

Abstract
Heat shock protein 90 (Hsp90) is a molecular chaperone that assists in the maturation of a limited set of substrate proteins that are collectively referred to as clients. The majority of identified Hsp90 clients are involved in signal transduction including many steroid hormone receptors and kinases. A handful of Hsp90 clients can be classified as non-signal transduction proteins including telomerase, cystic fibrosis transmembrane conductance regulator (CFTR), and antigenic peptides destined for major histocompatibility complex (MHC). Because Hsp90 clients are causative agents in cancer and cystic fibrosis, research on Hsp90 has intensified in recent years. We review the historical path of Hsp90 research within each class of client (kinase, hormone receptor, and non-signal transduction clients) and highlight current areas of active investigation.
Keywords: Hsp90, chaperone, ATPase, kinase, steroid hormone receptor, signal transduction, telomerase, CFTR, antigen presentation
Introduction
Hsp90, as with most heat shock proteins, was initially identified from changes in transcriptional pattern (1) and relative translational rate (2) upon shifting cells to elevated temperature. Hsp90 is named for its induction in response to temperature stress as well as its molecular weight of about 90 kD. Elevated temperature challenges the ability of proteins to fold efficiently placing an elevated burden on cellular chaperones. In response to elevated temperature, and many other stressful conditions, eukaryotes induce expression of the heat shock factor 1 (HSF1) transcription factor that in turn up-regulates expression of chaperones including Hsp90 (3). Many eukaryotes including budding yeast and humans have two genes encoding Hsp90 that encode nearly identical amino-acid sequences, one of which is constitutively expressed, while the other is HSF1 inducible (4). The constitutive expression level of Hsp90 under non-stress conditions is very high and Hsp90 is one of the most abundant proteins in the cell, accumulating primarily in the cytosol (5, 6). The high expression level of Hsp90 in the absence of stress is consistent with its requirement for the activity of many critical signal transduction clients in eukaryotes. Hsp90 is essential for viability in eukaryotes (4). In bacteria, which lack the wealth of signal transduction proteins present in eukaryotes, Hsp90 knockouts are viable (7).
The Hsp90 chaperone is highly conserved in eukaryotes both at the level of amino-acid sequence and biochemical function. The Saccharomyces cerevisiae and human Hsp90 proteins are 59% identical in amino acid sequence alignments. The conservation of Hsp90 in eukaryotes extends to the functional level as S. cerevisiae with both endogenous Hsp90 genes knocked out can be rescued by expression of human Hsp90 (8). Hsp90 serves as a protein interaction hub, binding to about 10% of the yeast proteome (9) including many co-chaperones necessary for client maturation. The observation that human Hsp90 complements budding yeast knockouts indicates that essential elements of this hub are evolutionarily conserved. Consistent with the functional conservation of Hsp90, steroid hormone receptors that are not natural to budding yeast function in an Hsp90-dependent manner when introduced into yeast (8).
Hsp90 contains three domains: the N-terminal (N) domain, middle (M) domain, and the C-terminal (C) domain. The N-domain binds to ATP and together with the M-domain catalyzes ATP hydrolysis (10, 11). The C-domain forms a stable dimer (12). Dimerization of Hsp90 is required for client maturation (13). Recent tour-de-force structural studies have revealed that flexible hinges between all three domains enables Hsp90 to adopt dramatically different 3-dimensional conformations (14–16). Isolated Hsp90 is a slow ATPase, ranging from about 1 min−1 for yeast Hsp90 to about 0.1 min−1 for human Hsp90, and ATP binding and hydrolysis cause Hsp90 to preferentially shift between an open-conformation with the N-domains separated, and a closed conformation with the N-domains associated (17–19). These open and closed structural transitions are known to drive the binding of many co-chaperones (14, 20–22) and are believed to drive the maturation of clients (Figure 8.1)
Figure 8.1. Model of the ATPase driven conformational cycle of Hsp90 that leads to client maturation.
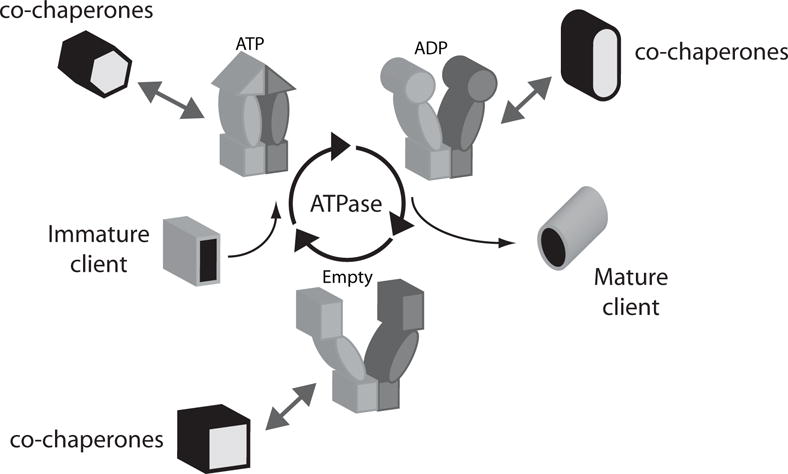
In addition to client and ATP, numerous co-chaperones bind to Hsp90 and influence its chaperone activity. Many of these Hsp90 binding interactions are inter-dependent. The nucleotide-bound state of Hsp90 influences the binding of both co-chaperones and clients. Understanding how the inter-dependent and often transient binding of co-chaperones and nucleotide lead to client maturation is an active area of investigation.
Hsp90 has a unique ATP binding site and the discovery of small-molecule inhibitors that specifically inhibit Hsp90 has provided useful research tools that also show promise as anticancer therapeutic agents. Hsp90 inhibitors were originally uncovered from a natural product screen for compounds that reversed the transformed phenotype of the v-src oncogenic kinase. One of the natural products from this screen, geldanamycin (GA) was found to bind specifically to Hsp90 and inhibit its ability to chaperone v-src (23). While Hsp90 function is essential for the viability of healthy cells, cancer cells tend to have a higher level of Hsp90-dependent clients and a higher required level of Hsp90 activity. For many types of cancer there appears to be a therapeutic window where cancer cells can be preferentially impacted by Hsp90 inhibiton compared to healthy cells. The medical interest in Hsp90 has spurred the synthesis of many small-molecule inhibitors as drug candidates (24).
Hsp90 is a specialized molecular chaperone that is required for the maturation of a limited set of clients. Many newly synthesized proteins require general molecular chaperones including Hsp60 (GroEL in bacteria), Hsp70 and Hsp40 in order to fold properly, but only a small fraction of these rely upon Hsp90. The limited proteins that require Hsp90 to mature to their active state are referred to as Hsp90 clients. These Hsp90 clients fall into three main categories: protein kinases, steroid hormone receptors, and non-signal transduction clients (Table 8.1). Within each greater client class, research has revealed important aspects of the mechanism of Hsp90-mediated client maturation, yet the molecular mechanism by which Hsp90 binds to and activates clients remains largely a mystery.
Table 8.1.
Classes of Hsp90 Clients.
| Client Class | Example Clients | Co-chaperones | References |
|---|---|---|---|
| Steroid Hormone Receptors | Progesterone & Glococorticoid Receptors | FKBP51, FKBP52, Hop, p23 | (38) |
| Kinase | v-src, B-Raf | Cdc37, Hop, p23 | (23, 50) |
| Non Signal Transduction | |||
| Telomerase | Telomerase | p23 | (54) |
| Antigen Presentation | MHC peptide loading | (59, 60) | |
| CFTR | CFTR | Aha1 | (61, 63, 64) |
Steroid Hormone Receptor Hsp90 Clients
Steroid hormone receptors are a class of transcription factors that are activated by binding to steroid agonist and were some of the first proteins actively studied as Hsp90 clients. In 1984, Baulieu and colleagues reported that monoclonal antibodies raised against progesterone receptor (PR) recognized a 90kD protein that tightly associated with apo-receptor, but not steroid-bound receptor (25), properties that indicated a regulatory role for the 90kD protein. Soon after, Pratt and colleagues reported that monoclonal antibodies raised against glucocorticoid receptor (GR) were capable of immuno-isolating the receptor as well as a 90kD protein that cross-reacted with antibodies raised against Hsp90 (26); thus identifying Hsp90 as a steroid hormone receptor chaperone.
Hsp90 was found to bind strongly to the apo-form of many other steroid-hormone receptors including the estrogen and androgen receptors; however some receptors such as those for retinoic acid and thyroid hormone showed little or no affinity for Hsp90 (27–30). Different DNA binding properties in the absence of steroid agonist were noted for Hsp90-dependent and Hsp90-independent receptors in cell-free expression systems (27). In the absence of steroid agonist, Hsp90-independent receptors were found to bind to target DNA in the presence and absence of Hsp90. In contrast, Hsp90-dependent receptors without steroid agonist only bound to DNA when Hsp90 was absent. In the absence of steroid agonist, Hsp90-dependent receptors were found in the cytoplasm while Hsp90-independent receptors were found in the nucleus. At the time, these results led to the hypothesis that Hsp90 functions to repress steroid hormone receptors by sequestering them in the cytoplasm. However, it is now appreciated that Hsp90-dependent receptors require Hsp90 in order to bind to steroid. Steroid binding controls the activity of all steroid receptors and for Hsp90-dependent receptors, this step requires Hsp90 activity.
Co-immunoprecipitation (co-IP) experiments revealed many components of the Hsp90 chaperone complexes involved in receptor maturation and paved the way for in vitro reconstitution of the chaperone reaction. Glutaraldehyde crosslinking studies revealed that the stoichiometry of receptor-Hsp90 complexes was 1:2 indicating that a receptor monomer binds to an Hsp90 dimer (31). From immuno-isolation of steroid receptors expressed in cell-free expression systems it was discovered that receptor-Hsp90 complexes included a number of co-chaperones including Hsp70, p23 and Hop (32, 33). A combination of these three co-chaperones along with the general chaperone Hsp40 and Hsp90 were found to be sufficient to develop a purified chaperone system for the maturation of progesterone receptor to the steroid-bound active form (34).
Structural and biochemical approaches have shown that steroid hormone receptors bind steroid ligand in an internal cavity and indicate that Hsp90 remodels this cavity to provide a solvent-accessible path for the hormone to enter. X-ray crystal (35, 36) structures clearly show that steroid ligands bind to receptors in an internal cavity that is completely inaccessible to solvent. Evidence for Hsp90 opening of this cleft comes from chemical modification studies that indicate modestly higher susceptibility of the steroid-binding cavity in the Hsp90-bound form of apo-GR (37). While the mechanism by which Hsp90 opens the steroid binding cleft remains unknown, the data clearly indicate that isolated apo-GR and apo-PR are not able to bind to steroid agonist, but that when complexed with Hsp90 the binding to steroid becomes efficient (38).
While much is understood about Hsp90 mediated steroid receptor maturation, many important aspects of the molecular mechanism remain open questions. It is clear that ATP hydrolysis by Hsp90 is required for efficient PR maturation (39). However, it is not clear how the ATPase driven conformational cycle of Hsp90 leads to opening of the steroid-binding cavity. What are the molecular interactions that cause apo-receptor to bind to Hsp90, Hsp90 to catalyze steroid binding, and steroid-bound receptor to dissociate from the chaperone complex? These questions are challenging to answer because of the dynamic nature of the Hsp90 conformational cycle, but may be addressed by structural and biochemical approaches that trap individual steps in the process.
Kinase Clients of Hsp90
The oncogenic v-src kinase was first shown to bind to Hsp90 through co-IP analysis (40), but it was not clear that Hsp90 was required for v-src kinase activity until it was reported that the natural compound geldanamycin (GA) prevents v-src activity indirectly by inhibiting Hsp90 (23). When v-src is expressed in the presence of GA, it accumulates in an inactive form indicating that Hsp90 is required for the maturation of v-src to an active form. In addition, treatment of cells with GA causes an increase in the proteosome mediated degradation rate of v-src and a decrease in the steady-state v-src level (41). Thus, Hsp90 is required for the maturation of v-src kinase to an active form and also protects v-src from degradation.
The maturation of kinases mediated by Hsp90 requires co-chaperones including the kinase-specific co-chaperone Cdc37. Cdc37 was originally identified in a screen for genes required for cell cycle progression (42). Genetic evidence for a functional interaction between Cdc37 and Hsp90 came from studies showing that either mutations in Cdc37 or a reduction in the level of the yeast Hsp90 protein suppresses the lethality of v-src overexpression (43, 44). Mutations in Hsp90 and Cdc37 were also found to exacerbate a defect in the Drosophila melanogaster sevenless receptor tyrosine kinase (45). Thus, Hsp90 and Cdc37 are both involved in the maturation of these model kinases. A physical connection between Cdc37 and Hsp90 was first indicated by co-IP analysis with the cyclin dependent kinase CDK4 (46). Both Hsp90 and Cdc37 were found to associate with CDK4 during co-IP analysis with an anti-CKD4 antibody suggesting there may be a direct mechanistic link between Hsp90 and Cdc37 in chaperoning kinases.
The direct interaction between Cdc37 and Hsp90 has recently been confirmed and described in atomic detail through structural analyses. Comparison of full-length yeast Hsp90 to truncated Hsp90 constructs indicated that the isolated Hsp90 N-domain was primarily responsible for mediating binding to the C-terminal domain of human Cdc37 (22). Co-crystallization of these domains revealed the atomic details of this interaction (22). More recently, Pearl and colleagues have successfully purified a homogenous complex of human Cdk4 and Cdc37 along with Hsp90 from the insect Spodoptera frugiperda expression system and analyzed the structure of this complex by negative stained electron microscopy (16). This tour de force work indicates that the kinase loaded complex consists of an Hsp90 dimer bound to single copies of Cdc37 and Cdk4. The observed electron density can be well fit with a structural model where Cdc37 binds to the Hsp90 N-domain and Cdk4 binds to the Hsp90 M-domain. Biochemical analyses of Cdk4-Cdc37 complexes in the absence of Hsp90 indicate a stoichiometry of one kinase to two Cdc37 molecules (16). The biochemical and structural data are consistent with an ordered model where a dimer of Cdc37 initially binds to kinases and subsequent binding of this complex to Hsp90 results in the release of one Cdc37 molecule.
The association of human Cdc37 with yeast Hsp90 observed biochemically and structurally (22) is consistent with the high amino acid conservation of Hsp90 between these two species. This conservation is also apparent at a functional level as metazoan Cdc37 (from flies) is capable of rescuing robust growth of S. cerevisiae made unhealthy by a point mutant in the yeast Cdc37 (45). Unlike Hsp90, the amino acid sequence of Cdc37 is divergent from yeast to humans with only 17% sequence identity after alignment. Of note, yeast Cdc37 binds to yeast Hsp90 with about 100-fold weaker affinity than human Cdc37 (47), indicating that while qualitative Cdc37 function may be similar across eukaryotes, the interaction details are quantitatively distinct.
The Hsp90 and Cdc37 chaperones stabilize the cellular accumulation of many kinases. Indeed, the observation that Hsp90 inhibition can lead to decreased cellular levels of client kinases has become a convenient screen to identify Hsp90-dependent kinases. In mammalian cells, inhibition of Hsp90 activity resulted in reduced levels of 80 out of 105 kinases analyzed (48). A recent study also found that more than 50% of the S. cerevisiae kinome was affected by the loss of functional Cdc37 (49) confirming the importance of a functional Hsp90:Cdc37 chaperone complex for the stability of cellular protein kinases.
Recent efforts have described the minimum chaperone requirements for the maturation of a kinase in vitro (50). Arlander et al. identified a five protein system for chaperoning the Chk1 kinase in vitro that consisted of Hsp90, Hsp70, Hsp40, Cdc37 and CK2 (50). The in vitro chaperone systems for kinases and steroid hormone receptors differ in their co-chaperone requirements and sensitivities, indicating the influential role that co-chaperones play in the client maturation process (34, 51). As with steroid hormone receptors, the in vitro chaperone system for kinases provides a promising route to carefully analyze the client maturation process in detail.
Non-signal transduction Hsp90 clients
Recent evidence indicates that Hsp90 is involved in the maturation of active telomerase. The catalytic core of telomerase consists of a protein component (TERT) and a template RNA (TR). Chaperones were originally implicated in the maturation of telomerase from the observation that reconstitution of active telomerase from isolated TERT and TR was stimulated by increasing concentration of chaperones present in reticulocyte lysates (52). A two-hybrid screen revealed that the Hsp90 co-chaperone p23 interacted with TERT and co-IP analyses revealed that both Hsp90 and p23 are involved in complexes with telomerase in mammalian cells (52). More recent biochemical dissections of telomerase function indicate that the interplay between Hsp90 and p23 influences the dynamics of telomerase binding to telomeres (53). Hsp90 activity in vivo is important for the proper maintenance of telomeric DNA length that decreases when Hsp90 activity is impaired or when Hsp90 is overexpressed (54, 55).
Hsp90 can bind to antigenic peptides and has been implicated in the delivery of these peptides to major histocompatability complexes (MHCs). Hsp90 was first connected with antigen presentation from analyses of antigens in tumors that could confer immunity in syngenic recipients (56). Fractionation studies revealed that Hsp90 from the tumors was important in the development of immunity (57, 58). Cellular studies indicated that antigenic peptides complexed with Hsp90 are loaded on to MHC class I with much higher efficiency than free peptides (59), which may otherwise be prone to rapid degradation. In addition, the delivery of peptides to MHC class II proteins is reduced upon treatment of cells with Hsp90 inhibitors (60). The molecular details of Hsp90 involvement in antigen presentation are a current area of active investigation.
In addition to soluble proteins, Hsp90 has recently been implicated in the maturation of CFTR, an integral membrane protein. Hsp90 was originally implicated in CFTR maturation from the observation that treatment of mammalian cells with the Hsp90-specific inhibitors prevented newly synthesized wild-type CFTR from transitioning to the maturely glycosylated cell-surface form (61). Cystic fibrosis-causing mutations in CFTR, including the most commonly occuring – deletion of a phenylalanine codon (ΔF508), cause defective folding and export of CFTR from the ER (62). Immunoprecipitation of wild-type and ΔF508 CFTR followed by mass spectrometry revealed that Hsp90 and many co-chaperones exhibited differential binding for wild-type and mutant CFTR (63). RNA interference screening of these co-chaperones demonstrated that silencing of the Hsp90 co-chaperone Aha1 rescued cell-surface expression and chloride ion transport of ΔF508 CFTR (63). Recent biochemical analyses indicate that Aha1 associates with ΔF508 CFTR nearly twice as much as with wild-type CFTR (64). The observation that reducing the association of ΔF508 CFTR with Aha1 corrects the function of this anion channel provides a promising avenue for the development of therapeutics to treat cystic fibrosis.
Hsp90 can function as a promiscuous anti-aggregation molecule in vitro for a large variety of hard-to-fold proteins, but the general anti-aggregation properties of Hsp90 do not appear important in cells. Following the discovery that Hsp90 expression increased in response to conditions that stress protein folding, a number of studies examined the capacity of Hsp90 to assist in the folding of proteins that were difficult to fold on their own. The anti-aggregation properties of Hsp90 for unfolded CFTR nucleotide binding domain 1 (65), citrate synthase (66), and p53 (67) do not require the binding and hydrolysis of ATP by Hsp90. In contrast both steroid hormone receptors and kinase clients of Hsp90 are dramatically impaired by mutations that destroy ATPase activity or competitive inhibition of the ATP binding site with drugs (23, 39). Thus, the anti-aggregation properties of Hsp90 do not seem to require the same ATPase driven Hsp90 conformational cycle that is implicated in the maturation of kinases and steroid hormone receptors. Multiple chaperones in addition to Hsp90 are capable of providing anti-aggregation properties in cells providing an explanation for the observation that Hsp90 is not required for the folding and solubility of most proteins in cells (68).
Current and future areas of active interest
Many important cellular roles of Hsp90 have been described and much of the current and future research in Hsp90 involves analyses of molecular mechanism. The molecular mechanism by which Hsp90 catalyzes the maturation of any client is an open and active area of research. Current structural models indicate that Hsp90 undergoes dramatic rearrangements in response to ATP binding and hydrolysis (14, 15, 19). However, how these structural rearrangements lead to client maturation is not clear. From the engineering of a monomeric Hsp90 variant, it is clear that dimerization of Hsp90 is critical for efficient ATP hydrolysis as well as the activation of a kinase and a steroid hormone receptor client (13). How each subunit in the Hsp90 dimer binds to and hydrolyzes ATP during the maturation of a client remains unclear. In the coming years, research will help to reveal the molecular mechanism by which Hsp90 binds to client proteins and along with co-chaperones and ATP catalyzes the maturation of these clients to active states.
References
- 1.Ritossa F. A new puffing pattern induced by temperature shock and DNP in drosophila. Experientia. 1962;18:571–573. [Google Scholar]
- 2.McKenzie SL, Henikoff S, Meselson M. Localization of RNA from heat-induced polysomes at puff sites in Drosophila melanogaster. Proc Natl Acad Sci U S A. 1975;72:1117–21. doi: 10.1073/pnas.72.3.1117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Lindquist S. The heat-shock response. Annu Rev Biochem. 1986;55:1151–91. doi: 10.1146/annurev.bi.55.070186.005443. [DOI] [PubMed] [Google Scholar]
- 4.Borkovich KA, Farrelly FW, Finkelstein DB, Taulien J, Lindquist S. hsp82 is an essential protein that is required in higher concentrations for growth of cells at higher temperatures. Mol Cell Biol. 1989;9:3919–30. doi: 10.1128/mcb.9.9.3919. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Ghaemmaghami S, Huh WK, Bower K, Howson RW, Belle A, Dephoure N, O’Shea EK, Weissman JS. Global analysis of protein expression in yeast. Nature. 2003;425:737–41. doi: 10.1038/nature02046. [DOI] [PubMed] [Google Scholar]
- 6.Huh WK, Falvo JV, Gerke LC, Carroll AS, Howson RW, Weissman JS, O’Shea EK. Global analysis of protein localization in budding yeast. Nature. 2003;425:686–91. doi: 10.1038/nature02026. [DOI] [PubMed] [Google Scholar]
- 7.Bardwell JC, Craig EA. Ancient heat shock gene is dispensable. J Bacteriol. 1988;170:2977–83. doi: 10.1128/jb.170.7.2977-2983.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Picard D, Khursheed B, Garabedian MJ, Fortin MG, Lindquist S, Yamamoto KR. Reduced levels of hsp90 compromise steroid receptor action in vivo. Nature. 1990;348:166–8. doi: 10.1038/348166a0. [DOI] [PubMed] [Google Scholar]
- 9.Zhao R, Davey M, Hsu YC, Kaplanek P, Tong A, Parsons AB, Krogan N, Cagney G, Mai D, Greenblatt J, Boone C, Emili A, Houry WA. Navigating the chaperone network: an integrative map of physical and genetic interactions mediated by the hsp90 chaperone. Cell. 2005;120:715–27. doi: 10.1016/j.cell.2004.12.024. [DOI] [PubMed] [Google Scholar]
- 10.Panaretou B, Prodromou C, Roe SM, O’Brien R, Ladbury JE, Piper PW, Pearl LH. ATP binding and hydrolysis are essential to the function of the Hsp90 molecular chaperone in vivo. Embo J. 1998;17:4829–36. doi: 10.1093/emboj/17.16.4829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Meyer P, Prodromou C, Hu B, Vaughan C, Roe SM, Panaretou B, Piper PW, Pearl LH. Structural and functional analysis of the middle segment of hsp90: implications for ATP hydrolysis and client protein and cochaperone interactions. Mol Cell. 2003;11:647–58. doi: 10.1016/s1097-2765(03)00065-0. [DOI] [PubMed] [Google Scholar]
- 12.Minami Y, Kimura Y, Kawasaki H, Suzuki K, Yahara I. The carboxy-terminal region of mammalian HSP90 is required for its dimerization and function in vivo. Mol Cell Biol. 1994;14:1459–64. doi: 10.1128/mcb.14.2.1459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Wayne N, Bolon DN. Dimerization of Hsp90 is required for in vivo function. Design and analysis of monomers and dimers. J Biol Chem. 2007;282:35386–95. doi: 10.1074/jbc.M703844200. [DOI] [PubMed] [Google Scholar]
- 14.Ali MM, Roe SM, Vaughan CK, Meyer P, Panaretou B, Piper PW, Prodromou C, Pearl LH. Crystal structure of an Hsp90-nucleotide-p23/Sba1 closed chaperone complex. Nature. 2006;440:1013–7. doi: 10.1038/nature04716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Shiau AK, Harris SF, Southworth DR, Agard DA. Structural Analysis of E. coli hsp90 reveals dramatic nucleotide-dependent conformational rearrangements. Cell. 2006;127:329–40. doi: 10.1016/j.cell.2006.09.027. [DOI] [PubMed] [Google Scholar]
- 16.Vaughan CK, Gohlke U, Sobott F, Good VM, Ali MM, Prodromou C, Robinson CV, Saibil HR, Pearl LH. Structure of an Hsp90-Cdc37-Cdk4 complex. Mol Cell. 2006;23:697–707. doi: 10.1016/j.molcel.2006.07.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Hessling M, Richter K, Buchner J. Dissection of the ATP-induced conformational cycle of the molecular chaperone Hsp90. Nat Struct Mol Biol. 2009;16:287–93. doi: 10.1038/nsmb.1565. [DOI] [PubMed] [Google Scholar]
- 18.Mickler M, Hessling M, Ratzke C, Buchner J, Hugel T. The large conformational changes of Hsp90 are only weakly coupled to ATP hydrolysis. Nat Struct Mol Biol. 2009;16:281–6. doi: 10.1038/nsmb.1557. [DOI] [PubMed] [Google Scholar]
- 19.Southworth DR, Agard DA. Species-dependent ensembles of conserved conformational states define the Hsp90 chaperone ATPase cycle. Mol Cell. 2008;32:631–40. doi: 10.1016/j.molcel.2008.10.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.McLaughlin SH, Sobott F, Yao ZP, Zhang W, Nielsen PR, Grossmann JG, Laue ED, Robinson CV, Jackson SE. The co-chaperone p23 arrests the Hsp90 ATPase cycle to trap client proteins. J Mol Biol. 2006;356:746–58. doi: 10.1016/j.jmb.2005.11.085. [DOI] [PubMed] [Google Scholar]
- 21.Panaretou B, Siligardi G, Meyer P, Maloney A, Sullivan JK, Singh S, Millson SH, Clarke PA, Naaby-Hansen S, Stein R, Cramer R, Mollapour M, Workman P, Piper PW, Pearl LH, Prodromou C. Activation of the ATPase activity of hsp90 by the stress-regulated cochaperone aha1. Mol Cell. 2002;10:1307–18. doi: 10.1016/s1097-2765(02)00785-2. [DOI] [PubMed] [Google Scholar]
- 22.Roe SM, Ali MM, Meyer P, Vaughan CK, Panaretou B, Piper PW, Prodromou C, Pearl LH. The Mechanism of Hsp90 regulation by the protein kinase-specific cochaperone p50(cdc37) Cell. 2004;116:87–98. doi: 10.1016/s0092-8674(03)01027-4. [DOI] [PubMed] [Google Scholar]
- 23.Whitesell L, Mimnaugh EG, De Costa B, Myers CE, Neckers LM. Inhibition of heat shock protein HSP90-pp60v-src heteroprotein complex formation by benzoquinone ansamycins: essential role for stress proteins in oncogenic transformation. Proc Natl Acad Sci U S A. 1994;91:8324–8. doi: 10.1073/pnas.91.18.8324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Whitesell L, Lindquist SL. HSP90 and the chaperoning of cancer. Nat Rev Cancer. 2005;5:761–72. doi: 10.1038/nrc1716. [DOI] [PubMed] [Google Scholar]
- 25.Joab I, Radanyi C, Renoir M, Buchou T, Catelli MG, Binart N, Mester J, Baulieu EE. Common non-hormone binding component in non-transformed chick oviduct receptors of four steroid hormones. Nature. 1984;308:850–3. doi: 10.1038/308850a0. [DOI] [PubMed] [Google Scholar]
- 26.Sanchez ER, Toft DO, Schlesinger MJ, Pratt WB. Evidence that the 90-kDa phosphoprotein associated with the untransformed L-cell glucocorticoid receptor is a murine heat shock protein. J Biol Chem. 1985;260:12398–401. [PubMed] [Google Scholar]
- 27.Dalman FC, Koenig RJ, Perdew GH, Massa E, Pratt WB. In contrast to the glucocorticoid receptor, the thyroid hormone receptor is translated in the DNA binding state and is not associated with hsp90. J Biol Chem. 1990;265:3615–8. [PubMed] [Google Scholar]
- 28.Dalman FC, Sturzenbecker LJ, Levin AA, Lucas DA, Perdew GH, Petkovitch M, Chambon P, Grippo JF, Pratt WB. Retinoic acid receptor belongs to a subclass of nuclear receptors that do not form “docking” complexes with hsp90. Biochemistry. 1991;30:5605–8. doi: 10.1021/bi00236a038. [DOI] [PubMed] [Google Scholar]
- 29.Redeuilh G, Moncharmont B, Secco C, Baulieu EE. Subunit composition of the molybdate-stabilized “8–9 S” nontransformed estradiol receptor purified from calf uterus. J Biol Chem. 1987;262:6969–75. [PubMed] [Google Scholar]
- 30.Veldscholte J, Berrevoets CA, Zegers ND, van der Kwast TH, Grootegoed JA, Mulder E. Hormone-induced dissociation of the androgen receptor-heat-shock protein complex: use of a new monoclonal antibody to distinguish transformed from nontransformed receptors. Biochemistry. 1992;31:7422–30. doi: 10.1021/bi00147a029. [DOI] [PubMed] [Google Scholar]
- 31.Denis M, Wikstrom AC, Gustafsson JA. The molybdate-stabilized nonactivated glucocorticoid receptor contains a dimer of Mr 90,000 non-hormone-binding protein. J Biol Chem. 1987;262:11803–6. [PubMed] [Google Scholar]
- 32.Smith DF, Schowalter DB, Kost SL, Toft DO. Reconstitution of progesterone receptor with heat shock proteins. Mol Endocrinol. 1990;4:1704–11. doi: 10.1210/mend-4-11-1704. [DOI] [PubMed] [Google Scholar]
- 33.Smith DF, Toft DO. Composition, assembly and activation of the avian progesterone receptor. J Steroid Biochem Mol Biol. 1992;41:201–7. doi: 10.1016/0960-0760(92)90345-j. [DOI] [PubMed] [Google Scholar]
- 34.Kosano H, Stensgard B, Charlesworth MC, McMahon N, Toft D. The assembly of progesterone receptor-hsp90 complexes using purified proteins. J Biol Chem. 1998;273:32973–9. doi: 10.1074/jbc.273.49.32973. [DOI] [PubMed] [Google Scholar]
- 35.Wagner RL, Apriletti JW, McGrath ME, West BL, Baxter JD, Fletterick RJ. A structural role for hormone in the thyroid hormone receptor. Nature. 1995;378:690–7. doi: 10.1038/378690a0. [DOI] [PubMed] [Google Scholar]
- 36.Williams SP, Sigler PB. Atomic structure of progesterone complexed with its receptor. Nature. 1998;393:392–6. doi: 10.1038/30775. [DOI] [PubMed] [Google Scholar]
- 37.Stancato LF, Silverstein AM, Gitler C, Groner B, Pratt WB. Use of the thiol-specific derivatizing agent N-iodoacetyl-3-[125I]iodotyrosine to demonstrate conformational differences between the unbound and hsp90-bound glucocorticoid receptor hormone binding domain. J Biol Chem. 1996;271:8831–6. doi: 10.1074/jbc.271.15.8831. [DOI] [PubMed] [Google Scholar]
- 38.Pratt WB, Toft DO. Regulation of signaling protein function and trafficking by the hsp90/hsp70-based chaperone machinery. Exp Biol Med (Maywood) 2003;228:111–33. doi: 10.1177/153537020322800201. [DOI] [PubMed] [Google Scholar]
- 39.Grenert JP, Johnson BD, Toft DO. The importance of ATP binding and hydrolysis by hsp90 in formation and function of protein heterocomplexes. J Biol Chem. 1999;274:17525–33. doi: 10.1074/jbc.274.25.17525. [DOI] [PubMed] [Google Scholar]
- 40.Brugge JS, Erikson E, Erikson RL. The specific interaction of the Rous sarcoma virus transforming protein, pp60src, with two cellular proteins. Cell. 1981;25:363–72. doi: 10.1016/0092-8674(81)90055-6. [DOI] [PubMed] [Google Scholar]
- 41.An WG, Schulte TW, Neckers LM. The heat shock protein 90 antagonist geldanamycin alters chaperone association with p210bcr-abl and v-src proteins before their degradation by the proteasome. Cell Growth Differ. 2000;11:355–60. [PubMed] [Google Scholar]
- 42.Reed SI. The selection of S. cerevisiae mutants defective in the start event of cell division. Genetics. 1980;95:561–77. doi: 10.1093/genetics/95.3.561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Dey B, Lightbody JJ, Boschelli F. CDC37 is required for p60v-src activity in yeast. Mol Biol Cell. 1996;7:1405–17. doi: 10.1091/mbc.7.9.1405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Xu Y, Lindquist S. Heat-shock protein hsp90 governs the activity of pp60v-src kinase. Proc Natl Acad Sci U S A. 1993;90:7074–8. doi: 10.1073/pnas.90.15.7074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Cutforth T, Rubin GM. Mutations in Hsp83 and cdc37 impair signaling by the sevenless receptor tyrosine kinase in Drosophila. Cell. 1994;77:1027–36. doi: 10.1016/0092-8674(94)90442-1. [DOI] [PubMed] [Google Scholar]
- 46.Dai K, Kobayashi R, Beach D. Physical interaction of mammalian CDC37 with CDK4. J Biol Chem. 1996;271:22030–4. doi: 10.1074/jbc.271.36.22030. [DOI] [PubMed] [Google Scholar]
- 47.Siligardi G, Panaretou B, Meyer P, Singh S, Woolfson DN, Piper PW, Pearl LH, Prodromou C. Regulation of Hsp90 ATPase activity by the co-chaperone Cdc37p/p50cdc37. J Biol Chem. 2002;277:20151–9. doi: 10.1074/jbc.M201287200. [DOI] [PubMed] [Google Scholar]
- 48.Citri A, Harari D, Shohat G, Ramakrishnan P, Gan J, Lavi S, Eisenstein M, Kimchi A, Wallach D, Pietrokovski S, Yarden Y. Hsp90 recognizes a common surface on client kinases. J Biol Chem. 2006;281:14361–9. doi: 10.1074/jbc.M512613200. [DOI] [PubMed] [Google Scholar]
- 49.Mandal AK, Lee P, Chen JA, Nillegoda N, Heller A, DiStasio S, Oen H, Victor J, Nair DM, Brodsky JL, Caplan AJ. Cdc37 has distinct roles in protein kinase quality control that protect nascent chains from degradation and promote posttranslational maturation. J Cell Biol. 2007;176:319–28. doi: 10.1083/jcb.200604106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Arlander SJ, Felts SJ, Wagner JM, Stensgard B, Toft DO, Karnitz LM. Chaperoning checkpoint kinase 1 (Chk1), an Hsp90 client, with purified chaperones. J Biol Chem. 2006;281:2989–98. doi: 10.1074/jbc.M508687200. [DOI] [PubMed] [Google Scholar]
- 51.Dittmar KD, Banach M, Galigniana MD, Pratt WB. The role of DnaJ-like proteins in glucocorticoid receptor.hsp90 heterocomplex assembly by the reconstituted hsp90.p60.hsp70 foldosome complex. J Biol Chem. 1998;273:7358–66. doi: 10.1074/jbc.273.13.7358. [DOI] [PubMed] [Google Scholar]
- 52.Holt SE, Aisner DL, Baur J, Tesmer VM, Dy M, Ouellette M, Trager JB, Morin GB, Toft DO, Shay JW, Wright WE, White MA. Functional requirement of p23 and Hsp90 in telomerase complexes. Genes Dev. 1999;13:817–26. doi: 10.1101/gad.13.7.817. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.DeZwaan DC, Toogun OA, Echtenkamp FJ, Freeman BC. The Hsp82 molecular chaperone promotes a switch between unextendable and extendable telomere states. Nat Struct Mol Biol. 2009;16:711–6. doi: 10.1038/nsmb.1616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Grandin N, Charbonneau M. Hsp90 levels affect telomere length in yeast. Mol Genet Genomics. 2001;265:126–34. doi: 10.1007/s004380000398. [DOI] [PubMed] [Google Scholar]
- 55.Toogun OA, Dezwaan DC, Freeman BC. The hsp90 molecular chaperone modulates multiple telomerase activities. Mol Cell Biol. 2008;28:457–67. doi: 10.1128/MCB.01417-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Prehn RT, Main JM. Immunity to methylcholanthrene-induced sarcomas. J Natl Cancer Inst. 1957;18:769–78. [PubMed] [Google Scholar]
- 57.DuBois GC, Law LW, Appella E. Purification and biochemical properties of tumor-associated transplantation antigens from methylcholanthrene-induced murine sarcomas. Proc Natl Acad Sci U S A. 1982;79:7669–73. doi: 10.1073/pnas.79.24.7669. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Srivastava PK, DeLeo AB, Old LJ. Tumor rejection antigens of chemically induced sarcomas of inbred mice. Proc Natl Acad Sci U S A. 1986;83:3407–11. doi: 10.1073/pnas.83.10.3407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Binder RJ, Blachere NE, Srivastava PK. Heat shock protein-chaperoned peptides but not free peptides introduced into the cytosol are presented efficiently by major histocompatibility complex I molecules. J Biol Chem. 2001;276:17163–71. doi: 10.1074/jbc.M011547200. [DOI] [PubMed] [Google Scholar]
- 60.Rajagopal D, Bal V, Mayor S, George A, Rath S. A role for the Hsp90 molecular chaperone family in antigen presentation to T lymphocytes via major histocompatibility complex class II molecules. Eur J Immunol. 2006;36:828–41. doi: 10.1002/eji.200535326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Loo MA, Jensen TJ, Cui L, Hou Y, Chang XB, Riordan JR. Perturbation of Hsp90 interaction with nascent CFTR prevents its maturation and accelerates its degradation by the proteasome. Embo J. 1998;17:6879–87. doi: 10.1093/emboj/17.23.6879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Riordan JR. Assembly of functional CFTR chloride channels. Annu Rev Physiol. 2005;67:701–18. doi: 10.1146/annurev.physiol.67.032003.154107. [DOI] [PubMed] [Google Scholar]
- 63.Wang X, Venable J, LaPointe P, Hutt DM, Koulov AV, Coppinger J, Gurkan C, Kellner W, Matteson J, Plutner H, Riordan JR, Kelly JW, Yates JR, 3rd, Balch WE. Hsp90 cochaperone Aha1 downregulation rescues misfolding of CFTR in cystic fibrosis. Cell. 2006;127:803–15. doi: 10.1016/j.cell.2006.09.043. [DOI] [PubMed] [Google Scholar]
- 64.Sun F, Mi Z, Condliffe SB, Bertrand CA, Gong X, Lu X, Zhang R, Latoche JD, Pilewski JM, Robbins PD, Frizzell RA. Chaperone displacement from mutant cystic fibrosis transmembrane conductance regulator restores its function in human airway epithelia. Faseb J. 2008;22:3255–63. doi: 10.1096/fj.07-105338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Youker RT, Walsh P, Beilharz T, Lithgow T, Brodsky JL. Distinct roles for the Hsp40 and Hsp90 molecular chaperones during cystic fibrosis transmembrane conductance regulator degradation in yeast. Mol Biol Cell. 2004;15:4787–97. doi: 10.1091/mbc.E04-07-0584. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Jakob U, Lilie H, Meyer I, Buchner J. Transient interaction of Hsp90 with early unfolding intermediates of citrate synthase. Implications for heat shock in vivo. J Biol Chem. 1995;270:7288–94. doi: 10.1074/jbc.270.13.7288. [DOI] [PubMed] [Google Scholar]
- 67.Muller L, Schaupp A, Walerych D, Wegele H, Buchner J. Hsp90 regulates the activity of wild type p53 under physiological and elevated temperatures. J Biol Chem. 2004;279:48846–54. doi: 10.1074/jbc.M407687200. [DOI] [PubMed] [Google Scholar]
- 68.Nathan DF, Vos MH, Lindquist S. In vivo functions of the Saccharomyces cerevisiae Hsp90 chaperone. Proc Natl Acad Sci U S A. 1997;94:12949–56. doi: 10.1073/pnas.94.24.12949. [DOI] [PMC free article] [PubMed] [Google Scholar]