

Abstract
The human oncogenic virus Kaposi's sarcoma herpesvirus (KSHV) is the most common cause of malignancies among AIDS patients. KSHV possesses over hundred genes, including 25 microRNAs (miRNAs). The roles of these miRNAs and many other viral genes in KSHV biology and pathogenesis remain largely unknown. Accordingly, the molecular mechanisms by which KSHV induces tumorigenesis are still poorly defined. Here, we identify KSHV miRNA K12-1 (miR-K12-1) as a novel viral oncogene by activating two important transcription factors, nuclear factor-κb (NF-κB) and signal transducer and activator of transcription 3 (STAT3). Interestingly, miR-K12-1 activates STAT3 indirectly through inducing NF-κB activation and NF-κB-dependent expression of the cytokine interleukin-6 (IL-6) by repressing the expression of the NF-κB inhibitor IκBα. Accordingly, expression of ectopic IκBα or knockdown of NF-κB RelA, IL-6 or STAT3 prevents expression of cell growth genes and suppresses the oncogenicities of both miR-K12-1 and KSHV. These data identify miR-K12-1/NF-κB/IL-6/STAT3 as a novel oncogenic signaling underlying KSHV tumorigenesis. These data also provide the first evidence showing that IL-6/STAT3 signaling acts as an essential mediator of NF-κB oncogenic actions. These findings significantly improve our understanding of KSHV pathogenesis and oncogenic interaction between NF-κB and STAT3.
Keywords: KSHV/HHV8, miR-K12-1, NF-κB, IL-6, STAT3
INTRODUCTION
Kaposi's sarcoma herpesvirus (KSHV), also known as human herpesvirus 8 (HHV8), is the etiological agent of three malignancies mainly associated with acquired immunodeficiency syndrome (AIDS) and immunosuppression: Kaposi's sarcoma (KS), primary effusion lymphoma (PEL) and multicentric Castleman's disease (MCD) [1–3]. Although the molecular biology of this oncogenic virus has been extensively studied during the past two decades, the mechanisms by which KSHV induces tumorigenesis still remain obscure [4]. Those characterized to date suggest that while its lytic infection contributes to tumor promotion in a paracrine manner, the latent infection of KSHV is the direct driving force in tumor formation and maintenance with the expression of a limited set of viral genes capable of hijacking cellular signaling pathways important for host cell proliferation and survival [5].
During latent infection, KSHV also expresses 12 pre-microRNAs (pre-miRNAs) that are processed to yield 25 mature miRNAs [6]. Mature miRNAs are approximately 22 nucleotides long, single-stranded non-coding RNAs that bind to mRNAs, resulting in their degradation and/or translational repression [7, 8]. Bioinformatic searches using the seed sequences of miRNAs reveal a large numbers of potential viral miRNA targets. However, so far only a few of them have been validated to be true targets of viral miRNAs [4]. Accordingly, the roles of those viral miRNAs in KSHV biology and pathogenesis remain largely unknown.
One KSHV miRNA, miRNA K12-1 (miR-K12-1), was reported to directly target and repress the expression of the cellular protein IκBα, the primary inhibitor of the transcription factor nuclear factor-κb (NF-κB) [9, 10]. NF-κB plays a causative role in the formation and therapeutic resistance of several tumor types, including those induced by KSHV [10–12]. Whereas its transient or low activation may contribute to KSHV lytic replication, constitutive hyper-activation of NF-κB plays a crucial role in the latency of KSHV as well as in the formation and maintenance of KSHV-associated tumors. Inhibition of NF-κB activation by chemical inhibitors or short hairpin RNAs (shRNAs) against the NF-κB activating kinase IKK induces reactivation of KSHV from latency, prevents human endothelial cell transformation by KSHV, and reverses malignant phenotype and tumorigenicity of KSHV-associated PEL cells both in vitro and in vivo [13–17]. Similarly, NF-κB specific inhibition by over-expressing a functional defective mutant of RelA (also known as p65), the prototypical member of the NF-κB family, or specific RelA shRNAs results in reactivation of latent KSHV and growth inhibition in PEL cells [5, 17].
The transcription factor signal transducer and activator of transcription 3 (STAT3) is another important cellular protein involved in the tumorigenesis of KSHV [5]. Like the inhibition of NF-κB, STAT3 inhibition by its dominant-negative mutant, pharmacological inhibitor or shRNAs results in death of KSHV-associated PEL cells [5, 18, 19]. However, it remains unknown whether and how these two key pro-tumorigenic factors crosstalk in KSHV tumorigenesis.
In this study, we demonstrate that miR-K12-1 functions as an oncogene by hijacking IκBα/NF-κB signaling to induce interleukin-6 (IL-6)-dependent activation of STAT3. We also demonstrate that the novel miR-K12-1/IκBα/NF-κB/IL-6/STAT3 oncogenic signaling pathway is an essential mechanism underlying KSHV tumorigenesis.
RESULTS
KSHV miR-K12-1 activates both NF-κB and STAT3 and functions as an oncogene
To investigate the significance of miR-K12-1 in KSHV oncogenicity, we first examined whether K12-1 represses the expression of IκBα in our system. In line with previous studies [9], we found that expression of miR-K12-1 repressed the expression of IκBα at both RNA and protein levels (Figure 1A). Following these studies, we examined whether miR-K12-1 induces nuclear translocation of RelA and NF-κB transcriptional activation. The role of IκBα is to sequester RelA and other NF-κB members in the cytoplasm, thereby preventing NF-κB transcriptional activation [10]. As expected, miR-K12-1 induced strong nuclear translocation of RelA and transcriptional activation of NF-κB (Figure 1B–1D).
Figure 1. KSHV miR-K12-1 induces NF-κB and STAT3 activation and cell transformation.
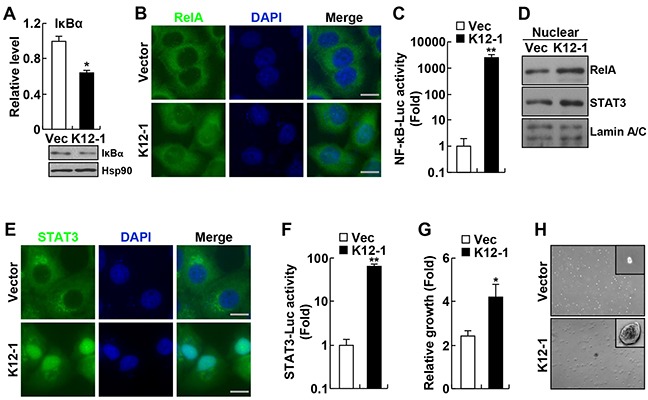
A. miR-K12-1 represses RNA and protein expression of IκBα. Real-time RT-PCR and immunoblotting analyses were performed to examine the IκBα RNA and protein levels, respectively, in Hela cells transfected with miR-K12-1 or an empty vector. B. miR-K12-1 induces RelA nuclear translocation. Immunofluorescence assay was performed to examine the nuclear translocation of RelA protein in Hela cells transfected with miR-K12-1 or an empty vector. Scale bar: 10 μm. C. miR-K12-1 induces transcriptional activation of NF-κB. Luciferase assay was performed to measure NF-κB transcriptional activity in Hela cells transfected with miR-K12-1 or an empty vector, together with NF-κB luciferase reporter. D. miR-K12-1 induces RelA and STAT3 nuclear expression. Nuclear fraction immunoblotting was performed to examine the nuclear expression of RelA and STAT3 proteins in Hela cells transfected with miR-K12-1 or an empty vector. E. miR-K12-1 induces STAT3 nuclear translocation. Immunofluorescence assay was performed to examine the nuclear translocation of STAT3 protein in Hela cells transfected with miR-K12-1 or an empty vector. Scale bar: 10 μm. F. miR-K12-1 induces transcriptional activation of STAT3. Luciferase assay was performed to measure STAT3 transcriptional activity in Hela cells transfected with miR-K12-1 or an empty vector, together with STAT3 luciferase reporter. G. miR-K12-1 promotes cell growth. Cell growth rates were determined for Rat-1 cells stably expressing miR-K12-1 or an empty vector. H. miR-K12-1 induces cell transformation. Soft agar colony formation assay was performed for Rat-1 cells stably expressing miR-K12-1 or an empty vector.
Interestingly, we found that miR-K12-1 is also a strong activator of STAT3, because miR-K12-1 induced strong nuclear translocation and transcriptional activation of STAT3 (Figure 1D–1F). Like NF-κB, STAT3 is an abundant latent cytoplasmic transcription factor that can translocate into the nucleus to induce gene transcription after being activated, although they are regulated by entirely different signaling mechanisms [20]. Also like NF-κB, STAT3 is often constitutively activated in many human cancers, including those induced by KSHV [5, 21].
Given the tumor-promoting role of both NF-κB and STAT3 activations, we hypothesized that miR-K12-1 is a viral oncogene that was not identified previously. In support of this hypothesis, we found that expression of miR-K12-1 significantly increased cell growth even in normal culture condition (Figure 1G). To directly test the hypothesis, we examined whether miR-K12-1 induces colony formation of the Rat-1 fibroblast cells in soft agar, a model system that has been widely used to test the oncogenic potential of candidate genes, particularly viral genes [22–25]. As expected, Rat-1 cells stably expressing an empty control vector did not form colonies in soft agar (Figure 1H, and Supplementary Figure S1). However, expression of miR-K12-1 alone was sufficient to induce Rat-1 cells to form colonies in soft agar. These data suggest that miR-K12-1 indeed has neoplastic transforming ability and functions as an oncogene. These data also suggest that the oncogenic potential of miR-K12-1 is associated with its ability in activating NF-κB and STAT3.
KSHV miR-K12-1 activates STAT3 indirectly through targeting the IκBα/NF-κB/IL-6 signaling pathway
To investigate the molecular mechanisms by which miR-K12-1 induces STAT3 activation, we analyzed whether miR-K12-1 targets the negative regulators of STAT3, such as suppressor of cytokine signaling 3 (SOCS3), protein inhibitor of activated STAT3 (PIAS3) or protein tyrosine phosphatases (PTPs). However, our bioinformatics analysis using miRNA target prediction software suggested that miR-K12-1 did not target those STAT3 negative regulators or any other known inhibitors of STAT3 signaling (data not shown), suggesting that miR-K12-1 activates STAT3 indirectly through activating STAT3 upstream activator(s) by repressing their inhibitor(s).
In this regard, we examined whether miR-K12-1 induces expression of the cytokine IL-6, a transcriptional target of NF-κB that functions as a potent STAT3 activator [26]. Indeed, miR-K12-1 induced IL-6 expression (Figure 2A). We then compared the activation dynamics of NF-κB and STAT3 induced by miR-K12-1, and found a four-hour delay in STAT3 activation in comparison to NF-κB activation (Figure 2B). These data suggested that the miR-K12-1 oncogene induces STAT3 activation through activating NF-κB and NF-κB-dependent IL-6 induction.
Figure 2. KSHV miR-K12-1 activates STAT3 via activating IκBα/NF-κB/IL-6 signaling.
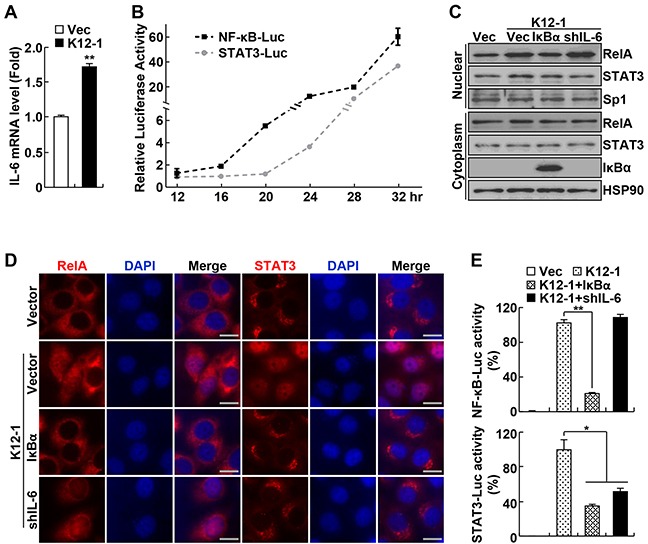
A. miR-K12-1 induces IL-6 expression. Real-time RT-PCR analysis was performed to examine the RNA levels of IL-6 in Hela cells transfected with miR-K12-1 or an empty vector. B. miR-K12-1 induces a delayed STAT3 activation compared to NF-κB activation. The dynamics of NF-κB and STAT3 transcriptional activation induced by miR-K12-1 were determined using luciferase assay for Hela cells co-transfected with miR-K12-1 and NF-κB or STAT3 luciferase reporter, respectively. C. IκBα expression or IL-6 knockdown inhibits STAT3 nuclear expression induced by miR-K12-1. Subcellular fraction immunoblotting was performed to examine the nuclear expression of RelA and STAT3 proteins in Hela cells transfected with the indicated constructs. D. IκBα expression or IL-6 knockdown prevents STAT3 nuclear translocation induced by miR-K12-1. Immunofluorescence assay was performed to examine the nuclear translocation of RelA and STAT3 proteins in Hela cells transfected with the indicated constructs. Scale bar: 10 μm. E. IκBα expression or IL-6 knockdown represses STAT3 transcriptional activation induced by miR-K12-1. Luciferase assay was performed to measure NF-κB and STAT3 transcriptional activity in Hela cells transfected with the indicated constructs together with NF-κB or STAT3 luciferase reporter, respectively.
To validate this, we examined the effects of IκBα expression and IL-6 knockdown on miR-K12-1 activation of NF-κB and STAT3. Expression of ectopic IκBα blocked miR-K12-1-induced nuclear translocation and transcriptional activation of both NF-κB RelA and STAT3 (Figure 2C–2E). On the other hand, shRNA knockdown of IL-6 only prevented STAT3 activation but had no effect on NF-κB activation by miR-K12-1. It should be pointed out that IκBα expression or IL-6 knockdown had no effect on the expression of transfected miR-K12-1 (Supplementary Figure S2). Consistent with its role in suppressing STAT3 activation by IL-6, SOCS3 over-expression inhibited STAT3 but not NF-κB activation induced by miR-K12-1 (Supplementary Figure S3). These data together indicate that the viral oncogene miR-K12-1 activates STAT3 through targeting IκBα and subsequent NF-κB activation and IL-6 induction.
The oncogenicity of miR-K12-1 requires IκBα repression and NF-κB/IL-6/STAT3 activation
To examine whether miR-K12-1 exerts its oncogenicity through the IκBα/NF-κB/IL-6/STAT3 signaling pathway, we generated miR-K12-1 stable Rat-1 cell lines that also express ectopic IκBα or shRNAs specifically against IL-6 or STAT3. Of note, expression of ectopic IκBα or knockdown of IL-6 or STAT3 did not affect the expression of the viral miRNA, because its expression levels in these stable cell lines were comparable (Supplementary Figure S4). Expression of IκBα or knockdown of IL-6 or STAT3 had no obvious effect on miR-K12-1-driven cell growth when 10% fetal bovine serum (FBS) was provided in cell culture medium (Figure 3A). However, the cell growth driven by miR-K12-1 was almost completely inhibited by IκBα expression when the supplemented FBS was decreased to 5% (Figure 3B). Similar inhibitions were also seen by IL-6 or STAT3 knockdown. More importantly, the neoplastic transforming ability of miR-K12-1 was also completely inhibited by IκBα expression, IL-6 knockdown or STAT3 knockdown (Figure 3C, and Supplementary Figure S5). These data suggest that miR-K12-1 functions as an oncogene via targeting the IκBα/NF-κB/IL-6/STAT3 signaling pathway.
Figure 3. The oncogenicity of KSHV miR-K12-1 depends on the activation of IκBα/NF-κB/IL-6/STAT3 signaling.
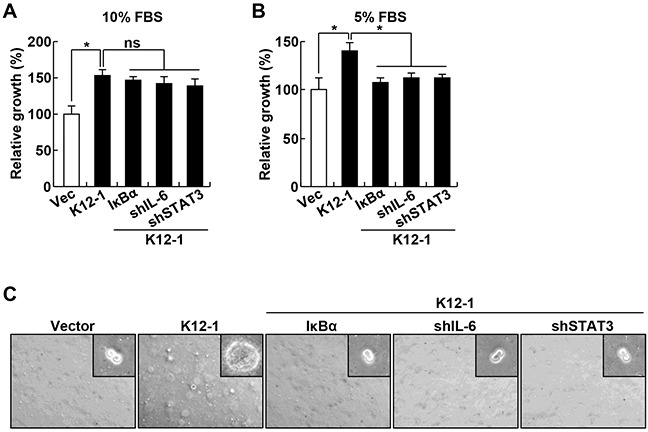
A. IκBα expression, IL-6 knockdown, or STAT3 knockdown has no obvious effect on miR-K12-1-expressing cells cultured in medium containing 10% FBS. Cell growth rates were determined for Rat-1 cells stably expressing an empty vector, miR-K12-1, or miR-K12-1 together with IκBα, IL-6 shRNA or STAT3 shRNA cultured in medium containing 10% FBS. B. IκBα expression, IL-6 knockdown, or STAT3 knockdown suppresses miR-K12-1-driven cell growth when the supplemented FBS is decreased to 5%. Cell growth rates were determined for Rat-1 cells stably expressing an empty vector, miR-K12-1, or miR-K12-1 together with IκBα, IL-6 shRNA or STAT3 shRNA cultured in medium containing 5% FBS. C. IκBα expression, IL-6 knockdown, or STAT3 knockdown suppresses miR-K12-1-driven cell transformation. Soft agar colony formation assay was performed for Rat-1 cells stably expressing an empty vector, miR-K12-1, or miR-K12-1 together with IκBα, IL-6 shRNA or STAT3 shRNA.
Activation of miR-K12-1/NF-κB/IL-6/STAT3 signaling is required for KSHV tumorigenesis
We next would like to investigate the pathophysiological relevance of the miR-K12-1/NF-κB/IL-6/STAT3 signaling pathway in KSHV tumorigenesis. We generated KSHV+ BCBL-1 PEL cell lines stably expressing shRNAs specifically against RelA, IL-6 or STAT3. Consistent with our recent studies showing high activations of NF-κB and STAT3 in various PEL cell lines [5], our nuclear fraction-immunoblotting (IB) and immunofluorescence assays indicated a strong nuclear expression of RelA and STAT3 in BCBL-1 cells stably expressing scrambled control shRNAs (Figure 4A, and Supplementary Figure S6). Knockdown of RelA significantly inhibited the nuclear expression of STAT3 in BCBL-1 cells. Of note, RelA knockdown had no effect on the STAT3 expression (Figure 4B). As expected, the expression of IL-6 in BCBL-1 cells was largely repressed by RelA knockdown. Interestingly, IL-6 knockdown, like the RelA knockdown, also inhibited STAT3 nuclear expression hence STAT3 activation in BCBL-1 cells (Figure 4C). Consistent with the fact that IL-6 only functions as an effector but not an activator of NF-κB, IL-6 knockdown did not affect RelA nuclear expression in BCBL-1 cells.
Figure 4. KSHV oncogenicity involves activation of miR-K12-1/NF-κB/IL-6/STAT3 signaling.
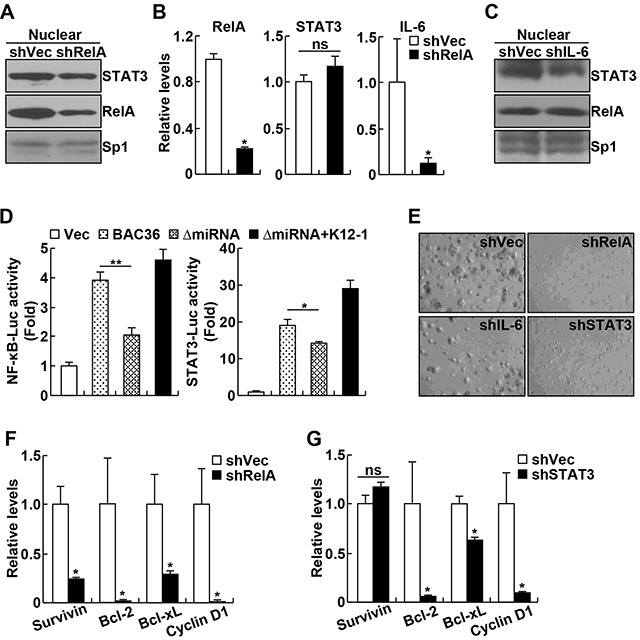
A. RelA knockdown decreases STAT3 nuclear expression in BCBL-1 cells. Nuclear fraction immunoblotting assay was performed to examine the nuclear expression of RelA and STAT3 proteins in BCBL-1 cells stably expressing RelA shRNA or an empty vector. B. RelA knockdown suppresses IL-6 but not STAT3 expression in BCBL-1 cells. Real time PCR analysis was performed to detect RelA, IL-6 and STAT3 RNA expression levels in BCBL-1 cells stably expressing RelA shRNA or an empty vector. C. IL-6 knockdown blocks STAT3 nuclear expression in BCBL-1 cells. Nuclear fraction immunoblotting assay was performed to examine the nuclear expression of RelA and STAT3 proteins in BCBL-1 cells stably expressing IL-6 shRNA or an empty vector. D. KSHV requires miR-K12-1 to induce NF-κB and STAT3 activation. Luciferase reporter activity assay was performed to examine the activation of NF-κB and STAT3 in 293FT cells with an empty vector, KSHVBac36, KSHVBac36 ΔmiRNA, or KSHVBac36 ΔmiRNA plus miR-K12-1. E. Knockdown of RelA, IL-6 or STAT3 suppresses the oncogenicty of BCBL-1 cells. Soft agar colony formation assay was performed for BCBL-1 cells stably expressing RelA shRNA, IL-6 shRNA, STAT3 shRNA, or an empty vector. F. RelA knockdown suppresses expression of cell survival and proliferation genes in BCBL-1 cells. Real time PCR analysis was performed to detect RNA expression levels of the indicated genes in BCBL-1 cells stably expressing RelA shRNA or an empty vector. G. STAT3 knockdown suppresses expression of cell survival and proliferation genes in BCBL-1 cells. Real time PCR analysis was performed to detect RNA expression levels of the indicated genes in BCBL-1 cells stably expressing STAT3 shRNA or an empty vector.
To repress miR-K12-1 expression in BCBL-1 cells, we initially used lock-nucleic acid (LNA)-based miR-K12-1 suppressor [9]. However, miR-K12-1 could not be repressed in BCBL-1 cells or other PEL cell lines to a detectable level, because of extremely low transfection of these cells. Thus, we changed the strategy and compared NF-κB and STAT3 activations induced by KSHV Bacterial Artificial Chromosome Clone (KSHVBac36) and its mutant (ΔmiRNA) in which miR-K12-1 and other 13 miRNAs are deleted [9]. As expected, expression of wildtype KSHVBac36 induced activation of both NF-κB and STAT3 (Figure 4D). However, the ΔmiRNA mutant lost the ability to activate either NF-κB or STAT3. More interestingly, re-expression of miR-K12-1 could efficiently rescue the ability of the ΔmiRNA mutant in activating NF-κB and STAT3. These data indicate that miR-K12-1/NF-κB/IL-6/STAT3 signaling is activated under KSHV pathogenic condition.
To further validate and also expand these studies, we also investigated the significance of the NF-κB/IL-6/STAT3 signaling pathway in KSHV oncogenesis. To this end, we examined the effect of RelA, IL-6 or STAT3 knockdown on the colony formation of BCBL-1 cells. In line with our recent studies [5], BCBL-1 cells expressing scrambled control shRNAs showed high oncogenicity, as evidenced by their high colony formation ability in soft agar (Figure 4E, and Supplementary Figure S7). However, the colony formation ability of BCBL-1 cells was significantly suppressed by RelA, IL-6 or STAT3 knockdown. In fact, knockdown of RelA, IL-6 or STAT3 was able to block the outgrowth of BCBL-1 cells in normal culture (Supplementary Figure S8). Highly consistent with these data, knockdown of RelA or STAT3 led to significant decreases in the expression of cell survival and proliferation genes, including Bcl-2, Bcl-xL, Survivin and/or Cyclin D1 (Figure 4F–4G). It should be pointed out that although the expressions of Bcl-2, Bcl-xL and Cyclin D1 in BCBL-1 cells were repressed by both RelA and STAT3 knockdowns, which have similar knockdown efficiency (Figure 4B and data not shown), the repression caused by RelA knockdown was stronger. Furthermore, Survivin, like IL-6, was only repressed by RelA but not STAT3 knockdown. These data suggest that activation of NF-κB/IL-6/STAT3 signaling is an important mechanism underlying KSHV oncogenesis. These data also suggest that in addition to activating STAT3 for gene induction, NF-κB also directly induces gene expression contributing to KSHV tumorigenesis.
DISCUSSION
KSHV encodes over 115 genes including 25 miRNAs [6]. However, the roles of most of these genes, particularly the viral miRNAs, in KSHV biology and pathogenesis remain unclear. The data presented here demonstrate miR-K12-1 as the first viral miRNA that possesses the oncogenic ability, and identify miR-K12-1/IκBα/NF-κB/IL-6/STAT3 as a novel oncogenic signaling pathway important for KSHV tumorigenesis (Supplementary Figure S9). These data also provide the first evidence showing that IL-6/STAT3 signaling acts as an essential downstream mediator of NF-κB oncogenic actions. These findings significantly improve our understanding of KSHV pathogenesis and oncogenic interaction between NF-κB and STAT3. These findings also set up a solid basis for us to target the miR-K12-1/IκBα/NF-κB/IL-6/STAT3 oncogenic signaling pathway for the prevention and treatment of KSHV-mediated cancers, for which no effective therapies exist currently.
Since their original cloning and identification in 2005, a role of KSHV miRNAs in viral pathogenesis has been expected. The miRNAs form a cluster that is located within the genomic region associated with KSHV latency and share the common promoter with several well-known latent genes, including the viral oncoprotein vFLIP [27–31]. Indeed, a virus mutant in which the viral miRNA cluster is deleted not only loses the transforming ability, but actually induces cell cycle arrest and apoptosis [32]. On the other hand, expression of the miRNA cluster decreases mitochondria biogenesis and induces aerobic glycolysis, a metabolic hallmark of cancer cells [33]. Interestingly, the metabolic shift is important for viral latency maintenance and provides a growth advantage. Moreover, several viral miRNAs have been shown to stabilize viral latency or to promote cell growth, migration and/or invasion [34–45]. However, whether any of viral miRNAs function as an oncogene has yet to be validated. Moreover, downstream functional targets of viral miRNAs remain largely unknown. Although recent comprehensive target analyses reveal nearly 1000 potential viral miRNA target genes, so far only a few of them have been validated to be true targets of viral miRNAs [4, 46–48]. Our studies not only validate IκBα as a bona fide target gene of miR-K12-1, but also demonstrate that miR-12-1 functions as an oncogene by targeting IκBα. In addition to IκBα, p21 and caspase-3 have also been suggested to be target genes of miR-K12-1 [49, 50]. Although it is interesting to test whether this function of miR-K12-1 is required for its oncogenic ability, our studies suggest that targeting these two and any other potential genes by miR-K12-1 is not sufficient to induce cell transformation.
Activation of NF-κB/IL-6/STAT3 signaling by miR-K12-1 is a critical mechanism underlying KSHV tumorigenesis, however, KSHV also uses other mechanisms to facilitate the activation of this pro-oncogenic signaling pathway. For example, the viral oncoproein vFLIP encoded by KSHV induces proteasomal degradation of IκBα, complementing miR-K12-1 in NF-κB activation [51]. It will be of interest to examine whether vFLIP is able to activate STAT3, and if so, whether through NF-κB and NF-κB-induced IL-6. In addition to inducing IκBα protein degradation to activate NF-κB, which is called ‘canonical NF-κB activation’, vFLIP also induces activation of the non-canonical NF-κB by inducing processing of p100, atypical NF-κB inhibitor that also serves as the precursor of the mature form of an important NF-κB member NF-κB2 p52 [52, 53]. Both NF-κB pathways have been linked to various cellular and viral tumorigenesis and often crosstalk with each other and other signaling pathways important to cancer pathogenesis, such as autophagy [25, 54–57]. It thus will be interesting to examine whether and how these important signaling pathways interact with each other in the tumorigenesis mediated by miRNA-K12-1, vFLIP or KSHV.
MATERIALS AND METHODS
Plasmids and reagents
KSHVBac36, KSHVBac36 ΔmiRNA, and KSHV miR-K12-1 expressing vector were kind gifts from Drs. Paul Lieberman and Rolf Renne. Vectors expressing shRNA against IL-6, RelA, or STAT3 (target sites are listed in Supplementary Table S1), IκBα, SOCS3, NF-κB–driven firefly luciferase reporter, and TK-driven Renilla luciferase reporter have been described before [5, 58]. The Sp1, Lamin A/C, and Hsp90 antibodies as well as the secondary antibodies were from Santa Cruz Biotechnology (Dallas, TX). The RelA and STAT3 antibodies were from Cell Signaling Technology (Danvers, MA). The anti-Myc antibody was generated from the 9E10 hybridoma as described [59].
Cell culture
Human PEL cell line BCBL-1 cells were maintained in suspension in RPMI 1640 medium supplemented with 10% FBS. Human cell lines Hela and 293FT and rat fiborblast Rat-1 cells were maintained in DMEM medium supplemented with 10% FBS.
Quantitative polymerase chain reaction (qPCR) analysis
Cells were subjected to RNA extraction, RNA reverse transcription and real-time PCR as described [60]. The expression levels of various genes were normalized to that of GAPDH. Primer pairs used for qPCR are listed in Supplementary Table S2.
Generation of stable transfectants
Rat-1 cells expressing an empty vector, miR-K12-1, or miR-K12-1 together with IκBα, IL-6 shRNA or STAT3 shRNA, and BCBL-1 cells expressing an empty vector, RelA shRNA, IL-6 shRNA, or STAT3 shRNA were generated as described before [5, 61].
Soft agar assays
Rat-1 (2.5 × 105) or BCBL-1 (5 × 104) cells suspended in culture medium containing 0.6% SeaPlaque low melting agarose were plated on the top of 1% agarose in culture medium as described before [5, 22–25]. Colonies in soft agar were counted 12 days after plating. Each determination was repeated in at least 3 independent experiments.
Immunofluorescence microscopy
Cells were fixed, permeabilized, and subsequently incubated with the indicated primary antibodies, followed by fluorescent dye-conjugated secondary antibodies [5, 62]. The stained proteins and their subcellular localizations were detected using a fluorescence microscope. The cells were also counterstained with DAPI for nuclear staining.
Immunoblotting (IB) analysis
Cytosolic and nuclear extracts or whole cell lysates were prepared as described previously [63, 64]. The whole-cell, cytosolic or nuclear extracts were then subjected to SDS-PAGE and immunoblotting using the indicated antibodies. The purity of the obtained cell nuclear fractions was confirmed by the detection of Sp1 or Lamin A/C (nuclear fraction) but no Hsp90 (cytoplasm) in IB assays.
Luciferase gene reporter assays
The indicated cell lines were transfected with NF-κB–driven firefly luciferase reporter and TK-driven Renilla luciferase reporter. At 24 h posttransfection, dual luciferase activities were measured as we described previously [58].
Statistical analysis
Data were reported as mean ± standard deviation (SD). As described previously [65], Student's t test (two tailed) was used to assess significance of differences between two groups, and p values < 0.05 and 0.01 were considered statistically significant and highly statistically significant, and indicated by * and **, respectively.
SUPPLEMENTARY FIGURES AND TABLES
Acknowledgments
The authors thank Gutian Xiao for his critical reading and helpful suggestions, Rolf Renne and Paul Lieberman for providing KSHV miR-K12-1.
Footnotes
CONFLICTS OF INTEREST
The authors declare no conflict of interest.
GRANT SUPPORT
This study was supported in part by the National Institute of Health (NIH)/National Cancer Institute (NCI) grants R01CA172090, R21CA189703, P30CA047904-25S for AIDS-associated malignancies, and University of Pittsburgh Competitive Medical Research Fund.
REFERENCES
- 1.Chang Y, Cesarman E, Pessin MS, Lee F, Culpepper J, Knowles DM, Moore PS. Identification of herpesvirus-like DNA sequences in AIDS-associated Kaposi's sarcoma. Science. 1994;265:1865–1869. doi: 10.1126/science.7997879. [DOI] [PubMed] [Google Scholar]
- 2.Cesarman E, Chang Y, Moore PS, Said JW, Knowles DM. Kaposi's sarcoma-associated herpesvirus-like DNA sequences in AIDS-related body-cavity-based lymphomas. N Engl J Med. 1995;332:1186–1191. doi: 10.1056/NEJM199505043321802. [DOI] [PubMed] [Google Scholar]
- 3.Soulier J, Grollet L, Oksenhendler E, Cacoub P, Cazals-Hatem D, Babinet P, d'Agay MF, Clauvel JP, Raphael M, Degos L, et al. Kaposi's sarcoma-associated herpesvirus-like DNA sequences in multicentric Castleman's disease. Blood. 1995;86:1276–1280. [PubMed] [Google Scholar]
- 4.Cousins E, Nicholas J. Molecular biology of human herpesvirus 8: novel functions and virus-host interactions implicated in viral pathogenesis and replication. Recent Results Cancer Res. 2014;193:227–268. doi: 10.1007/978-3-642-38965-8_13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Sun F, Xiao Y, Qu Z. Oncovirus Kaposi sarcoma herpesvirus (KSHV) represses tumor suppressor PDLIM2 to persistently activate nuclear factor kB (NF-κB) and STAT3 transcription factors for tumorigenesis and tumor maintenance. J Biol Chem. 2015;290:7362–7368. doi: 10.1074/jbc.C115.637918. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Zhu Y, Huang Y, Jung JU, Lu C, Gao SJ. Viral miRNA targeting of bicistronic and polycistronic transcripts. Curr Opin Virol. 2014;7:66–72. doi: 10.1016/j.coviro.2014.04.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Ambros V. The functions of animal microRNAs. Nature. 2004;431:350–355. doi: 10.1038/nature02871. [DOI] [PubMed] [Google Scholar]
- 8.Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116:281–297. doi: 10.1016/s0092-8674(04)00045-5. [DOI] [PubMed] [Google Scholar]
- 9.Lei X, Bai Z, Ye F, Xie J, Kim CG, Huang Y, Gao SJ. Regulation of NF-κB inhibitor IκBα and viral replication by a KSHV microRNA. Nat Cell Biol. 2010;12:193–199. doi: 10.1038/ncb2019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Xiao G, Fu J. NF-κB and cancer: a paradigm of Yin-Yang. Am J Cancer Res. 2011;1:192–221. [PMC free article] [PubMed] [Google Scholar]
- 11.Qu Z, Xiao G. Human T-cell lymphotropic virus: a model of NF-κB-associated tumorigenesis. Viruses. 2011;3:714–749. doi: 10.3390/v3060714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.de Oliveira DE, Ballon G, Cesarman E. NF-κB signaling modulation by EBV and KSHV. Trends Microbiol. 2010;18:248–257. doi: 10.1016/j.tim.2010.04.001. [DOI] [PubMed] [Google Scholar]
- 13.Keller SA, Schattner EJ, Cesarman E. Inhibition of NF-κB induces apoptosis of KSHV-infected primary effusion lymphoma cells. Blood. 2000;96:2537–2542. [PubMed] [Google Scholar]
- 14.Brown HJ, Song MJ, Deng H, Wu TT, Cheng G, Sun R. NF-κB inhibits γherpesvirus lytic replication. J Virol. 2003;77:8532–8540. doi: 10.1128/JVI.77.15.8532-8540.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Keller SA, Hernandez-Hopkins D, Vider J, Ponomarev V, Hyjek E, Schattner EJ, Cesarman E. NF-κB is essential for the progression of KSHV- and EBV-infected lymphomas in vivo. Blood. 2006;107:3295–3302. doi: 10.1182/blood-2005-07-2730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Matta H, Surabhi RM, Zhao J, Punj V, Sun Q, Schamus S, Mazzacurati L, Chaudhary PM. Induction of spindle cell morphology in human vascular endothelial cells by human herpesvirus 8-encoded viral FLICE inhibitory protein K13. Oncogene. 2007;26:1656–1660. doi: 10.1038/sj.onc.1209931. [DOI] [PubMed] [Google Scholar]
- 17.He Z, Zhao J, Zhang J, Jung JU, Feng P. NF-κB activation coordinated by IKKβ and IKKε enables latent infection of Kaposi's sarcoma-associated herpesvirus. J Virol. 2014;88:444–455. doi: 10.1128/JVI.01716-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Aoki Y, Feldman GM, Tosato G. Inhibition of STAT3 signaling induces apoptosis and decreases survivin expression in primary effusion lymphoma. Blood. 2003;101:1535–1542. doi: 10.1182/blood-2002-07-2130. [DOI] [PubMed] [Google Scholar]
- 19.Cousins E, Nicholas J. Role of human herpesvirus 8 interleukin-6-activated gp130 signal transducer in primary effusion lymphoma cell growth and viability. J Virol. 2013;87:10816–10827. doi: 10.1128/JVI.02047-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Grivennikov SI, Karin M. Dangerous liaisons: STAT3 and NF-κB collaboration and crosstalk in cancer. Cytokine Growth Factor Rev. 2010;21:11–19. doi: 10.1016/j.cytogfr.2009.11.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Zhou J, Qu Z, Yan S, Sun F, Whitsett JA, Shapiro SD, Xiao G. Differential roles of STAT3 in the initiation and growth of lung cancer. Oncogene. 2015;34:3804–3814. doi: 10.1038/onc.2014.318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Qing G, Qu Z, Xiao G. Endoproteolytic processing of C-terminally truncated NF-κB2 precursors at kB-containing promoters. Proc Natl Acad Sci U S A. 2007;104:5324–5329. doi: 10.1073/pnas.0609914104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Yan P, Fu J, Qu Z, Li S, Tanaka T, Grusby MJ, Xiao G. PDLIM2 suppresses HTLV-I Tax-mediated tumorigenesis by targeting Tax into the nuclear matrix for proteasomal degradation. Blood. 2009;113:4370–4380. doi: 10.1182/blood-2008-10-185660. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Fu J, Yan P, Li S, Qu Z, Xiao G. Molecular determinants of PDLIM2 in suppressing HTLV-I Tax-mediated tumorigenesis. Oncogene. 2010;29:6499–6507. doi: 10.1038/onc.2010.374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Fu J, Qu Z, Yan P, Ishikawa C, Aqeilan RI, Rabson AB, Xiao G. The tumor suppressor gene WWOX links the canonical and noncanonical NF-κB pathways in HTLV-I Tax-mediated tumorigenesis. Blood. 2011;117:1652–1661. doi: 10.1182/blood-2010-08-303073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Qu Z, Sun F, Zhou J, Li L, Shapiro SD, Xiao G. Interleukin-6 prevents the initiation but enhances the progression of lung cancer. Cancer Res. 2015;75:3209–3215. doi: 10.1158/0008-5472.CAN-14-3042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Samols MA, Hu J, Skalsky RL, Renne R. Cloning and identification of a microRNA cluster within the latency-associated region of Kaposi's sarcoma-associated herpesvirus. J Virol. 2005;79:9301–9305. doi: 10.1128/JVI.79.14.9301-9305.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Cai X, Lu S, Zhang Z, Gonzalez CM, Damania B, Cullen BR. Kaposi′s sarcoma-associated herpesvirus expresses an array of viral microRNAs in latently infected cells. Proc Natl Acad Sci U S A. 2005;102:5570–5575. doi: 10.1073/pnas.0408192102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Pearce M, Matsumura S, Wilson AC. Transcripts encoding K12, v-FLIP, v-cyclin, and the microRNA cluster of Kaposi's sarcoma-associated herpesvirus originate from a common promoter. J Virol. 2005;79:14457–14464. doi: 10.1128/JVI.79.22.14457-14464.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Pfeffer S, Sewer A, Lagos-Quintana M, Sheridan R, Sander C, Grasser FA, van Dyk LF, Ho CK, Shuman S, Chien M, et al. Identification of microRNAs of the herpesvirus family. Nat Methods. 2005;2:269–276. doi: 10.1038/nmeth746. [DOI] [PubMed] [Google Scholar]
- 31.Grundhoff A, Sullivan CS, Ganem D. A combined computational and microarray-based approach identifies novel microRNAs encoded by human gamma-herpesviruses. RNA. 2006;12:733–750. doi: 10.1261/rna.2326106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Moody R, Zhu Y, Huang Y, Cui X, Jones T, Bedolla R, Lei X, Bai Z, Gao SJ. KSHV microRNAs mediate cellular transformation and tumorigenesis by redundantly targeting cell growth and survival pathways. PLoS Pathog. 2013;9:e1003857. doi: 10.1371/journal.ppat.1003857. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Yogev O, Lagos D, Enver T, Boshoff C. Kaposi's sarcoma herpesvirus microRNAs induce metabolic transformation of infected cells. PLoS Pathog. 2014;10:e1004400. doi: 10.1371/journal.ppat.1004400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Bellare P, Ganem D. Regulation of KSHV lytic switch protein expression by a virus-encoded microRNA: an evolutionary adaptation that fine-tunes lytic reactivation. Cell Host Microbe. 2009;6:570–575. doi: 10.1016/j.chom.2009.11.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Lu F, Stedman W, Yousef M, Renne R, Lieberman PM. Epigenetic regulation of Kaposi's sarcoma-associated herpesvirus latency by virus-encoded microRNAs that target Rta and the cellular Rbl2-DNMT pathway. J Virol. 2010;84:2697–2706. doi: 10.1128/JVI.01997-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Lin X, Liang D, He Z, Deng Q, Robertson ES, Lan K. miR-K12-7-5p encoded by Kaposi's sarcoma-associated herpesvirus stabilizes the latent state by targeting viral ORF50/RTA. PLoS One. 2011;6:e16224. doi: 10.1371/journal.pone.0016224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Lin HR, Ganem D. Viral microRNA target allows insight into the role of translation in governing microRNA target accessibility. Proc Natl Acad Sci U S A. 2011;108:5148–5153. doi: 10.1073/pnas.1102033108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Dahlke C, Maul K, Christalla T, Walz N, Schult P, Stocking C, Grundhoff A. A microRNA encoded by Kaposi sarcoma-associated herpesvirus promotes B-cell expansion in vivo. PLoS One. 2012;7:e49435. doi: 10.1371/journal.pone.0049435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Hu M, Wang C, Li W, Lu W, Bai Z, Qin D, Yan Q, Zhu J, Krueger BJ, Renne R, Gao SJ, Lu C. A KSHV microRNA Directly Targets G Protein-Coupled Receptor Kinase 2 to Promote the Migration and Invasion of Endothelial Cells by Inducing CXCR2 and Activating AKT Signaling. PLoS Pathog. 2015;11:e1005171. doi: 10.1371/journal.ppat.1005171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Forte E, Raja AN, Shamulailatpam P, Manzano M, Schipma MJ, Casey JL, Gottwein E. MicroRNA-mediated transformation by the Kaposi's sarcoma-associated herpesvirus Kaposin locus. J Virol. 2015;89:2333–2341. doi: 10.1128/JVI.03317-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Lei X, Zhu Y, Jones T, Bai Z, Huang Y, Gao SJ. A Kaposi's sarcoma-associated herpesvirus microRNA and its variants target the transforming growth factor β pathway to promote cell survival. J Virol. 2012;86:11698–11711. doi: 10.1128/JVI.06855-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Liu Y, Sun R, Lin X, Liang D, Deng Q, Lan K. Kaposi's sarcoma-associated herpesvirus-encoded microRNA miR-K12-11 attenuates transforming growth factor beta signaling through suppression of SMAD5. J Virol. 2012;86:1372–1381. doi: 10.1128/JVI.06245-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Boss IW, Nadeau PE, Abbott JR, Yang Y, Mergia A, Renne R. A Kaposi's sarcoma-associated herpesvirus-encoded ortholog of microRNA miR-155 induces human splenic B-cell expansion in NOD/LtSz-scid IL2Rγnull mice. J Virol. 2011;85:9877–9886. doi: 10.1128/JVI.05558-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Gottwein E, Mukherjee N, Sachse C, Frenzel C, Majoros WH, Chi JT, Braich R, Manoharan M, Soutschek J, Ohler U, Cullen BR. A viral microRNA functions as an orthologue of cellular miR-155. Nature. 2007;450:1096–1099. doi: 10.1038/nature05992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Qin Z, Dai L, Defee M, Findlay VJ, Watson DK, Toole BP, Cameron J, Peruzzi F, Kirkwood K, Parsons C. Kaposi's sarcoma-associated herpesvirus suppression of DUSP1 facilitates cellular pathogenesis following de novo infection. J Virol. 2013;87:621–635. doi: 10.1128/JVI.01441-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Bai Z, Huang Y, Li W, Zhu Y, Jung JU, Lu C, Gao SJ. Genomewide mapping and screening of Kaposi's sarcoma-associated herpesvirus (KSHV) 3′ untranslated regions identify bicistronic and polycistronic viral transcripts as frequent targets of KSHV microRNAs. J Virol. 2014;88:377–392. doi: 10.1128/JVI.02689-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Gottwein E, Corcoran DL, Mukherjee N, Skalsky RL, Hafner M, Nusbaum JD, Shamulailatpam P, Love CL, Dave SS, Tuschl T, et al. Viral microRNA targetome of KSHV-infected primary effusion lymphoma cell lines. Cell Host Microbe. 2011;10:515–526. doi: 10.1016/j.chom.2011.09.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Haecker I, Gay LA, Yang Y, Hu J, Morse AM, McIntyre L, Renne R. Ago-HITS-CLIP expands understanding of Kaposi's sarcoma-associated herpesvirus miRNA function in primary effusion lymphomas. PLoS Pathog. 2012;8:54. doi: 10.1371/journal.ppat.1002884. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Gottwein E, Cullen BR. A human herpesvirus microRNA inhibits p21 expression and attenuates p21-mediated cell cycle arrest. J Virol. 2010;84:5229–5237. doi: 10.1128/JVI.00202-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Suffert G, Malterer G, Hausser J, Viiliäinen J, Fender A, Contrant M, Ivacevic T, Benes V, Gros F, Voinnet O, Zavolan M, Ojala PM, Haas JG, Pfeffer S. Kaposi's Sarcoma herpesvirus microRNAs target caspase 3 and regulate apoptosis. PLoS Pathog. 2011;7:e1002405. doi: 10.1371/journal.ppat.1002405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Liu L, Eby MT, Rathore N, Sinha SK, Kumar A, Chaudhary PM. The human herpes virus 8-encoded viral FLICE inhibitory protein physically associates with and persistently activates the IκB kinase complex. J Biol Chem. 2002;277:13745–13751. doi: 10.1074/jbc.M110480200. [DOI] [PubMed] [Google Scholar]
- 52.Matta H, Chaudhary PM. Activation of alternative NF-κB pathway by human herpes virus 8-encoded Fas-associated death domain-like IL-1 b-converting enzyme inhibitory protein (vFLIP) Proc Natl Acad Sci U S A. 2004;101:9399–9404. doi: 10.1073/pnas.0308016101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Xiao G, Rabson AB, Young W, Qing G, Qu Z. Alternative pathways of NF-κB activation: a double-edged sword in health and disease. Cytokine Growth Factor Rev. 2006;17:281–293. doi: 10.1016/j.cytogfr.2006.04.005. [DOI] [PubMed] [Google Scholar]
- 54.Qing G, Yan P, Xiao G. Hsp90 inhibition results in autophagy-mediated proteasome-independent degradation of IκB kinase (IKK) Cell Res. 2006;16:895–901. doi: 10.1038/sj.cr.7310109. [DOI] [PubMed] [Google Scholar]
- 55.Qing G, Yan P, Qu Z, Liu H, Xiao G. Hsp90 regulates processing of NF-κB2 p100 involving protection of NF-κB-inducing kinase (NIK) from autophagy-mediated degradation. Cell Res. 2007;17:520–530. doi: 10.1038/cr.2007.47. [DOI] [PubMed] [Google Scholar]
- 56.Yan P, Qing G, Qu Z, Wu CC, Rabson A, Xiao G. Targeting autophagic regulation of NF-κB in HTLV-I transformed cells by geldanamycin: implications for therapeutic interventions. Autophagy. 2007;3:600–603. doi: 10.4161/auto.4761. [DOI] [PubMed] [Google Scholar]
- 57.Xiao G. Autophagy and NF-κB: fight for fate. Cytokine Growth Factor Rev. 2007;18:233–243. doi: 10.1016/j.cytogfr.2007.04.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Xiao G, Cvijic ME, Fong A, Harhaj EW, Uhlik MT, Waterfield M, Sun SC. Retroviral oncoprotein Tax induces processing of NF-κB2/p100 in T cells: evidence for the involvement of IKKα. EMBO J. 2001;20:6805–6815. doi: 10.1093/emboj/20.23.6805. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Qing G, Qu Z, Xiao G. Regulation of NF-κB2 p100 processing by its cis-acting domain. J Biol Chem. 2005;280:18–27. doi: 10.1074/jbc.M406619200. [DOI] [PubMed] [Google Scholar]
- 60.Qu Z, Yan P, Fu J, Jiang J, Grusby MJ, Smithgall TE, Xiao G. DNA methylation-dependent repression of PDZ-LIM domain-containing protein 2 in colon cancer and its role as a potential therapeutic target. Cancer Res. 2010;70:1766–1772. doi: 10.1158/0008-5472.CAN-09-3263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Qu Z, Fu J, Yan P, Hu J, Cheng SY, Xiao G. Epigenetic repression of PDZ-LIM domain-containing protein 2: implications for the biology and treatment of breast cancer. J Biol Chem. 2010;285:11786–11792. doi: 10.1074/jbc.M109.086561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Qing G, Xiao G. Essential role of IκB kinase a in the constitutive processing of NF-κB2 p100. J Biol Chem. 2005;280:9765–9768. doi: 10.1074/jbc.C400502200. [DOI] [PubMed] [Google Scholar]
- 63.Qu Z., Qing G, Rabson A, Xiao G. Tax deregulation of NF-κB2 p100 processing involves both b-TrCP-dependent and -independent mechanisms. J Biol Chem. 2004;279:44563–44572. doi: 10.1074/jbc.M403689200. [DOI] [PubMed] [Google Scholar]
- 64.Qu Z, Sun D, Young W. Lithium promotes neural precursor cell proliferation: evidence for the involvement of the non-canonical GSK-3β-NF-AT signaling. Cell Biosci. 2011;1:18. doi: 10.1186/2045-3701-1-18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Sun F, Qu Z, Xiao Y, Zhou J, Burns TF, Stabile LP, Siegfried JM, Xiao G. NF-κB1 p105 suppresses lung tumorigenesis through the Tpl2 kinase but independently of its NF-κB function. Oncogene. 2015 Aug 24; doi: 10.1038/onc.2015.299. [Epub ahead of print] [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.