

Abstract
Background:
Chlorophytum borivilianum is an industrially valued medicinal crop. Propagation through seeds is not feasible because of low germination percentage and long dormancy period. Therefore, callus culture and plant regeneration can be an alternative to improve this crop production. Also, callus can serve as an alternative source of bioactive compounds.
Objective:
To evaluate the effect of different phytohormones on callus induction, subculture cycle, and regeneration studies of callus in C. borivilianum.
Materials and Methods:
Young shoot buds of C. borivilianum were inoculated on Murashige and Skoog medium fortified with 3% sucrose and different concentrations (0, 1, 5, 10, and 15 mg/L) of either naphthalene acetic acid or 2,4-dichlorophenoxyacetic acid or indole-3-acetic acid and callus induction was evaluated up to four subcultures cycles. Shoot regeneration from callus was studied on Murashige and Skoog media fortified with 6-benzylaminopurine andkinetin or thidiazuron at varied levels (0, 0.5, 1, 2, and 3 mg/L). Microshoots were rooted on Murashige and Skoog media supplemented with 1.0 mg/L indole-3-butyric acid and plantlets were acclimatized before transferred to the natural conditions.
Results:
Callus induction was better evidenced on Murashige and Skoog media containing 5 mg/L 2,4-dichlorophenoxyacetic acid up to fourth subculture. Callus differentiated into shoots on Murashige and Skoog media fortified with 6-benzylaminopurine or kinetin, whereas thidiazuron completely failed to regenerate shoots. Furthermore, microshoots rooted on 1.0 mg/L indole-3-butyric acid containing Murashige and Skoog media. The rooted plantlets were successfully acclimatized and established in soil with 88.3% survivability.
Conclusion:
The type of auxins played an important role in inducing callus tissue from shoot bud explants of Safed musli. In future, this in vitro protocol could benefit in crop improvement programs and serve as a new source of bioactive compounds from Safed musli callus tissue for various therapeutic applications.
SUMMARY
Explants de-differentiated to form callus on Murashige and Skoog media containing 5 mg/L 2,4-D up to fourth subculture.
Callus re-differentiated into shoots on Murashige and Skoog media fortified with 0.5 mg/L BAP.
In vitro rooting of shoots was achieved on 1.0 mg/L IBA containing Murashige and Skoog media.
The rooted plantlets were successfully acclimatized and established in soil with 88.3% survivability.
Abbreviations used: MS: Murashige and Skoog, NAA: naphthalene acetic acid, 2,4-D: 2,4-dichlorophenoxyacetic acid, IAA: indole-3-acetic acid, BAP: 6-benzylaminopurine, Kn: Kinetin, TDZ: thidiazuron, IBA: indole-3-butyric acid, RCBD: Randomized Complete Block Design, DMRT: Duncan's Multiple Range Test
Keywords: Callus, regeneration, phytohormones, saponins, Safed musli
INTRODUCTION
Safed musli (Chlorophytum borivilianum), which originates from India, is an industrially valued medicinal plant belonging to the family Liliaceae. Among more than 300 species identified under this genus, Safed musli contains the highest amount of saponins, which is known for its therapeutic effect in the Ayurvedic medicine. Its tuberous root extract is used as an alternative to viagra, cure diabetes, arthritis, chronic leucorrhoea, and delay menopause.[1] The tuberous roots of this crop contain numerous active ingredients, such as saponins, alkaloids, flavonoids and phenolic acids, steroids, triterpenoids, tannins, and vitamins.[2] There is certainly a huge demand for Safed musli in India. In addition, because of increasing awareness and appreciation on the health benefits of herbal products, the demand for Safed musli has bolstered across the world, forcing establishment of new farms. Although Safed musli is able to produce seeds, the viability is low with only 14–16% germination and has long dormancy period.[3] In addition, tuber formation only occurs in the second year when the plant is grown using seeds. However, tubers have been used to propagate and establish the crop. Although the use of tubers is more amenable, tubers have dormancy and often have asynchronous germination causing difficulties in management of the agronomic aspects. In addition, the valuable tubers that can be utilized for secondary metabolite extraction has to be sacrificed for use as planting materials. Because of the abovementioned constraints, micropropagation can be an alternative to supply planting materials for production of Safed musli in relatively short time and produce disease free, genetically uniform plants with higher yield.
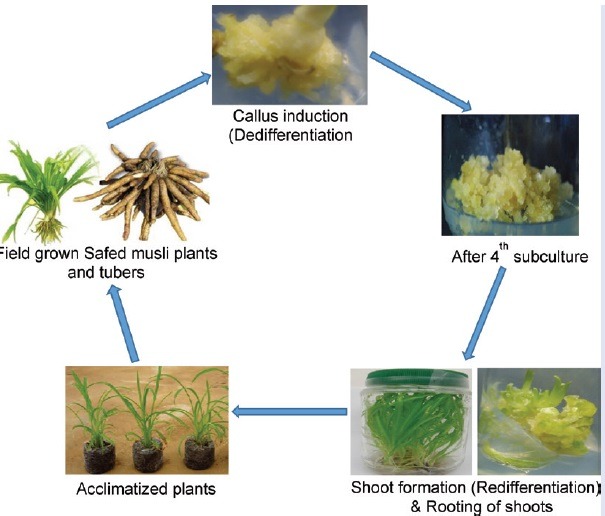
Under in vitro conditions, plants can be regenerated via direct organogenesis or indirect organogenesis through the formation of unorganized callus tissues induced by the application of exogenous growth regulators.[4] It is a prerequisite to establish a suitable callus induction and shoot regeneration method for every plant species in order to make use of tissue culture practices in plant improvement programs including transformation technology.[5] The plant growth regulators play a significant role in inducing callus and followed by their subsequent shoot regeneration into plantlet. The response to plant growth regulators depend on the type of explant tissues and plant species. According to Narayanaswamy,[6] an auxin such as 2,4-dichlorophenoxyacetic acid (2,4-D) or naphthalene acetic acid (NAA) is very ideal for callus induction. Among the auxin types, 2,4-D is the most frequently used hormone to initiate callus and somatic embryogenesis in various plants, whereas indole-3-acetic acid (IAA), indole-3-butyric acid (IBA), NAA, picloram, and dicamba could also be used as stimulants but in a lesser extent.[7,8,9] The regeneration of shoot is the next step after achieving callus in indirect organogenesis method. Plant regeneration from callus is possible by de novo organogenesis or somatic embryogenesis.[10] Among cytokinins, 6-benzylaminopurine (BAP) interacts actively when added to media along with 2,4-D or IAA for callus initiation and its maintenance.[6] According to Ignacimuthu,[11] cytokinin at low concentrations is known to promote shoot differentiation from callus. The proliferation of callus depends on the rate of growth and media composition. Accordingly, it is necessary to subculture periodically on a fresh medium for maintaining its growth. Moreover, repeated subculture cycles of callus could reduce its regenerative capacity. Therefore, optimization of subculture period is essential in order to obtain higher callus biomass with regenerative potency. Tissue culture studies including somatic embryogenesis, clonal propagation, liquid culture, and in vitro tuberization have been reported in Safed musli using different explants.[12,13,14,15,16] However, further research efforts are required to establish an efficient and reliable in vitro culture system in Safed musli crop improvements through other biotechnological means. Moreover, so far limited efforts are made to observe the effect of various plant growth regulators on in vitro callus induction and maintenance. Callus can serve as alternatives to medicinally valued compounds from plants and can serve as supplement to conventional practice of field propagation for large-scale production of phytocompounds. As plant tissue culture approach is independent from environmental conditions, there is a possibility of producing desirable bioactive compounds continuously.[17] Therefore, the present investigation was undertaken with an objective to develop an efficient callus induction and shoot organogenesis protocol for Safed musli by employing various plant growth regulators. Also, the effect of subculture frequencies on Safed musli callus culture is described.
MATERIALS AND METHODS
Plant material
Tubers of Safed musli were harvested at maturity and stored in the dark at 28°C. The tubers were taken out after a month of storage, and tubers which had sprouted during the storage with a shoot length of about 1–2 cm were selected for this study. The selected tubers were thoroughly washed by using tap water to facilitate the removal of adhering particles and later immersed in 5 g/L of bavistin (fungicide) for 2 h. Explants were then immersed in tap water and washed three times using sterilized double-distilled water and placed for 20 min in 0.1% sodium hypochlorite (NaOCl) containing two to three drops of Tween-20. Under the laminar air flow, explants were treated with 0.1% HgCl2 solution for 15 min to complete the surface sterilization procedure. Finally, explants were washed three times using sterilized double distilled water and the young shoot buds were incised vertically (size) and the these explants were inoculated on the culture medium for callus induction.
Callus induction and subculture
Full strength Murashige and Skoog basal medium[18] containing 3% sucrose was used for the callus induction study. Three types of auxins (NAA, 2,4-D, and IAA) were added in the media at different concentration each (0, 1, 5, 10, and 15 mg/L). Using either 1M HCl or 1M NaOH, pH of the medium was adjusted to 5.8, and finally 3.9 g/L agar (Difco, USA) was added to the medium. After inoculation, all culture flasks were kept at 25 ± 2°C under fluorescent lighting for 16 h (light intensity at 45 μmoL/m2 /s). Once callus formation was noticed, they were isolated and transferred onto the same induction media for further growth. The callus was subcultured at every 28-day interval up to four subculture cycles and subsequently transferred to shoot regeneration media. The data on various parameters such as callus morphological features, percentage of callus formation, number of days required to induce callus, and mean fresh weight (g) of callus were recorded.
Shoot regeneration
Shoot regeneration was studied by using full strength Murashige and Skoog basal media fortified with various types of cytokinins such as BAP, kinetin (Kn), and thidiazuron (TDZ) at varied levels (0, 0.5, 1, 2, and 3 mg/L). Approximately 1 g of callus was placed in each culture tube and data were collected after 28 days of culture. The parameters included in this experiment were percentage of shoot regeneration and number of shoots formed per culture tube.
In vitro rooting and acclimatization
In vitro rooting was achieved by following the protocol of Kemat et al.[19] Briefly, about 3–4 cm long in vitro regenerated shoots derived from calli were inoculated on to full strength Murashige and Skoog medium supplemented with 1.0 mg/L IBA. Later, well-rooted plantlets were carefully washed using sterile distilled water to remove agar and placed in benlate solution (3%) (fungicide) for 5 min. Later, the plantlets were transferred to small polycups containing vermiculite and organic matter of plant and animal origin at the ratio of 1?:?1 (w/w). The transplants were kept under greenhouse condition for about 3 weeks. Plantlets were watered twice a day and to ensure high humidity pots were covered with plastic polythene bags. After acclimatization, the plants were then moved to the larger polythene bags containing normal garden soil and allowed to grow under natural condition and data on the percentage survival were recorded after 5 weeks of transfer.
Statistical analysis
All studies were organized in a randomized complete block design with a single factor. All experiments were replicated three times and each replication consisted of 10 samples. Data were analyzed using statistical software (SAS program version 9.1). Differences between the treatment means were analyzed by means of Duncan's multiple range test at level α = 5%.
RESULTS
Effect of auxins on the induction of callus
In this study, different types of auxins (NAA, 2,4-D, and IAA) with various concentrations were used for inducing callus in Safed musli. The results showed that there was a considerable influence of 2,4-D on callus induction, whereas both NAA and IAA supplemented media showed no response on callus formation [Table 1]. Callus initiation was triggered in all treatments of 2,4-D supplemented media. The explants started to form callus on the eighth week (56 days) of culture on Murashige and Skoog media containing 1 to 5 mg/L 2,4-D. However, the highest percentage of callus induction (66.67%) and mean callus fresh weight (10.65 ± 0.78 g) was observed on 5 mg/L 2,4-D supplemented media after 56 days of inoculation. This was followed by treatment with 1 mg/L 2,4-D, where 40% of callus induction with mean callus fresh weight of 4.49 ± 0.22 g was noticed [Figure 1a and b]. Higher concentrations of 2,4-D were not effective in inducing callus [Figure 1c and d]. All calli appeared to be friable and yellowish in color during the initial stage of callus formation (56–63 days). There was no evidence of callus on Murashige and Skoog media devoid of growth regulators (control), although Murashige and Skoog media containing different concentrations of NAA and IAA produced only roots and did not show any signs of callusing [Figure 1e].
Table 1.
The effect of different auxin types (NAA, 2,4-D, and IAA) on callus induction from young shoot bud explants of Safed musli after 8 weeks of culture
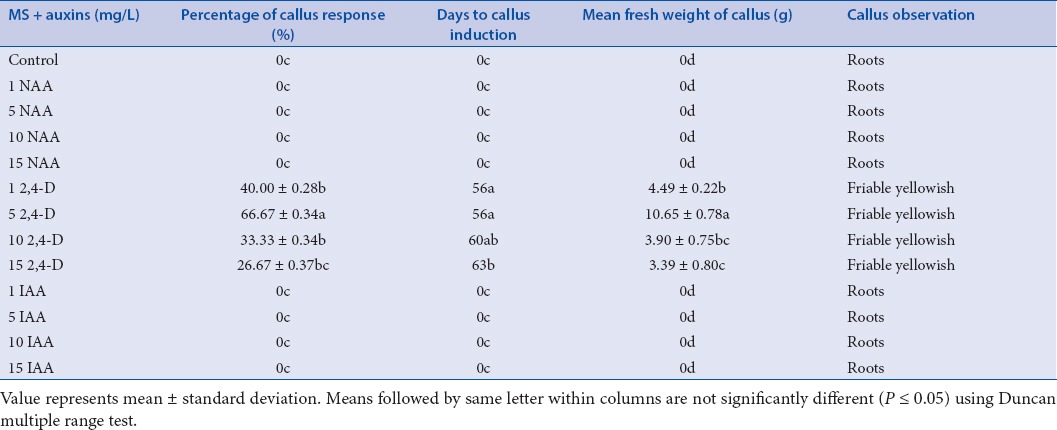
Figure 1.

Callus induction and shoot organogenesis in Safed musli: Explants forming callus on MS + 1.0 mg/L (A); 5.0 mg/L 2,4-D (B); 10 mg/L 2,4-D (C); 15 mg/L 2,4-D (D) after 8 weeks; Explants showing roots on MS + 0.5 mg/L NAA after 8 weeks (E) (Scale bar = 0.5 cm); Callus after fourth sub culture cycle on MS + 05mg/L 2,4-D (F) (Scale bar = 1.0 cm); Shoots regenerated on MS + 0.5 mg/L BAP (G) after 4 weeks; Green callus on MS + 1.0 mg/L TDZ (H) after 6 weeks (Scale bar = 0.5 cm); Multiplication of shoots on MS + 0.5mg/L BA (I) after 15 days of subculture (Scale bar = 1.0 cm).
Table 2.
The effect of subcultures on mean weight of Safed musli callus from first subculture until fourth subculture
Subculture of callus
As callusing was only found in media supplemented with 2,4-D, subculture experiment was carried out only for the different concentration of 2,4-D. The callus growth was remarkable and occurred at the surface not in contact with agar media. There was a significant difference between all the treatments from first subculture until fourth subculture. During the first subculture on Murashige and Skoog medium containing 5 mg/L 2,4-D, there was a higher growth of callus with mean fresh weight of 10.65 ± 0.78 g within 4 weeks of culture. However, increased levels of 2,4-D showed no significant effect in enhancing callus growth. As the subculture continued from first to fourth, there was a noticeable increase in mean weight of callus in all the treatments. 2,4-D at 5 mg/L concentration exhibited the highest callus formation at all subculture MS, Murashige and Skoog. periods and the highest callus fresh weight (26.13 ± 3.18 g) was observed at the fourth subculture. Although the number of subcultures and concentration of 2,4-D affected the growth and development of callus, morphologically all calluses were friable yellowish [Figure 1f].
Indirect shoot regeneration from callus
In this study, callus from the fourth subculture were used to initiate shoot regeneration. The callus cultures displayed tendency to differentiate into shoot on medium fortified with either BAP or Kinetin. However, morphogenesis was found to vary considerably at varied concentrations of cytokinins [Table 3]. The highest percentage of shoot induction (66.7 ± 0.24) and mean number of shoots per tube (9.6 ± 1.51) were established when 0.5 mg/L BAP was supplemented to Murashige and Skoog media [Figure 1g]. The next best result was noticed on Murashige and Skoog media supplemented with 0.5 mg/L Kn, where 53.33 ± 0.18% of shoot induction and 5.6 ± 0.54 mean number of shoots per tube was observed. However, increased concentrations of both BAP and Kn decreased the shoot formation frequency and shoot numbers. Higher concentrations of both BAP and Kn were only able to induce 20–27% of shoots. The regeneration process was found to vary significantly among the treatments. The green spots were noticed as early as 7 days after inoculation in treatments containing 0.5 mg/L of BAP. The green spot later formed shoot primordia, and developed into a young shoot bud. The process of regeneration starting from greenish callus until shoot development took about 7–28 days, depending on the treatments. Although, all concentrations of TDZ failed to regenerate shoots, morphologically callus changed into a friable and were green in colour [Figure 1h]. The present study clearly indicates that Safed musli shoot regeneration from callus can be achieved by using low concentration of either BAP or Kn (0.5 mg/L each) in the media.
Table 3.
The effect of cytokinin types on callus derived shoot regeneration in Safed musli after 4 weeks of culture
In vitro rooting and acclimatization
In this study, we have used the in vitro rooting protocol established earlier by Kemat et al.[19] in Safed musli. It was observed that Murashige and Skoog media supplemented with IBA was able to induce better rooting of callus derived microshoots. Murashige and Skoog medium supplemented with 1.0 mg/L IBA was found to be effective in inducing in vitro roots (100%) with 50.33 ± 0.98 mean number of roots having mean length of 5.96 ± 0.56 cm. The acclimatized plantlets showed a higher percentage of survival (88.34 ± 1.97%) under normal condition. All plants were observed to be normal and grew well with no signs of morphological variations.
DISCUSSION
Various plant growth regulators have been reported to be successful in enhancing the initiation of callus in the family, Liliaceae.[10,21] The morphological characteristics of callus initiated from young shoot buds of Safed musli was found to be similar to the results obtained by Suman et al.[22] in Safed musli, using pedicel and inflorescence parts the explant. Also, they have proposed that optimized callus growth can be accomplished on Murashige and Skoog medium containing 20 μmol/L 2,4-D, 4 μmol/L NAA, and 0.067 μmol/L BAP, which is contradictory to our results where use of NAA and IAA did not show any signs of callus growth; however, induced rooting. According to Narayanaswamy,[6] callus producing roots will never differentiate into shoots because shoot morphogenesis gets terminated with the formation of roots within a callus. Hence, the prospect of plantlet production from callus with roots is not feasible. It has also been stated that 2,4-D is more active in triggering meristemic activity leading to cell proliferation, compared with NAA and IAA.[18] The ability of 2,4-D to enhance callus regeneration was also found in Aloe vera.[9] Similarly, many researchers have used 2,4-D in the media to encourage callus formation in different plant species.[6,22,23] The subculture processes in callus is important to ensure good growth and development. The failure to do subculture in callus will cause the tissue to gradually die. Narayanaswamy[6] recommended that callus has to be subcultured at every 4–6 weeks. In this study, subculture carried out at every fourth week up to four subculture cycles played a significant role in enhancing the callus growth with no morphological variations. Suman et al.[22] have reported that after 6–8 weeks of inoculation and subculture, the callus becomes yellowish, compact, hard, and nodular when pedicel and inflorescence parts of Safed musli were used as explants. However, callus obtained in our study using shoot tips remained to be yellowish and friable. Callus tissue has the ability to induce adventitious shoots and it is considered as a good source for introducing genetic variability in crops.[23] Shoot differentiation was observed from the calli obtained after the fourth subculture cycle when inoculated on Murashige and Skoog media supplemented with either BAP or Kn. However, all concentrations of TDZ failed to induce shoots from callus but morphologically calli color turned from friable yellowish to nodular green color. The levels of plant growth hormones play an important role in inducing morphogenic changes during in vitro regeneration.[24] Similarly, shoot regeneration frequency in our study was found to be higher only when lower concentrations of BAP or Kn were used in the media. As the price of BAP is cheaper than Kn, it is more convenient to use BAP instead of Kn.
Correspondingly, it is reported that higher concentrations of either BAP or Kn (>0.5 mg/L) leads to the formation of hyperhydric shoots with morphological abnormalities in Leptadenia reticulata and Pogostemon cablin.[24,25,26] The necessity of cytokinins for shoot initiation was also documented in the fish tail fern[27] and Sarcostemma acidum.[28] Hence, it can be mentioned that in many plant species including Safed musli, cytokinins change the mode of development from callus to differentiate into shoots. Murashige and Skoog media fortified with 1.0 mg/L IBA effectively induced better rooting frequency, which is in corroboration with the previous study reports in Safed musli.[19,29,30] The survival percentage of the acclimatized plantlets were found to be higher under normal condition, which was concurrent with the previous reports in the same plant, Safed musli.[29,30] Also there were no morphological variations observed after transfer and all plants showed normal development. Hence, this protocol can be very useful in regenerating plantlets through callus phase for the production of planting materials or for various biotechnological applications.
CONCLUSION
The type of auxins played an important role in inducing callus tissue from shoot bud explants of Safed musli. Both NAA and IAA were inefficient to induce callus, whereas 2,4-D at 5 mg/L promoted high frequency of callus induction. Subculture cycle showed enhanced callus multiplication rate. The maximum shoot regeneration frequency from callus was observed on Murashige and Skoog medium supplemented with 0.5 mg/L BAP, whereas the use of TDZ failed to exhibit shoot differentiation. Rooting was well established on Murashige and Skoog medium containing 1.0 mg/L IBA and the plantlets showed higher percentage of survival (88.34 ± 1.97%) under field condition. Thus, the present protocol is highly useful for transformation or genetic engineering studies. In addition, this study facilitates the possible extraction of bioactive compounds from Safed musli callus tissue for various therapeutic applications without planting in the field.
Financial support and sponsorship
Nil
Conflicts of interest
There are no conflicts of interest.
Acknowledgement
The authors thank Yayasan Felda for financing this work through a research grant and Universiti Putra Malaysia for all the facilities to successfully carry out this work.
REFERENCES
- 1.Kaushik N. Saponins of Chlorophytum species. Phytochem Rev. 2005;4:4–6. [Google Scholar]
- 2.Khanam Z, Singh O, Singh R, Bhat IUH. Safed musli (Chlorophytum borivilianum): a review of its botany, ethnopharmacology and phytochemistry. J Ethnopharmacol. 2013;150:4–41. doi: 10.1016/j.jep.2013.08.064. [DOI] [PubMed] [Google Scholar]
- 3.Kaur R, Arora S, Thukral AK. Enhancing seed germination of Chlorophytum borivilianum Sant. Et Fernand. with PGRs, steroidal hormones and zinc. Res J Seed Sci. 2009;2:2 32–9. [Google Scholar]
- 4.Gaspar T, Kevers C, Penel C, Greppin H, Reid DM, Thorpe TA. Plant hormones and plant growth regulators in plant tissue culture. In Vitro Cell Dev Biol Plant. 1990;32:4272–89. [Google Scholar]
- 5.Wang XD, Nolan KE, Irwanto RR, Sheahan MB, Rose R. Ontogeny of embryogenic callus in Medica gotruncatula: the fate of the pluripotent and totipotent stem cells. Ann Bot. 2011;107:4599–609. doi: 10.1093/aob/mcq269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Narayanaswamy S. Plant cell and tissue culture 1994 9th reprint India Tata McGraw-Hill Education [Google Scholar]
- 7.Ipekci Z, Gozukirmizi N. Indirect somatic embryogenesis and plant regeneration from leaf and internode explants of Paulownia elongata. Plant Cell Tissue Organ Cult. 2004;79:341–45. [Google Scholar]
- 8.Rai MK, Akhtar N, Jaiswal VS. Somatic embryogenesis and plant regeneration in Psidium guajava L. cv. Banarasi local. Sci Horticult. 2007;113:129–33. [Google Scholar]
- 9.Rathore MS, Chikara J, Shekhawat NS. Plantlet regeneration from callus cultures of selected genotype of Aloe vera L. -an ancient plant for modern herbal industries. Appl Biochem Biotechnol. 2011;163:7860–68. doi: 10.1007/s12010-010-9090-1. [DOI] [PubMed] [Google Scholar]
- 10.Raghavan V. Embryogenesis in angiosperms 1986 Cambridge, UK Cambridge University Press [Google Scholar]
- 11.Ignacimuthu I. Plant biotechnology. New Delhi: India Oxford and IBH Pub. Co; 1997. [Google Scholar]
- 12.Dave A, Joshi N, Purohit SD. In vitro propagation of Chlorophytum borivilianum using encapsulated shoot buds. Eur J Hort Sci. 2014;69:4–42. [Google Scholar]
- 13.Rizvi MZ, Kukreja AK, Khanuja SPS. In vitro culture of Chlorophytum borivilianum Sant. et Fernand in liquid culture medium as a cost effective measure. Curr Sci. 2007;92:87–90. [Google Scholar]
- 14.Haque R, Saha S, Bera T. Micropropagation of an important medicinal plant Chlorophytum borivilianum. Int J Ph Sci. 2009;1:4–63. [Google Scholar]
- 15.Kumar A, Aggarwal D, Gupta P, Reddy MS. Factors affecting in vitro propagation and field establishment of Chlorophytum borivilianum. Biol Plantarum. 2010;54:4–6. [Google Scholar]
- 16.Ashraf MF, Aziz MA, Kadir MA, Stanslas J, Farokhian E. In vitro tuberization of Chlorophytum borivilianum Sant & Fern (Safed musli) as influenced by sucrose, CCC and culture systems. Plant Cell Physiol. 2013;54:4–64. doi: 10.1093/pcp/pct083. [DOI] [PubMed] [Google Scholar]
- 17.López-Laredo AR, Ramírez-Flores FD, Sepúlveda-Jiménez G, Trejo-Tapia G. Comparison of metabolite levels in callus of Tecoma stans (L.) Juss. ex Kunth. cultured in photoperiod and darkness. In Vitro Cell Dev Biol Plant. 2009;45:4–58. [Google Scholar]
- 18.Murashige T, Skoog F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant. 1962;15:4–97. [Google Scholar]
- 19.Kemat N, Kadir M. A, Abdullah NAP, Ashraf F, Rapid multiplication of Safed musli (Chlorophytum borivilianum) through shoot proliferation. Afr J Biotechnol. 2010;9:294595–600. [Google Scholar]
- 20.Yun SY, Lee MS, Lim SC, Shin JD. Micropropagation of Heloniopsis orientalis (Thunb.) C. Tanaka in vitro. Kor J Plant Tissue Culture. 2000;27:197–202. [Google Scholar]
- 21.Wawrosch C, Malla PR, Kopp B. Clonal propagation of Lilium nepalense D. Don, a threatened medicinal plant of Nepal. Plant Cell Rep. 2001;20:285–88. [Google Scholar]
- 22.Suman K, Anuradha P, Shaifali G, Pavan VVS, Shashwat S. Isolation and characterization of secondary metabolites “Saponins” from explants of Indian medicinal plant Chlorophytum borivilianum. Int J Pharm Front. 2011;1:214–22. [Google Scholar]
- 23.Sen MK, Nasrin S, Rahman S, Jamal AHM. In vitro callus induction and plantlet regeneration of Achyranthes aspera L., a high value medicinal plant. A Pac J Trop Biomed. 2014;4:140–6. doi: 10.1016/S2221-1691(14)60206-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Sudipta KM, Kumara Swamy M, Balasubramanya S, Anuradha M. Cost effective approach for in vitro propagation of (Leptadenia reticulata Wight & Arn.)---a threatened plant of medicinal importance. J Phytol. 2011;3:272–9. [Google Scholar]
- 25.Kumaraswamy M, Anuradha M. Micropropagation of Pogostemon cablin Benth. through direct regeneration for production of true to type plants. Plant Tissue Cult Biotechnol. 2010;20:181–9. [Google Scholar]
- 26.Swamy MK, Mohanty SK, Anuradha M. The effect of plant growth regulators and natural supplements on in vitro propagation of Pogostemon cablin Benth. J Crop Sci Biotechnol. 2014;17:4–8. [Google Scholar]
- 27.Beck MJ, Caponetti JD. The effects of kinetin and naphthalene acetic acid on in vitro shoot multiplication and rooting in the fish tail fern. Am J Bot. 1983;70:4–7. doi: 10.1002/j.1537-2197.1983.tb12425.x. [DOI] [PubMed] [Google Scholar]
- 28.Rathore MS, Shekhawat NS. In vitro regeneration in Sarcostemma acidum (Roxb.)-an important medicinal plant of semi-arid ecosystem of Rajasthan, India. Physiol Mol Biol Plants. 2013;19:2269–75. doi: 10.1007/s12298-012-0158-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Meenakshi P, Neeraj S, Sushila K, Milind P. Plant regeneration studies in Safed musli (Chlorophytum. sp.) Int J Res Ayuveda Pharm. 2014;5:195–98. [Google Scholar]
- 30.Sharma U, Mohan JSS. In vitro clonal propagation of Chlorophytum borivilianum Sant. et Fernand., a rare medicinal herb from immature floral buds along with inflorescence axis. Indian J Exp Biol. 2006;44:177–82. [PubMed] [Google Scholar]