

Abstract
Natural products can be a source of biomolecules with antioxidant activity which are able to prevent oxidative stress-induced diseases and show antitumor activity, making them important sources of new anticancer drug prototypes. In this context, this study aimed to analyze the chemical composition of an ethanol extract of Senna velutina leaves and to assess its antioxidant and cytotoxic activities in leukemic cells. The antioxidant properties were evaluated using a DPPH free radical scavenging assay and by examining the extract's inhibition of AAPH-induced lipid peroxidation in human erythrocytes. Its cytotoxicity and possible mechanisms of action were assessed in Jurkat and K562 leukemic cell lines. The ethanol extract contained flavonoids, such as epigallocatechin, epicatechin, kaempferol heteroside, rutin, and dimeric and trimeric proanthocyanidin derivatives. The extract exhibited antioxidant activity by scavenging free radicals and antihemolytic action, and it decreased malondialdehyde content in human erythrocytes. Furthermore, the extract also induced leukemic cell death by activating intracellular calcium and caspase-3, decreasing mitochondrial membrane potential, and arresting the cell cycle in S and G2 phases. Hence, S. velutina leaf extract contains antioxidant and antileukemic biomolecules with potential applications in diseases associated with oxidative stress and in the inhibition of tumor cell proliferation.
1. Introduction
Several diseases, including cancer, diabetes, atherosclerosis, inflammatory diseases, and premature aging, are related to oxidative stress [1]. Oxidative stress stems from excess of free radicals in the body and low antioxidant activity, resulting in damage to essential biomolecules such as nucleic acids, proteins, and lipids [2].
Cancer is an oxidative stress-related disease that causes high rates of morbidity and mortality in the global population [3]. Leukemias are cancers that affect the cells of the hematopoietic system; depending on their cellular origin and maturity stage, leukemias can be classified as either myeloid or lymphoid and as acute or chronic [4]. Surgery, radiotherapy, and chemotherapy [5] are among the main types of treatment for these cancers.
Biomolecules with anticancer activity at low therapeutic doses and with reduced side effects have been increasingly sought in recent decades [6]. Between 1940 and 2014, 49% of the 174 anticancer drugs that were made available on the market were either natural products or their derivatives [7]. Thus, there is a trend among the general population and the medical community to regard medicinal plants as alternative sources of antitumor drugs, provided that the therapeutic properties of such plants have been scientifically researched and proven [8]. The discoveries of paclitaxel [9], an anticancer drug, and of homoharringtonine [10], which is used in the treatment of acute and chronic myeloid leukemia, are examples of successful cases in the development of medicinal plant-derived drugs.
The search for new molecules with therapeutic properties, including antioxidant and anticarcinogenic activities, is facilitated by the vast biodiversity and bioprospecting potential in Brazil. Senna genus has been used in Brazilian folk medicine for its antioxidant, antimicrobial [11], anti-inflammatory [12], antidiabetic [13], and antitumor [14] activities, among other uses.
Taxonomically, some species have been transferred from Cassia genus to Senna genus [15]. This taxon currently comprises 500–600 species [14, 16], of which many have not been yet characterized with respect to their chemical compositions and biological properties as the arboreal species Senna velutina (Vogel) H. S. Irwin & Barneby (Fabaceae, Caesalpinioideae). In this context, the aim of the present study was to determine the chemical composition of an ethanol extract of S. velutina leaves and to evaluate its antioxidant and antileukemic activities.
2. Materials and Methods
2.1. Plant Material and Extract Preparation
S. velutina leaves were collected following the identification of the plant and authorization of the SISBIO (Sistema de Autorização e Informação em Biodiversidade, permit number 54470-1) in Dourados, Mato Grosso do Sul (S 22°05′545′′ and W 055°20′746′′), Brazil, oven-dried with the air circulation at a temperature of 45 ± 5°C, and then ground in a Willy-type knife mill. An exsiccated sample was deposited in the Herbarium of the Federal University of Grande Dourados, Mato Grosso do Sul, Brazil, with registration number 4665.
The extract was then prepared by macerating the plant material in an ethanol 95% mixture at room temperature in the dark for 7 days. Then, the extract was filtered, and the residue was further extracted twice using the same process. After 21 days, the filtrate was concentrated in a rotary vacuum evaporator (Gehaka, São Paulo, SP, Brazil) to obtain the ethanol extract of S. velutina leaves (ESV). The dry extract yield was 29%, calculated using the following formula: extraction yield (%) = (weight of the freeze-dried extract × 100)/(weight of the original sample). The ESV was stored at −20°C protected from light.
2.2. Chemical Analysis
The extract was analyzed by Ultra Fast Liquid Chromatography (UFLC) (Shimadzu) coupled to Diode Array Detector (DAD) (240–800 nm, Shimadzu) and electrospray ionization time-of-flight (ESI-QTOF-micrOTOF QII) (operating in positive and negative mode, 120–1200 Da, Bruker Daltonics). A C-18 column was used (Kinetex, 2.6 μm, 150 × 2.2 mm, Phenomenex) protected by a guard column of the same material. The mobile phase was as follows: water (solvent A) and acetonitrile (solvent B) both with 0.1% of formic acid in a gradient of 0–2 min 3% B, 2–25 min 3–25% B, and 25–40 min 25–80% B followed by washing and reconditioning of the column (8 minutes). The flow rate was 0.3 mL/min and 1 μL (1 mg/mL) of extract was injected. The other micrOTOF-QII parameters were as follows: temperature, 200°C; N2 drying gas flow rate, 9 L/min; Nebulizer, 4.0 bar; capillary voltage, −3500 V (negative) and +4500 V (positive); and internal calibration with TFA-NA injected at the end of the chromatographic analysis. The rutin and epicatechin standards were obtained from Sigma-Aldrich with a purity of ≥95%.
2.3. Antioxidant Activity
2.3.1. DPPH Free Radical Scavenging Activity
The 2,2-diphenyl-1-picrylhydrazyl (DPPH) radical scavenging activity of ESV was evaluated as described in D. Gupta and R. K. Gupta [17], with modifications. In this assay, 0.2 mL of ESV at different concentrations (1–1000 μg/mL) was added to 1.8 mL of DPPH solution (0.11 mM) in 80% ethanol. The mixture was incubated for 30 minutes at room temperature in the dark. Absorbance at 517 nm was then measured spectrophotometrically. Ascorbic acid and butylhydroxytoluene (BHT) were used as reference antioxidants. As a control, 0.2 mL of solvent used to dilute the extract was added to 1.8 mL of DPPH solution (0.11 mM) in 80% ethanol. Two independent experiments were performed in triplicate. The percentage inhibition was calculated relative to the control using the following equation:
| (1) |
2.3.2. Inhibition of Lipid Peroxidation in Human Erythrocytes
(1) Preparation of Erythrocyte Suspension. After approval by the Research Ethics Committee (Comitê de Ética em Pesquisa, CEP) of the University Center of Grande Dourados (UNIGRAN, Brazil (CEP process number: 123/12)), peripheral blood from healthy donors was collected into tubes containing sodium citrate which were then centrifuged at 400 ×g for 10 min. The plasma and leukocyte layer were discarded, and the erythrocytes were washed 3 times with 0.9% sodium chloride solution (NaCl) and centrifuged. Finally, 10% erythrocyte suspension was prepared in 0.9% NaCl solution to attain a 2.5% final concentration for further analysis.
(2) Hemolytic Activity and Inhibition of Oxidative Hemolysis. The ability of ESV to protect against lipid peroxidation was evaluated using an antihemolytic assay in human erythrocytes that were incubated with the oxidant 2,2′-azobis-(2-amidinopropane) dihydrochloride (AAPH), a thermoinducible initiator of lipid peroxidation, as described by Campos et al. [18]. To evaluate hemolytic activity and inhibition of oxidative hemolysis, erythrocytes were preincubated with either ESV or ascorbic acid (25–125 μg/mL) at 37°C for 30 min before the addition of 0.5 mL of either 0.9% NaCl or 50 mM AAPH (dissolved in 0.9% NaCl). In the assay, 1% ethanol was used as a solvent control, while 0.9% NaCl was used as the baseline hemolysis control. Total hemolysis was induced by incubating erythrocytes with distilled water. Samples were incubated at 37°C with regular stirring. The extent of hemolysis was determined at 60-minute intervals for an incubation period of 5 h. Tubes were centrifuged at 700 ×g for 5 minutes; 0.2 mL of the supernatant was collected and added to 1.8 mL of 0.9% NaCl for spectrophotometric reading at 540 nm. The hemolysis percentage was calculated using the formula A/B × 100, where A is the sample absorbance and B is the absorbance of the total hemolysis sample. Three independent experiments were performed in triplicate.
(3) Malondialdehyde (MDA) Measurements. The inhibition of malondialdehyde (MDA) production, which is a byproduct of lipid peroxidation, was evaluated according to the method described by Campos et al. [18]. Erythrocytes were preincubated at 37°C for 30 min with either ESV or ascorbic acid (25–125 μg/mL) before the addition of 0.5 mL of 50 mM AAPH solution. The mixtures were incubated at 37°C with regular stirring; 1% ethanol was used as a negative control. The MDA concentration was determined at 60-minute intervals for a total of 5 h. To determine the MDA concentration, samples were centrifuged at 700 ×g for 5 min; 0.5 mL of each supernatant was collected and transferred to a tube containing 1 mL of 10 nM thiobarbituric acid (TBA), dissolved in 75 mM monobasic potassium phosphate buffer at pH 2.5. The standard controls used were 500 μL of a 20 mM MDA solution and 1 mL of TBA. Samples were incubated at 96°C for 45 min and allowed to cool before adding 4 mL of n-butyl alcohol and centrifuging at 1600 ×g for 5 min. The resulting supernatant was read at 532 nm in a spectrophotometer. Two independent experiments were performed in triplicate. Sample MDA levels were expressed in nM/mL, according to the following formula:
| (2) |
2.4. Cytotoxic Activity
2.4.1. Cell Culture
Leukemia human cell lines Jurkat and K562 were cultivated in RPMI 1640 (Sigma-Aldrich, Germany) culture medium, supplemented with 10% fetal bovine serum, 100 U/mL penicillin (Sigma-Aldrich, Germany), and 100 μg/mL streptomycin (Sigma-Aldrich, Germany). Cells were cultured in a humidified incubator containing 5% CO2 at 37°C.
2.4.2. Cytotoxicity and Cell Death Profile
Cytotoxic activity and the cell death profile were evaluated according to the method described by Paredes-Gamero et al. [19]. Leukemic cells were seeded at 105 cells/mL in 96-well microplates and treated with ESV (0–100 μg/mL) for 24 h. Then, the cells were centrifuged 600 ×g for 6 min and resuspended in binding buffer (0.14 M NaCl, 2.5 mM CaCl2, 0.01 M HEPES, and pH 7.4) and incubated at room temperature with 1 μL of Annexin V-FITC (BD Biosciences, San Diego, CA, USA) and 5 μg/mL propidium iodide (PI) Becton Dickinson, USA for 20 min. Sample analysis was performed using Accuri C6 flow cytometer (Becton Dickinson, San Diego, CA, USA), with acquisition of 3,000 events.
2.4.3. Measurement of Mitochondrial Membrane Potential
To evaluate the possible effects of ESV on mitochondrial membrane potential, leukemic cells were incubated with the fluorescent marker JC-1 (5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethylbenzimidazolylcarbocyanine iodide; Molecular Probes, Eugene, OR, USA) according to the method described by Moraes et al. [20]. JC-1 probe accumulates in mitochondria in a potential-dependent manner. Viable cells with high mitochondrial membrane potential are stained red. Upon reduction of the mitochondrial membrane potential, cells appear green. In this assay, cells were seeded into 24-well plates (105 cells/mL) containing supplemented media and were then incubated with 27.6 μg/mL (Jurkat cells) or 67.5 μg/mL (K562 cells) ESV for 24 h. The cells were then centrifuged and incubated with JC-1 (1 μg/mL) for 15 min at room temperature. Fluorescence readings were performed in a FACSCalibur flow cytometer using CellQuest software (Becton Dickinson, San Diego, CA, USA). A total of 10,000 events were collected per sample.
2.4.4. Caspase-3 Activity
Caspase-3 activity was assessed according to the method described by Moraes et al. [20], with minor modifications. Caspase activity was measured by flow cytometer. Leukemic lineages were treated with ESV (27.6 μg/mL) in 24-well microplates (105 cells/mL) for 24 h. Then the cells were fixed with 2% paraformaldehyde in PBS for 30 min and permeabilized with 0.01% saponin for 15 min at room temperature. Next, the cells were incubated for 1 h at 37°C with anti-cleaved-caspase 3-FITC antibody (Becton Dickinson, USA). After incubation for 40 min, the fluorescence was analyzed by Accuri C6 flow cytometer (Becton Dickinson, USA). A total of 10,000 events were acquired.
2.4.5. Intracellular Calcium and Pan-Caspase Inhibitors
The roles of intracellular calcium and caspases in ESV-promoted cytotoxicity were evaluated according to Bechara et al. [21], with minor modifications. Jurkat cells were pretreated for 1 h at 37°C under a 5% CO2 atmosphere with either the intracelullar calcium chelator BAPTA-AM or the pan-caspase inhibitor Z-VAD-FMK; ESV (27.6 μg/mL) was then added to the cells and allowed to incubate for 24 h. Control and treated cells were resuspended in culture medium containing 0.05% trypan blue and were counted in a hemocytometer chamber to determine cell viability (trypan blue exclusion assay).
2.4.6. Cell Cycle Phases
Distribution of cell cycle was determined by PI staining and flow cytometry analysis. Leukemic lineages (105 cells/mL) were treated with ESV (27.6 μg/mL) for 24 h and then were fixed and permeabilized as previously described and treated with 4 mg/mL RNase (Sigma-Aldrich, Germany) for 45 min at 37°C. For DNA labeling, cells were incubated with 5 μg/mL of PI (Sigma Aldrich, Germany). Percentages of cells within cell cycle compartments (G1, S, and G2/M) were determined by Accuri C6 flow cytometer (Becton Dickinson, USA). A total of 10,000 events were acquired.
2.5. Statistical Analyses
The data are shown as the mean ± standard error of the mean (SEM) and were analyzed for statistical significant differences between the groups using Student's t-test for comparison between two groups and one-way analysis of variance (ANOVA) followed by Dunnett's test for comparison of more than two groups using Prism 5 GraphPad Software. The results were considered significant when P < 0.05.
3. Results
3.1. Chemical Composition of ESV
The metabolites in ESV were identified by interpreting the UV absorption and mass spectra and comparing them with data in the literature. When available, compounds were compared with authentic standards for confirmation.
The identification of compound 2, with m/z 305.0660 [M-H]−, was based on the fragmentation pattern of epigallocatechin proposed by Dou et al. [22], namely, m/z 261 [M-CO2]−, m/z 219 [M-C4H6O2]−, m/z 179 [M-C6H6O3]−, m/z 167 [M-C7H6O3]−, and m/z 165 [M-C7H8O3]−, and also on the detection of UV absorption at 270 nm.
Compounds 4, 8, and 9 showed UV absorption patterns that were characteristic of flavonols (270 and 340 nm). Their fragmentation patterns were consistent with those of heteroside derivatives of kaempferol, whose main fragment consists of m/z 285 [C15H9O6]−. Several compounds of this class have been described in Senna genus [23, 24].
Dimeric (compounds 6, 7, and 10–14) and trimeric (compounds 15–22) proanthocyanidins, comprising cassiaflavan, afzelechin, epicatechin, epigallocatechin, and naringenin subunits, were also observed. These derivatives, although rare in nature, are often reported in Senna genus; some authors consider them to be chemical markers for the genus [25–27]. The UV absorption maxima at 280 nm and the increase in reverse-phase retention time concomitant with decreased hydroxyl group content or increased degree of polymerization are in agreement with the study by Callemien and Collin [28]. Fragmentation patterns obtained by retro-Diels-Alder (RDA) fission, heterocyclic ring fission (HRF), and quinone methide (QM) can be used to characterize the subunits that make up proanthocyanidins [29]. A complete discussion on the elucidation of this class of compounds may be found in the literature [29, 30]. Thus, based on retention times, fragmentation profiles, and comparisons with previously published data, 22 compounds in ESV were characterized (Figure 1 and Table 1).
Figure 1.

UFLC-DAD-ESI-QTOF-micrOTOF QII chemical profiling (negative mode) of an ethanol extract of S. velutina leaves. (a) Base peak chromatograms (BPC). (b) Extract ion chromatogram (EIC) of m/z 305. (c) EIC of m/z 289. (d) EIC of m/z 593. (e) EIC of m/z 609. (f) EIC of m/z 545. (g) EIC of m/z 529. (h) EIC of m/z 513. (i) EIC of m/z 785. (j) EIC of m/z 769.
Table 1.
Compounds identified in ESV by UFLC-DAD-ESI-QTOF-micrOTOF QII.
| ID | Time (min) | UV | [M-H]−
(m/z) |
Molecular formula | Error (ppm) | MS/MS | Compound |
|---|---|---|---|---|---|---|---|
| 1 | 1.1 | — | 341.1086 | C12H20O11 | 0.6 | 341: 179 | Sugar derivative |
| 2 | 8.6 | 270 | 305.0660 | C15H14O7 | 2.3 | 305: 261, 219, 179, 167, 165 | Epigallocatechin |
| 3 | 12.5 | 280 | 289.0714 | C15H14O6 | 1.3 | 289: 245, 205, 203 | Epicatechin |
| 4 | 19.1 | 270/346 | 593.1524 | C27H30O15 | 2.1 | 593: 447, 285 | Kaempferol-O-hexoside-deoxyhexoside |
| 5 | 19.7 | 270/346 | 609.1450 | C27H30O16 | 1.9 | 609: 463, 301 | Rutin |
| 6 | 20.5 | 280 | 545.1440 | C30H26O10 | 2.5 | 545: 305, 239, 219, 167, 165 | Cassiaflavan-epigallocatechin |
| 7 | 20.8 | 280 | 545.1440 | C30H26O10 | 2.5 | 545: 305, 239, 219, 167, 165 | Cassiaflavan-epigallocatechin |
| 8 | 21.2 | 268/338 | 593.1522 | C27H30O15 | 1.6 | 593: 447, 285 | Kaempferol-O-hexoside-deoxyhexoside |
| 9 | 22.3 | 270/342 | 593.1521 | C27H30O15 | 1.5 | 593: 447, 285 | Kaempferol-O-hexoside-deoxyhexoside |
| 10 | 22.6 | 280 | 529.1489 | C30H26O9 | 2.9 | 529: 289, 245, 239, 203 | Cassiaflavan-epicatechin |
| 11 | 23.5 | 280 | 529.1484 | C30H26O9 | 3.7 | 529: 289, 245, 239, 203 | Cassiaflavan-epicatechin |
| 12 | 24.9 | 280 | 529.1489 | C30H26O9 | 2.9 | 529: 267, 257, 239, 151 | Naringenin-afzelechin |
| 13 | 28.3 | 280 | 513.1551 | C30H26O8 | 0.8 | 513: 267, 255, 239 | Cassiaflavan-afzelechin |
| 14 | 28.8 | 280 | 513.1541 | C30H26O8 | 2.8 | 513: 267, 255, 239 | Cassiaflavan-afzelechin |
| 15 | 30.2 | 280 | 785.2266 | C45H38O13 | 3.4 | 785: 435, 305, 239 | Cassiaflavan-cassiaflavan-epigallocatechin |
| 16 | 30.4 | 280 | 785.2285 | C45H38O13 | 1.7 | 785: 435, 305, 239 | Cassiaflavan-cassiaflavan-epigallocatechin |
| 17 | 30.5 | 280 | 785.2255 | C45H38O13 | 2.0 | 785: 435, 305, 239 | Cassiaflavan-cassiaflavan-epigallocatechin |
| 18 | 31.1 | 280 | 769.2310 | C45H38O12 | 2.6 | 769: 529, 419, 289 | Cassiaflavan-cassiaflavan-epicatechin |
| 19 | 31.3 | 280 | 769.2303 | C45H38O12 | 1.6 | 769: 529, 419, 289 | Cassiaflavan-cassiaflavan-epicatechin |
| 20 | 31.5 | 280 | 769.2295 | C45H38O12 | 0.6 | 769: 377, 267, 239 | Cassiaflavan-naringenin-afzelechin |
| 21 | 31.7 | 280 | 769.2310 | C45H38O12 | 2.6 | 769: 377, 267, 239 | Cassiaflavan-naringenin-afzelechin |
| 22 | 31.8 | 280 | 769.2297 | C45H38O12 | 0.9 | 769: 377, 267, 239 | Cassiaflavan-naringenin-afzelechin |
3.2. DPPH Free Radical Scavenging Activity
ESV was able to scavenge the DPPH free radical, with a 2.5-fold higher IC50 and maximum activity values relative to ascorbic acid; however, these values were lower than those obtained with BHT (Table 2).
Table 2.
IC50 and maximal DPPH radical scavenging activity of standard antioxidants and of ESV.
| Sample | IC50 (µg/mL) | Maximal inhibition | |
|---|---|---|---|
| % | µg/mL | ||
| Ascorbic acid | 2.6 ± 0.8 | 90.9 ± 1.6 | 10 |
| BHT | 21.3 ± 1.2 | 92.4 ± 1.2 | 250 |
| ESV | 6.3 ± 1.3 | 92.4 ± 0.4 | 25 |
Values are means ± SEM.
3.3. Hemolytic Activity and Inhibition of Oxidative Hemolysis
Over the range of tested concentrations, ESV showed no hemolytic activity in human erythrocytes, as no hemolysis was observed after up to 5 h of incubation (Figure 2(a)).
Figure 2.

Hemolysis and MDA content in human erythrocytes incubated for 5 hours with ascorbic acid (AA) and ESV (50–125 μg/mL). (a) Hemolytic activity of ESV in the absence of AAPH. (b) Antihemolytic activity after addition of AAPH. (c) Malondialdehyde (MDA) concentration (nM/mL) after addition of the oxidizing agent. ∗ P < 0.05 compared to the AAPH-only control (erythrocytes incubated with oxidant only).
The control antioxidant, ascorbic acid, was able to protect erythrocytes from hemolysis for up to 4 h of incubation when they were exposed to the oxidant AAPH (data not shown). ESV was able to protect erythrocytes for 5 h over the tested 50–125 μg/mL concentration range (Figure 2(b)), demonstrating its powerful antihemolytic activity.
3.4. MDA Measurements
The degree of protection conferred by ESV against AAPH-induced lipid peroxidation in human erythrocytes was evaluated by measuring MDA levels. At ESV concentrations of 100 and 125 μg/mL, the MDA levels were decreased throughout the course of the assay (data not shown) and after 5 h of incubation (Figure 2(c)).
3.5. Cytotoxic Activity and Cell Death Profile
ESV promoted cell death in both tested cell lines. IC50 values indicated that ESV was more effective in Jurkat cells than in the erythroleukemic cell line K562 (IC50 = 27.6 μg/mL and 67.5 μg/mL, resp.) (Figures 3(a) and 3(b)). ESV treatment promoted double staining in both cell lines. This type of death was evident in 71.9 ± 5.7% of Jurkat cells and 30.1 ± 2.8% of K562 cells after treatment with 40 and 80 μg/mL of extract, respectively (Figures 4(a) and 4(b)).
Figure 3.

Viability of leukemic Jurkat (a) and K562 (b) cells after treatment with different concentrations of ESV. ∗ P < 0.05 compared to the untreated control group.
Figure 4.

Cell death profiles of ESV-treated Jurkat (a) and K562 (b) cells. Anx−/PI−, viable cells; Anx+/PI−, apoptotic cells; Anx−/PI+, necrotic cells; and Anx+/PI+, late apoptotic cells. ∗ P < 0.05 compared to the respective control groups.
3.6. Mitochondrial Membrane Potential
The mitochondrial membrane potentials of Jurkat and K562 leukemic cells decreased after 24 h of incubation with ESV, as evidenced by a decrease in red fluorescence and an increase in green fluorescence compared to untreated cells. The mitochondrial membrane potential was reduced by 91.0 ± 4.3% in Jurkat cells and by 74.7 ± 7.3% in K562 cells after treatment with 27.6 and 67.5 μg/mL of extract, respectively (Figures 5(a) and 5(b)).
Figure 5.

Mitochondrial membrane potential of leukemic Jurkat (a) and K562 (b) cells treated with different ESV concentrations. ∗∗∗ P < 0.0001 compared to the untreated control group.
3.7. Caspase-3 Activity
The Jurkat cell line, which was more sensitive to ESV activity, was used to investigate ESV-promoted cell death mechanisms. The fluorescence histogram (Figure 6(a)) showed a rightward shift (greater fluorescence values), indicating the activation of the apoptosis-inducing enzyme caspase-3. Cleaved caspase-3 levels increased 4.5-fold in ESV-treated cells relative to untreated cells (Figure 6(b)).
Figure 6.

Histogram (a) and representative graph (b) of caspase-3 activation in ESV-treated Jurkat cells. ∗∗∗ P < 0.0001 compared to the untreated control group.
3.8. Pan-Caspase Inhibition and Intracellular Calcium Chelation
Jurkat cells were preincubated with the pan-caspase inhibitor Z-VAD-FMK to assess whether ESV cytotoxicity was mainly mediated by caspase activation. Z-VAD-FMK pretreatment did not decrease ESV-induced cell death (Figure 7). However, the intracellular Ca2+ chelator BAPTA-AM partially inhibited ESV-induced death in Jurkat cells (Figure 7).
Figure 7.
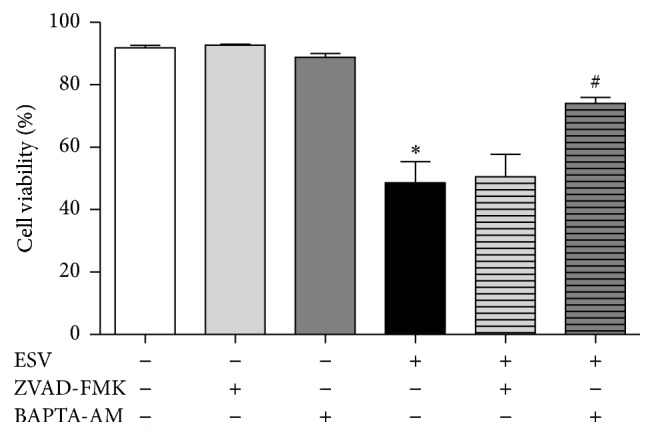
Involvement of caspases (via the pan-caspase inhibitor Z-VAD-FMK) and intracellular calcium (using the chelator BAPTA-AM) in ESV-induced Jurkat cell cytotoxicity. ∗ P < 0.05 compared to the untreated control group. # P < 0.05 compared to the ESV group.
3.9. Cell Cycle Phases
Histograms were used to show the distributions of cell cycle phases in control and ESV-treated Jurkat cells after 24 h of incubation (Figure 8(a)). The results show a decreased portion of cells in G0/G1 phase (17.4 ± 0.6%) and increased portions of cells in S and G2/M phases (30.7 ± 1.8% and 26.5 ± 1.9%, resp.) (Figure 8(b)). These results indicate that ESV inhibits the progression of cell cycle transitions.
Figure 8.

Histogram (a) and representative graph (b) of cell cycle distribution after 24 h of treatment with ESV. ∗ P < 0.05 and ∗∗ P < 0.001 compared to the untreated control group.
4. Discussion
Brazilian biodiversity is rich in active compounds with high potential for development of new therapeutic drugs, particularly antioxidants and anticancer agents. Several plant species found in Brazil have been characterized for their antioxidant and cytotoxic activities in several tumor cell lines [31–33].
In the present study, an ethanol extract of S. velutina leaves exhibited antioxidant activity and showed cytotoxic effects against two leukemic cell lines, Jurkat and K562. The antioxidant activity of ESV was demonstrated in AAPH-incubated human erythrocytes; DPPH free radical scavenging and inhibition of lipid peroxidation led to decreases in oxidative hemolysis and malondialdehyde production. This activity of ESV is likely related to the presence of flavone derivatives with antioxidant activity, such as epigallocatechin, epicatechin, rutin, kaempferol glycosides, and dimeric and trimeric proanthocyanidins, in the leaves [34–37].
Flavonoids can donate hydrogen atoms to radicals, protecting against lipid peroxidation, and this ability is associated with the presence of a dihydroxylated B-ring [38]. Similar to other phenolic compounds, the antioxidant activity of flavonoids is ascribed to the presence of free hydroxyl groups in the molecule, and the level of antioxidant activity increases concomitantly with the number of hydroxyl groups [39].
Excess free radicals in the body can promote not only lipid peroxidation but also oxidative DNA damage, leading to the development of early stages of mutagenesis and carcinogenesis [40, 41]. Thus, compounds with antioxidant properties have fundamental roles in preventing diseases such as cancer.
The cytotoxic activity of ESV against leukemic lines was evaluated with an Annexin/PI-stain cell death assay. Furthermore, activation of caspase-3 was observed but was not confirmed to be a main mechanism of cell death when the pan-caspase inhibitor Z-VAD-FMK was used. Different extracts can promote several different death mechanisms simultaneously because of their different compositions. This fact complicates the analysis and identification of specific cell death pathways.
The ESV-induced decrease in mitochondrial membrane potential is consistent with the observed cytotoxic effect of ESV against leukemic cells. The changes in potential are likely due to increased mitochondrial membrane permeability arising from an increase in intracellular calcium levels, a characteristic of necrotic cell death [42, 43].
The present study demonstrated the involvement of calcium in cell death, as the reduced cell viability of extract-treated cells was reversed by incubation with the calcium chelator BAPTA-AM. High calcium levels promote the opening of mitochondrial permeability transition pores; these pores are nonselective and thus release the contents of the intermembrane space of the mitochondrion [44].
Another mechanism of the ESV-induced cytotoxicity against Jurkat cells consists of cell cycle arrest. ESV promoted a decrease in the number of cells in G0/G1 phase and an increase in the number of cells in S and G2 phases. Established anticancer drugs, such as cisplatin and doxorubicin, exert a similar profile change in the tumor cell cycle [14, 45]. Mueller et al. [46] observed that cisplatin is likely to be active in G2/M phases because cells in these phases are more sensitive to DNA damage, as DNA repair mechanisms are less active than in G1/S phases. Flavonoids are phytochemicals that are known to induce cell cycle arrest by decreasing cellular levels of cyclin B and cyclin-dependent kinase 1, which are responsible for controlling cell cycle progression between S and M stages [47]. Furthermore, one of the major anticancer mechanisms ascribed to flavonoids is their ability to induce cell cycle changes in tumor cell lines [48]. Thus, cyclin-dependent kinase inhibitors (CDKIs) have generated great interest for their ability to arrest the tumor cell cycle and prevent tumor cell proliferation; such cell cycle arrest may be the main mechanism through which ESV operates [49]. However, plant extracts are natural products with complex chemical composition, and biologically active compounds in extracts may act alone or synergistically through different pathways.
In conclusion, an ethanol extract of S. velutina leaves exhibited antioxidant activity and showed cytotoxic effects on leukemic cells by activating intracellular calcium and caspase-3, decreasing mitochondrial membrane potential, and arresting the cell cycle in S and G2 phases.
Acknowledgments
This work was supported by grants of the Fundação de Apoio ao Desenvolvimento do Ensino, Ciência e Tecnologia do Mato Grosso do Sul (FUNDECT), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES). Edson Lucas dos Santos, Carlos Alexandre Carollo, and Edgar J. Paredes-Gamero are recipients of fellowships from Brazilian National Research Council (CNPq), Brazil.
Abbreviations
- AA:
Ascorbic acid
- AAPH:
2,2′-Azobis-(2-amidinopropane) dihydrochloride
- Abs:
Absorbance
- Anx:
Annexin V-FITC
- BHT:
Butylhydroxytoluene
- Ca2+:
Calcium
- CaCl2:
Calcium chloride
- DAD:
Diode Array Detector
- DPPH:
2,2-Diphenyl-1-picrylhydrazyl
- ESI-QTOF-micrOTOF QII:
Electrospray ionization time-of-flight
- ESV:
Ethanol extract of S. velutina leaves
- JC-1:
5,5′,6,6′-Tetrachloro-1,1′,3,3′-tetraethylbenzimidazolylcarbocyanine iodide
- MDA:
Malondialdehyde
- NaCl:
Sodium chloride
- PI:
Propidium iodide
- SEM:
Standard error of the mean
- TBA:
Thiobarbituric acid
- UFLC:
Ultra Fast Liquid Chromatography.
Competing Interests
The authors declare that they have no competing interests.
References
- 1.Sultan S. Reviewing the protective role of antioxidants in oxidative stress caused by free radicals. Asian Pacific Journal of Health Science. 2014;1:401–406. [Google Scholar]
- 2.Lobo V., Patil A., Phatak A., Chandra N. Free radicals, antioxidants and functional foods: impact on human health. Pharmacognosy Reviews. 2010;4(8):118–126. doi: 10.4103/0973-7847.70902. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Siegel R., Naishadham D., Jemal A. Cancer statistics, 2013. CA: A Cancer Journal for Clinicians. 2013;63(1):11–30. doi: 10.3322/caac.21166. [DOI] [PubMed] [Google Scholar]
- 4.American Cancer Society. Leukemia-Chronic Myeloid (Myelogenous) Overview. 2013. [Google Scholar]
- 5.Asmaa M. J. S., Al-Jamal H. A. N., Ang C. Y., Asan J. M., Seeni A., Johan M. F. Apoptosis induction in MV4-11 and K562 human leukemic cells by Pereskia sacharosa (Cactaceae) leaf crude extract. Asian Pacific Journal of Cancer Prevention. 2014;15(1):475–481. doi: 10.7314/apjcp.2014.15.1.475. [DOI] [PubMed] [Google Scholar]
- 6.Srdic-Rajic T., Tisma-Miletic N., Cavic M., et al. Sensitization of K562 leukemia cells to doxorubicin by the Viscum album extract. Phytotherapy Research. 2016;30(3):485–495. doi: 10.1002/ptr.5554. [DOI] [PubMed] [Google Scholar]
- 7.Newman D. J., Cragg G. M. Natural products as sources of new drugs from 1981 to 2014. Journal of Natural Products. 2016;79(3):629–661. doi: 10.1021/acs.jnatprod.5b01055. [DOI] [PubMed] [Google Scholar]
- 8.Balunas M. J., Kinghorn A. D. Drug discovery from medicinal plants. Life Sciences. 2005;78(5):431–441. doi: 10.1016/j.lfs.2005.09.012. [DOI] [PubMed] [Google Scholar]
- 9.Nagle A., Hur W., Gray N. S. Antimitotic agents of natural origin. Current Drug Targets. 2006;7(3):305–326. doi: 10.2174/138945006776054933. [DOI] [PubMed] [Google Scholar]
- 10.Prakash O., Kumar A., Kumar P., Ajeet Anticancer potential of plants and natural products: a review. American Journal of Pharmacological Sciences. 2013;1(6):104–115. doi: 10.12691/ajps-1-6-1. [DOI] [Google Scholar]
- 11.Mak Y. W., Chuah L. O., Ahmad R., Bhat R. Antioxidant and antibacterial activities of hibiscus (Hibiscus rosa-sinensis L.) and Cassia (Senna bicapsularis L.) flower extracts. Journal of King Saud University-Science. 2013;25:275–282. [Google Scholar]
- 12.Susunaga-Notario A. C., Pérez-Gutiérrez S., Zavala-Sánchez M. Á., et al. Bioassay-guided chemical study of the anti-inflammatory effect of Senna villosa (Miller) H.S. Irwin & Barneby (Leguminosae) in TPA-induced ear edema. Molecules. 2014;19(7):10261–10278. doi: 10.3390/molecules190710261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Thilagam E., Parimaladevi B., Kumarappan C., Mandal S. C. α-Glucosidase and α-amylase inhibitory activity of Senna surattensis . Journal of Acupuncture and Meridian Studies. 2013;6(1):24–30. doi: 10.1016/j.jams.2012.10.005. [DOI] [PubMed] [Google Scholar]
- 14.Pereira R. M., Ferreira-Silva G. Á., Pivatto M., et al. Alkaloids derived from flowers of Senna spectabilis, (–)-cassine and (–)-spectaline, have antiproliferative activity on HepG2 cells for inducing cell cycle arrest in G1/S transition through ERK inactivation and downregulation of cyclin D1 expression. Toxicology in Vitro. 2016;31:86–92. doi: 10.1016/j.tiv.2015.11.018. [DOI] [PubMed] [Google Scholar]
- 15.Viegas Junior C., de Rezende A., Silva D. H. S., et al. Aspectos químicos, biológicos e etnofarmacológicos do gênero Cassia . Química Nova. 2006;29(6):1279–1286. doi: 10.1590/s0100-40422006000600025. [DOI] [Google Scholar]
- 16.Delumlle L., Demeyer K. Anthraquinones in Plants—Source, Safety and Applications in Gastrointestinal Health. Nottingham University Press; 2008. [Google Scholar]
- 17.Gupta D., Gupta R. K. Bioprotective properties of Dragon's blood resin: in vitro evaluation of antioxidant activity and antimicrobial activity. BMC Complementary and Alternative Medicine. 2011;11, article 13:1–9. doi: 10.1186/1472-6882-11-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Campos J. F., Das Santos U. P., Da Rocha P. D. S., et al. Antimicrobial, antioxidant, anti-inflammatory, and cytotoxic activities of propolis from the stingless bee Tetragonisca fiebrigi (Jataí) Evidence-Based Complementary and Alternative Medicine. 2015;2015:11. doi: 10.1155/2015/296186.296186 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Paredes-Gamero E. J., Martins M. N. C., Cappabianco F. A. M., Ide J. S., Miranda A. Characterization of dual effects induced by antimicrobial peptides: regulated cell death or membrane disruption. Biochimica et Biophysica Acta—General Subjects. 2012;1820(7):1062–1072. doi: 10.1016/j.bbagen.2012.02.015. [DOI] [PubMed] [Google Scholar]
- 20.Moraes V. W. R., Caires A. C. F., Paredes-Gamero E. J., Rodrigues T. Organopalladium compound 7b targets mitochondrial thiols and induces caspase-dependent apoptosis in human myeloid leukemia cells. Cell Death & Disease. 2013;4, article e658:8. doi: 10.1038/cddis.2013.190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Bechara A., Barbosa C. M. V., Paredes-Gamero E. J., et al. Palladacycle (BPC) antitumour activity against resistant and metastatic cell lines: the relationship with cytosolic calcium mobilisation and cathepsin B activity. European Journal of Medicinal Chemistry. 2014;79:24–33. doi: 10.1016/j.ejmech.2014.03.073. [DOI] [PubMed] [Google Scholar]
- 22.Dou J., Lee V. S. Y., Tzen J. T. C., Lee M.-R. Identification and comparison of phenolic compounds in the preparation of oolong tea manufactured by semifermentation and drying processes. Journal of Agricultural and Food Chemistry. 2007;55(18):7462–7468. doi: 10.1021/jf0718603. [DOI] [PubMed] [Google Scholar]
- 23.Rao K. V., Damu A. G., Jayaprakasam B., Gunasekar D. Flavonol glycosides from Cassia hirsuta . Journal of Natural Products. 1999;62(2):305–306. doi: 10.1021/np980195c. [DOI] [PubMed] [Google Scholar]
- 24.El-Sayed N. H., Dooh A. M. A., El-Khrisy S. A. M., Mabry T. J. Flavonoids of Cassia italica . Phytochemistry. 1992;31(6):p. 2187. doi: 10.1016/0031-9422(92)80400-9. [DOI] [Google Scholar]
- 25.Morimoto S., Nonaka G., Chen R., Nishioka I. Tannins and related compounds. LXI. Isolation and structures of novel bi- and triflavanoids from the leaves of Cassia fistula L. Chemical and Pharmaceutical Bulletin. 1988;36(1):39–47. doi: 10.1248/cpb.36.39. [DOI] [Google Scholar]
- 26.Hatano T., Yamashita A., Hashimoto T., et al. Flavan dimers with lipase inhibitory activity from Cassia nomame . Phytochemistry. 1997;46(5):893–900. doi: 10.1016/s0031-9422(97)00367-1. [DOI] [Google Scholar]
- 27.Coetzee J., Mciteka L., Malan E., Ferreira D. Structure and synthesis of butiniflavan-epicatechin and -epigallocatechin probutinidins. Phytochemistry. 1999;52(4):737–743. doi: 10.1016/s0031-9422(99)00261-7. [DOI] [Google Scholar]
- 28.Callemien D., Collin S. Use of RP-HPLC-ESI (-)-MS/MS to differentiate various proanthocyanidin isomers in lager beer extracts. The Journal of the American Society of Brewing Chemists. 2008;66:109–115. [Google Scholar]
- 29.Gu L., Kelm M. A., Hammerstone J. F., et al. Liquid chromatographic/electrospray ionization mass spectrometric studies of proanthocyanidins in foods. Journal of Mass Spectrometry. 2003;38(12):1272–1280. doi: 10.1002/jms.541. [DOI] [PubMed] [Google Scholar]
- 30.Lazarus S. A., Adamson G. E., Hammerstone J. F., Schmitz H. H. High-performance liquid chromatography/mass spectrometry analysis of proanthocyanidins in foods and beverages. Journal of Agricultural and Food Chemistry. 1999;47(9):3693–3701. doi: 10.1021/jf9813642. [DOI] [PubMed] [Google Scholar]
- 31.Casagrande J. C., Macorini L. F. B., Antunes K. A., et al. Antioxidant and cytotoxic activity of hydroethanolic extract from Jacaranda decurrens leaves. PLoS ONE. 2014;9(11):1–8. doi: 10.1371/journal.pone.0112748.e112748 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Serpeloni J. M., Leal Specian A. F., Ribeiro D. L., et al. Antimutagenicity and induction of antioxidant defense by flavonoid rich extract of Myrcia bella Cambess. in normal and tumor gastric cells. Journal of Ethnopharmacology. 2015;176:345–355. doi: 10.1016/j.jep.2015.11.003. [DOI] [PubMed] [Google Scholar]
- 33.Formagio A. S. N., Ramos D. D., Vieira M. C., et al. Phenolic compounds of Hibiscus sabdariffa and influence of organic residues on its antioxidant and antitumoral properties. Brazilian Journal of Biology. 2015;75(1):69–76. doi: 10.1590/1519-6984.07413. [DOI] [PubMed] [Google Scholar]
- 34.Yang J., Guo J., Yuan J. In vitro antioxidant properties of rutin. LWT—Food Science and Technology. 2008;41(6):1060–1066. doi: 10.1016/j.lwt.2007.06.010. [DOI] [Google Scholar]
- 35.Chua L. S. A review on plant-based rutin extraction methods and its pharmacological activities. Journal of Ethnopharmacology. 2013;150(3):805–817. doi: 10.1016/j.jep.2013.10.036. [DOI] [PubMed] [Google Scholar]
- 36.Pauli E. D., Malta G. B., Sanchez P. M., Moreira I. C., Scarminio I. S. Mixture design analysis of solvent extractor effects on epicatechin, epigallocatechin gallate, epigallocatechin and antioxidant activities of the Camellia sinensis L. leaves. Analytical Chemistry Research. 2014;2:23–29. doi: 10.1016/j.ancr.2014.08.001. [DOI] [Google Scholar]
- 37.Plumb G. W., De Pascual-Teresa S., Santos-Buelga C., Cheynier V., Williamson G. Antioxidant properties of catechins and proanthocyanidins: effect of polymerisation, galloylation and glycosylation. Free Radical Research. 1998;29(4):351–358. doi: 10.1080/10715769800300391. [DOI] [PubMed] [Google Scholar]
- 38.Valko M., Rhodes C. J., Moncol J., Izakovic M., Mazur M. Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chemico-Biological Interactions. 2006;160(1):1–40. doi: 10.1016/j.cbi.2005.12.009. [DOI] [PubMed] [Google Scholar]
- 39.Kurek-Górecka A., Rzepecka-Stojko A., Górecki M., Stojko J., Sosada M., Swierczek-Zieba G. Structure and antioxidant activity of polyphenols derived from propolis. Molecules. 2014;19(1):78–101. doi: 10.3390/molecules19010078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Frei B. Reactive oxygen species and antioxidant vitamins: mechanisms of action. The American Journal of Medicine. 1994;97(3):5–13. doi: 10.1016/0002-9343(94)90292-5. [DOI] [PubMed] [Google Scholar]
- 41.Valko M., Leibfritz D., Moncol J., Cronin M. T. D., Mazur M., Telser J. Free radicals and antioxidants in normal physiological functions and human disease. The International Journal of Biochemistry & Cell Biology. 2007;39(1):44–84. doi: 10.1016/j.biocel.2006.07.001. [DOI] [PubMed] [Google Scholar]
- 42.Lemasters J. J., Qian T., Bradham C. A., et al. Mitochondrial dysfunction in the pathogenesis of necrotic and apoptotic cell death. Journal of Bioenergetics and Biomembranes. 1999;31(4):305–319. doi: 10.1023/A:1005419617371. [DOI] [PubMed] [Google Scholar]
- 43.Festjens N., Vanden Berghe T., Vandenabeele P. Necrosis, a well-orchestrated form of cell demise: signalling cascades, important mediators and concomitant immune response. Biochimica et Biophysica Acta (BBA)—Bioenergetics. 2006;1757(9-10):1371–1387. doi: 10.1016/j.bbabio.2006.06.014. [DOI] [PubMed] [Google Scholar]
- 44.Kinnally K. W., Peixoto P. M., Ryu S.-Y., Dejean L. M. Is mPTP the gatekeeper for necrosis, apoptosis, or both? Biochimica et Biophysica Acta—Molecular Cell Research. 2011;1813(4):616–622. doi: 10.1016/j.bbamcr.2010.09.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Wang W., Chen K., Liu Q., et al. Suppression of tumor growth by Pleurotus ferulae ethanol extract through induction of cell apoptosis, and inhibition of cell proliferation and migration. PLoS ONE. 2014;9(7):1–14. doi: 10.1371/journal.pone.0102673.e102673 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Mueller S., Schittenhelm M., Honecker F., et al. Cell-cycle progression and response of germ cell tumors to cisplatin in vitro . International Journal of Oncology. 2006;29(2):471–479. [PubMed] [Google Scholar]
- 47.Carocho M., Ferreira I. C. F. R. The role of phenolic compounds in the fight against cancer—a review. Anti-Cancer Agents in Medicinal Chemistry. 2013;13(8):1236–1258. doi: 10.2174/18715206113139990301. [DOI] [PubMed] [Google Scholar]
- 48.Singh R. P., Agarwal R. Natural flavonoids targeting deregulated cell cycle progression in cancer cells. Current Drug Targets. 2006;7(3):345–354. doi: 10.2174/138945006776055004. [DOI] [PubMed] [Google Scholar]
- 49.Schwartz G. K., Shah M. A. Targeting the cell cycle: a new approach to cancer therapy. Journal of Clinical Oncology—Biology of Neoplasia. 2005;23(36):9408–9421. doi: 10.1200/jco.2005.01.5594. [DOI] [PubMed] [Google Scholar]