

Abstract
Study Objectives:
To estimate genetic and environmental influences on the associations between insomnia and depression symptoms concurrently and longitudinally.
Methods:
Behavioral genetic analyses were conducted on data from the British longitudinal G1219 twin/sibling study. One thousand five hundred fiftysix twins and siblings participated at Time 1 (mean age = 20.3 years, SD = 1.76). Eight hundred sixty-two participated at Time 2 (mean age = 25.2 years, SD = 1.73 years). Participants completed the Insomnia Symptoms Questionnaire and the Short Mood and Feelings Questionnaire to assess symptoms of insomnia and depression respectively.
Results:
Genetic effects accounted for 33% to 41% of the variance of the phenotypes. The phenotypic correlations were moderate (r = 0.34 to r = 0.52). The genetic correlations between the variables were high (0.73–1.00). Genetic effects accounted for a substantial proportion of the associations between variables (50% to 90%). Non-shared environmental effects explained the rest of the variance and covariance of the traits.
Conclusions:
While genetic effects play a modest role in insomnia and depression symptoms separately, they appear to play a more central role in concurrent and longitudinal associations between these phenotypes. This should be acknowledged in theories explaining these common associations.
Citation:
Gregory AM, Rijsdijk FV, Eley TC, Buysse DJ, Schneider MN, Parsons M, Barclay NL. A longitudinal twin and sibling study of associations between insomnia and depression symptoms in young adults. SLEEP 2016;39(11):1985–1992.
Keywords: insomnia, depression, genetics, twins, longitudinal
Significance.
In this twin/sibling study, we show that insomnia and depression symptoms share common genetic influences. This study is the first to demonstrate substantial genetic contributions to the concurrent and longitudinal associations between insomnia and depression in young adults. Non-shared environmental influences for insomnia and depression symptoms also overlapped, but to a lesser extent. The non-shared environment contributed to both concurrent and longitudinal associations between variables. This knowledge can be usefully employed in studies attempting to identify genetic variants, and specify non-shared environmental influences involved in insomnia and depression.
INTRODUCTION
Longitudinal associations between insomnia and depression symptoms are well established.1–3 While there is some suggestion that insomnia symptoms are more likely to predict depression than vice versa, a recent review of the literature concluded that the best evidence to date suggests that, within adulthood, the associations are likely bidirectional.1
What is less well established is the reason for the association between poor sleep and depression. Explanations are diverse and include the suggestion that insomnia and depression symptoms are due to hyperarousal4 or heightened emotional reactivity.5 Neural processes underlying the association include the possibility that poor sleep moderates the association between amygdala response to threat stimuli and depression.6,7 Sleep disruption may also disturb prefrontal cortex functioning,8 which plays an important role in the emotional and cognitive processing central to depression.9 Other explanations have highlighted the possibility that fragmentation of rapid eye movement (REM) sleep could lead to both the subjective sense of having slept poorly and depression.10
The extent to which genetic predisposition underlies such processes and explains the association between insomnia symptoms and depression has received limited attention.11 Nonetheless, initial work in this domain has been enlightening and genetically informative studies may further clarify the association between insomnia and depression.4 Twin studies to date typically focus on concurrent associations between sleep disturbances and depression symptoms. Genetic effects appear to play a role in explaining these associations in childhood, adolescence, early adulthood, and in adulthood more generally.12–15 These studies suggest that sleep disturbances are perhaps part of the same genetic cluster of disorders as depression (which is also considered to have strong genetic links to anxiety16).
Fewer twin studies have attempted to establish the direction of effects between sleep disturbances and depression or have tried to understand the longitudinal links between the pheno-types. One used cross-sectional genetic analyses to attempt to establish the direction of causation between poor sleep and depression in twins aged 18–87 years old.17 The authors concluded that while depression caused poor sleep (and not vice versa) in young females, there was evidence for reciprocal causation between the phenotypes in older females. The direction of causation was less clear for males, who were underrepresented in the sample. A longitudinal study by our own team found that within childhood, generally defined sleep problems at 8 years of age predicted depression symptoms at 10 years.18 Overall it was concluded that genetic influences may be the most important source of variance that is carried over across time points, although the sample was too small to draw definitive conclusions.
Understanding the mechanisms underlying longitudinal links between phenotypes may help to identify those at risk for later problems, with the ultimate aim of developing preventative programs or improving treatment. Given the importance of the research question and the limited research addressing this topic to date, the aim of this study was to assess a sample of young adult twins and siblings19 over a 5-year period in order to examine both concurrently and longitudinally:
associations between insomnia and depression symptoms;
genetic and environmental influences on the associations between these phenotypes.
METHODS
Participants
The present analyses focus on waves 4 and 5 of the G1219 twin/ sibling study.19 At wave 4 (for ease of presentation referred to herein as Time 1), a total of 1,556 individuals participated in the G1219 study. At wave 5 (referred to as Time 2), a total of 862 individuals participated. At these waves of the study, written informed consent was obtained from the participants themselves. The mean age at Time 1 was 20.33 years (SD = 1.76, range = 18–27 years), and 62% of the sample were female. The mean age at Time 2 was 25.18 years (SD = 1.73, range = 22–32 years), and 66% of the sample were female. The majority of participants were close in age within each time-point (at Time 1, 90% of the participants were aged 18–22 years, and at Time 2, 90% of the participants were aged 22–27 years), but the inclusion of siblings inevitably created some age spread.
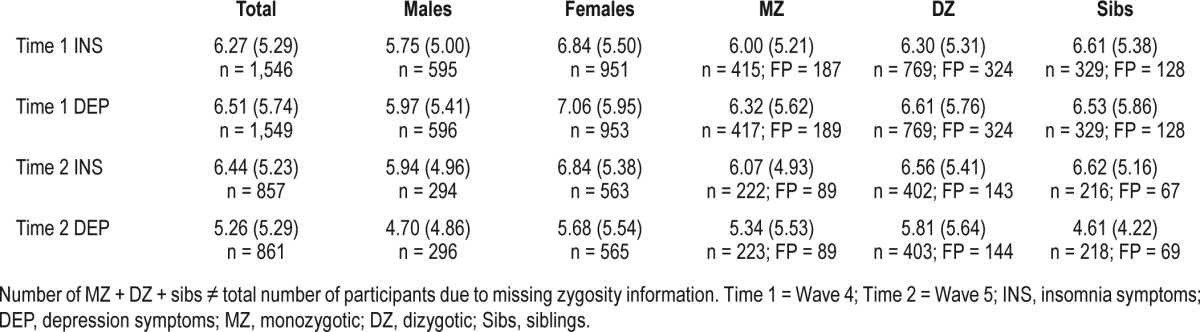
Zygosity was established through a questionnaire measure completed by mothers at waves 2 and 3 assessing physical similarities between twins.20 Table 1 shows the number of monozygotic (MZ), dizygotic (DZ) and sibling individuals as well as the number of full pairs for each variable.
Table 1.
Means (SD) for raw insomnia and depression scores and number of individual (n) and full pairs (FP).
Ethics Approval
Ethics approval for the collection of data at different waves comes from the Research Ethics Committees of the Institute of Psychiatry, South London and Maudsley NHS trust, and/ or Goldsmiths, University of London Ethics Committee. The protocol for data collection conformed to international ethical standards.21
Predictors of Attrition
Logistic regression, focussing on variables which have previously been associated with attrition in this sample, demonstrated that responders at Time 2 were significantly more likely than non-responders to be female (odds ratio = 0.60, P < 0.001) and have more highly educated mothers (odds ratio = 1.14, P < 0.001, where mother's education was rated from the lowest level 0 to 8). There were no significant differences between respondents and non-respondents in terms of housing, insomnia symptoms, depression symptoms, or prosocial behaviour at earlier waves (all P's > 0.05).
Measures
Insomnia Symptoms
Insomnia symptoms were measured using 6 items from the Insomnia Symptoms Questionnaire22 (e.g., difficulty falling asleep; difficulty staying asleep). Items measure the frequency of insomnia symptoms occurring per week during the past month on a 5-point Likert scale (0 = never/don't know; 1 = rarely; 2 = sometimes; 3 = frequently; 4 = always). The total scale score is the sum of these responses (ranging from 0 to 24; higher scores = more severe insomnia). The total score demonstrated good internal consistency in the total sample (Cronbach α = 0.87 at both time-points).
Depression
Depression symptoms were measured using the Short Mood and Feelings Questionnaire (SMFQ).23 This 13-item measure assesses key symptoms of depression (e.g. “I felt miserable or unhappy”) over the previous 2 weeks. Responses are given on a 3-point Likert scale (0 = not true; 1 = sometimes; 2 = true). The psychometric properties of the SMFQ are well established24,25 and in the current study demonstrated good internal consistency (Cronbach α = 0.90 at both time points). While this measure was originally designed for use with children, it has been validated in older adolescents/young adults.26 Of note, none of the items in the depression scale measured sleep. One item, “I felt so tired I just sat around and did nothing” tapped fatigue, which could be related to a plethora of different conditions including depression, diabetes, or anemia.27 As this item could also have reflected a sleep disorder, we conducted preliminary sensitivity analyses, by excluding this item from our measure of depression at Time 2. Correlations with insomnia symptoms did not change meaningfully. We therefore focused on the full depression scale in the analyses reported here.
Statistical Analyses
Data Preparation
Skew was not problematic for insomnia symptoms from the total sample at either time-point (Time 1 insomnia symptoms mean = 6.27 [SD = 5.29]; skew = 0.79, [SE = 0.06]; Time 2 insomnia symptoms mean = 6.44 [5.23]; skew = 0.73, [SE = 0.08]). Depression symptom scores were positively skewed at both time-points (Time 1 depression symptoms mean = 6.44 [SD = 5.23]; skew = 1.26, [SE = 0.06]; Time 2 depression symptoms mean = 5.26 [5.29]; skew = 1.42 [SE = 0.08]). Depression symptom scores were therefore log transformed prior to model fitting analyses. This successfully reduced skew (Time 1 depression skew = −0.35 [SE = 0.06]; Time 2 depression symptoms skew = −0.15 [SE = 0.08]). Outliers of ≥ 3 standard deviations above or below the means on all variables were omitted (n = 6 cases). Additionally, prior to analysis the variables were age and sex regressed as a standard procedure for twin modelling.28
Analytic Principles using Twin and Sibling Data
Twin studies compare the similarity within MZ twin pairs to the similarity within DZ twin pairs (with additional full sibling pairs) to estimate genetic and environmental influences on traits. Since MZ twins share 100% of their segregating genes while DZ twins and full siblings share on average half, the difference in covariance is used to estimate the relative contribution of 3 sources of variance impacting on a phenotype: additive genetic influences (A) (where alleles at all loci “add up” to influence behavior); shared environmental influences (C) (environmental influences that act to make individuals within a family similar); and non-shared environmental influences, (E) (environmental influences that act to make individuals within a family different; this source of variance also incorporates measurement error). If the correlation between MZ pairs is greater than that of DZ/sibling pairs, genetic influences are indicated. MZ twin correlations equivalent to DZ/sibling correlations indicate the importance of the shared environment, and the extent to which MZ twin correlations are smaller than 1 indicates the magnitude of non-shared environmental factors.29
Descriptive Statistics, Phenotypic, and Twin Correlations
Descriptive statistics for the untransformed scores are first presented (Table 1). Then phenotypic correlations are estimated in a constrained model which specifies: one overall set of means and within-individual cross-trait correlations (Table 2, i.e. equated across groups and twin/sibling order). Next, a set of MZ, DZ, and sibling cross-twins/siblings, within-trait correlations; and a set of MZ, DZ, and sibling cross-twins/siblings, cross-trait correlations are presented (Table 3). The twin/sibling correlations provide an indication of the relative sources of variance on individual phenotypes and their associations.
Table 2.
Within person phenotypic correlations between insomnia and depression symptoms at times 1 and 2 (95% confidence intervals in parentheses).

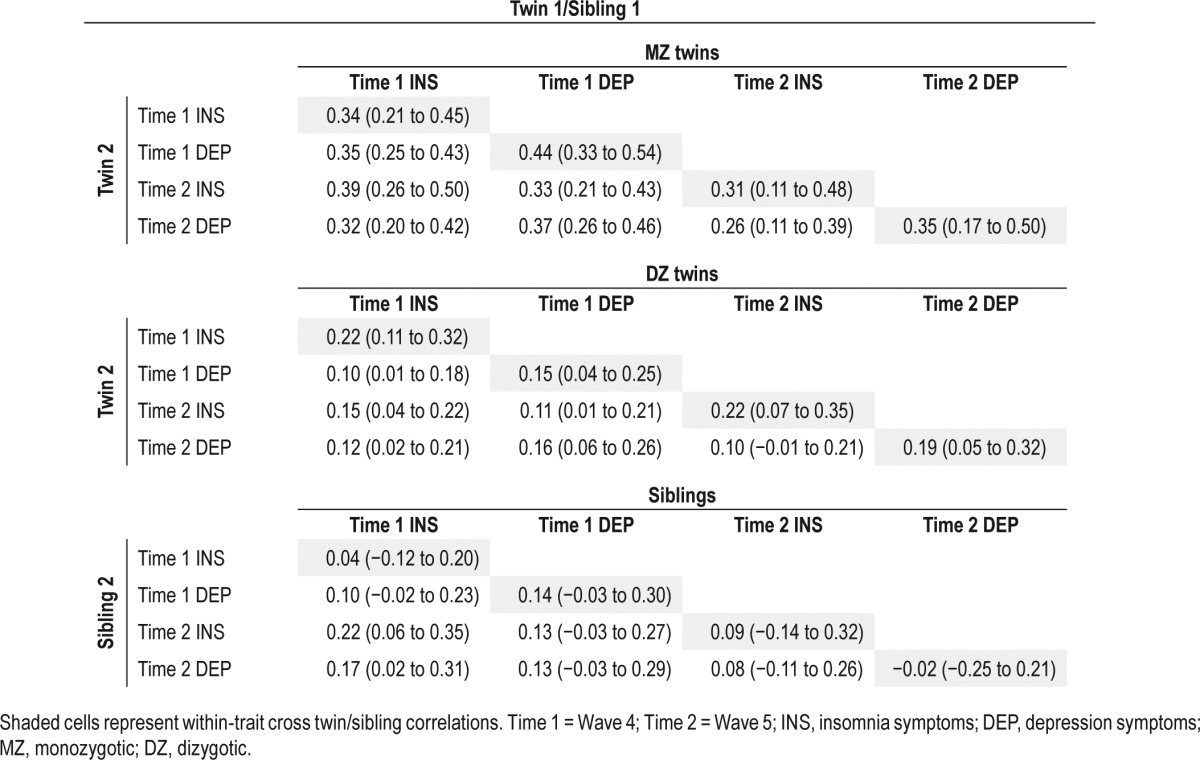
Table 3.
Twin correlations (within and across traits) for insomnia and depression symptoms at times 1 and 2 (95% confidence intervals in parentheses).
Multivariate Genetic Model Fitting
A correlated factors model was fitted to the 2-time point data to examine genetic and environmental influences on the association between phenotypes (Figure 1). This model specifies a different set of male and female loadings for the genetic, shared, and non-shared environmental factors influencing each variable in the model (qualitative sex differences), while the correlations between the A, C, and E factors are the same across sex. This specification is necessary when modelling multivariate data containing opposite sex pairs.30 This was followed by a model in which the male and female parameters were constrained to be equal, providing a 12 df sex-differences test. Finally, we tested an AE model to see whether we could drop the C parameters without a significant decrease in fit.
Figure 1.
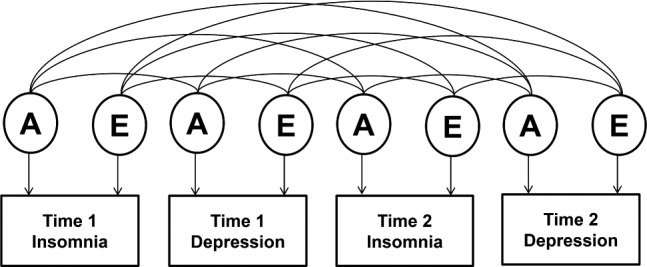
Multivariate correlated factors model. A, additive genetic influence; E, non-shared environmental influence.
Analyses were conducted in the program OpenMx, a widely used program for analyzing genetically sensitive data, using raw maximum likelihood estimation.31 The fit statistic provided for raw data analysis is minus twice the log likelihood of the observations (−2LL). Differences in fit between models are evaluated by likelihood ratio testing since the difference in −2LL is χ2 distributed. The 95% confidence intervals provide information about the precision of parameter estimates.
RESULTS
Descriptives
Table 1 shows the means and standard deviations of the raw insomnia symptoms and depression symptom scores for both Time 1 and 2, split by sex and zygosity. Compared to males, females experienced greater insomnia symptoms at Time 1 (χ2 = 14.38, df = 1, P < 0.001) and at Time 2 (χ2 = 6.82, df = 1, P < 0.01). Compared to males, females also experienced greater depressive symptoms at Time 1 (χ2 = 13.09, df = 1, P < 0.001) and at Time 2 (χ2 = 8.7, df = 1, P < 0.01). Apart from Time 2 insomnia symptoms, the standard deviation was significantly higher in females compared to males for Time 1 insomnia and depression and Time 2 depression (χ2 = 4.74, 5.91, 6.57, P = 0.02, 0.02, and 0.01, respectively). There were no significant zygosity differences for any of the variables. Comparing twins and siblings, there were no significant differences in means and standard deviations, apart from depression symptoms at Time 2 which showed a lower mean (χ2 = 5.95, df = 1, P < 0.02), and standard deviation (χ2 = 5.95, df = 1, P < 0.02) in siblings. The male-female differences in means are accounted for by specifying a free set of mean parameters for each group (even in the no-sex-difference variance components model).
Correlations
The within-individual cross-trait correlations are presented in Table 2. At both time-points, greater insomnia symptoms were associated with more depression symptoms (Time 1: r = 0.50 [95% confidence intervals, (CI) = 0.46–0.54]; Time 2: r = 0.52 [CI = 0.47–0.56]). There was a substantial degree of stability in the traits across time (insomnia symptoms: r = 0.50 [CI = 0.44– 0.54]; depression symptoms: r = 0.50 [CI = 0.44–0.54]). The cross-trait longitudinal correlations indicated that insomnia symptoms at Time 1 were significantly associated with depression symptoms at Time 2 (r = 0.34 [CI = 0.28–0.39]). Likewise, depression symptoms at Time 1 were significantly associated with insomnia symptoms at Time 2 (r = 0.38 [CI = 0.32–0.44]).
Twin Correlations
The cross-member correlations for MZ and DZ twins and siblings are presented in Table 3. This model is interpreted due to the lack of sex differences detected. For the correlation matrices given by group (zygosity) and sex, please see Tables S1 and S2 in the supplemental material. All MZ and DZ within-trait twin correlations were significant: (range r = 0.31–0.44 and r = 0.15–0.22, respectively). All within-trait sibling correlations were non-significant (range r = −0.02 to 0.14). However, overlapping 95% CI indicate that they were not significantly different from the DZ twin pairs: equating the sibling and DZ correlations resulted in a non-significant drop in fit (χ2 = 17.50, df = 10, P = 0.06), justifying modelling sibling data as DZ twin pairs in the genetic models. The cross-twin cross-trait correlations for insomnia symptoms and depression symptoms at both time-points were, in general, higher in MZ compared to DZ pairs and siblings, suggesting the possibility of genetic effects on the insomnia-depression association. The extent to which this is the case is formally estimated and tested in the genetic model fitting analysis.
Genetic Analyses
The model equating the ACE parameter estimates across males and females (−2LL = 27641.05, df = 4668) did not show a significant decline in fit (χ2 = 12.09, df = 12, P = 0.44) when compared to the quantitative sex-differences model (−2LL = 27628.95, df = 4656). Furthermore, it was possible to drop C (factor loadings and correlations) from the model without decrease in fit (χ2 = 2.04, df = 10, P = 0.99). For simplicity, the more parsimonious AE no-sex-differences model is interpreted here (Table 4), but the full ACE model estimates are provided in Tables S3 and S4 in the supplemental material.
Table 4.
Parameter estimates from best fitting multivariate AE model (95% confidence intervals).

The genetic and non-shared environmental correlations between the variables are presented in the top part of Table 4. For example, the genes influencing insomnia symptoms at Time 1 are highly correlated with those influencing depression symptoms at Time 1 (rA = 0.80, CI = 0.65–0.94). All of the genetic correlation between traits were high (ranging from 0.73 for the genetic correlation between insomnia and depression symptoms at Time 2 to 1.00 for the correlation between insomnia symptoms at Times 1 and 2). The non-shared environmental correlations were smaller (ranging from 0.05 for the association between insomnia symptoms at Time 1 and depression symptoms at Time 2; to 0.40 for the association between insomnia and depression symptoms at Time 2). The lower part of Table 4 shows the standardized estimates for genetic and non-shared environmental influences within each trait (diagonal elements). Additive genetic influences accounted for between 33% to 41% of the variance. The off-diagonal elements represent the proportion of covariance determined by correlating genetic and non-shared environmental factors. Most of the association between variables is due to genetic factors (explaining 50% to 90% of the covariance).
DISCUSSION
Phenotypic Associations
This was the first twin study to examine the longitudinal associations between insomnia and depression symptoms within adulthood. As expected, there were moderate phenotypic associations between insomnia and depression symptoms both within and across time-points, in line with the plethora of literature highlighting links between these phenotypes.1–4
Genetic and Environmental Influences on Associations
When decomposing the concurrent associations between insomnia and depression symptoms into genetic and environmental influences we found a strong genetic correlation between the two phenotypes and a large proportion of the phenotypic correlations could be explained by genes. This is in line with a handful of comparable studies in this area.13,15 Of note, we have previously reported on the concurrent associations between sleep quality and depression and anxiety symptoms at time 1—unsurprisingly producing similar results to the time 1 analyses reported in this current manuscript.14
What is novel here is that when considering the longitudinal associations between the phenotypes, the genetic overlap appeared to be just as strong as for the concurrent associations—and overall, genes appeared to play an even larger role in the association between the phenotypes longitudinally than concurrently.
Persistence of Symptoms over Time
While not a key focus of the current study, these data also provided an understanding of why symptoms of insomnia and depression may persist over time. Again, genetic factors appear to play the largest role—with a perfect correlation between genetic factors influencing insomnia symptoms at Times 1 and 2—suggesting that the same genetic factors may be influencing insomnia symptoms across one's 20s. There was also a high genetic correlation for depression symptoms at Times 1 and 2, and overall, genetic factors appeared to explain roughly three quarters of the persistence between each of the pheno-types over time. These sleep data are largely consistent with previous behavioural genetic work examining the persistence of sleep disturbances and insomnia symptoms over time by highlighting an important role for genetic factors—although previous research also suggests that new genetic influences may also come into play over time.18,32 Similarly, the depression symptom data reported here fits with previous research from our group (including Time 1 data reported here),33 and from others showing that genes account for stability in depression symptoms over time although new genetic influences may also come into play over time.34
Genetic Influences
The general conclusion that genetic factors are important in the persistence and associations between disorders chimes well with the “generalist genes hypothesis” which proposes that genes are important in explaining the overlap between disorders and over time—but that environmental influences are perhaps more important in explaining differences between disorders and across time-points.35
The obvious next step of specifying genes involved in the associations between insomnia and depression symptoms has proven to be much more difficult than expected, with ‘missing heritability’ reported for all complex phenotypes.36 Large scale Genome Wide Association Study (GWAS) collaborations and decreasing costs of next-generation sequencing will help to move forward the currently poor understanding of the genetics of sleep difficulties.37
Environmental Influences
While genetic factors appeared to be key in explaining the associations between insomnia and depression symptoms both concurrently and longitudinally, it is important to note that the non-shared environment also played a role. Univariately, non-shared environmental influences (which include error) accounted for roughly two-thirds of the variance of each phenotype. There were also small to moderate overlaps between the non-shared environmental influences for different bivariate combinations of phenotypes and overall, non-shared environment accounted for a small to moderate proportion of the associations between phenotypes, with the greatest influence for the concurrent associations between phenotypes at each wave. While it was beyond the scope of this study to specify environmental influences involved in the association, candidates come from previous work. For example, important environmental influences might include negative life events, which are plausibly associated with poorer sleep quality38 and depression symptoms.39 Such influences are able to explain both concurrent and longitudinal relationships between variables, given the dynamic nature of the associations between stress and depression over time.39 How these environmental influences “get under the skin” to lead to both insomnia and depression symptoms is an area for further research, and our group has recently used the MZ differences approach to examine the possibility that epigenetic differences can account for extreme differences in phenotypes between twins who are genetically identical (for work focused on depression and diurnal preference see 40,41). Our future work will explore differences in insomnia symptoms between MZ twins.42
Limitations
The results of this study need to be considered alongside the limitations. First, insomnia and depression symptoms were assessed using symptom count measures, which focused on frequency, but not severity of symptoms. We also focused on symptoms rather than diagnoses. This approach is standard when dealing with twin data.43 Furthermore, the convergence of results found here and in previous work using a more clinical approach is reassuring. For example, genetic influence on insomnia symptoms are here estimated at 35% at time 1 and 37% at time 2, which corresponds well with estimates from other twin samples (e.g., 31%13). Furthermore, the estimates for depressive symptoms at time 1 (41%) and time 2 (33%) correspond to a genetic estimate from a meta-analysis of the genetic epidemiology of major depression (37% 44). While research on a plethora of phenotypes suggests that the same influences are likely to account for individual differences in the full range as at the extremes,29 this assumption is yet to be tested for insomnia symptoms.
A second limitation is that we did not systematically assess the multiple factors which could have caused insomnia in our participants (such as circadian sleep disorders and medical conditions). Furthermore, we did not assess the use of drugs prescribed for depression in our sample. It is possible that such drug use could have been important in accounting for some of the association between the variables reported here. For example, sleep problems can be a side-effect of selective serotonin reuptake inhibitors.45 We did assess past-month use of prescribed or over the counter drugs to aid sleep, and found that only 9% of our sample had used such drugs at Time 2. When we re-ran phenotypic analyses excluding those who had taken these drugs at Time 2, we found very similar results.
A third limitation concerns the sample which was relatively small for a twin study due to inevitable attrition across the waves of data collection. Nonetheless, the sample size reported here is comparable with other twin studies of sleep difficul-ties.18 While the data need to be interpreted considering the large confidence intervals, it was reassuring that results reported here are in line with comparable studies using larger sample sizes. Furthermore, there was no selective attrition for the key variables assessed here, namely insomnia and depression symptoms. However, those who stayed in the study from Time 1 to Time 2 were more likely than those who dropped out to be female and have more highly educated mothers. This is noteworthy given that the G1219 study as a whole initially comprised families including parents who are more educated and more likely to own their own homes than representative samples.19 Heritability is a population statistic, meaning that our results may not apply to other cohorts. This needs to be considered when interpreting the results of this study.
Finally, standard twin limitations need to be considered.29 Years of careful consideration of these limitations leads to the realisation that different limitations influence genetic and environmental estimates unsystematically and that estimates should be interpreted generally.
CONCLUSIONS
The overall conclusion that genes are important in the association between insomnia and depression symptoms goes to reinforce the general conclusion that genes are important for all complex traits,46 and therefore need to be acknowledged when developing theories describing the mechanisms underlying phenotypes and their associations. Genome-wide approaches to these questions will be the next step,47 as we consider whether genetic markers across the genome associated with, for example insomnia, also influence major depressive disorders. This approach is highly complementary to the one used here, and has the added benefit of producing data that applies to an individual, not just population variance. However, it also requires huge sample sizes so progress is inevitably somewhat slow. Another way to explore this question further is to see whether reducing one difficulty (for example using a psychological therapy for insomnia) influences the other (e.g., depression), and examining the role of genetic effects on such changes. We have been studying genetic influences on outcome following psychological therapy in anxiety disorders. As with any area of psychiatric genetics, the work requires large sample sizes and most studies to date have been underpowered (e.g., work on pediatric anxiety48). However there are preliminary indications that there are genetic influences on response to psychological treatment,49 indicating that this is a fruitful avenue for further research. Future work by our group will examine genetic prediction of response to CBT for insomnia symptoms and whether improving those symptoms has positive effects on depression symptoms.
DISCLOSURE STATEMENT
This was not an industry supported study. Waves 1–3 funded by the W T Grant Foundation, the University of London Central Research fund and a Medical Research Council Training Fellowship (G81/343) and Career Development Award to Thalia C. Eley. Wave 4 supported by the Economic and Social Research Council (RES-000-22-2206) and the Institute of Social Psychiatry (06/07–11) to Alice M. Gregory. Wave 5 supported by funding from Goldsmiths, University of London to Alice M. Gregory. The authors have indicated no financial conflicts of interest. The work was performed at Goldsmiths, University of London and the Institute of Psychiatry, Psychology & Neuroscience, King's College London.
ACKNOWLEDGMENTS
The authors thank the families for their participation as well as numerous staff and students from the Social Genetic Developmental Psychiatry Centre, Institute of Psychiatry, Psychology & Neuroscience, King's College London and Goldsmiths, University of London. The authors thank Monika Waszczuk for her input and help with Figure 1. Authorship responsibilities: TCE is the founder of the G1219 study. AMG ran waves 4 and 5 of the G1219 study. FR and NB ran the analyses for this study. All authors contributed to the drafting of this manuscript.
REFERENCES
- 1.Alvaro PK, Roberts RM, Harris JK. A systematic review assessing bidirectionality between sleep disturbances, anxiety, and depression. Sleep. 2013;36:1059–68. doi: 10.5665/sleep.2810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Baglioni C, Battagliese G, Feige B, et al. Insomnia as a predictor of depression: a meta-analytic evaluation of longitudinal epidemiological studies. J Affect Disord. 2011;135:10–9. doi: 10.1016/j.jad.2011.01.011. [DOI] [PubMed] [Google Scholar]
- 3.Lovato N, Gradisar M. A meta-analysis and model of the relationship between sleep and depression in adolescents: recommendations for future research and clinical practice. Sleep Med Rev. 2014;18:521–9. doi: 10.1016/j.smrv.2014.03.006. [DOI] [PubMed] [Google Scholar]
- 4.Staner L. Comorbidity of insomnia and depression. Sleep Med Rev. 2010;14:35–46. doi: 10.1016/j.smrv.2009.09.003. [DOI] [PubMed] [Google Scholar]
- 5.Baglioni C, Spiegelhalder K, Lombardo C, Riemann D. Sleep and emotions: a focus on insomnia. Sleep Med Rev. 2010;14:227–38. doi: 10.1016/j.smrv.2009.10.007. [DOI] [PubMed] [Google Scholar]
- 6.Prather AA, Bogdan R, Hariri AR. Impact of sleep quality on amygdala reactivity, negative affect, and perceived stress. Psychosom Med. 2013;75:350–8. doi: 10.1097/PSY.0b013e31828ef15b. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Yoo SS, Gujar N, Hu P, Jolesz FA, Walker MP. The human emotional brain without sleep - a prefrontal amygdala disconnect. Curr Biol. 2007;17:R877–8. doi: 10.1016/j.cub.2007.08.007. [DOI] [PubMed] [Google Scholar]
- 8.Muzur A, Pace-Schott EF, Hobson JA. The prefrontal cortex in sleep. Trends Cogn Sci. 2002;6:475–81. doi: 10.1016/s1364-6613(02)01992-7. [DOI] [PubMed] [Google Scholar]
- 9.Hariri AR. Sunderland, MA: Sinauer; 2015. Looking inside the disordered brain. [Google Scholar]
- 10.Riemann D, Spiegelhalder K, Nissen C, Hirscher V, Baglioni C, Feige B. REM sleep instability - a new pathway for insomnia? Pharmacopsychiatry. 2012;45:167–76. doi: 10.1055/s-0031-1299721. [DOI] [PubMed] [Google Scholar]
- 11.Barclay NL, Gregory AM. Quantitative genetic research on sleep: a review of normal sleep, sleep disturbances and associated emotional, behavioural, and health-related difficulties. Sleep Med Rev. 2013;17:29–40. doi: 10.1016/j.smrv.2012.01.008. [DOI] [PubMed] [Google Scholar]
- 12.Gregory AM, Rijsdijk FV, Dahl RE, McGuffin P, Eley TC. Associations between sleep problems, anxiety and depression in twins at 8 years of age. Pediatrics. 2006;118:1124–32. doi: 10.1542/peds.2005-3118. [DOI] [PubMed] [Google Scholar]
- 13.Gehrman PR, Meltzer LJ, Moore M, et al. Heritability of insomnia symptoms in youth and their relationship to depression and anxiety. Sleep. 2011;34:1641–6. doi: 10.5665/sleep.1424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Gregory AM, Buysse DJ, Willis TA, et al. Associations between sleep quality and anxiety and depression symptoms in a sample of young adult twins and siblings. J Psychosom Res. 2011;71:250–5. doi: 10.1016/j.jpsychores.2011.03.011. [DOI] [PubMed] [Google Scholar]
- 15.Hur YM, Burri A, Spector TD. The genetic and environmental structure of the covariation among the symptoms of insomnia, fatigue, and depression in adult females. Twin Res Human Gen. 2012;15:720–6. doi: 10.1017/thg.2012.60. [DOI] [PubMed] [Google Scholar]
- 16.Middeldorp CM, Cath DC, van Dyck R, Boomsma DI. The comorbidity of anxiety and depression in the perspective of genetic epidemiology. A review of twin and family studies. Psychol Med. 2005;35:611–24. doi: 10.1017/s003329170400412x. [DOI] [PubMed] [Google Scholar]
- 17.Gillespie NA, Gehrman P, Byrne EM, Kendler KS, Heath AC, Martin NG. Modeling the direction of causation between cross-sectional measures of disrupted sleep, anxiety and depression in a sample of male and female Australian twins. J Sleep Res. 2012;21:675–83. doi: 10.1111/j.1365-2869.2012.01026.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Gregory AM, Rijsdijk FV, Lau JF, Dahl RE, Eley TC. The direction of longitudinal associations between sleep problems and depression symptoms: a study of twins aged 8 and 10 years. Sleep. 2009;32:189–99. doi: 10.1093/sleep/32.2.189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.McAdams TA, Gregory AM, Rowe R, et al. The genesis 12-19 (G1219) study: a twin and sibling study of gene-environment interplay and adolescent development in the UK. Twin Res Human Gen. 2013;16:134–43. doi: 10.1017/thg.2012.83. [DOI] [PubMed] [Google Scholar]
- 20.Cohen DJ, Dibble E, Grawe JM, Pollin W. Reliably separating identical from fraternal twins. Arch Gen Psychiatry. 1975;32:1371–5. doi: 10.1001/archpsyc.1975.01760290039004. [DOI] [PubMed] [Google Scholar]
- 21.Portaluppi F, Smolensky MH, Touitou Y. Ethics and methods for biological rhythm research on animals and human beings. Chronobiol Int. 2010;27:1911–29. doi: 10.3109/07420528.2010.516381. [DOI] [PubMed] [Google Scholar]
- 22.Okun ML, Kravitz HM, Sowers MF, Moul DE, Buysse DJ, Hall M. Psychometric evaluation of the Insomnia Symptoms Questionnaire: a self-report measure to identify chronic insomnia. J Clin Sleep Med. 2009;5:41–51. [PMC free article] [PubMed] [Google Scholar]
- 23.Chorpita BF, Yim L, Moffitt C, Umemoto LA, Francis SE. Assessment of symptoms of DSM-IV anxiety and depression in children: a revised child anxiety and depression scale. Behav Res Ther. 2000;38:835–55. doi: 10.1016/s0005-7967(99)00130-8. [DOI] [PubMed] [Google Scholar]
- 24.Costello EJ, Angold A. Scales to assess child and adolescent depression: checklists, screens, and nets. J Am Acad Child Adolesc Psychiatry. 1988;27:726–37. doi: 10.1097/00004583-198811000-00011. [DOI] [PubMed] [Google Scholar]
- 25.Costello EJ, Benjamin R, Angold A, Silver D. Mood variability in adolescents - a study of depressed, nondepressed and comorbid patients. J Affect Disord. 1991;23:199–212. doi: 10.1016/0165-0327(91)90101-w. [DOI] [PubMed] [Google Scholar]
- 26.Turner N, Joinson C, Peters TJ, Wiles N, Lewis G. Validity of the Short Mood and Feelings Questionnaire in late adolescence. Psychol Assessment. 2014;26:752–62. doi: 10.1037/a0036572. [DOI] [PubMed] [Google Scholar]
- 27.Finsterer J, Mahjoub SZ. Fatigue in healthy and diseased individuals. Am J Hosp Palliat Med. 2014;31:562–75. doi: 10.1177/1049909113494748. [DOI] [PubMed] [Google Scholar]
- 28.McGue M, Bouchard TJ., Jr Adjustment of twin data for the effects of age and sex. Behav Genet. 1984;14:325–43. doi: 10.1007/BF01080045. [DOI] [PubMed] [Google Scholar]
- 29.Plomin R, DeFries JC, Knopik VS, Neiderhiser JM. Behavioral Genetics. 6th ed. New York, NY: Worth Publishers; 2013. [Google Scholar]
- 30.Neale MC, Roysamb E, Jacobson K. Multivariate genetic analysis of sex limitation and G x E interaction. Twin Res Human Gen. 2006;9:481–9. doi: 10.1375/183242706778024937. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Boker SM, Neale MC, Maes HH, et al. OpenMx: an open source extended structural equation modelling framework. Psychometrika. 2011;76:306–17. doi: 10.1007/s11336-010-9200-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Barclay NL, Gehrman PR, Gregory AM, Eaves LJ, Silberg JL. The heritability of insomnia progression during childhood/adolescence: results from a longitudinal twin study. Sleep. 2015;38:109–18. doi: 10.5665/sleep.4334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Waszczuk MA, Zavos HMS, Gregory AM, Eley TC. The stability and change of etiological influences on depression, anxiety symptoms and their co-occurrence across adolescence and young adulthood. Psychol Med. 2016;46:161–75. doi: 10.1017/S0033291715001634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Gillespie NA, Kirk KM, Evans DM, Heath AC, Hickie IB, Martin NG. Do the genetic or environmental determinants of anxiety and depression change with age? A longitudinal study of Australian twins. Twin Res. 2004;7:39–53. doi: 10.1375/13690520460741435. [DOI] [PubMed] [Google Scholar]
- 35.Eley TC. General genes: a new theme in developmental psychopathology. Curr Directions in Psych Sci. 1997;6:90–5. [Google Scholar]
- 36.Manolio TA, Collins FS, Cox NJ, et al. Finding the missing heritability of complex diseases. Nature. 2009;461:747–53. doi: 10.1038/nature08494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Parsons MJ. On the genetics of sleep disorders: genome-wide association studies and beyond. Adv Genom Genetics. 2015;5:293–303. [Google Scholar]
- 38.Barclay NL, Eley TC, Rijsdijk FV, Gregory AM. Dependent negative life events and sleep quality: an examination of gene-environment interplay. Sleep Med. 2011;12:403–9. doi: 10.1016/j.sleep.2010.09.009. [DOI] [PubMed] [Google Scholar]
- 39.Hammen C. Stress and depression. Ann Rev Clin Psychol. 2005;1:293–319. doi: 10.1146/annurev.clinpsy.1.102803.143938. [DOI] [PubMed] [Google Scholar]
- 40.Dempster EL, Wong CC, Lester KJ, et al. Genome-wide methylomic analysis of monozygotic twins discordant for adolescent depression. Biol Psychiatry. 2014;76:977–83. doi: 10.1016/j.biopsych.2014.04.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Wong CCY, Parsons MJ, Lester KJ, et al. Epigenome-wide DNA methylation analysis of monozygotic twins discordant for diurnal preference. Twin Res Human Gen. 2015;18:662–9. doi: 10.1017/thg.2015.78. [DOI] [PubMed] [Google Scholar]
- 42.Palagini L, Biber K, Riemann D. The genetics of insomnia - Evidence for epigenetic mechanisms? Sleep Med Rev. 2014;18:225–35. doi: 10.1016/j.smrv.2013.05.002. [DOI] [PubMed] [Google Scholar]
- 43.Taylor MJ, Gregory AM, Freeman D, Ronald A. Do sleep disturbances and psychotic experiences in adolescence share genetic and environmental influences? J Abnorm Psychol. 2015;124:674–84. doi: 10.1037/abn0000057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Sullivan PF, Neale MC, Kendler KS. Genetic epidemiology of major depression: review and meta-analysis. Am J Psychiatry. 2000;157:1552–62. doi: 10.1176/appi.ajp.157.10.1552. [DOI] [PubMed] [Google Scholar]
- 45.Wilson S, Argyropoulos S. Antidepressants and sleep - A qualitative review of the literature. Drugs. 2005;65:927–47. doi: 10.2165/00003495-200565070-00003. [DOI] [PubMed] [Google Scholar]
- 46.Polderman TJ, Benyamin B, de Leeuw CA, et al. Meta-analysis of the heritability of human traits based on fifty years of twin studies. Nat Genet. 2015;47:702–9. doi: 10.1038/ng.3285. [DOI] [PubMed] [Google Scholar]
- 47.Wray NR, Maier R. Genetic basis of complex genetic disease: the contribution of disease heterogeneity to missing heritability. Curr Epidemiol Rep. 2014;1:220–7. [Google Scholar]
- 48.Lester KJ, Roberts S, Keers R, et al. Non-replication of the association between 5HTTLPR and response to psychological therapy for child anxiety disorders. Br J Psychiatry. 2016;208:182–8. doi: 10.1192/bjp.bp.114.154997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Coleman JR, Lester KL, Keers R, et al. Genome-wide association study of response to cognitive behavioural therapy in children with anxiety disorders. Br J Psychiatry. 2016 Mar 17; doi: 10.1192/bjp.bp.115.168229. [Epub ahead of print] [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.