

Abstract
Background
African trypanosomes cause lethal diseases in humans and animals and escape host immune attack by switching the expression of Variant Surface Glycoprotein (VSG) genes. The expressed VSGs are located at the ends of telomeric, polycistronic transcription units known as VSG expression sites (VSG-ESs). Each cell has many VSG-ESs but only one is transcribed in bloodstream-form parasites and all of them are inactive upon transmission to the insect vector mid-gut; a subset of monocistronic metacyclic VSG-ESs are then activated in the insect salivary gland. Deep-sequence analyses have been informative but assigning sequences to individual VSG-ESs has been challenging because they each contain closely related expression-site associated genes, or ESAGs, thought to contribute to virulence.
Results
We utilised ART, an in silico short read simulator to demonstrate the feasibility of accurately aligning reads to VSG-ESs. Then, using high-resolution transcriptomes from isogenic bloodstream and insect-stage Lister 427 Trypanosoma brucei, we uncover increased abundance in the insect mid-gut stage of mRNAs from metacyclic VSG-ESs and of mRNAs from the unusual ESAG, ESAG10. Further, we show that the silencing associated with allelic exclusion involves repression focussed at the ends of the VSG-ESs. We also use the approach to report relative fitness costs following ESAG RNAi from a genome-scale screen.
Conclusions
By assigning sequences to individual VSG-ESs we provide new insights into VSG-ES transcription control, allelic exclusion and impacts on fitness. Thus, deeper insights into the expression and function of regulated multi-gene families are more accessible than previously anticipated.
Electronic supplementary material
The online version of this article (doi:10.1186/s12864-016-3154-8) contains supplementary material, which is available to authorized users.
Keywords: Allelic exclusion, Antigenic variation, Gene expression, RNA-seq, Trypanosoma brucei, VSG
Background
African trypanosomes are protozoan parasites that cause devastating diseases known as human African trypanosomiasis and a livestock disease known as nagana. These parasites are transmitted by the bite of an infected tsetse-fly, the distribution of which restricts the geographic spread of the disease. The parasite exists extracellularly, and is continually exposed to immune attack in the mammalian host [1]. To persist in the host bloodstream, the parasite has evolved a sophisticated strategy of antigenic variation and immune evasion. The trypanosome surface is coated in a dense layer of 107 copies of a single variant surface glycoprotein (VSG) [2]. Switching of this VSG coat is central to adaptive immune evasion, and operates at a rate of approximately 10−6 per parasite cell division in culture [3]. In vivo, this leads to the recrudescent parasitaemia characteristic of T. brucei infection [1], where un-switched parasites are removed by antibody mediated killing.
VSG expression sites (VSG-ESs) are the key subtelomeric polycistronic units involved in antigenic variation in bloodstream African trypanosomes [4]. Understanding the expression and function of these units is critical to understanding virulence. VSG-ES transcription, mediated by RNA polymerase I, initiates at multiple VSG-ES promoters but is attenuated in all but one to prevent multi-VSG expression in individual cells [5]. The polycistronic VSG-ESs contain a number of Expression Site Associated Genes (ESAGs), several of which are of unknown function, but those that have been characterised are involved in nutrient acquisition; ESAG6 and ESAG7 [6], and innate immune evasion; ESAG4 and SRA [7, 8]. In trypanosomes, maturation of mRNA from nascent transcripts occurs via the linked processes of trans-splicing, the addition of a 39-nt capped leader sequence, and poly-adenylation [9, 10]. RNA Pol-II transcribes the spliced leader from a repetitive array as a primary 135 b transcript [11], that is processed and 5′ capped before association with the spliceosome [12], which mediates trans-splicing to nascent transcripts [10].
Antigenic variation is specifically required for immune evasion in the bloodstream and, consistent with this, VSG-ESs are subject to developmental regulation. Upon parasite differentiation in the tsetse mid-gut, VSG transcription stops and the VSG coat is shed in the fly mid-gut, where recent evidence shows it interferes with fly innate immunity [13]. Procyclins, a family of repetitive proteins containing either EP or GPEET amino acid repeats, replace the VSG coat in the mid-gut [14]. Following migration to the fly salivary gland, a distinct sub-set of VSGs are expressed on the surface of metacyclic cells from monocistronic VSG-ESs, and are required for re-infection of the mammalian host [15, 16].
Next-generation sequencing (NGS) and RNA-seq approaches in particular, have been used in African trypanosomes to examine a range of features of genome organisation and gene expression, including developmentally regulated transcript expression [17], alternative splicing [18], control by RNA-binding proteins [19] and translation control [20, 21]. The approach has also been used to analyse relative expression levels for transcripts mapping to the active VSG-ES, revealing that most ESAG transcripts are present at 1–0.01 % the level of the active VSG transcript [17, 18].
NGS analysis of VSG-ESs presents several unique challenges. In particular, VSG-ESs are closely related [22] and, although increased mapping stringency can improve the alignment [23], the accuracy of assigning sequence-reads to the correct and specific sites has not been assessed in detail. Genes related to ESAGs (GRESAGs) are also found at non-telomeric locations; copies of GRESAG4 are particularly prevalent and copies of GRESAG2 are present at procyclin loci [24, 25]. In addition, VSG-ESs are under-represented in reference genome-sequence assemblies. Fortunately, the full set of VSG-ESs have been isolated and sequenced [22], and the subset of VSG-ESs expressed in the metacyclic stage has also been identified [15, 26], in the widely studied Lister 427 strain. However, developmental control of VSG-ESs has not yet been analysed in any detail in this strain.
We generated transcriptome data from sub-cloned populations of Lister 427 cells expressing a defined VSG (VSG-2) and from differentiated insect mid-gut stage cultures directly derived from those sub-clones. We then developed computational approaches to determine how accurately short reads derived from NGS can be aligned to VSG-ESs. We find that the differences between VSG-ESs are sufficient to allow 100-b reads to be accurately aligned to specific loci. Subsequent high-stringency mapping revealed a number of unanticipated features regarding VSG-ESs and their developmental control. High-stringency mapping was also applied to published NGS datasets. This revealed specific perturbations to VSG-ES transcriptomes following knockdown or over-expression of the allelic exclusion regulator VEX1 [27], and relative fitness costs following knockdown of individual ESAGs [28].
Results
Transcriptomes from isogenic bloodstream and insect-stage T. brucei
We derived transcriptomes from a pair of Lister 427 bloodstream form sub-clones expressing VSG-2 and from directly derived, differentiated, insect-stage cultures, separated from the bloodstream form sub-clones by only 10 days. Since the core, non sub-telomeric portion of the TREU-927 reference and Lister 427 genomes are closely related, sequence reads were aligned to the 11 megabase chromosomes from the TREU-927 genome [29], plus a non-redundant set of the 14 VSG-ESs [22] and 5 metacyclic VSG-ESs [15] from the Lister 427 strain. We aligned 26.4 and 26.5 million bloodstream form and 49.9 and 53.5 million insect-stage, 100 b, paired end reads using Bowtie2 [30], yielding approximately 180x genome coverage for bloodstream form and 350x for insect-stage transcriptomes. We calculated reads per kilobase of transcript per million mapped reads (RPKM), and differential expression using edgeR [31]; an additional table file shows these values (Additional file 1, ‘BSF v PCF mapq > 0’ tab). This revealed excellent reproducibility between clones, Pearson correlation coefficients of > 0.99 (Additional file 2), and robust developmental control of known bloodstream-form and insect-stage specific genes (Fig. 1a). For example, upon differentiation, the EP and GPEET procyclins increased 306 (EP1 and EP2 average) and 153-fold, respectively, while VSG-2 decreased 1.2 × 106 fold; BES1 ESAG transcripts were also decreased (Fig. 1a). In addition, phosphoglycerate kinase A (PGKA) displayed little change while PGKB increased (3.3-fold) and PGKC decreased (4.7-fold), as expected [32].
Fig. 1.
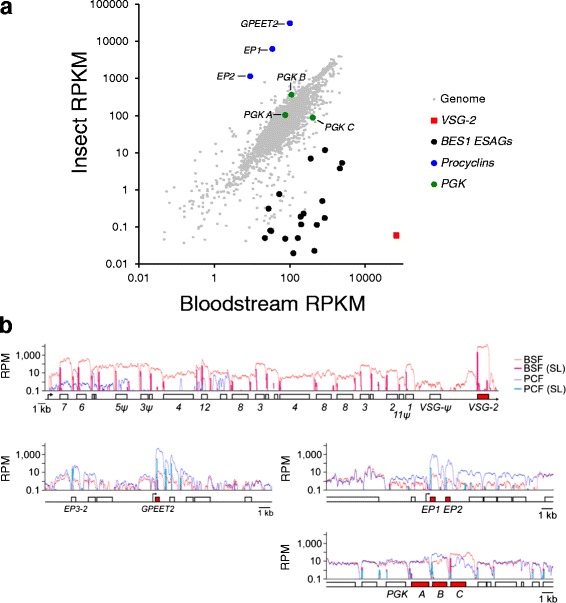
T. brucei transcriptomes from isogenic bloodstream and insect stage cultures. a The scatter plot shows RNA-seq data for bloodstream form and insect stage cultures. Values are calculated as RPKM (Reads Per Kilobase of transcript per Million mapped reads) and are averages for a pair of independent sub-clones. Selected developmentally regulated genes are highlighted. b Base-pair resolution plots of the active VSG-ES (top panel), GPEET2 and EP1/EP2 loci (middle panels) and the phosphoglycerate kinase locus (lower panel). Read counts are normalised using reads per million mapped (RPM) and trans-spliced reads are also indicated. Beneath each panel is a schematic map for each locus. Boxes, coding sequences; arrowsheads, Pol-I promoters
We next analysed reads from bloodstream and insect-stage cells that aligned to the active VSG-ES. A single base resolution BES1 plot (Fig. 1b) revealed a strikingly compact transcription-unit, incorporating little inter-transcript DNA sequence. Reads associated with a trans-spliced leader sequence, found associated with all trypanosomatid mRNAs [10], revealed trans-splicing at discrete points for each gene (Fig. 1b), as expected [33]. We observed multiple trans-splicing events within the VSG gene, but the dominant splice-site was used >1000-fold more frequently than other sites. As also expected, we see bloodstream-specific over-representation (266 fold on average) of transcripts for every ESAG present in the active VSG-ES (Fig. 1b), consistent with transcription attenuation following differentiation [34]. In the bloodstream-form, the VSG transcript itself is 141-fold more abundant than the mean value of the other VSG-ES-derived transcripts. We do see some isolated ESAGs that display higher expression relative to upstream ESAGs following differentiation to the insect stage but, rather than VSG-ES internal transcription initiation, this likely reflects incorrect assignment of reads from GRESAGs that are transcribed by RNA pol-II [24, 25]. Analysis of procyclin loci (also transcribed by RNA Pol-I) and the PGK locus (transcribed by RNA Pol-II) revealed similarly compact transcription units and the expected developmental controls (Fig. 1b). Thus, our RNA-seq datasets from isogenic bloodstream and insect-stage cultures are suitable for more detailed VSG-ES transcriptome analysis.
‘Short’ reads can be accurately assigned to VSG-ESs
We next considered the challenge of accurately assigning 100-b sequence reads from RNA-seq datasets to individual VSG-ESs. Analysis of ESAG7 genes from the Lister 427 strain highlighted the challenge in terms of distinguishing among individual ESAGs (Additional file 3). In this case, a high level of identity was observed throughout the coding-sequence. There are differences however, which can be exploited.
In order to determine the feasibility of accurate short read alignment to VSG-ESs, we used ART [35] to simulate the Illumina sequencing platform in silico and to generate 105 single end, 100-b reads derived exclusively from individual VSG-ESs. We initially simulated Illumina sequencing runs of the sequence isolated from the active VSG-ES in our experimental strain and from three additional VSG-ESs (Fig. 2a). These reads were then aligned to the VSG-ES sequences using Bowtie2 [30], with the same settings used above. We observed reads aligning inappropriately to other VSG-ESs, however the signal to noise ratio could be increased significantly by filtering reads by the uniqueness (MapQ value) of the alignment, assigned to each read by Bowtie2 (Fig. 2a). This filters reads with a higher probability of being mis-aligned to the genome. We went on to repeat this in silico analysis with the entire set of assembled bloodstream and metacyclic VSG-ESs from the Lister 427 strain (Fig. 2b).
Fig. 2.
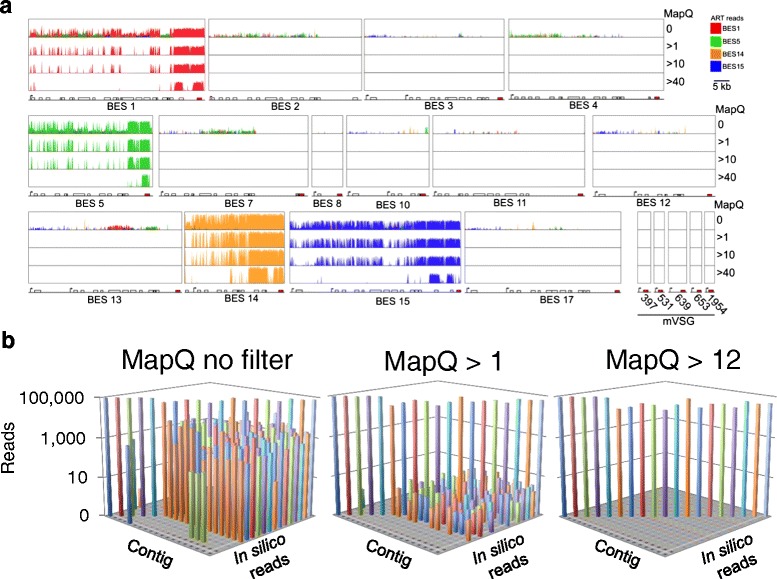
Simulations identity the optimal MapQ setting for accurate VSG-ES mapping. a. In silico reads generated from the ART program for a subset of VSG-ESs were aligned to the genome; only the VSG-ESs are shown on a linear scale. Varying the MapQ value (either no filter – 0, or incrementally > 1, > 10, > 40) to filter non-uniquely mapping reads improves the signal to noise ratio. mVSG, metacyclic VSGs. Beneath each panel is a schematic map for each locus. Boxes, coding sequences; arrowsheads, Pol-I promoters. Red reads BES1, Green reads BES5, Orange reads BES14, Blue reads BES15. b Analysis of all VSG-ESs using the ART method. Plots show reads derived from each contig, and aligned to the genome; only genes in VSG-ESs are shown. MapQ > 12 is the lowest value at which no noise is detected. Metacyclic VSGs are on the left
The analysis indicated that Bowtie2 aligns 75.5 % of in silico generated reads to the correct bloodstream VSG-ES with a MapQ > = 0 (98.7 % to metacyclic VSG-ESs) such that mis-aligned reads, as expected, can have a significant negative impact on transcriptome analysis. A MapQ value of > 1 removes 99.9 % of inappropriately aligned reads and retains 81 % of the signal while a MapQ > 12 eradicated all noise from the data and retained 65.7 % of the signal (Fig. 2b). We selected a MapQ cutoff > 1 as optimal for accurately assigning short sequence reads to individual VSG-ESs; the vast majority of VSG-ES associated genes retain short reads using this approach; an additional table file shows these values (Additional file 1, ‘BSF v PCF mapq > 1’ tab).
During this analysis, we also observed a distinct trend in the distribution of in silico-generated reads. Specifically, reads were more effectively retained closer to the telomere as we increased the uniqueness of alignment (Additional file 4A), indicating that sequences closer to telomeres are more divergent/unique. The alignment map for promoter-proximal ESAG7 genes from Additional file 3 is also compared to an alignment map for the telomere-proximal ESAG1 to further illustrate this point (Additional file 4B). Although a contested hypothesis [36], error-prone VSG gene-conversion was previously suggested as a mechanism contributing to antigenic variation [37, 38]. Our observation is consistent with this hypothesis when taken together with the inherent fragility of sub-telomeres and subsequent telomere-directed gene-conversion events [39].
Differential controls affecting specific ESAGs and VSGs
Using a MapQ > 1, we filtered our RNA-seq data for high-confidence read alignments to VSG-ESs. The major developmental changes reported above (Fig. 1a) were also observed for this filtered dataset (Additional file 5). Filtering allowed us to assign reads to ‘silent’ VSG-ESs (Fig. 3a-b) and this revealed transcripts originating from the majority of VSGs in bloodstream-form cells (Additional file 5). This may reflect low-level expression of other VSG-ESs in cells expressing VSG-2 or may equally reflect low-frequency activation of alternate VSG-ESs. Since our RNA samples were prepared using approximately 100 million cells and switching occurs in approximately one in every 106 cells, we expect approximately 100 distinct switching events to be represented, although this does not so readily explain the expression of metacyclic VSG genes. As transcription is initiated at all VSG-ESs [5], proximity to the promoter may explain this low-level transcription of metacyclic VSGs that we observe in bloodstream form cells (Fig. 3c).
Fig. 3.
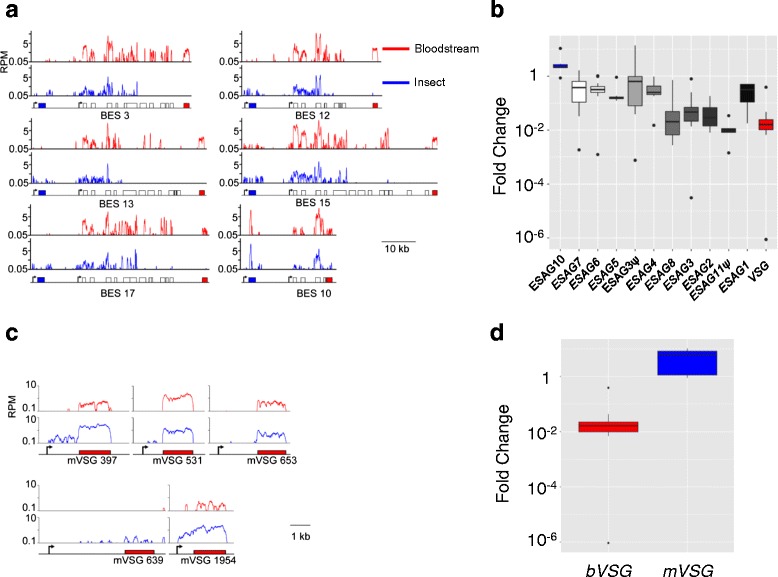
VSG-ES transcriptomes reveal unexpected features. a Base-pair resolution plots of six silent VSG-ESs containing ESAG10 (blue boxes). Read counts were normalised using RPM. Beneath each panel is a schematic map for each locus. Boxes, coding sequences; arrowsheads, Pol-I promoters. Red lines, bloodstream-form; blue lines, insect-stage. b Box plot showing fold-change (log scale) from bloodstream to insect stage, of active and silent ESAGs grouped by type and position in VSG-ESs according to [22]. c Base-pair resolution plots of all five metacyclic-specific VSG-ESs. Red lines, bloodstream-form; blue lines, insect-stage. d Box plot showing fold-change (log scale) from bloodstream to insect stage, of bloodstream (bVSG) and metacyclic (mVSG) VSGs
Reads mapping to ‘silent’ VSG-ESs are 2.4 × 104-fold lower on average relative to those mapping to the active VSG-ES and are further reduced when cells differentiate to the insect stage (Fig. 3a). Again, we see some isolated ESAGs that display higher expression relative to upstream genes, likely reflecting reads from RNA pol-II transcribed GRESAGs [24, 25]. VSG-ESs share a generic structure, with similar ESAGs in similar positions. When grouped and represented according to their position, VSG-ES associated genes closer to telomeres display greater down-regulation in the insect-stage (Fig. 3b). For instance, average VSG expression level decreases 21-fold upon differentiation to insect stage cells, while ESAG7 expression decreased only 2 fold. In contrast to other ESAGs, four of six ESAG10 genes were significantly (p < 0.05) upregulated (average 3.9 fold) in the insect-stage (Fig. 3a-b). This was unexpected since ESAG expression has been considered bloodstream stage-specific [40]. Thus, ESAG10 may be an unconventional ESAG in terms of developmental expression-control.
Another unexpected observation was that, while VSGs in polycistronic VSG-ESs were down-regulated, three of the five VSGs located within the monocistronic metacyclic VSG-ESs were expressed at a significantly higher level (8.4 fold average, p < 10−22) in insect-stage cells (Fig. 3c-d, Additional file 5). VSG expression has been considered to be specific to the tsetse salivary gland stage and the bloodstream-stages, due to promoter control (the VSG-ES promoters active in bloodstream-form cells are distinct from the VSG promoters active in metacyclic cells) and stage-specific stabilisation of transcripts driven by a conserved element in the VSG mRNA 3′ untranslated sequence [41]. We speculate that the unexpected increase in expression of monocistronic metacyclic VSGs reflects progression to a metacyclic (−like) stage by small numbers of cells present in differentiated cultures. Indeed, increased expression of a single RNA binding-protein, RBP6, can trigger this progression through the life cycle [15]. Alternatively, sub-telomeric silencing may be less pronounced in insect-stage cells.
Our analysis of silent VSG-ESs allowed us to identify the trans-splicing sites for twelve VSG-ES linked VSGs and all five metacyclic VSGs from our RNA-seq data (Additional file 6A-B). We counted polypyrimidine tract lengths for VSG splice sites and compared these to the genes in the RNA Pol-I transcribed procyclin loci, counting the number of consecutive pyrimidines and allowing for a single purine interruption. We found that the VSG genes are associated with significantly shorter polypyrimidine tracts (11.5 b, n = 17) compared to genes in the procyclin loci (19.0 b, n = 13, p < 4 × 10−4) or the 20 most abundant RNA Pol-II transcripts in our dataset (20.4 b, n = 20, p < 5 × 10−4) (Additional file 6C). Notably, ESAG7 genes also possess shorter polypyrimidine tracts, suggesting that VSG-ES associated genes do not require extensive polypyrimidine tracts to form abundant mature messenger RNAs. Identification of splice-sites also allowed us to predict 5′-untranslated sequences and we note that there does not appear to be a consensus here; these sequences range in size from 15 to 91 b.
Regulation of VSG-ES transcripts by VEX1
VEX1 (VSG exclusion 1) is an allelic exclusion regulator required for the control of VSG gene expression [27]. In order to further our understanding of control by VEX1, we analysed the RNA-seq datasets generated following VEX1 overexpression (Fig. 4a) or RNAi knockdown (Fig. 4b), which both lead to increased expression of ‘silent’, ES-associated VSG genes [27]; an additional table file shows these values (Additional file 1, ‘VEX1’ tabs). High confidence (MapQ > 1) read alignments, of genes with > 10 reads when VEX1 was overexpressed, revealed an average of 5.4 fold more active VSG-ES ESAG transcripts, with 77 % of genes tested significantly increasing (> 3 fold change, p < 0.05) (Fig. 4a, Additional file 7, red bars), and an average 93-fold increase in silent ESAG transcripts, with 86 % of genes tested increasing significantly (> 3 fold change, p < 0.05) (Fig. 4a, Additional file 7, blue bars). These results are consistent with the previously reported positive control by ectopically expressed VEX1 [27].
Fig. 4.
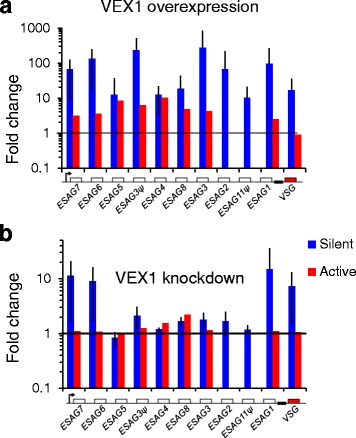
VSG-ES transcriptome analysis following VEX1 perturbation. a Average change of ‘generic’ [22] ESAG expression at either active (red) or silent (blue) VSG-ESs following overexpression of VEX1. b As in A but showing change in gene expression following knockdown of VEX1. Beneath each panel is a schematic map for each locus. Boxes, coding sequences; arrowsheads, Pol-I promoters. Black line crosses at 1 fold change (no change in expression)
We next analysed the RNA-seq data derived following VEX1 RNAi (Fig. 4b, Additional file 7). In this case, our analysis shows minimal impact at the active VSG-ES (Fig. 4b, red bars) but differential behaviour of the ‘silent’ VSGs and the ESAGs at either end of VSG-ESs relative to the centrally located ESAGs. Specifically, the promoter-adjacent ESAG6 and 7 were increased 10-fold; 75 % of genes tested increasing significantly (> 3 fold, p < 0.05), while the telomere-proximal VSG and ESAG1 increased 10 to 15-fold; 50 % of genes tested increasing significantly (> 3 fold, p < 0.05). This is in contrast to several centrally located ESAGs, which increased only 1.5 fold on average, with just 2 % of genes tested increasing significantly (> 3 fold, p < 0.05).
Loss-of-fitness associated with ESAG knockdown
Our understanding of the functions of the ESAGs remains incomplete and, because of difficulties with accurate mapping as outlined above, the ESAGs were not analysed as part of prior high-throughput phenotyping analysis using RNA interference (RNAi) in in vitro culture [28]. We revisited these data and mapped RIT-seq reads (MapQ > 1) from bloodstream and insect stage cells to the VSG-ES active in the bloodstream-form (Fig. 5a); an additional table file shows these values (Additional file 1, ‘BES1 RIT-seq’ tab). Although the RNAi library used in the Lister 427 strain was derived from the TREU 927 strain, the closely related RNAi target fragments are still expected to mediate efficient ESAG knockdown [42]. Mapped reads were quantified and analysed relative to the uninduced control (Fig. 5b) revealing loss-of-fitness following ESAG2, ESAG3 and ESAG8 RNAi that achieved significance following ESAG2 RNAi; other ESAGs failed to register a loss-of-fitness. A previous study reported loss-of-fitness for every ESAG knockdown tested [43] but ESAG2 also registered the greatest loss-of-fitness in that study. This phenotype may reflect the knockdown of RNA pol-II transcribed GRESAG2 transcripts [24, 25] rather than VSG-ES associated ESAG2 transcripts, however. None of the ESAGs, except for ESAG8, registered a loss-of-fitness in the insect-stage, consistent with developmental stage-specific ESAG expression. Notably, several ESAGs may be specifically required for host-parasite interactions, functions that are dispensable in in vitro culture.
Fig. 5.
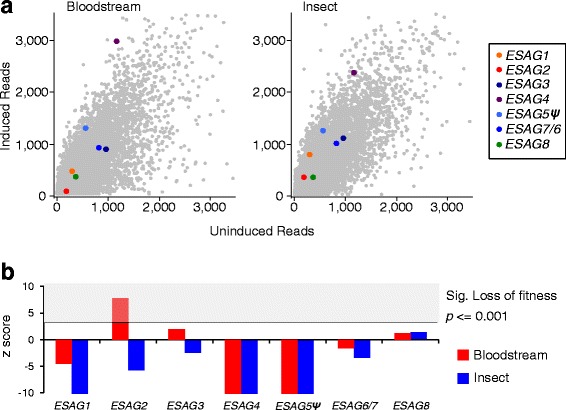
ESAG fitness profiles following high-throughput RNAi. a Scatter plots show genome wide quantification of fitness following ESAG knockdown as defined by RIT-seq in bloodstream (3 days of RNAi induction) and insect-stage cells. b Bar plots show z-scores reporting significant loss of fitness in bloodstream and insect RIT-seq experiments [28]
Discussion
Massive parallel NGS approaches have revolutionised the study of gene regulation and expression, producing data with an unrivalled depth [44], and have been applied with great success in African trypanosomes; see [17, 18, 33] for just a few examples. There are, however, a number of challenges associated with the analysis of subtelomeric sequences in a range of eukaryotes. These regions often incorporate highly repetitive and plastic components of the genome. This is particularly true of parasites, in which subtelomeric genes function in the processes of antigenic variation and immune evasion, such as malaria parasites [45] and African trypanosomes [4].
Analysis of these repetitive loci has proven challenging, a fact exemplified by our in silico analyses. Indeed, we note the use of orthogonal methods such as qRT-PCR and genetic tagging of VSG-ESs [46, 47], despite the availability of NGS approaches and datasets. We now report high-coverage transcriptomes from isogenic T. brucei cultures and from two major life cycle stages, namely the mammal-infective bloodstream form and the tsetse fly mid-gut stage, with a focus on the regulation of VSG-ES transcription. Simulated Illumina sequence data allowed us to gauge an appropriate filter that maximises the signal-to-noise ratio of sequence alignments at these loci. We find the expected extreme developmental regulation of VSG and EP/GPEET surface antigen genes and VSG-ES attenuation in insect stage cells but also, using high-stringency mapping, uncover additional and unexpected features.
Previous reports indicate that alignment of short reads to VSG-ESs can be problematic due to the similarity between these loci [22, 46, 48]. Our analyses show that an average of 24.5 % of VSG-ES derived reads are typically incorrectly aligned to VSG-ESs, and filtering reads with a MapQ value > 1 greatly reduces mis-mapping. Further analyses of VSG-ESs suggest that this is particularly useful for the closely related ESAG sequences. Improvements in sequencing technologies, such as quality (our sequence data has mean per-base quality scores > 34) and read-length now facilitate accurate high-stringency mapping. Specifically, we believe that 100–150 b reads that are commonly produced by current Illumina technologies incorporate sufficient SNPs to allow specific assignment to individual VSG-ESs.
In bloodstream cells, monoallelic expression ensures that a single subtelomere is productively transcribed [1]. However, in mid-gut stage cells, VSG-ESs are silenced [34]. In our populations, we find that transcripts for twelve silent VSG-ES linked and all five metacyclic VSGs are detectable, although at a level approximately 26,000 times lower than the active VSG; illustrating the impressive dynamic range of RNA-seq. In addition, our analysis reveals that ESAG10 mRNAs, encoding putative folate transporters [49], are more abundant in the mid-gut stage cultures than the bloodstream form. This surprising finding is suggestive of less effective silencing in the insect-stage of ESAG10-associated VSG-ES promoters relative to the almost identical VSG-ES promoters located downstream [22]. Alternatively, ESAG10 transcripts may display increased stability in insect-stage cells. Notably, additional RNA pol-II transcribed genes on chromosome 8 (Tb927.8.3620, 3630 and 3650) encode folate transporters and whether the ESAG10-associated promoters or transcripts are ‘activated’ at any point in the life cycle remains unknown.
High-stringency mapping using transcriptomic datasets derived following knockdown of the allelic exclusion regulator, VEX1 [27], revealed derepression of promoter and telomere proximal VSG-ES genes. In another study, ectopic overexpression of a second VSG gene resulted in VSG-ES silencing spreading from the telomere towards the promoter in a disruptor of telomeric silencing 1B (DOT1B) dependent manner [43]. Our analysis indicates that VEX1-mediated silencing is directed at the telomeric VSG and ESAG1 genes, and at the VSG-ES promoter-adjacent ESAG6 and ESAG7 genes. This is in contrast to VEX1 overexpression, which upregulates all silent ESAGs and VSGs. Thus, our current data indicate that VEX1-mediated silencing primarily affects the ends of silent VSG-ESs, suggesting that subtelomere conformation may be important in the control of these genes. Finally, high-stringency phenotyping data confirm (GR)ESAG2 as the ESAG associated with the greatest fitness cost when knocked-down in in vitro culture.
Conclusions
By distinguishing between closely related transcription units, we have been able to enhance our understanding of the behaviour of VSG-ESs in terms of VSG silencing, developmental regulation and contributions to fitness in culture. NGS approaches, coupled to high-stringency mapping, such as RNA-seq, ChIP-seq, RIT-seq and the growing list of ‘seq’ technologies will undoubtedly improve our understanding of the organisation and expression of these virulence gene loci and indeed closely related gene families in a range of other organisms.
Methods
T. brucei
Two subclones of wild type bloodstream-form Lister 427 strain T. brucei expressing VSG-2 (VSG-221, Mitat1.2) were differentiated to insect mid-gut stage cells as previously described [50]. Briefly, cells were collected by centrifugation and resuspended in differentiation medium (DTM) [50] supplemented with 3 mM citrate and 3 mM cis-aconitate and maintained for 10 days at 27 °C, 0 % CO2 (ambient).
RNA-seq
For RNA extraction, 5 × 107 cells were collected and RNA prepared using the Qaigen RNeasy kit, according to the manufacturer’s instructions. Poly-A+ RNA was enriched using oligo-dT beads, and reverse transcribed. Second strand synthesis was randomly primed. Sequencing was performed on the HiSeq platform (Illumina) at the University of Dundee generating 100-b paired-end reads. This yielded insect-stage RNA-seq data, using identical processing, that were only 10-days removed from our bloodstream-form RNA-seq data [27].
In order to align reads we generated a hybrid genome assembly consisting of the 11 megabase chromosomes from the T. brucei 927 reference genome [29], the non-redundant set of 14 bloodstream expression sites [22] and the 5 metacyclic expression sites from our Lister 427 strain [15, 26]. Read alignment was performed using Bowtie2 [30] as previously described [27] using the parameters --very-sensitive --no-discordant. Approximately 25 million bloodstream-form and 50 million insect stage reads were aligned for each clone. Alignment files were manipulated using SAMtools [51], and visualized in the Artemis genome browser [52]. Single base resolution plots were generated using the pysam API (https://github.com/pysam-developers/pysam) in an in-house script that filters reads based on alignment quality (MapQ) and corrects for library size (available on request). Trans-spliced reads were extracted using a previously published script [53], using the partial spliced leader sequence ‘TCTGTACTATATTG’ and it’s reverse complement to search. This is the shortest sequence that returns only spliced leader sequences following BLAST search of the TREU-927 genome sequence on TriTrypDB. Differential expression analysis was performed with edgeR [31] as previously described [27]. When analysing VEX1 perturbation, we excluded genes with <10 reads averaged across replicates in both uninducing or inducing conditions.
ART simulation
ART is a software package that simulates next-generation sequencing runs using empirical error models utilized by the 1000 genomes project [35]. Illumina sequencing runs were simulated for all 19 of the Lister 427 VSG-ES contigs in the hybrid genome using the parameters art_illumina -i contigX.fa -len 100 -ss MS -c 100000. This produced 105 single-end reads for each contig; as the longest contig is 59,781 bp, this provided coverage of every base in each VSG-ES. These in silico reads were then aligned back to the complete genome with Bowtie2 using the parameters --very-sensitive [30]. Alignment files were manipulated using SAMtools [51]. Read counts were generated using the Artemis genome browser [52].
Sequence and data analysis
Clustal alignment analysis and visualisation of ESAG7 sequences was performed using CLC workbench using settings: gap open cost = 0.0, gap extension cost = 0.0, end gap cost = free, alignment mode = very accurate, redo alignments = no, use fixedpoints = yes. A non-redundant gene list was from [28] and VSG-ES sequences [22] were retrieved from TriTrypDB. ‘Generic’ ESAG lists are derived from [22]; ESAGs from each VSG-ES were compiled based on relative position within each VSG-ES.
Acknowledgments
Funding
The work was supported by The Wellcome Trust (Investigator Award 100320/Z/12/Z to D.H. and Strategic Award 100476/Z/12/Z supporting Biological Chemistry & Drug Discovery).
Availability of data and materials
The RNA-seq sequence data reported in this paper have been deposited in the European Nucleotide Archive, www.ebi.ac.uk/ena (accession no. PRJEB8747). Genome sequences assembled from publicly available data (tritrypdb.org) for this paper, and any scripts used are provided on request without condition.
Authors’ contributions
SH and DH designed the study and analysed the data, LG generated samples for RNA-seq. SH, LG and DH wrote the paper. All authors read and approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Consent for publication
Not applicable.
Ethics approval and consent to participate
Not applicable.
Abbreviations
- BES
Bloodstream expression site
- BSF
Bloodstream form
- bVSG
Bloodstream VSG
- ChIP-seq
Chromatin immunoprecipitation sequencing
- DOT1B
Disruptor of telomeric silencing 1B
- DTM
Differentiation trypanosome medium
- ESAG
Expression-site associated gene
- GRESAG
Gene related to ESAG
- mVSG
Metacyclic VSG
- NGS
Next-generation sequencing
- PCF
Procyclic (insect) form
- PGK
Phosphoglycerate kinase
- qRT-PCR
Quantitative reverse transcription polymerase chain reaction
- RBP
RNA binding-protein
- RIT-seq
RNA interference target sequencing
- RNA Pol
RNA polymerase
- RNAi
RNA interference
- RNA-seq
RNA sequencing
- RPKM
Reads per kilobase of transcript per million mapped reads
- RPM
Reads per million mapped
- SRA
Serum resistance-associated
- TREU
Trypanosomiasis research Edinburgh University
- VEX1
VSG exclusion 1
- VSG
Variant surface glycoprotein
- VSG-ES
VSG expression site
Additional files
Table containing RNA-seq and RIT-seq data. (Excel 7530 kb)
Comparison of RNA-seq replicates. Scatter plots of bloodstream-form and insect-stage RPKM values for each condition. Data used to derive plots in Fig. 1 (MapQ > =0). (PDF 28 kb)
ESAG7 sequences are highly similar between VSG-ESs. Clustal alignment between ESAG7 nucleotide sequence in BES1 (Tb429.BES40.2) and other ESAG7 sequences. BES2, 4 and 8 have multiple ESAG7 genes. (PDF 238 kb)
Divergence increases towards the telomere. A. RPKM values for each ESAG and VSG in the non-redundant set plotted against the distance from the start-codon to the telomere. B. ESAG7 Clustal alignment plot from Additional file 3 compared to ESAG1 Clustal alignment plot. (PDF 169 kb)
High stringency RNA-seq analysis. Scatter plot of RPKM values of each condition with MapQ value > 1. Values are calculated as RPKM (Reads Per Kilobase of transcript per Million mapped reads) and are averages for a pair of independent sub-clones. Selected developmentally regulated genes are highlighted. Metacyclic (mVSG) and bloodstream (bVSG) specific VSGs are also shown. (PDF 23 kb)
VSG genes have significantly shorter polypyrimidine tracts upstream. A. Logo plots of the 17 VSG trans-splice sites identified from our data set (top) and genes in the RNA Pol-I transcribed procyclin loci. Pyrimidines, red; purines, grey. B. Individual trans-slice sites from beta-tubulin (top), bloodstream VSG-ES linked VSGs and metacyclic VSGs. Pyrimidines, red; purines, grey. C. Bar charts show average polypyrimidine tract length comparisons over several gene groups. ***, P <0.0005. (PDF 1442 kb)
Individual VSG-ES transcriptome analysis following VEX1 perturbation. Bar-charts show fold-changes for ESAG expression in each VSG-ES following either VEX1 knockdown (blue bars) or overexpression (red bars). (PDF 32 kb)
Contributor Information
Sebastian Hutchinson, Email: s.y.hutchinson@dundee.ac.uk.
Lucy Glover, Email: lucy.glover@pasteur.fr.
David Horn, Phone: +44 1382 386286, Email: d.horn@dundee.ac.uk.
References
- 1.Horn D. Antigenic variation in African trypanosomes. Mol Biochem Parasitol. 2014;195(2):123–129. doi: 10.1016/j.molbiopara.2014.05.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Cross GA. Identification, purification and properties of clone-specific glycoprotein antigens constituting the surface coat of Trypanosoma brucei. Parasitology. 1975;71(3):393–417. doi: 10.1017/S003118200004717X. [DOI] [PubMed] [Google Scholar]
- 3.Boothroyd CE, Dreesen O, Leonova T, Ly KI, Figueiredo LM, Cross GA, Papavasiliou FN. A yeast-endonuclease-generated DNA break induces antigenic switching in Trypanosoma brucei. Nature. 2009;459(7244):278–281. doi: 10.1038/nature07982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Glover L, Hutchinson S, Alsford S, McCulloch R, Field MC, Horn D. Antigenic variation in African trypanosomes: the importance of chromosomal and nuclear context in VSG expression control. Cell Microbiol. 2013;15(12):1984–1993. doi: 10.1111/cmi.12215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Kassem A, Pays E, Vanhamme L. Transcription is initiated on silent variant surface glycoprotein expression sites despite monoallelic expression in Trypanosoma brucei. Proc Natl Acad Sci U S A. 2014;111(24):8943–8948. doi: 10.1073/pnas.1404873111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Schell D, Evers R, Preis D, Ziegelbauer K, Kiefer H, Lottspeich F, Cornelissen AW, Overath P. A transferrin-binding protein of Trypanosoma brucei is encoded by one of the genes in the variant surface glycoprotein gene expression site. EMBO J. 1991;10(5):1061–1066. doi: 10.1002/j.1460-2075.1991.tb08045.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Salmon D, Vanwalleghem G, Morias Y, Denoeud J, Krumbholz C, Lhomme F, Bachmaier S, Kador M, Gossmann J, Dias FB, et al. Adenylate cyclases of Trypanosoma brucei inhibit the innate immune response of the host. Science. 2012;337(6093):463–466. doi: 10.1126/science.1222753. [DOI] [PubMed] [Google Scholar]
- 8.Xong HV, Vanhamme L, Chamekh M, Chimfwembe CE, Van Den Abbeele J, Pays A, Van Meirvenne N, Hamers R, De Baetselier P, Pays E. A VSG expression site-associated gene confers resistance to human serum in Trypanosoma rhodesiense. Cell. 1998;95(6):839–846. doi: 10.1016/S0092-8674(00)81706-7. [DOI] [PubMed] [Google Scholar]
- 9.Matthews KR, Tschudi C, Ullu E. A common pyrimidine-rich motif governs trans-splicing and polyadenylation of tubulin polycistronic pre-mRNA in trypanosomes. Genes Dev. 1994;8(4):491–501. doi: 10.1101/gad.8.4.491. [DOI] [PubMed] [Google Scholar]
- 10.Parsons M, Nelson RG, Watkins KP, Agabian N. Trypanosome mRNAs share a common 5′ spliced leader sequence. Cell. 1984;38(1):309–316. doi: 10.1016/0092-8674(84)90552-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Milhausen M, Nelson RG, Sather S, Selkirk M, Agabian N. Identification of a small RNA containing the trypanosome spliced leader: a donor of shared 5′ sequences of trypanosomatid mRNAs? Cell. 1984;38(3):721–729. doi: 10.1016/0092-8674(84)90267-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Tschudi C, Ullu E. Destruction of U2, U4, or U6 small nuclear RNA blocks trans splicing in trypanosome cells. Cell. 1990;61(3):459–466. doi: 10.1016/0092-8674(90)90527-L. [DOI] [PubMed] [Google Scholar]
- 13.Aksoy E, Vigneron A, Bing X, Zhao X, O’Neill M, Wu YN, Bangs JD, Weiss BL, Aksoy S. Mammalian African trypanosome VSG coat enhances tsetse’s vector competence. Proc Natl Acad Sci U S A. 2016;113(25):6961–6966. doi: 10.1073/pnas.1600304113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Roditi I, Schwarz H, Pearson TW, Beecroft RP, Liu MK, Richardson JP, Buhring HJ, Pleiss J, Bulow R, Williams RO, et al. Procyclin gene expression and loss of the variant surface glycoprotein during differentiation of Trypanosoma brucei. J Cell Biol. 1989;108(2):737–746. doi: 10.1083/jcb.108.2.737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kolev NG, Ramey-Butler K, Cross GA, Ullu E, Tschudi C. Developmental progression to infectivity in Trypanosoma brucei triggered by an RNA-binding protein. Science. 2012;338(6112):1352–1353. doi: 10.1126/science.1229641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Sharma R, Gluenz E, Peacock L, Gibson W, Gull K, Carrington M. The heart of darkness: growth and form of Trypanosoma brucei in the tsetse fly. Trends Parasitol. 2009;25(11):517–524. doi: 10.1016/j.pt.2009.08.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Siegel TN, Hekstra DR, Wang X, Dewell S, Cross GA. Genome-wide analysis of mRNA abundance in two life-cycle stages of Trypanosoma brucei and identification of splicing and polyadenylation sites. Nucleic Acids Res. 2010;38(15):4946–4957. doi: 10.1093/nar/gkq237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Nilsson D, Gunasekera K, Mani J, Osteras M, Farinelli L, Baerlocher L, Roditi I, Ochsenreiter T. Spliced leader trapping reveals widespread alternative splicing patterns in the highly dynamic transcriptome of Trypanosoma brucei. PLoS Pathog. 2010;6(8):e1001037. doi: 10.1371/journal.ppat.1001037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Wurst M, Seliger B, Jha BA, Klein C, Queiroz R, Clayton C. Expression of the RNA recognition motif protein RBP10 promotes a bloodstream-form transcript pattern in Trypanosoma brucei. Mol Microbiol. 2012;83(5):1048–1063. doi: 10.1111/j.1365-2958.2012.07988.x. [DOI] [PubMed] [Google Scholar]
- 20.Jensen BC, Ramasamy G, Vasconcelos EJ, Ingolia NT, Myler PJ, Parsons M. Extensive stage-regulation of translation revealed by ribosome profiling of Trypanosoma brucei. BMC Genomics. 2014;15:911. doi: 10.1186/1471-2164-15-911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Vasquez JJ, Hon CC, Vanselow JT, Schlosser A, Siegel TN. Comparative ribosome profiling reveals extensive translational complexity in different Trypanosoma brucei life cycle stages. Nucleic Acids Res. 2014;42(6):3623–3637. doi: 10.1093/nar/gkt1386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Hertz-Fowler C, Figueiredo LM, Quail MA, Becker M, Jackson A, Bason N, Brooks K, Churcher C, Fahkro S, Goodhead I, et al. Telomeric expression sites are highly conserved in Trypanosoma brucei. PLoS One. 2008;3(10):e3527. doi: 10.1371/journal.pone.0003527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Schulz D, Mugnier MR, Paulsen EM, Kim HS, Chung CW, Tough DF, Rioja I, Prinjha RK, Papavasiliou FN, Debler EW. Bromodomain proteins contribute to maintenance of bloodstream form stage identity in the African trypanosome. PLoS Biol. 2015;13(12):e1002316. doi: 10.1371/journal.pbio.1002316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Berberof M, Pays A, Pays E. A similar gene is shared by both the variant surface glycoprotein and procyclin gene transcription units of Trypanosoma brucei. Mol Cell Biol. 1991;11(3):1473–1479. doi: 10.1128/MCB.11.3.1473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Paindavoine P, Rolin S, Van Assel S, Geuskens M, Jauniaux JC, Dinsart C, Huet G, Pays E. A gene from the variant surface glycoprotein expression site encodes one of several transmembrane adenylate cyclases located on the flagellum of Trypanosoma brucei. Mol Cell Biol. 1992;12(3):1218–1225. doi: 10.1128/MCB.12.3.1218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Cross GA, Kim HS, Wickstead B. Capturing the variant surface glycoprotein repertoire (the VSGnome) of Trypanosoma brucei Lister 427. Mol Biochem Parasitol. 2014;195(1):59–73. doi: 10.1016/j.molbiopara.2014.06.004. [DOI] [PubMed] [Google Scholar]
- 27.Glover L, Hutchinson S, Alsford S, Horn D. VEX1 controls the allelic exclusion required for antigenic variation in trypanosomes. Proc Natl Acad Sci U S A. 2016;113:7225–7230. doi: 10.1073/pnas.1600344113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Alsford S, Turner DJ, Obado SO, Sanchez-Flores A, Glover L, Berriman M, Hertz-Fowler C, Horn D. High-throughput phenotyping using parallel sequencing of RNA interference targets in the African trypanosome. Genome Res. 2011;21(6):915–924. doi: 10.1101/gr.115089.110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Berriman M, Ghedin E, Hertz-Fowler C, Blandin G, Renauld H, Bartholomeu DC, Lennard NJ, Caler E, Hamlin NE, Haas B, et al. The genome of the African trypanosome Trypanosoma brucei. Science. 2005;309(5733):416–422. doi: 10.1126/science.1112642. [DOI] [PubMed] [Google Scholar]
- 30.Langmead B, Salzberg SL. Fast gapped-read alignment with Bowtie 2. Nat Methods. 2012;9(4):357–359. doi: 10.1038/nmeth.1923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Robinson MD, McCarthy DJ, Smyth GK. edgeR: a bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics. 2010;26(1):139–140. doi: 10.1093/bioinformatics/btp616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Blattner J, Clayton CE. The 3′-untranslated regions from the Trypanosoma brucei phosphoglycerate kinase-encoding genes mediate developmental regulation. Gene. 1995;162(1):153–156. doi: 10.1016/0378-1119(95)00366-E. [DOI] [PubMed] [Google Scholar]
- 33.Kolev NG, Franklin JB, Carmi S, Shi H, Michaeli S, Tschudi C. The transcriptome of the human pathogen Trypanosoma brucei at single-nucleotide resolution. PLoS Pathog. 2010;6(9):e1001090. doi: 10.1371/journal.ppat.1001090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Rudenko G, Blundell PA, Taylor MC, Kieft R, Borst P. VSG gene expression site control in insect form Trypanosoma brucei. EMBO J. 1994;13(22):5470–5482. doi: 10.1002/j.1460-2075.1994.tb06882.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Huang W, Li L, Myers JR, Marth GT. ART: a next-generation sequencing read simulator. Bioinformatics. 2012;28(4):593–594. doi: 10.1093/bioinformatics/btr708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Graham VS, Barry JD. Is point mutagenesis a mechanism for antigenic variation in Trypanosoma brucei? Mol Biochem Parasitol. 1996;79(1):35–45. doi: 10.1016/0166-6851(96)02642-4. [DOI] [PubMed] [Google Scholar]
- 37.Lu Y, Hall T, Gay LS, Donelson JE. Point mutations are associated with a gene duplication leading to the bloodstream reexpression of a trypanosome metacyclic VSG. Cell. 1993;72(3):397–406. doi: 10.1016/0092-8674(93)90116-8. [DOI] [PubMed] [Google Scholar]
- 38.Rice-Ficht AC, Chen KK, Donelson JE. Point mutations during generation of expression-linked extra copy of trypanosome surface glycoprotein gene. Nature. 1982;298(5875):676–679. doi: 10.1038/298676a0. [DOI] [PubMed] [Google Scholar]
- 39.Glover L, Alsford S, Horn D. DNA break site at fragile subtelomeres determines probability and mechanism of antigenic variation in African trypanosomes. PLoS Pathog. 2013;9(3):e1003260. doi: 10.1371/journal.ppat.1003260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Gottesdiener K, Chung HM, Brown SD, Lee MG, Van der Ploeg LH. Characterization of VSG gene expression site promoters and promoter-associated DNA rearrangement events. Mol Cell Biol. 1991;11(5):2467–2480. doi: 10.1128/MCB.11.5.2467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Berberof M, Vanhamme L, Tebabi P, Pays A, Jefferies D, Welburn S, Pays E. The 3′-terminal region of the mRNAs for VSG and procyclin can confer stage specificity to gene expression in Trypanosoma brucei. EMBO J. 1995;14(12):2925–2934. doi: 10.1002/j.1460-2075.1995.tb07292.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Durand-Dubief M, Kohl L, Bastin P. Efficiency and specificity of RNA interference generated by intra- and intermolecular double stranded RNA in Trypanosoma brucei. Mol Biochem Parasitol. 2003;129(1):11–21. doi: 10.1016/S0166-6851(03)00071-9. [DOI] [PubMed] [Google Scholar]
- 43.Batram C, Jones NG, Janzen CJ, Markert SM, Engstler M. Expression site attenuation mechanistically links antigenic variation and development in Trypanosoma brucei. eLife. 2014;3:e02324. doi: 10.7554/eLife.02324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Goodwin S, McPherson JD, McCombie WR. Coming of age: ten years of next-generation sequencing technologies. Nat Rev Genet. 2016;17(6):333–351. doi: 10.1038/nrg.2016.49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Guizetti J, Scherf A. Silence, activate, poise and switch! mechanisms of antigenic variation in plasmodium falciparum. Cell Microbiol. 2013;15(5):718–726. doi: 10.1111/cmi.12115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Reynolds D, Hofmeister BT, Cliffe L, Alabady M, Siegel TN, Schmitz RJ, Sabatini R. Histone H3 variant regulates RNA polymerase II transcription termination and dual strand transcription of siRNA loci in trypanosoma brucei. PLoS Genet. 2016;12(1):e1005758. doi: 10.1371/journal.pgen.1005758. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Pena AC, Pimentel MR, Manso H, Vaz-Drago R, Pinto-Neves D, Aresta-Branco F, Rijo-Ferreira F, Guegan F, Pedro Coelho L, Carmo-Fonseca M, et al. Trypanosoma brucei histone H1 inhibits RNA polymerase I transcription and is important for parasite fitness in vivo. Mol Microbiol. 2014;93(4):645–663. doi: 10.1111/mmi.12677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Nguyen TN, Muller LS, Park SH, Siegel TN, Gunzl A. Promoter occupancy of the basal class I transcription factor A differs strongly between active and silent VSG expression sites in Trypanosoma brucei. Nucleic Acids Res. 2014;42(5):3164–3176. doi: 10.1093/nar/gkt1301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Gottesdiener KM. A new VSG expression site-associated gene (ESAG) in the promoter region of Trypanosoma brucei encodes a protein with 10 potential transmembrane domains. Mol Biochem Parasitol. 1994;63(1):143–151. doi: 10.1016/0166-6851(94)90017-5. [DOI] [PubMed] [Google Scholar]
- 50.Overath P, Czichos J, Haas C. The effect of citrate/cis-aconitate on oxidative metabolism during transformation of Trypanosoma brucei. Eur J Biochem. 1986;160(1):175–182. doi: 10.1111/j.1432-1033.1986.tb09955.x. [DOI] [PubMed] [Google Scholar]
- 51.Li H, Handsaker B, Wysoker A, Fennell T, Ruan J, Homer N, Marth G, Abecasis G, Durbin R, Genome Project Data Processing S The sequence alignment/Map format and SAMtools. Bioinformatics. 2009;25(16):2078–2079. doi: 10.1093/bioinformatics/btp352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Carver T, Harris SR, Berriman M, Parkhill J, McQuillan JA. Artemis: an integrated platform for visualization and analysis of high-throughput sequence-based experimental data. Bioinformatics. 2012;28(4):464–469. doi: 10.1093/bioinformatics/btr703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Glover L, Alsford S, Baker N, Turner DJ, Sanchez-Flores A, Hutchinson S, Hertz-Fowler C, Berriman M, Horn D. Genome-scale RNAi screens for high-throughput phenotyping in bloodstream-form African trypanosomes. Nat Protoc. 2015;10(1):106–133. doi: 10.1038/nprot.2015.005. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The RNA-seq sequence data reported in this paper have been deposited in the European Nucleotide Archive, www.ebi.ac.uk/ena (accession no. PRJEB8747). Genome sequences assembled from publicly available data (tritrypdb.org) for this paper, and any scripts used are provided on request without condition.