol. 2013;2013:942030–942036. doi: 10.1155/2013/942030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 238.Ríos R, Carneiro I, Arce VM, Devesa J. Myostatin is an inhibitor of myogenic differentiation. Am J Physiol Cell Physiol. 2002;282:C993–C999. doi: 10.1152/ajpcell.00372.2001. [DOI] [PubMed] [Google Scholar]
- 239.Langley B, Thomas M, Bishop A, Sharma M, Gilmour S, Kambadur R. Myostatin inhibits myoblast differentiation by down-regulating MyoD expression. Journal of Biological Chemistry. 2002;277:49831–49840. doi: 10.1074/jbc.M204291200. [DOI] [PubMed] [Google Scholar]
- 240.Thomas M, Langley B, Berry C, et al. Myostatin, a negative regulator of muscle growth, functions by inhibiting myoblast proliferation. Journal of Biological Chemistry. 2000;275:40235–40243. doi: 10.1074/jbc.M004356200. [DOI] [PubMed] [Google Scholar]
- 241.Trendelenburg AU, Meyer A, Rohner D, Boyle J, Hatakeyama S, Glass DJ. Myostatin reduces Akt/TORC1/p70S6K signaling, inhibiting myoblast differentiation and myotube size. American Journal of Physiology. 2009;296:C1258–C1270. doi: 10.1152/ajpcell.00105.2009. [DOI] [PubMed] [Google Scholar]
- 242.Yakar S, Courtland HW, Clemmons D. IGF-1 and bone: New discoveries from mouse models. J Bone Miner Res. 2010;25:2543–2552. doi: 10.1002/jbmr.234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 243.Yakar S, Liu JL, Stannard B, et al. Normal growth and development in the absence of hepatic insulin-like growth factor I. Proc Natl Acad Sci U S A. 1999;96:7324–7329. doi: 10.1073/pnas.96.13.7324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 244.Yakar S, Rosen CJ, Beamer WG, et al. Circulating levels of IGF-1 directly regulate bone growth and density. J Clin Invest. 2002;110:771–781. doi: 10.1172/JCI15463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 245.Edwall D, Schalling M, Jennische E, Norstedt G. Induction of insulin-like growth factor I messenger ribonucleic acid during regeneration of rat skeletal muscle. Endocrinology. 1989;124:820–825. doi: 10.1210/endo-124-2-820. [DOI] [PubMed] [Google Scholar]
- 246.Bikle DD, Sakata T, Leary C, et al. Insulin-like growth factor I is required for the anabolic actions of parathyroid hormone on mouse bone. J Bone Miner Res. 2002;17:1570–1578. doi: 10.1359/jbmr.2002.17.9.1570. [DOI] [PubMed] [Google Scholar]
- 247.Zhao G, Monier-Faugere MC, Langub MC, et al. Targeted overexpression of insulin-like growth factor I to osteoblasts of transgenic mice: increased trabecular bone volume without increased osteoblast proliferation. Endocrinology. 2000;141:2674–2682. doi: 10.1210/endo.141.7.7585. [DOI] [PubMed] [Google Scholar]
- 248.Govoni KE, Wergedal JE, Florin L, Angel P, Baylink DJ, Mohan S. Conditional deletion of insulin-like growth factor-I in collagen type 1α2-expressing cells results in postnatal lethality and a dramatic reduction in bone accretion. Endocrinology. 2007;148:5706–5715. doi: 10.1210/en.2007-0608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 249.Hirschberg R, Kopple JD. Effects of growth hormone and IGF-I on renal function. Kidney Int Suppl. 1989;27:S20–S26. [PubMed] [Google Scholar]
- 250.Bach LA, Hale LJ. Insulin-like growth factors and kidney disease. Am J Kidney Dis. 2015;65:327–336. doi: 10.1053/j.ajkd.2014.05.024. [DOI] [PubMed] [Google Scholar]
- 251.Müller-Wiefel D, Frisch H, Tulassay T, Bell L, Zadik Z. Treatment of growth failure with growth hormone in children with chronic kidney disease: an open-label long-term study. Clin Nephrol. 2010;74:97–105. doi: 10.5414/cnp74097. [DOI] [PubMed] [Google Scholar]
- 252.Sicuro A, Mahlbacher K, Hulter HN, Krapf R. Effect of growth hormone on renal and systemic acid-base homeostasis in humans. Am J Physiol. 1998;274:F650–F657. doi: 10.1152/ajprenal.1998.274.4.F650. [DOI] [PubMed] [Google Scholar]
- 253.Meazza C, Bonomelli I, Pagani S, et al. Effect of human recombinant growth hormone therapy on circulating levels of erythropoietin and granulocyte-colony stimulating factor in short children. J Pediatr Endocrinol Metab. 2009;22:837–843. doi: 10.1515/jpem.2009.22.9.837. [DOI] [PubMed] [Google Scholar]
- 254.Sohmiya M, Kato Y. Effect of long-term administration of recombinant human growth hormone (rhGH) on plasma erythropoietin (EPO) and haemoglobin levels in anaemic patients with adult GH deficiency. Clin Endocrinol (Oxf) 2001;55:749–754. doi: 10.1046/j.1365-2265.2001.01417.x. [DOI] [PubMed] [Google Scholar]
- 255.Sohmiya M, Kanazawa I, Kato Y. Effect of recombinant human GH on circulating granulocyte colony-stimulating factor and neutrophils in patients with adult GH deficiency. Eur J Endocrinol. 2005;152:211–215. doi: 10.1530/eje.1.01831. [DOI] [PubMed] [Google Scholar]
- 256.Esposito A, Capalbo D, De Martino L, et al. Long-term effects of growth hormone (GH) replacement therapy on hematopoiesis in a large cohort of children with GH deficiency. Endocrine. 2016;53:192–198. doi: 10.1007/s12020-015-0781-9. [DOI] [PubMed] [Google Scholar]
- 257.Carlo-Stella C, Di Nicola M, Milani R, et al. Age- and irradiation-associated loss of bone marrow hematopoietic function in mice is reversed by recombinant human growth hormone. Exp Hematol. 2004;32:171–178. doi: 10.1016/j.exphem.2003.11.007. [DOI] [PubMed] [Google Scholar]
- 258.Weigent DA. Lymphocyte GH-axis hormones in immunity. Cell Immunol. 2013;285:118–132. doi: 10.1016/j.cellimm.2013.10.003. [DOI] [PubMed] [Google Scholar]
- 259.Liu QL, Zhang J, Liu X, Gao JY. Role of growth hormone in maturation and activation of dendritic cells via miR-200a and the Keap1/Nrf2 pathway. Cell Prolif. 2015;48:573–581. doi: 10.1111/cpr.12206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 260.Weigent DA, Blalock JE, LeBoeuf RD. An antisense oligodeoxynucleotide to growth hormone messenger ribonucleic acid inhibits lymphocyte proliferation. Endocrinology. 1991;128:2053–2057. doi: 10.1210/endo-128-4-2053. [DOI] [PubMed] [Google Scholar]
- 261.Baxter JB, Blalock JE, Weigent DA. Characterization of immunoreactive insulin-like growth factor-I from leukocytes and its regulation by growth hormone. Endocrinology. 1991;129:1727–1734. doi: 10.1210/endo-129-4-1727. [DOI] [PubMed] [Google Scholar]
- 262.Arnold RE, Weigent DA. The inhibition of superoxide production in EL4 lymphoma cells overexpressing growth hormone. Immunopharm Immunotoxicol. 2003;25:159–177. doi: 10.1081/iph-120020467. [DOI] [PubMed] [Google Scholar]
- 263.Arnold RE, Weigent DA. The production of nitric oxide in EL4 lymphoma cells overexpressing growth hormone. J Neuroimmunol. 2003;134:82–94. doi: 10.1016/s0165-5728(02)00420-4. [DOI] [PubMed] [Google Scholar]
- 264.Arnold RE, Weigent DA. The inhibition of apoptosis in EL4 lymphoma cells overexpressing growth hormone. Neuroimmunomodulation. 2004;11:149–159. doi: 10.1159/000076764. [DOI] [PubMed] [Google Scholar]
- 265.Weigent DA, Arnold RE. Expression of insulin-like growth factor-1 and insulin-like growth factor-1 receptors in EL4 lymphoma cells overexpressing growth hormone. Cell Immunol. 2005;234:54–66. doi: 10.1016/j.cellimm.2005.04.016. [DOI] [PubMed] [Google Scholar]
- 266.Farmer JT, Weigent DA. Expression of insulin-like growth factor-2 receptors on EL4 cells overexpressing growth hormone. Brain Behav Immun. 2007;21:79–85. doi: 10.1016/j.bbi.2006.02.006. [DOI] [PubMed] [Google Scholar]
- 267.Farmer JT, Weigent DA. TGF-beta-1 expression in EL4 lymphoma cells overexpressing growth hormone. Cell Immunol. 2006;240:22–30. doi: 10.1016/j.cellimm.2006.06.003. [DOI] [PubMed] [Google Scholar]
- 268.Weigent DA. Regulation of Id2 expression in EL4 T lymphoma cells overexpressing growth hormone. Cell Immunol. 2009;255:46–54. doi: 10.1016/j.cellimm.2008.10.003. [DOI] [PubMed] [Google Scholar]
- 269.Malarkey WB, Wang J, Cheney C, Glaser R, Nagaraja H. Human lymphocyte growth hormone stimulates interferon gamma production and is inhibited by cortisol and norepinephrine. J Neuroimmunol. 2002;123:180–187. doi: 10.1016/s0165-5728(01)00489-1. [DOI] [PubMed] [Google Scholar]
- 270.Kao TL, Harbour DV, Meyer WJ. 3d. Immunoreactive growth hormone production by cultured lymphocytes. Ann NY Acad Sci. 1992;650:179–181. doi: 10.1111/j.1749-6632.1992.tb49117.x. [DOI] [PubMed] [Google Scholar]
- 271.Weigent DA, Baxter JB, Blalock JE. The production of growth hormone and insulin-like growth factor-I by the same subpopulation of rat mononuclear leukocytes. Brain Behav Immun. 1992;6:365–376. doi: 10.1016/0889-1591(92)90035-m. [DOI] [PubMed] [Google Scholar]
- 272.Re RN, Cook JL. Senescence, apoptosis, and stem cell biology: the rationale for an expanded view of intracrine action. Am J Physiol Heart Circ Physiology. 2009;297:H893–H901. doi: 10.1152/ajpheart.00414.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 273.Weigent DA. High Molecular Weight Isoforms of Growth Hormone In Cells of the Immune System. Cell Immunol. 2011;271:44–52. doi: 10.1016/j.cellimm.2011.06.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 274.Lobie PE, Garcia-Aragon J, Lincoln DT, Barnard R, Wilcox JN, Waters MJ. Localization and ontogeny of growth hormone receptor gene expression in the central nervous system. Brain Des Dev Brain Res. 1993;74:225–233. doi: 10.1016/0165-3806(93)90008-x. [DOI] [PubMed] [Google Scholar]
- 275.Harvey S, Kakebeeke M, Murphy AE, Sanders EJ. Growth hormone in the nervous system: autocrine or paracrine roles in retinal function? Can J Physiol Pharmacol. 2003;81:371–384. doi: 10.1139/y03-034. [DOI] [PubMed] [Google Scholar]
- 276.Laron Z, Galatzer A. Growth hormone, somatomedin and prolactin—relationship to brain function. Brain Dev. 1985;7:559–567. doi: 10.1016/s0387-7604(85)80001-2. [DOI] [PubMed] [Google Scholar]
- 277.Nyberg F, Burman P. Growth hormone and its receptors in the central nervous system—location and functional significance. Horm Res. 1996;45:18–22. doi: 10.1159/000184753. [DOI] [PubMed] [Google Scholar]
- 278.Lobie PE, Zhu T, Graichen R, Goh EL. Growth hormone, insulin-like growth factor I and the CNS: localization, function and mechanism of action. Growth Hormone and IGF Research. 2000;10(Suppl B):S51–S56. doi: 10.1016/s1096-6374(00)80010-6. [DOI] [PubMed] [Google Scholar]
- 279.Pathipati P, Gorba T, Scheepens A, Goffin V, Sun Y, Fraser M. Growth hormone and prolactin regulate human neural stem cell regenerative activity. Neuroscience. 2011;190:409–427. doi: 10.1016/j.neuroscience.2011.05.029. [DOI] [PubMed] [Google Scholar]
- 280.Scheepens A, Williams CE, Breier BH, Guan J, Gluckman PD. A role for the somatotropic axis in neural development, injury and disease. J Pediatr Endocrinol Metab. 2000;13(Suppl 6):1483–1491. doi: 10.1515/jpem-2000-s623. [DOI] [PubMed] [Google Scholar]
- 281.García-Aragón J, Lobie PE, Muscat GE, Gobius KS, Norstedt G, Waters MJ. Prenatal expression of the growth hormone (GH) receptor/binding protein in the rat: a role for GH in embryonic and fetal development? Development. 1992;114:869–876. doi: 10.1242/dev.114.4.869. [DOI] [PubMed] [Google Scholar]
- 282.Turnley AM, Faux CH, Rietze RL, Coonan JR, Bartlett PF. Suppressor of cytokine signaling 2 regulates neuronal differentiation by inhibiting growth hormone signaling. Nat Neurosci. 2002;5:1155–1162. doi: 10.1038/nn954. [DOI] [PubMed] [Google Scholar]
- 283.Donahue CP, Jensen RV, Ochiishi T, et al. Transcriptional profiling reveals regulated genes in the hippocampus during memory formation. Hippocampus. 2002;12:821–833. doi: 10.1002/hipo.10058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 284.Donahue CP, Kosik KS, Shors TJ. Growth hormone is produced within the hippocampus where it responds to age, sex, and stress. Proc Natl Acad Sci U S A. 2006;103:6031–6036. doi: 10.1073/pnas.0507776103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 285.Sun LY, Al-Regaiey K, Masternak MM, Wang J, Bartke A. Local expression of GH and IGF-1 in the hippocampus of GH-deficient long-lived mice. Neurobiol Aging. 2005;26:929–937. doi: 10.1016/j.neurobiolaging.2004.07.010. [DOI] [PubMed] [Google Scholar]
- 286.Sun LY, Evans MS, Hsieh J, Panici J, Bartke A. Increased neurogenesis in dentate gyrus of long-lived Ames dwarf mice. Endocrinology. 2005;146:1138–1144. doi: 10.1210/en.2004-1115. [DOI] [PubMed] [Google Scholar]
- 287.Parent JM. Injury-induced neurogenesis in the adult mammalian brain. Neuroscientist. 2003;9:261–272. doi: 10.1177/1073858403252680. [DOI] [PubMed] [Google Scholar]
- 288.Ajo R, Cacicedo L, Navarro C, Sanchez-Franco F. Growth hormone action on proliferation and differentiation of cerebral cortical cells from fetal rat. Endocrinology. 2003;144:1086–1097. doi: 10.1210/en.2002-220667. [DOI] [PubMed] [Google Scholar]
- 289.Lyuh E, Kim HJ, Kim M, et al. Dose-specific or dose-dependent effect of growth hormone treatment on the proliferation and differentiation of cultured neuronal cells. Growth Horm IGF Res. 2007;17:315–322. doi: 10.1016/j.ghir.2007.03.002. [DOI] [PubMed] [Google Scholar]
- 290.Blackmore DG, Golmohammadi MG, Large B, Waters MJ, Rietze RL. Exercise increases neural stem cell number in a growth hormone-dependent manner, augmenting the regenerative response in aged mice. Stem Cells. 2009;27:2044–2052. doi: 10.1002/stem.120. [DOI] [PubMed] [Google Scholar]
- 291.Devesa P, Reimunde P, Gallego R, Devesa J, Arce V. Growth hormone (GH) treatment may cooperate with locally-produced GH in increasing the proliferative response of hippocampal progenitors to kainate-induced injury. Brain Inj. 2011;25:503–510. doi: 10.3109/02699052.2011.559611. [DOI] [PubMed] [Google Scholar]
- 292.Devesa P, Agasse F, Xapelli S, et al. Growth hormone pathways signaling for cell proliferation and survival in hippocampal neural precursors from postnatal mice. BMC Neurosci. 2014;15:100. doi: 10.1186/1471-2202-15-100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 293.David Aberg N, Lind J, Isgaard J, Georg KH. Peripheral growth hormone induces cell proliferation in the intact adult rat brain. Growth Horm IGF Res. 2010;20:264–269. doi: 10.1016/j.ghir.2009.12.003. [DOI] [PubMed] [Google Scholar]
- 294.Reimunde P, Rodicio C, López N, Alonso A, Devesa P, Devesa J. Effects of recombinant growth hormone replacement and physical rehabilitation in recovery of gross motor function in children with cerebral palsy. Ther Clin Risk Manag. 2010;30:585–592. doi: 10.2147/TCRM.S14919. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 295.Devesa J, Alonso B, Casteleiro N, et al. Effects of recombinant growth hormone (GH) replacement and psychomotor and cognitive stimulation in the neurodevelopment of GH-deficient (GHD) children with cerebral palsy: a pilot study. Ther Clin Risk Manag. 2011;7:199–206. doi: 10.2147/TCRM.S21403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 296.High WM, Briones-Galang M, Clark JA, et al. Effect of growth hormone replacement therapy on cognition after traumatic brain injury. J Neurotrauma. 2010;27:1565–1575. doi: 10.1089/neu.2009.1253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 297.Reimunde P, Quintana A, Castañón B, et al. Effects of growth hormone (GH) replacement and cognitive rehabilitation in patients with cognitive disorders after traumatic brain injury. Brain Inj. 2011;25:65–73. doi: 10.3109/02699052.2010.536196. [DOI] [PubMed] [Google Scholar]
- 298.Devesa J, Reimunde P, Devesa A, et al. Recovery from neurological sequelae secondary to oncological brain surgery in an adult growth hormone-deficient patient after growth hormone treatment. J Rehabil Med. 2009;41:775–777. doi: 10.2340/16501977-0416. [DOI] [PubMed] [Google Scholar]
- 299.Heredia M, Fuente A, Criado J, Yajeya J, Devesa J, Riolobos AS. Early growth hormone (GH) treatment promotes relevant motor functional improvement after severe frontal cortex lesion in adult rats. Behav Brain Res. 2013;247:48–58. doi: 10.1016/j.bbr.2013.03.012. [DOI] [PubMed] [Google Scholar]
- 300.Devesa J, Díaz-Getino G, Rey P, et al. Brain Recovery after a Plane Crash: Treatment with Growth Hormone (GH) and Neurorehabilitation: A Case Report. Int J Mol Sci. 2015;16:30470–30482. doi: 10.3390/ijms161226244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 301.Devesa J, Lema H, Zas E, Munín B, Taboada P, Devesa P. Learning and Memory Recoveries in a Young Girl Treated with Growth Hormone and Neurorehabilitation. J Clin Med. 2016;5:pii:E14. doi: 10.3390/jcm5020014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 302.Pathipati P, Surus A, Williams CE, Scheepens A. Delayed and chronic treatment with growth hormone after endothelin-induced stroke in the adult rat. Behav Brain Res. 2009;204:93–101. doi: 10.1016/j.bbr.2009.05.023. [DOI] [PubMed] [Google Scholar]
- 303.Song J, Park K, Lee H, Kim M. The effect of recombinant human growth hormone therapy in patients with completed stroke: a pilot trial. Ann Rehabil Med. 2012;36:447–457. doi: 10.5535/arm.2012.36.4.447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 304.Christophidis LJ, Gorba T, Gustavsson M, et al. Growth hormone receptor immunoreactivity is increased in the subventricular zone of juvenile rat brain after focal ischemia: a potential role for growth hormone in injury-induced neurogenesis. Growth Horm IGF Res. 2009;19:497–506. doi: 10.1016/j.ghir.2009.05.001. [DOI] [PubMed] [Google Scholar]
- 305.Li RC, Guo SZ, Raccurt M, Moudilou E, et al. Exogenous growth hormone attenuates cognitive deficits induced by intermittent hipoxia in rats. Neuroscience. 2011;196:237–250. doi: 10.1016/j.neuroscience.2011.08.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 306.Aberg D. Role of the growth hormone/insulin-like growth factor 1 axis in neurogenesis. Endocr Dev. 2010;17:63–76. doi: 10.1159/000262529. [DOI] [PubMed] [Google Scholar]
- 307.Nyberg F, Hallberg M. Growth hormone and cognitive function. Nat Rev Endocrinol. 2013;9:357–365. doi: 10.1038/nrendo.2013.78. [DOI] [PubMed] [Google Scholar]
- 308.Le Grèves M, Zhou Q, Berg M, et al. Growth hormone replacement in hypophysectomized rats affects spatial performance and hippocampal levels of NMDA receptor subunit and PSD-95 gene transcript levels. Exp Brain Res. 2006;173:267–273. doi: 10.1007/s00221-006-0438-2. [DOI] [PubMed] [Google Scholar]
- 309.Enhamre-Brolin E, Carlsson A, Hallberg M, Nyberg F. Growth hormone reverses streptozotocin-induced cognitive impairments in male mice. Behav Brain Res. 2012;238:273–278. doi: 10.1016/j.bbr.2012.10.036. [DOI] [PubMed] [Google Scholar]
- 310.Jørgensen JO, Müller J, Møller J, et al. Adult growth hormone deficiency. Horm Res. 1994;42:235–241. doi: 10.1159/000184199. [DOI] [PubMed] [Google Scholar]
- 311.Rosén T, Wirén L, Wilhelmsen L, Wiklund I, Bengtsson BA. Decreased psychological well-being in adult patients with growth hormone deficiency. Clin Endocrinol (Oxf) 1994;40:111–116. doi: 10.1111/j.1365-2265.1994.tb02452.x. [DOI] [PubMed] [Google Scholar]
- 312.McGauley G, Cuneo R, Salomon F, Sönksen PH. Growth hormone deficiency and quality of life. Horm Res. 1996;45:34–37. doi: 10.1159/000184756. [DOI] [PubMed] [Google Scholar]
- 313.Burman P, Johansson AG, Siegbahn A, Vessby B, Karlsson FA. Growth hormone (GH)-deficient men are more responsive to GH replacement therapy than women. J Clin Endocrinol Metab. 1997;82:550–555. doi: 10.1210/jcem.82.2.3776. [DOI] [PubMed] [Google Scholar]
- 314.Falletti MG, Maruff P, Burman P, Harris A. The effects of growth hormone (GH) deficiency and GH replacement on cognitive performance in adults: a meta-analysis of the current literature. Psychoneuroendocrinology. 2006;31:681–691. doi: 10.1016/j.psyneuen.2006.01.005. [DOI] [PubMed] [Google Scholar]
- 315.Webb SM. Measurements of quality of life in patients with growth hormone deficiency. J Endocrinol Invest. 2008;31(9 Suppl):52–55. [PubMed] [Google Scholar]
- 316.Geisler A, Lass N, Reinsch N, et al. Quality of life in children and adolescents with growth hormone deficiency: association with growth hormone treatment. Horm Res Paediatr. 2012;78:94–99. doi: 10.1159/000341151. [DOI] [PubMed] [Google Scholar]
- 317.Chaplin JE, Kriström B, Jonsson B, et al. Improvements in behaviour and self-esteem following growth hormone treatment in short prepubertal children. Horm Res Paediatr. 2011;75:291–303. doi: 10.1159/000322937. [DOI] [PubMed] [Google Scholar]
- 318.Deijen JB, de Boer H, Blok GJ, van der Veen EA. Cognitive impairments and mood disturbances in growth hormone deficient men. Psychoneuroendocrinology. 1996;21:313–322. doi: 10.1016/0306-4530(95)00050-x. [DOI] [PubMed] [Google Scholar]
- 319.Arwert LI, Veltman DJ, Deijen JB, van Dam PS, Delemarre-van deWaal HA, Drent ML. Growth hormone deficiency and memory functioning in adults visualized by functional magnetic resonance imaging. Neuroendocrinology. 2005;82:32–40. doi: 10.1159/000090123. [DOI] [PubMed] [Google Scholar]
- 320.Arwert LI, Veltman DJ, Deijen JB, van Dam PS, Drent ML. Effects of growth hormone substitution therapy on cognitive functioning in growth hormone deficient patients: a functional MRI study. Neuroendocrinology. 2006;83:12–19. doi: 10.1159/000093337. [DOI] [PubMed] [Google Scholar]
- 321.Li E, Kim DH, Cai M, Lee S. Hippocampus-dependent spatial learning and memory are impaired in growth hormone-deficient spontaneous dwarf rats. Endocr J. 2011;58:257–267. doi: 10.1507/endocrj.k11e-006. [DOI] [PubMed] [Google Scholar]
- 322.Park SW, Shin S, Kim CH, et al. Differential effects of insufflated, subcutaneous, and intravenous growth hormone on bone growth, cognitive function, and NMDA receptor subunit expression. Endocrinology. 2010;151:4418–4427. doi: 10.1210/en.2010-0152. [DOI] [PubMed] [Google Scholar]
- 323.Le Grevès M, Steensland P, Le Grevès P, Nyberg F. Growth hormone induces age-dependent alteration in the expression of hippocampal growth hormone receptor and N-methyl-D-aspartate receptor subunits gene transcripts in male rats. Proc Natl Acad Sci U S A. 2002;99:7119–7123. doi: 10.1073/pnas.092135399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 324.Mahmoud GS, Grover LM. Growth hormone enhances excitatory synaptic transmission in area CA1 of rat hippocampus. J Neurophysiol. 2006;95:2962–2974. doi: 10.1152/jn.00947.2005. [DOI] [PubMed] [Google Scholar]
- 325.Lichtenwalner RJ, Forbes ME, Bennett SA, Lynch CD, Sonntag WE, Riddle DR. Intracerebroventricular infusion of insulin-like growth factor-I ameliorates the age-related decline in hippocampal neurogenesis. Neuroscience. 2001;107:603–613. doi: 10.1016/s0306-4522(01)00378-5. [DOI] [PubMed] [Google Scholar]
- 326.Lichtenwalner RJ, Forbes ME, Sonntag WE, Riddle DR. Adult-onset deficiency in growth hormone and insulin-like growth factor-I decreases survival of dentate granule neurons: insights into the regulation of adult hippocampal neurogenesis. J Neurosci Res. 2006;83:199–210. doi: 10.1002/jnr.20719. [DOI] [PubMed] [Google Scholar]
- 327.Aberg MA, Aberg ND, Hedbacker H, Oscarsson J, Eriksson PS. Peripheral infusion of IGF-I selectively induces neurogenesis in the adult rat hippocampus. J Neurosci. 2000;20:2896–2903. doi: 10.1523/JNEUROSCI.20-08-02896.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 328.Darnaudery M, Perez-Martin M, Belizaire G, Maccari S, Garcia-Segura LM. Insulin-like growth factor 1 reduces age-related disorders induced by prenatal stress in female rats. Neurobiol Aging. 2006;27:119–127. doi: 10.1016/j.neurobiolaging.2005.01.008. [DOI] [PubMed] [Google Scholar]
- 329.Marrone DF, Ramirez-Amaya V, Barnes CA. Neurons generated in senescence maintain capacity for functional integration. Hippocampus. 2012;22:1134–1142. doi: 10.1002/hipo.20959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 330.Azcoitia I, Pérez Martín M, Salazar V, et al. Growth hormone prevents neuronal loss in the aged rat hippocampus. Neurobiol Aging. 2005;5:697–703. doi: 10.1016/j.neurobiolaging.2004.06.007. [DOI] [PubMed] [Google Scholar]
- 331.Aberg ND, Johansson UE, Aberg MA, et al. Peripheral infusion of insulin-like growth factor-I increases the number of newborn oligodendrocytes in the cerebral cortex of adult hypophysectomized rats. Endocrinology. 2007;148:3765–3772. doi: 10.1210/en.2006-1556. [DOI] [PubMed] [Google Scholar]
- 332.Pang Y, Zheng B, Fan LW, Rhodes PG, Cai Z. IGF-1 protects oligodendrocyte progenitors against TNF alpha-induced damage by activation of PI3K/Akt and interruption of the mitochondrial apoptotic pathway. Glia. 2007;55:1099–1107. doi: 10.1002/glia.20530. [DOI] [PubMed] [Google Scholar]
- 333.Gonzalez-Perez O, Alvarez-Buylla A. Oligodendrogenesis in the subventricular zone and the role of epidermal growth factor. Brain Res Rev. 2011;67:147–156. doi: 10.1016/j.brainresrev.2011.01.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 334.Pan SN, Ma HM, Su Z, Zhang CX, Zhu SY, Du ML. Epidermal growth factor receptor signaling mediates growth hormone-induced growth of chondrocytes from sex hormone-inhibited adolescent rats. Clin Exp Pharmacol Physiol. 2011;38:534–542. doi: 10.1111/j.1440-1681.2011.05547.x. [DOI] [PubMed] [Google Scholar]
- 335.Sim FJ, Zhao C, Penderis J, Franklin RJ. The age-related decrease in CNS remyelination efficiency is attributable to an impairment of both oligodendrocyte progenitor recruitment and differentiation. J Neurosci. 2002;22:2451–2459. doi: 10.1523/JNEUROSCI.22-07-02451.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 336.Stern WC, Miller M, Jalowiec JE, Forbes WB, Morgane PJ. Effects of growth hormone on brain biogenic amine levels. Pharmacol Biochem Behav. 1975;3:1115–1118. doi: 10.1016/0091-3057(75)90025-8. [DOI] [PubMed] [Google Scholar]
- 337.Söderpalm B, Ericson M, Bohlooly M, Engel JA, Tornell J. Bovine growth hormone transgenic mice display alterations in locomotor activity and brain monoamine neurochemistry. Endocrinology. 1999;140:5619–5625. doi: 10.1210/endo.140.12.7181. [DOI] [PubMed] [Google Scholar]
- 338.Andersson K, Fuxe K, Eneroth P, Isaksson O, Nyberg F, Roos P. Rat growth hormone and hypothalamic catecholamine nerve terminal systems. Evidence for rapid and discrete reductions in dopamine and noradrenaline levels and turnover in the median eminence of the hypophysectomized male rat. Eur J Pharmacol. 1983;95:271–275. doi: 10.1016/0014-2999(83)90645-3. [DOI] [PubMed] [Google Scholar]
- 339.Le Grevès M, Le Grevès P, Nyberg F. Age-related effects of IGF-1 on the NMDA-, GH- and IGF-1-receptor mRNA transcripts in the rat hippocampus. Brain Res Bull. 2005;65:369–374. doi: 10.1016/j.brainresbull.2005.01.012. [DOI] [PubMed] [Google Scholar]
- 340.Arce VM, Cella SG, Locatelli V, Müller EE. Studies of growth hormone secretion in calorically restricted dogs: effect of cholinergic agonists and antagonists, glucose and thyrotropin-releasing hormone. Neuroendocrinology. 1991;53:467–472. doi: 10.1159/000125759. [DOI] [PubMed] [Google Scholar]
- 341.Ghigo E, Goffi S, Arvat E, et al. A neuroendocrinological approach to evidence an impairment of central cholinergic function in aging. J Endocrinol Invest. 1992;15:665–670. doi: 10.1007/BF03345812. [DOI] [PubMed] [Google Scholar]
- 342.Araujo DM, Lapchak PA, Collier B, Chabot JG, Quirion R. Insulin-like growth factor-1 (somatomedin-C) receptors in the rat brain: distribution and interaction with the hippocampal cholinergic system. Brain Res. 1989;484:130–138. doi: 10.1016/0006-8993(89)90355-7. [DOI] [PubMed] [Google Scholar]
- 343.Devesa J, Devesa P, Reimunde P, Arce V. Growth Hormone and Kynesitherapy for Brain Injury Recovery. In: Agrawal Amit., editor. Brain Injury—Pathogenesis, Monitoring, Recovery and Management. 2012. pp. 417–454. [Google Scholar]
- 344.Devesa P, Gelabert M, González-Mosquera T, et al. Growth hormone treatment enhances the functional recovery of sciatic nerves after transection and repair. Muscle Nerve. 2012;45:385–392. doi: 10.1002/mus.22303. [DOI] [PubMed] [Google Scholar]
- 345.Tuffaha SH, Budihardjo JD, Sarhane KA, et al. Growth Hormone Therapy Accelerates Axonal Regeneration, Promotes Motor Reinnervation, and Reduces Muscle Atrophy following Peripheral Nerve Injury. Plast Reconstr Surg. 2016;137:1771–1780. doi: 10.1097/PRS.0000000000002188. [DOI] [PubMed] [Google Scholar]
- 346.Ray BN, Kweon HK, Argetsinger LS, Fingar DC, Andrews PC, Carter-Su C. Research resource: identification of novel growth hormone-regulated phosphorylation sites by quantitative phosphoproteomics. Mol Endocrinol. 2012;26:1056–1073. doi: 10.1210/me.2011-1258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 347.Carter-Su C, Schwartz J, Argetsinger LS. Growth hormone signaling pathways. Growth Horm IGF Res. 2016;28:11–15. doi: 10.1016/j.ghir.2015.09.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 348.Scheinman EJ, Damouni R, Caspi A, Shen-Orr Z, Tiosano D, LeRoith D. The beneficial effect of growth hormone treatment on islet mass in streptozotocin-treated mice. Diabetes Metab Res Rev. 2015;31:492–449. doi: 10.1002/dmrr.2631. [DOI] [PubMed] [Google Scholar]