

Abstract
GoxA is a glycine oxidase bearing a protein-derived cysteine tryptophylquinone (CTQ) cofactor that is formed by posttranslational modifications catalyzed by a flavoprotein, GoxB. Two forms of GoxA were isolated. An active form with mature CTQ and an inactive precursor protein that lacked CTQ. The active GoxA was present as a homodimer with no detectable affinity for GoxB, whereas the precursor was isolated as a monomer in a tight complex with one GoxB. Thus, the interaction of GoxA with GoxB and subunit assembly of mature GoxA are each dependent on the extent of CTQ biosynthesis.
Keywords: quinoprotein, flavoprotein, posttranslational modification, protein biosynthesis
Quinoproteins may contain exogenous quinone cofactors such as pyrroloquinoline quinone1 or protein-derived2 quinone cofactors that are formed by irreversible posttranslational modification of amino acid residues, most notably Tyr and Trp (Figure 1)3. Tyrosine-derived cofactors include trihydroxyphenylalanine (Topa) quinone (TPQ)4 which is found in several copper-containing amine oxidases5 and lysyl tyrosylquinone (LTQ) which is found in lysyl oxidase6. Tryptophan-derived cofactors include tryptophan tryptophylquinone (TTQ) which is found in amine dehydrogenases and cysteine tryptophylquinone (CTQ) which was been found in quinohemoprotein amine dehydrogenase (QHNDH)7, in an L-lysine-ε-oxidase (LodA)8 and in a glycine oxidase (GoxA)9, which catalyzes the oxidative deamination of glycine (eq 1).
| (1) |
Figure 1.
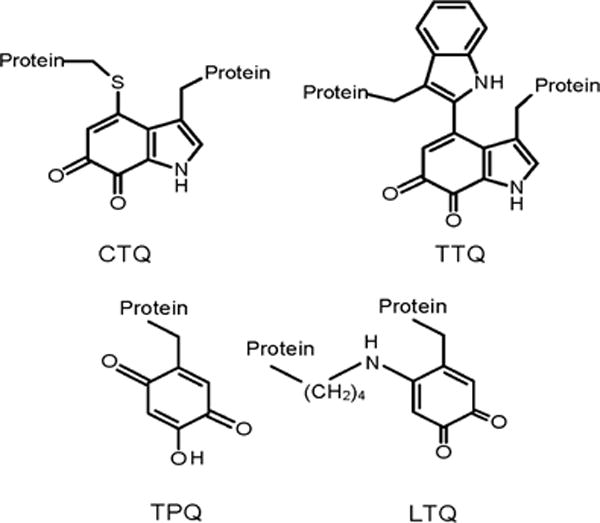
Protein-derived quinone cofactors formed by posttranslational modification. CTQ is cysteine tryptophylquinone cofactor. TTQ is tryptophan tryptophylquinone cofactor. TPQ is 2,4,5-trihydroxyphenylalanine quinone or Topaquinone. LTQ is lysine tyrosylquinone.
The mechanism of the biosynthesis of protein-derived quinone cofactors differs from cofactor to cofactor10. For the Tyr-derived cofactors it is a self-processing event which is dependent on the presence of copper and O2. The copper facilitates oxygen insertion into the Tyr side chain and subsequently remains in the active site to assist in catalysis11, 12. The biosynthesis of TTQ in methylamine dehydrogenase (MADH) includes posttranslational modifications that are catalyzed by a diheme enzyme called MauG13, 14. The mechanism of the biosynthesis of CTQ in QHNDH has yet to be fully characterized, but it does require one or more other gene products10. In the case of LodA and GoxA it has been shown that flavoproteins are required for the biosynthesis of CTQ, although no mechanistic studies have been thus far reported. The posttranslational modifications required for CTQ biosynthesis are the incorporation of two oxygen atoms into the side-chain of a specific Trp residue and the covalent cross-link formation between that residue and a Cys residue (Figure 1). LodA and GoxA were isolated from the marine bacterium Marinomonas mediterranea8, 9. The gene clusters which encode LodA and GoxA each contain only one other gene, lodB and goxB15. Each of these latter two genes is predicted to encode a putative flavoprotein. Expression of either active LodA or GoxA with the mature CTQ cofactor requires that lodA and goxA each be co-expressed with lodB and goxB, respectively16. Expression of either lodA or goxA in the absence of lodB or goxB, respectively, leads to the production of inactive precursor forms of the proteins that lack CTQ17.
This study reports the isolation and characterization of a complex of a GoxA precursor protein and GoxB from induced cultures of recombinant E. coli. It is shown that a flavin is indeed present in this complex with the putative flavoprotein, GoxB, and that mature CTQ is not present in the precursor form of GoxA. An active GoxA with mature CTQ, which is not in complex with GoxB, is also isolated from the same cultures. Evidence is presented that the affinity between GoxA and GoxB is dependent upon the extent of CTQ biosynthesis. Furthermore, it is shown that the oligomerization state of GoxA is dependent on the extent of CTQ biosynthesis. These results are compared and contrasted with the features of the biosynthesis of TTQ and TPQ in their respective enzymes.
During purification of the His-tagged recombinant GoxA, two protein fractions eluted from the Ni-NTA resin affinity column, one at approximately 60 mM imidazole and the other at approximately 150 mM imidazole. When assayed for glycine oxidase activity9 the first fraction exhibited no activity, whereas the second fraction was active and exhibited a kcat of 14.90 ± 0.02 s−1 under aerobic conditions (252μM O2) with 2 mM glycine at 30°C (see SI for experimental details). Each of the fractions exhibited a different visible absorption spectrum. The fraction that was active exhibited spectral features in the 350–420 nm range with peaks that are consistent with the presence of CTQ (Figure 2A). Similar visible absorbance spectra have been reported for LodA18, the γ subunit of QHNDH19 and a synthetic CTQ model compound20. In contrast, the inactive fraction exhibited an absorbance spectrum with features characteristic of a flavin. (Figure 2B). This suggested that the fraction contained GoxB, although that protein did not have a His-tag. From this spectrum it could not be concluded whether or not CTQ was also present as the absorbance of the flavin would mask the CTQ absorbance.
Figure 2.
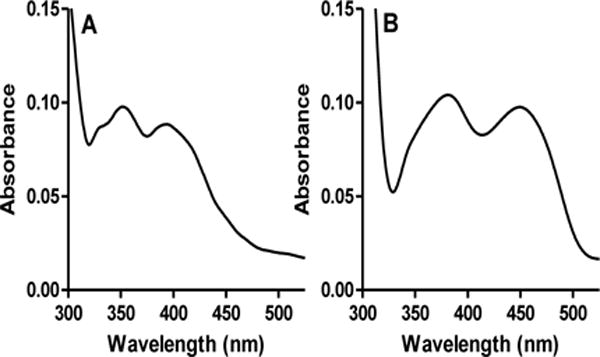
Visible absorption spectra of the two protein fractions that eluted from the Ni-NTA affinity column. A. Absorption spectrum of the active fraction which exhibits a spectrum characteristic of CTQ. B. Absorption spectrum of the inactive fraction which exhibits a spectrum characteristic of flavin.
The two protein fractions were analyzed by SDS-PAGE (Figure 3A). The inactive fraction (lanes 6,7) contained two species. The larger band migrated as expected for GoxA which has a predicted mass from the gene (Marme_1655) of 76284.8 Da. The other migrated as would be expected for GoxB which has a predicted mass from the gene (Marme_1654) of 41856.3 Da9. That active protein fraction (lane 1) contained only one major species which migrated as expected for GoxA. A second lower band is seen. A similar gel was also subjected to Western Blot using an anti His-tag antibody and this minor band also reacted. Thus, this most likely is a minor degradation product.
Figure 3.
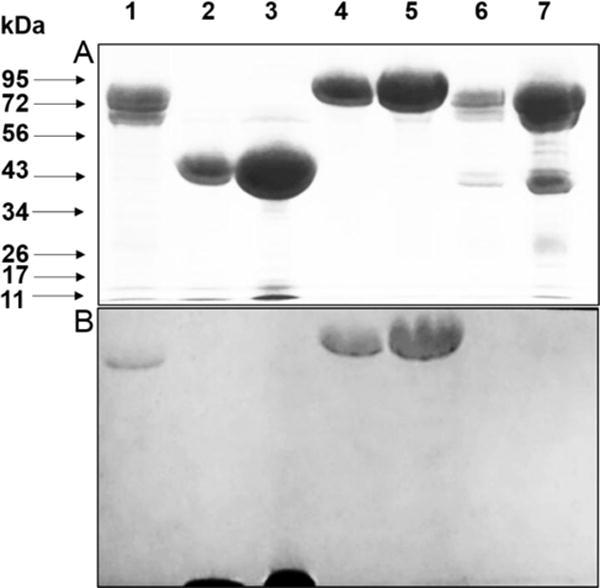
SDS-PAGE and quinoprotein staining of protein fractions. A. 7.5% gel stained for protein. B. Electroblot which was stained for quinoproteins. A gel identical to that in A was subjected to electrophoretic transfer of the proteins and stained for quinoproteins (see SI for experimental details). Lanes: (1) 20 μg active protein fraction, (2) 20 μg MADH, (3) 80 μg MADH (4) 20 μg LodA, (5) 40 μg LodA (6) 20 μg inactive protein fraction, (7) 100 μg inactive protein fraction.
Analysis by mass spectrometry indicated that the inactive fraction contained a protein with a molecular weight of the untagged GoxB. The much larger intact GoxA species did not fly in the mass spectrometer and so it was not possible to determine from the mass whether or not CTQ was present. It was not possible to separate the two proteins in the inactive fraction by other chromatographic techniques. The sample was subjected to anion exchange (DEAE) chromatography, hydrophobic interaction chromatography using butyl sepharose and size exclusion chromatography. In each case the proteins co-eluted indicating that the two proteins interact with each other to form a tight complex.
Two possible reasons for the lack of activity in the fraction containing the GoxA-GoxB complex are that either the presence of the second protein interferes with the reaction of GoxA, or that GoxA in that fraction is actually a precursor lacking the mature CTQ cofactor. As the presence or absence of CTQ could not be confirmed by the absorbance spectrum or mass spectrometry, an alternative method was used to determine whether CTQ was present. The proteins were subjected to SDS-PAGE followed by electroblot to a nitrocellulose membrane which was subjected to a staining protocol that is specific for covalent quinoproteins21 (Figure 3B).
The active GoxA that eluted alone (Lane 1) stained positive for the presence of quinone. The GoxA which co-eluted with GoxB (Lanes 6,7) did not stain indicating that it does not have the mature CTQ cofactor. Thus, it is actually a precursor protein of GoxA. Additional proteins were included on the gel as positive and negative controls. Two proteins were included in which the presence of the quinone cofactor has been verified in crystal structures. MADH contains two subunits with TTQ present in the smaller subunit22. The smaller subunit stained positive and the larger one did not (Lanes 2,3). LodA, which contains CTQ23 also stained positive (Lanes 4,5). The GoxB present in complex with the precursor form of GoxA did not stain. Comparable amounts of GoxA, MADH, LodA, and the precursor GoxA-GoxB complex were loaded on the gel. GoxA and LodA did not transfer to the nitrocellulose as efficiently as the other proteins. For that reason the gel in Figure 3A, which is identical to the one which was used in the electroblot transfer to a nitrocellulose membrane, was intentionally overloaded to ensure that a negative result was not because of insufficient transfer of proteins. For further confirmation a dot blot was performed in which the protein sample was spotted directly on the nitrocellulose membrane, dried and then subjected to the staining procedure. Again the active protein stained positive and the inactive did not, indicating that whereas the active GoxA has mature CTQ, the GoxA in complex with GoxB is a precursor protein lacking mature CTQ.
The inactive and active fractions were also characterized by size exclusion chromatography. The active GoxA with CTQ eluted from the column primarily as a dimer (Figure 4A). Two peaks were observed. The relative mass of the minor peak (16%) was approximately 76 kDa which is the predicted molecular weight of the monomer. The major peak (84%) was approximately 140 kDa consistent with the mass of a homodimer. The inactive fraction containing precursor of GoxA and GoxB eluted 100% as a single peak with a relative mass of approximately 120 kDa which is approximately the sum of the predicted molecular weights of GoxA and GoxB (Figure 4B). When goxA was expressed alone a precursor of GoxA was identify which lacked CTQ. However, it was not possible to analyze the subunit composition of this species for comparison as it was unstable in solution. It tended to aggregate as evidenced by sticking to membranes during concentration and to column resins during purification.
Figure 4.
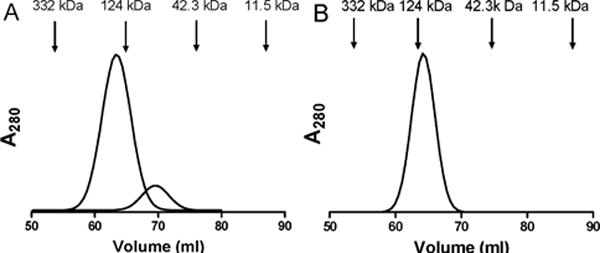
Size exclusion chromatography of the active protein fraction containing GoxA (A) and the inactive protein fraction containing the GoxA precursor in complex with GoxB (B). The positions of elution and molecular weights of standards used to calibrate the column are indicated.
The goxB gene does not exhibit a sequence motif characteristic of an NADH binding site. Nevertheless, to test the possibility that flavin-dependent CTQ biosynthesis within the precursor GoxA-GoxB complex was NADH dependent, NADH and NADPH were each incubated with the complex. This did not result in formation of active GoxA or alteration of the complex. Elucidation of the mechanism of activation of the flavin to enable it to catalyze the CTQ biosynthetic reactions will be a topic of future studies.
It is worth noting that in the native gox operon, as well as in the construct used for the recombinant expression, the stop codon of goxA overlaps with the start codon of goxB, indicating that both genes are translationally coupled (SI Fig. 1). This is true of the majority of the genes encoding proteins similar to GoxA17, 24. This genetic organization may be relevant for the expression of proteins forming complexes25, and in particular for the biosynthesis and assembly of GoxA.
The implications of these results are profound. Together the results indicate that the GoxA precursor forms a tight complex with GoxB, whereas GoxA with mature CTQ does not. Thus, complex formation between the two is strongly dependent upon the extent of CTQ biosynthesis. Furthermore, the native dimeric structure of active GoxA is not achieved until after CTQ formation and release of GoxB. Thus, the oligomerization state of GoxA is also dependent upon the extent of CTQ biosynthesis.
These results are in stark contrast to what was observed for TTQ biosynthesis in MADH by the diheme enzyme MauG26. It was shown that the oxidation of the residues on the precursor protein of MADH (preMADH) did not occur by direct reaction with the hemes but instead by long-range electron transfer from the substrate to the high-valent hemes27. It was possible to crystallize a catalytically competent complex of preMADH and MauG28, but the proteins did not form a stable complex in solution, as do the GoxA precursor and GoxB. Kinetic studies demonstrated that the affinity of MauG for preMADH was approximately 10-fold greater than for MADH with mature TTQ, but the interactions were transient29. In the present study the difference in affinity of GoxB for the GoxA precursor protein is far greater than for the post-translationally modified mature GoxA. In one case a complex forms which cannot be separated and in the other the complex formation is undetectable in solution. Furthermore, the crystallographic studies revealed that prior to TTQ biosynthesis, preMADH existed as a heterotetramer and formed a complex with two molecules of MauG, one associated with each half of the heterotetramer28. Thus, in contrast to the results obtained for GoxA, the subunit assembly of MADH showed no dependence on the extent of TTQ biosynthesis. In that case it preceded CTQ biosynthesis.
LodA which is isolated from the same bacterium as GoxA also contains a CTQ cofactor. The biosynthesis of the cofactor is also dependent upon the flavoprotein LodB. In contrast to what was observed with GoxA, no complex of LodA or LodA precursors with LodB was observed during purification. Interestingly, the preparation of pure LodA was found to contain a significant fraction of a precursor that lacked CTQ but no LodB18. Thus, while preLodA and LodB must also interact to achieve CTQ biosynthesis, the complex formation between those two proteins must be transient as was the case with preMADH and MauG, and different from what was observed for precursor GoxA-GoxB complex. This observation is also consistent with the previous observation that LodB and GoxB cannot substitute for each other and are specific for their respective LodA and GoxA precursor proteins. It seems that it may not just be a matter of specificity, but the mechanisms of action of LodB-dependent and GoxB-dependent CTQ biosynthesis may be significantly different.
The mechanisms of the biosynthesis of protein-derived cofactors are complex and diverse. Most common protein posttranslational modifications, such as phosphorylation and glycosylation, occur at the protein surface which enables the substrate residues to directly contact the modifying enzyme. In contrast, the residues that undergo posttranslational modification in protein-derived quinone cofactors typically reside within the protein. Some of these modifications are self-processing events, such as the biogenesis of the TPQ cofactor of copper-containing amine oxidases. In this case the host protein binds copper which is used to activate oxygen for insertion into the side-chain of a specific Tyr residue11 and then participates in catalysis30, 31. For TTQ and CTQ biosynthesis modifying enzymes are required. This initial characterization of the tight complex formed by the GoxA precursor and GoxB is an important step towards future studies to characterize the details of the mechanism of CTQ formation. As GoxB is a flavoprotein, this mechanism will likely describe novel reactions, the catalysis of which are unprecedented for a flavin cofactor.
Supplementary Material
Acknowledgments
The authors thank Dr, Carrie Wilmot and Chao Li at the University of Minnesota for performing mass spectrometry. We also thank Yu Tang and Dr. Patricia Lucas-Elío for technical assistance.
Funding Sources
This research was supported by internal funds from the Burnett School of Biomedical Sciences and in part the National Institute of General Medical Sciences of the National Institutes of Health under award number R37GM41574 (V.L.D.).
ABBREVIATIONS
- CTQ
cysteine tryptophylquinone
- LTQ
lysine tyrosylquinone
- MADH
methylamine dehydrogenase
- QHNDH
quinohemoprotein amine dehydrogenase
- TPQ
2,4,5-trihydroxyphenylalanine quinone
- TTQ
tryptophan tryptophylquinone
Footnotes
ASSOCIATED CONTENT
The Supporting Information is available. Detailed experimental Methods and Figure S1.
Author Contributions
The manuscript was written through contributions of all authors. All authors have approved the final version of the manuscript.
Notes
The authors declare no competing financial interests.
References
- 1.Davidson VL. Pyrroloquinoline quinone (PQQ) from methanol dehydrogenase and tryptophan tryptophylquinone (TTQ) from methylamine dehydrogenase. Adv Protein Chem. 2001;58:95–140. doi: 10.1016/s0065-3233(01)58003-1. [DOI] [PubMed] [Google Scholar]
- 2.Davidson VL. Protein-derived cofactors. Expanding the scope of post-translational modifications. Biochemistry. 2007;46:5283–5292. doi: 10.1021/bi700468t. [DOI] [PubMed] [Google Scholar]
- 3.Davidson VL. Generation of protein-derived redox cofactors by posttranslational modification. Mol Biosyst. 2011;7:29–37. doi: 10.1039/c005311b. [DOI] [PubMed] [Google Scholar]
- 4.Janes SM, Mu D, Wemmer D, Smith AJ, Kaur S, Maltby D, Burlingame AL, Klinman JP. A new redox cofactor in eukaryotic enzymes: 6-hydroxydopa at the active site of bovine serum amine oxidase. Science. 1990;248:981–987. doi: 10.1126/science.2111581. [DOI] [PubMed] [Google Scholar]
- 5.Mure M. Tyrosine-derived quinone cofactors. Acct Chem Res. 2004;37:131–139. doi: 10.1021/ar9703342. [DOI] [PubMed] [Google Scholar]
- 6.Wang SX, Mure M, Medzihradszky KF, Burlingame AL, Brown DE, Dooley DM, Smith AJ, Kagan HM, Klinman JP. A crosslinked cofactor in lysyl oxidase: Redox function for amino acid side chains. Science. 1996;273:1078–1084. doi: 10.1126/science.273.5278.1078. [DOI] [PubMed] [Google Scholar]
- 7.Datta S, Mori Y, Takagi K, Kawaguchi K, Chen ZW, Okajima T, Kuroda S, Ikeda T, Kano K, Tanizawa K, Mathews FS. Structure of a quinohemoprotein amine dehydrogenase with an uncommon redox cofactor and highly unusual crosslinking. Proc Natl Acad Sci U S A. 2001;98:14268–14273. doi: 10.1073/pnas.241429098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Gomez D, Lucas-Elio P, Sanchez-Amat A, Solano F. A novel type of lysine oxidase: L-lysine-epsilon-oxidase. Biochim Biophys Acta. 2006;1764:1577–1585. doi: 10.1016/j.bbapap.2006.08.014. [DOI] [PubMed] [Google Scholar]
- 9.Campillo-Brocal JC, Lucas-Elio P, Sanchez-Amat A. Identification in Marinomonas mediterranea of a novel quinoprotein with glycine oxidase activity. MicrobiologyOpen. 2013;2:684–694. doi: 10.1002/mbo3.107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Klinman JP, Bonnot F. Intrigues and intricacies of the biosynthetic pathways for the enzymatic quinocofactors: PQQ, TTQ, CTQ, TPQ, and LTQ. Chem Rev. 2014;114:4343–4365. doi: 10.1021/cr400475g. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Cai D, Klinman JP. Evidence of a self-catalytic mechanism of 2,4,5-trihydroxyphenylalanine quinone biogenesis in yeast copper amine oxidase. J Biol Chem. 1994;269:32039–32042. [PubMed] [Google Scholar]
- 12.Bollinger JA, Brown DE, Dooley DM. The formation of lysine tyrosylquinone (LTQ) is a self-processing reaction. Expression and characterization of a drosophila lysyl oxidase. Biochemistry. 2005;44:11708–11714. doi: 10.1021/bi0504310. [DOI] [PubMed] [Google Scholar]
- 13.Yukl ET, Liu F, Krzystek J, Shin S, Jensen LM, Davidson VL, Wilmot CM, Liu A. Diradical intermediate within the context of tryptophan tryptophylquinone biosynthesis. Proc Natl Acad Sci U S A. 2013;110:4569–4573. doi: 10.1073/pnas.1215011110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Wang Y, Graichen ME, Liu A, Pearson AR, Wilmot CM, Davidson VL. MauG, a novel diheme protein required for tryptophan tryptophylquinone biogenesis. Biochemistry. 2003;42:7318–7325. doi: 10.1021/bi034243q. [DOI] [PubMed] [Google Scholar]
- 15.Gomez D, Lucas-Elio P, Solano F, Sanchez-Amat A. Both genes in the Marinomonas mediterranea lodAB operon are required for the expression of the antimicrobial protein lysine oxidase. Mol Microbiol. 2010;75:462–473. doi: 10.1111/j.1365-2958.2009.07000.x. [DOI] [PubMed] [Google Scholar]
- 16.Chacon-Verdu MD, Gomez D, Solano F, Lucas-Elio P, Sanchez-Amat A. LodB is required for the recombinant synthesis of the quinoprotein L-lysine-epsilon-oxidase from Marinomonas mediterranea. Appl Microbiol Biot. 2014;98:2981–2989. doi: 10.1007/s00253-013-5168-3. [DOI] [PubMed] [Google Scholar]
- 17.Chacon-Verdu MD, Campillo-Brocal JC, Lucas-Elio P, Davidson VL, Sanchez-Amat A. Characterization of recombinant biosynthetic precursors of the cysteine tryptophylquinone cofactors of L-lysine-epsilon-oxidase and glycine oxidase from Marinomonas mediterranea. Biochim Biophys Acta. 2015;1854:1123–1131. doi: 10.1016/j.bbapap.2014.12.018. [DOI] [PubMed] [Google Scholar]
- 18.Sehanobish E, Chacon-Verdu MD, Sanchez-Amat A, Davidson VL. Roles of active site residues in LodA, a cysteine tryptophylquinone dependent epsilon-lysine oxidase. Arch Biochem Biophys. 2015;579:26–32. doi: 10.1016/j.abb.2015.05.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Fujieda N, Mori M, Kano K, Ikeda T. Spectroelectrochemical evaluation of redox potentials of cysteine tryptophylquinone and two hemes c in quinohemoprotein amine dehydrogenase from Paracoccus denitrificans. Biochemistry. 2002;41:13736–13743. doi: 10.1021/bi026387v. [DOI] [PubMed] [Google Scholar]
- 20.Murakami Y, Yoshimoto N, Fujieda N, Ohkubo K, Hasegawa T, Kano K, Fukuzumi S, Itoh S. Model studies of 6,7-indolequinone cofactors of quinoprotein amine dehydrogenases. J Org Chem. 2007;72:3369–3380. doi: 10.1021/jo0700272. [DOI] [PubMed] [Google Scholar]
- 21.Paz MA, Fluckiger R, Boak A, Kagan HM, Gallop PM. Specific detection of quinoproteins by redox-cycling staining. J Biol Chem. 1991;266:689–692. [PubMed] [Google Scholar]
- 22.Chen L, Doi M, Durley RC, Chistoserdov AY, Lidstrom ME, Davidson VL, Mathews FS. Refined crystal structure of methylamine dehydrogenase from Paracoccus denitrificans at 1.75 A resolution. J Mol Biol. 1998;276:131–149. doi: 10.1006/jmbi.1997.1511. [DOI] [PubMed] [Google Scholar]
- 23.Okazaki S, Nakano S, Matsui D, Akaji S, Inagaki K, Asano Y. X-Ray crystallographic evidence for the presence of the cysteine tryptophylquinone cofactor in L-lysine epsilon-oxidase from Marinomonas mediterranea. J Biochem. 2013;154:233–236. doi: 10.1093/jb/mvt070. [DOI] [PubMed] [Google Scholar]
- 24.Campillo-Brocal JC, Chacon-Verdu MD, Lucas-Elio P, Sanchez-Amat A. Distribution in microbial genomes of genes similar to lodA and goxA which encode a novel family of quinoproteins with amino acid oxidase activity. BMC Genomics. 2015;16:231. doi: 10.1186/s12864-015-1455-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Sneppen K, Pedersen S, Krishna S, Dodd I, Semsey S. Economy of operon formation: cotranscription minimizes shortfall in protein complexes. MBio. 2010;1 doi: 10.1128/mBio.00177-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Davidson VL, Wilmot CM. Posttranslational biosynthesis of the protein-derived cofactor tryptophan tryptophylquinone. Annu Rev Biochem. 2013;82:531–550. doi: 10.1146/annurev-biochem-051110-133601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.AbuTarboush N, Jensen LM, Yukl ET, Geng J, Liu A, Wilmot CM, Davidson VL. Mutagenesis of tryptophan199 suggests that hopping is required for MauG-dependent tryptophan tryptophylquinone biosynthesis. Proc Natl Acad Sci U S A. 2011;108:16956–16961. doi: 10.1073/pnas.1109423108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Jensen LM, Sanishvili R, Davidson VL, Wilmot CM. In crystallo posttranslational modification within a MauG/pre-methylamine dehydrogenase complex. Science. 2010;327:1392–1394. doi: 10.1126/science.1182492. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Shin S, Abu Tarboush N, Davidson VL. Long-range electron transfer reactions between hemes of MauG and different forms of tryptophan tryptophylquinone of methylamine dehydrogenase. Biochemistry. 2010;49:5810–5816. doi: 10.1021/bi1004969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Mukherjee A, Smirnov VV, Lanci MP, Brown DE, Shepard EM, Dooley DM, Roth JP. Inner-sphere mechanism for molecular oxygen reduction catalyzed by copper amine oxidases. J Am Chem Soc. 2008;130:9459–9473. doi: 10.1021/ja801378f. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Dooley DM, Mcguirl MA, Brown DE, Turowski PN, Mcintire WS, Knowles PF. A Cu (I)-semiquinone state in substrate-reduced amine oxidases. Nature. 1991;349:262–264. doi: 10.1038/349262a0. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.