

Abstract
In typical functional connectivity studies, connections between voxels or regions in the brain are represented as edges in a network. Networks for different subjects are constructed at a given graph density and are summarized by some network measure such as path length. Examining these summary measures for many density values yields samples of connectivity curves, one for each individual. This has led to the adoption of basic tools of functional data analysis, most commonly to compare control and disease groups through the average curves in each group. Such group differences, however, neglect the variability in the sample of connectivity curves. In this article, the use of functional principal component analysis (FPCA) is demonstrated to enrich functional connectivity studies by providing increased power and flexibility for statistical inference. Specifically, individual connectivity curves are related to individual characteristics such as age and measures of cognitive function, thus providing a tool to relate brain connectivity with these variables at the individual level. This individual level analysis opens a new perspective that goes beyond previous group level comparisons. Using a large data set of resting-state functional magnetic resonance imaging scans, relationships between connectivity and two measures of cognitive function—episodic memory and executive function—were investigated. The group-based approach was implemented by dichotomizing the continuous cognitive variable and testing for group differences, resulting in no statistically significant findings. To demonstrate the new approach, FPCA was implemented, followed by linear regression models with cognitive scores as responses, identifying significant associations of connectivity in the right middle temporal region with both cognitive scores.
Keywords: : connectivity curves, episodic memory, executive function, fMRI, functional data analysis, functional principal component analysis, network, network density, resting state
Introduction
In recent years, spatial patterns of distributed neural activity in the brain have been increasingly modeled in terms of graphs or networks (Fallani et al., 2014). Data arrive in the form of a time-varying signal collected at a spatial array of locations: either a regular 3D array as in blood oxygenation level dependent (BOLD) functional magnetic resonance imaging (fMRI) or an irregular 2D array as in magnetoencephalography and electroencephalography. Each signal represents the time course of brain activity at one location in the brain. A key scientific problem of interest is to characterize functional connectivity: spatiotemporal patterns of time course similarity across the spatial locations.
In network-based approaches to this problem, groupings of spatial locations are represented as nodes in a graph, and edges between nodes represent a high level of time series similarity between the constituent nodes. We consider connectivity at the local level, where the nodes are individual voxels within a specific brain region (Meskaldji et al., 2011; Kaiser, 2011), complementing previous quantifications of short-range connectivity such as regional homogeneity (Zang et al., 2004) and local functional connectivity density mapping (Tomasi and Volkow, 2010). Similarity is quantified by the Pearson correlation, as is common in fMRI analyses (Achard et al., 2006; Bassett and Bullmore, 2006; Buckner et al., 2009; Tomasi and Volkow, 2011; van den Heuvel et al., 2008; Worsley et al., 2005; Zalesky et al., 2012). Once such a graph is constructed, functional connectivity can be quantified in terms of a wide variety of summary measures of graph characteristics, which we refer to as network connectivity measures (Sporns, 2011). These measures are useful for identifying common topological properties in functional brain networks. In addition, it is of great interest to study relationships between the connectivity measures and age, cognitive functioning, genetics, and physical health of the individuals receiving fMRI scans, thus investigating the relationships between brain organization and other characteristics of the individual.
Because the similarity between two nodes in a network is measured on a continuous scale, network edges are often instantiated by comparing these similarities with a threshold (Buckner et al., 2009; Tomasi and Volkow, 2011; van den Heuvel et al., 2008). When network connectivity measures are used to compare the networks of different subjects, the prevailing approach is to use subject-specific thresholds so that the resulting networks have the same density, that is, an equal number of edges (Bassett et al., 2012; Hosseini et al., 2012a; Van Wijk et al., 2010). While some studies have conducted network analyses by choosing a single density (Bassett et al., 2009), it is more common to construct and compare networks over a range of density values (Bassett et al., 2006; Ginestet et al., 2011; Hosseini and Kesler, 2013; Hosseini et al., 2012a; Klimm et al., 2014; Lynall et al., 2010; Siebenhühner et al., 2013; Singhet al., 2013; Yu et al., 2011). In this case, the connectivity measure used to summarize the network is a function of the density, so that methods of functional data analysis (FDA) are applicable [for an introduction to FDA, see, e.g., Ramsay and Silverman (2005)]. The usefulness of these methods in connectivity studies was first observed in Bassett et al. (2012).
However, the full power of functional data analysis in connectivity and other neuroimaging studies is yet to be utilized, especially in the context of relating connectivity to other variables. Specifically, the ubiquitous approach for comparing two groups (e.g., healthy/diseased or young/elderly) is to compare their mean connectivity curves through a permutation test (Bassett et al., 2012; Hosseini and Kesler, 2013; Hosseini et al., 2012a; Klimm et al., 2014; Siebenhühner et al., 2013; Singh et al., 2013). An alternative is the so-called “area under the curve” or cost integration approach (Ginestet et al., 2011), although there is a consensus that the resulting inference is less powerful as it is less sensitive to curve shape. While such tests of group mean differences have yielded interesting scientific findings, they are insufficient for identifying differences in variance, for example, or, more importantly, for inferring relationships with a continuous covariate such as age. For example, it is of great interest to study the magnitude and direction of associations between individual connectivity and various outcomes, such as test scores of episodic memory, executive function, and other cognitive measures.
Since permutation tests for group differences are inadequate to address these questions, we propose to use functional principal component analysis (FPCA), which transforms the connectivity of each subject into a subject-specific vector of so-called functional principal component (FPC) scores. In turn, this allows one to formulate regression models where these scores that quantify subject-specific connectivity serve as predictors or responses. Another advantage of these subject-specific FPC scores is the enhanced ability to visualize the variability in connectivity between subjects. For example, examining the joint and marginal distributions of the FPC scores can aid in detecting subgroups and identifying outliers, among other applications, which are extremely difficult tasks to perform with the raw sample of curves. In addition, the effects of the FPC scores on curve shape can be easily visualized, lending interpretability to the regression models.
Due to the known effect of normal aging on connectivity (Betzel et al., 2014; Cao et al., 2014; Ferreira and Busatto, 2013), differences in connectivity between disease and control groups are only meaningful if these groups are age matched. For data that are not age matched, there is no convenient method for incorporating age as a nuisance covariate when testing for group differences in mean curves. However, when implementing the proposed FPCA regression approach to investigate the effects of network connectivity on cognitive performance, one must consider that cognitive performance also declines with age. In this setting, the confounding effect of age can be adjusted for easily by including age as covariate in the regression model. We describe the different aspects of FPCA regression analysis in the Materials and Methods section and demonstrate its various advantages, including the convenience of the adjustment for age and other covariates, in the Results section.
Materials and Methods
Participants
This study included 341 elderly participants in a longitudinal study of cognitive impairment that has been described previously (Hinton et al., 2010). All participants were evaluated within the research program of the University of California, Davis Alzheimer's Disease Center (UCD ADC). Clinical evaluation of this cohort, including the neuropsychological test battery, has been described previously (He et al., 2012). Tables 1 and 2 contain statistical summaries of the groups under comparison in the Group Differences section.
Table 1.
Statistical Summaries of Episodic Memory Groups
| Group | Low | High |
|---|---|---|
| Gender | ||
| Female | 76 | 99 |
| Male | 85 | 45 |
| Education, years | ||
| Mean (SD) | 13.23 (4.56) | 13.88 (4.14) |
| Age | ||
| Mean (SD) | 78.47 (7.68) | 76.51 (6.96) |
| Ethnicity | ||
| White | 82 | 71 |
| Hispanic | 44 | 29 |
| African American | 25 | 34 |
| Other | 10 | 10 |
| Episodic memory score | ||
| Mean (SD) | −1.07 (0.52) | 0.48 (0.64) |
Table 2.
Statistical Summaries of Executive Function Groups
| Group | Low | High |
|---|---|---|
| Gender | ||
| Female | 80 | 98 |
| Male | 74 | 56 |
| Education, years | ||
| Mean (SD) | 12.63 (4.52) | 14.37 (4.17) |
| Age | ||
| Mean (SD) | 78.70 (7.83) | 76.37 (6.69) |
| Ethnicity | ||
| White | 64 | 90 |
| Hispanic | 46 | 29 |
| African American | 28 | 31 |
| Other | 16 | 4 |
| Episodic memory score | ||
| Mean (SD) | −0.86 (0.51) | 0.31 (0.42) |
fMRI acquisition
Participants received one fMRI scan (He et al., 2012) at the UC Davis Imaging Research Center on a 1.5T GE Signa Horizon LX Echospeed system. Participants received an 8-min axial echo-planar imaging BOLD fMRI scan and were provided with no specific instructions before the acquisition other than to keep their eyes open. Scan parameters were as follows: TR 2.0 sec, TE 40 msec, FOV 22 cm, Flip angle 90°, 24 5-mm thick contiguous slices with bandwidth 62.5 KHz, and 64 × 64 matrix with R-L frequency encode direction. This sequence provided 240 time points of data at each voxel.
fMRI preprocessing
The preprocessing steps for each scan include correction for differences in slice timing, correction for head motion, and coregistration to the subject's 3DT1 MRI scan. Multiple linear regression was applied to the signal at each voxel to remove the global linear trend (to account for signal drift) along with two other global signals corresponding to cerebral spinal fluid and white matter. Finally, each signal was band-pass filtered to preserve frequency components between 0.01 and 0.08 Hz. Temporal and spatial processing of the fMRI data were performed in Statistical Parametric Mapping (SPM8, www.fil.ion.ucl.ac.uk/spm) and Resting-State fMRI Data Analysis Toolkit V1.8 (REST1.8, http://restfmri.net/forum/?q=rest). The first four time points were discarded to eliminate nonequilibrium effects of magnetization. Time points with large head motion, defined as translation greater than 1.5 mm and/or rotation greater than 1.5°, were then identified, and participants with any such time points were excluded, resulting in a total of 341 participants for this study.
Graph construction and connectivity measures
In our experiments, we considered connectivity within local hubs corresponding to 10 anatomic regions previously identified in Buckner et al. (2009) as functional connectivity hubs. The identification of these hubs, or regions with an abnormally large number of functional connections, is important to understand the integration of distinct processes and networks in the brain, as well as their relationships with disease, for example. A natural follow-up question is how to quantify the strength or integrity of a particular hub at the subject or group level and how to relate it to external variables such as age or cognitive function. To address this in the current study, for each of the 10 hubs mentioned, a local network was constructed by isolating a 11 × 11 × 11 cube of voxels centered at the corresponding seed voxel listed in Table 3 of Buckner et al., 2009; nongray matter voxels were then discarded using a template. This resulted in 10 networks for each subject, with the number of nodes ranging between 610 and 1331 voxels. We remark that, while distinct hub locations may differ in the number of network nodes, the number of nodes in any particular hub is the same across all subjects. This is an important consideration, since it is known that networks with an unequal number of nodes cannot be compared equitably (Van Wijk et al., 2010).
For each of these sets of voxels, we used the Pearson correlation between the signals at two voxels as a similarity measure. In previous work (Ginestet et al., 2011), it has been noted that positive correlations imply different biological processes than negative correlations. In our analyses, we focused on positive correlations only and, accordingly, all negative correlations were effectively set to 0, as has been done in other studies (Bruno et al., 2012; Fan et al., 2011; He et al., 2007; Hosseini et al., 2012b). Denoting by N the total number of possible connections and by M the number of positive correlations in the network, for a given graph density value 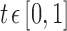 , the largest R correlations were identified, where R is the largest integer such that
, the largest R correlations were identified, where R is the largest integer such that 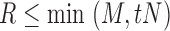 . The network at density t is then constructed by instantiating R edges corresponding to these correlations. The global network connectivity measures (Rubinov and Sporns, 2009) of small-worldness, average characteristic path length, average clustering coefficient, and average efficiency were then computed using the graph-theoretical analysis toolbox (GAT) for Matlab (Hosseini et al., 2012a).
. The network at density t is then constructed by instantiating R edges corresponding to these correlations. The global network connectivity measures (Rubinov and Sporns, 2009) of small-worldness, average characteristic path length, average clustering coefficient, and average efficiency were then computed using the graph-theoretical analysis toolbox (GAT) for Matlab (Hosseini et al., 2012a).
For each of the 10 distinct networks, the range of network densities considered was an interval 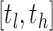 . In this study, tl was the smallest possible density so that every node had at least one connection for all subjects. By considering the total number of positive correlations, the maximum possible density for each subject was calculated, and th was taken as the smallest of these maximum densities. Thus, the range of densities considered is the largest such that all networks are fully connected (all nodes have a connection) and also unsaturated (not all edges are present). In addition, we remark that, while the networks are discrete in nature, the large number of nodes N renders the resulting network connectivity curves approximately continuous and smooth.
. In this study, tl was the smallest possible density so that every node had at least one connection for all subjects. By considering the total number of positive correlations, the maximum possible density for each subject was calculated, and th was taken as the smallest of these maximum densities. Thus, the range of densities considered is the largest such that all networks are fully connected (all nodes have a connection) and also unsaturated (not all edges are present). In addition, we remark that, while the networks are discrete in nature, the large number of nodes N renders the resulting network connectivity curves approximately continuous and smooth.
Before carrying out the analyses, subjects with poor data quality (e.g., presence of constant signals after preprocessing) were identified and removed. For each subject, the percentage of signals that were constant was computed separately for each of the 10 hubs. If any of these percentages were greater than 1%, the individual was considered an outlier and removed. This resulted in the removal of 9 subjects, leaving 332 subjects for the analysis.
Functional principal component analysis
For a fixed hub, let 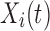 be the value of a particular network connectivity measure for the network of the i th subject with density
be the value of a particular network connectivity measure for the network of the i th subject with density 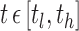 . In order for these functions to be used in subsequent analysis, these infinite-dimensional functional objects are reduced to a sequence of one or more scalar values. In this study, Xi is a random function with pointwise mean
. In order for these functions to be used in subsequent analysis, these infinite-dimensional functional objects are reduced to a sequence of one or more scalar values. In this study, Xi is a random function with pointwise mean 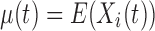 and covariance
and covariance 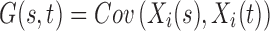 . According to the Karhunen–Loève expansion, Xi can be written as
. According to the Karhunen–Loève expansion, Xi can be written as
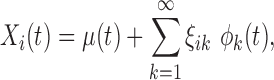 |
where the functions 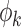 form an orthonormal basis and are eigenfunctions of the integral operator
form an orthonormal basis and are eigenfunctions of the integral operator 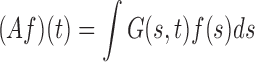 , and
, and 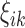 are the FPC scores given by
are the FPC scores given by 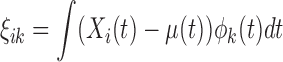 . The eigenvalues
. The eigenvalues 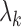 corresponding to the eigenfunctions
corresponding to the eigenfunctions 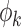 are positive and decreasing in k. Hence, like standard principal component analysis, the FPC scores are uncorrelated and give an optimal representation of the data in terms of accounting for the total variability in the function Xi. In fact, one way of viewing FPCA is that it learns the linear representation for the data that minimizes the amount of lost information when truncated to a finite number of terms.
are positive and decreasing in k. Hence, like standard principal component analysis, the FPC scores are uncorrelated and give an optimal representation of the data in terms of accounting for the total variability in the function Xi. In fact, one way of viewing FPCA is that it learns the linear representation for the data that minimizes the amount of lost information when truncated to a finite number of terms.
To reduce the function Xi to a finite sequence of scores 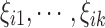 , the truncation point K can be chosen so as to account for a particular percentage of the total variability. In our experiments, this was set at 95%. For all hubs and network connectivity measures, this resulted in reducing the sample of functions to just the first FPC score. For more information on FPCA, and the estimation of the FPC scores from a sample, see Müller (2011).
, the truncation point K can be chosen so as to account for a particular percentage of the total variability. In our experiments, this was set at 95%. For all hubs and network connectivity measures, this resulted in reducing the sample of functions to just the first FPC score. For more information on FPCA, and the estimation of the FPC scores from a sample, see Müller (2011).
Relationships between network connectivity and cognition
The goal of the experiments in the Results section is to relate the network connectivity measures to scientifically relevant outcomes of interest: two continuous standardized measures of cognitive functioning that assess episodic memory and executive function. We compare traditional group difference methodology and the proposed FPCA technique in their ability to capture and quantify these associations.
For each of the 10 hubs considered for network construction, we obtained samples of network connectivity measure functions, as outlined in the Graph Construction and Connectivity Measures section, using all 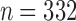 subjects. These functions were reduced to a single number, the first FPC score, as described in the Functional Principal Component Analysis section. To assess the relationship between the connectivity measures within the given hub and the two cognitive measurements of interest, we considered the standard multiple linear regression model with the first FPC score and age as predictors. The models were fitted using ordinary least squares, and the null hypothesis that the connectivity measure has no effect was tested using the standard partial F-test, after checking the assumptions of homoscedasticity and Gaussianity of the errors. As some subjects had missing values for the cognitive scores (27 for episodic memory and 24 for executive function), such subjects were removed from the regression analyses. As a comparison, for each of the two cognitive scores, the subjects were divided into low- and high-performing groups by a median split. For statistical summaries of these groups, see Tables 1 and 2.
subjects. These functions were reduced to a single number, the first FPC score, as described in the Functional Principal Component Analysis section. To assess the relationship between the connectivity measures within the given hub and the two cognitive measurements of interest, we considered the standard multiple linear regression model with the first FPC score and age as predictors. The models were fitted using ordinary least squares, and the null hypothesis that the connectivity measure has no effect was tested using the standard partial F-test, after checking the assumptions of homoscedasticity and Gaussianity of the errors. As some subjects had missing values for the cognitive scores (27 for episodic memory and 24 for executive function), such subjects were removed from the regression analyses. As a comparison, for each of the two cognitive scores, the subjects were divided into low- and high-performing groups by a median split. For statistical summaries of these groups, see Tables 1 and 2.
Results
Group differences
For both episodic memory and executive function, the low- and high-performing groups were compared for each combination of hub and network connectivity measure, for a total of 40 tests per cognitive score. After correcting these p-values using false discovery rate (FDR) with 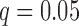 (this correction was made separately for the two cognitive scores), there were no significant findings. Notably, in the right middle temporal (RMT) hub, which was found to be significant in the FPCA regression analyses detailed below, the low- and high-performing episodic memory groups had virtually identical mean curves for all network measures (Fig. 1) and similarly for the groups defined by executive function performance.
(this correction was made separately for the two cognitive scores), there were no significant findings. Notably, in the right middle temporal (RMT) hub, which was found to be significant in the FPCA regression analyses detailed below, the low- and high-performing episodic memory groups had virtually identical mean curves for all network measures (Fig. 1) and similarly for the groups defined by executive function performance.
FIG. 1.

Average network connectivity curves in the RMT hub for low (solid line) and high (dashed line) episodic memory score groups. Permutation tests revealed no significant difference, as the means are virtually identical, while the FPCA analyses were significant. Corresponding curve comparisons for groups defined by executive function are indistinguishable, so they are not shown in this study. FPCA, functional principal component analysis; RMT, right middle temporal.
Functional principal component analysis
For each of the four network connectivity measures, the distribution of connectivity functions in the sample for the RMT hub is visualized in Figure 2 by a functional version of the box plot (Hyndman and Shang, 2012). Figure 3 shows the mean network connectivity curves for each measure for all subjects combined, as opposed to the group means in Figure 1. Plots for the other hubs can be done similarly; these are shown due to the significance of this hub in the ensuing regression models. The minimum and maximum densities for this hub were 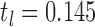 and
and 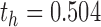 .
.
FIG. 2.

Functional box plot of network connectivity curves in the RMT hub as a function of density. Similar to the box in a standard box plot, the gray region marks the area covered by the middle 50% of curves. The dashed line corresponds to the “median” curve, while the “whiskers” correspond to the dash-dotted lines.
FIG. 3.

Average network connectivity curves in the RMT hub using all subjects.
Unlike group differences, where the mean curve in each group is most relevant (Fig. 1), the important objects in FPCA are the eigenfunctions 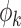 and the FPC scores
and the FPC scores 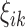 . The eigenfunctions for all network connectivity measures reflect higher variability in connectivity for low densities, since each eigenfunction gets closer to zero for high densities (Fig. 4). In addition, since these functions represent variability of the connectivity around the corresponding mean functions (Fig. 3), the fact that the eigenfunctions are strictly positive leads to the conclusion that high FPC scores for each network measure are associated with connectivity curves that are above average in the sample, that is, they lie above the mean function. This is especially so for lower densities.
. The eigenfunctions for all network connectivity measures reflect higher variability in connectivity for low densities, since each eigenfunction gets closer to zero for high densities (Fig. 4). In addition, since these functions represent variability of the connectivity around the corresponding mean functions (Fig. 3), the fact that the eigenfunctions are strictly positive leads to the conclusion that high FPC scores for each network measure are associated with connectivity curves that are above average in the sample, that is, they lie above the mean function. This is especially so for lower densities.
FIG. 4.

First FPCA eigenfunction for each network connectivity measure in the RMT hub. The first eigenfunctions for path length, efficiency, small worldness, and clustering accounted for 98%, 97%, 96%, and 99% of the total variability, respectively.
Similar to group differences, 40 regression models were fit for each cognitive score, with age included as one of the predictors. For each cognitive score, the 40 p-values were corrected using FDR with 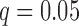 . Four significant relationships were discovered, all within the RMT hub. After accounting for age, the first FPC score for path length is positively correlated with episodic memory (slope = 6.53, FDR-corrected p-value = 0.0075) and also with executive function (slope = 4.64, FDR-corrected p-value = 0.0164), while the first FPC score for efficiency is negatively correlated with episodic memory (slope = −41.69, FDR-corrected p-value = 0.0075) and executive function (slope = −30.32, FDR-corrected p-value = 0.0164). Given the interpretation of the eigenfunctions, this shows that above average path length and below average efficiency in this hub are associated with superior episodic memory and executive function.
. Four significant relationships were discovered, all within the RMT hub. After accounting for age, the first FPC score for path length is positively correlated with episodic memory (slope = 6.53, FDR-corrected p-value = 0.0075) and also with executive function (slope = 4.64, FDR-corrected p-value = 0.0164), while the first FPC score for efficiency is negatively correlated with episodic memory (slope = −41.69, FDR-corrected p-value = 0.0075) and executive function (slope = −30.32, FDR-corrected p-value = 0.0164). Given the interpretation of the eigenfunctions, this shows that above average path length and below average efficiency in this hub are associated with superior episodic memory and executive function.
Discussion
We have broadened the application of functional data analysis tools in neuroimaging studies by utilizing FPCA to fit regression models with cognitive scores as the dependent variable, as opposed to simple two-sample tests for detecting group differences. There are three main findings.
First, these models are an improvement in terms of their power to detect significant relationships between connectivity measures and continuous measures of cognitive function. Even after controlling for age and multiple testing, the FPCA regressions found four significant associations, whereas group comparisons did not reveal any significance.
Second, by interpreting the FPC scores using their corresponding eigenfunction, it is possible to identify shape characteristics in subject-specific network connectivity curves that relate to the desired outcomes; this aspect is absent in testing for group differences. In our experiments, we found that above average path length and below average efficiency of local connectivity within the RMT hub, particularly for smaller network densities, are positively associated with episodic memory and executive function. The importance of this hub with regard to episodic memory and executive function is consistent with the existing cognitive neuroscience literature (Banich et al., 2000; Milham et al., 2003a,b; Nyberg et al., 1996; Ragland et al., 2004; Spaniol et al., 2009; van Veen and Carter, 2005). To our knowledge, however, the specific relationship to these network connectivity measures is novel, as is the nature of the relationship. The associations between graph metrics (efficiency and path length) in this local region and cognitive function (episodic memory and executive function) may appear counterintuitive at first sight. However, we remark that previous studies have identified regions in which local connectivity increases with age [Ferreira and Busatto (2013) and references therein], providing further evidence that stronger local connectivity may be linked to cognitive decline.
Finally, nuisance variables such as age can easily be controlled in the FPCA regression models, whereas analysis based on group differences does not readily allow to adjust for such factors. While age is a prominent and ubiquitous example of a nuisance covariate, there are many others that matter in neuroscience studies, such as medication doses and education levels.
As mentioned in the Introduction the utility of FPCA for quantifying connectivity extends beyond regression. As an example, scatterplots of FPC scores can provide interesting visualizations of the distribution of network connectivity curves, which could be useful for subgroup detection, classification, and differentiating groups by variability, among other statistical techniques.
The key strength of this study is its thorough experimental evaluation of samples of functional connectivity networks through FPCA regression, as well as comparison with traditional group difference testing, on a large, real-world fMRI data set that included state-of-the-art measurement of cognitive functioning in two relevant domains. One potential limitation of our approach is that there is no guarantee that FPCA will provide eigenfunctions that are easy to interpret. In our experiments, the eigenfunctions were relatively easy to interpret in terms of what characteristics of connectivity change the eigenfunction captures over the range of densities. This was mostly due to the smooth nature of the underlying samples of curves. However, FPCA may feature more complex eigenfunctions that are harder to interpret.
Future work of interest will move beyond FPCA to more advanced functional data analysis methods, especially methods of dimensionality reduction. One such promising method is the ISOMAP (Tenenbaum et al., 2000), which has been extended to functional data and can often provide insightful interpretations for samples of functions. In addition, there are many other network connectivity measures besides the four considered in our experiments that may be useful in characterizing these networks for the purpose of relating connectivity to other observed variables. Finally, while we have focused on a small subset of local brain networks, the same approach can be used for any network-based connectivity study.
Acknowledgments
This work was supported by National Science Foundation grant DMS-1228369 and DMS-1407852, National Institutes of Health grants AG10220, AG10129, AG 030514, AG031252, and AG021028, and California Department of Public Health Alzheimer's Disease Program Contracts 06-55311 and 06-55312.
Author Disclosure Statement
No competing financial interests exist.
References
- Achard S, Salvador R, Whitcher B, Suckling J, Bullmore E. 2006. A resilient, low-frequency, small-world human brain functional network with highly connected association cortical hubs. J Neurosci 26:63–72 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Banich M, Milham M, Atchley R, Cohen N, Webb A, Wszalek T, et al. 2000. fMRI studies of stroop tasks reveal unique roles of anterior and posterior brain systems in attentional selection. J Cogn Neurosci 12:988–1000 [DOI] [PubMed] [Google Scholar]
- Bassett DS, Bullmore E. 2006. Small-world brain networks. Neuroscientist 12:512–523 [DOI] [PubMed] [Google Scholar]
- Bassett DS, Bullmore ET, Meyer-Lindenberg A, Apud JA, Weinberger DR, Coppola R. 2009. Cognitive fitness of cost-efficient brain functional networks. Proc Natl Acad Sci U S A 106:11747–11752 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bassett DS, Meyer-Lindenberg A, Achard S, Duke T, Bullmore E. 2006. Adaptive reconfiguration of fractal small-world human brain functional networks. Proc Natl Acad Sci U S A 103:19518–19523 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bassett DS, Nelson BG, Mueller BA, Camchong J, Lim KO. 2012. Altered resting state complexity in schizophrenia. Neuroimage 59:2196–2207 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Betzel RF, Byrge L, He Y, Goñi J, Zuo X-N, Sporns O. 2014. Changes in structural and functional connectivity among resting-state networks across the human lifespan. Neuroimage 102:345–357 [DOI] [PubMed] [Google Scholar]
- Bruno J, Hosseini SH, Kesler S. 2012. Altered resting state functional brain network topology in chemotherapy-treated breast cancer survivors. Neurobiol Dis 48:329–338 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buckner RL, Sepulcre J, Talukdar T, Krienen FM, Liu H, Hedden T, et al. 2009. Cortical hubs revealed by intrinsic functional connectivity: mapping, assessment of stability, and relation to Alzheimer's disease. J Neurosci 29:1860–1873 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cao M, Wang JH, Dai ZJ, Cao XY, Jiang LL, Fan FM, et al. 2014. Topological organization of the human brain functional connectome across the lifespan. Dev Cogn Neurosci 7:76–96 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fallani FDV, Richiardi J, Chavez M, Achard S. 2014. Graph analysis of functional brain networks: practical issues in translational neuroscience. Philos Trans R Soc B Biol Sci 369:20130521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fan Y, Shi F, Smith JK, Lin W, Gilmore JH, Shen D. 2011. Brain anatomical networks in early human brain development. Neuroimage 54:1862–1871 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferreira LK, Busatto GF. 2013. Resting-state functional connectivity in normal brain aging. Neurosci Biobehav Rev 37:384–400 [DOI] [PubMed] [Google Scholar]
- Ginestet CE, Nichols TE, Bullmore ET, Simmons A. 2011. Brain network analysis: separating cost from topology using cost-integration. PLoS One 6:e21570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- He J, Carmichael O, Fletcher E, Singh B, Iosif A-M, Martinez O, et al. 2012. Influence of functional connectivity and structural MRI measures on episodic memory. Neurobiol Aging 33:2612–2620 [DOI] [PMC free article] [PubMed] [Google Scholar]
- He Y, Chen ZJ, Evans AC. 2007. Small-world anatomical networks in the human brain revealed by cortical thickness from MRI. Cereb Cortex 17:2407–2419 [DOI] [PubMed] [Google Scholar]
- Hinton L, Carter K, Reed BR, Beckett L, Lara E, DeCarli C, Mungas D. 2010. Recruitment of a community-based cohort for research on diversity and risk of dementia. Alzheimer Dis Assoc Disord 24:234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hosseini S, Hoeft F, Kesler SR. 2012a. GAT: a graph-theoretical analysis toolbox for analyzing between-group differences in large-scale structural and functional brain networks. PLoS One 7:e40709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hosseini SH, Kesler SR. 2013. Comparing connectivity pattern and small-world organization between structural correlation and resting-state networks in healthy adults. Neuroimage 78:402–414 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hosseini SH, Koovakkattu D, Kesler SR. 2012b. Altered small-world properties of gray matter networks in breast cancer. BMC Neurol 12:28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hyndman RJ, Shang HL. 2012. Rainbow plots, bagplots, and boxplots for functional data. J Comp Graph Stat 19:29–45 [Google Scholar]
- Kaiser M. 2011. A tutorial in connectome analysis: topological and spatial features of brain networks. Neuroimage 57:892–907 [DOI] [PubMed] [Google Scholar]
- Klimm F, Bassett DS, Carlson JM, Mucha PJ. 2014. Resolving structural variability in network models and the brain. PLoS Comput Biol 10:e1003491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lynall M-E, Bassett DS, Kerwin R, McKenna PJ, Kitzbichler M, Muller U, Bullmore E. 2010. Functional connectivity and brain networks in schizophrenia. J Neurosci 30:9477–9487 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meskaldji DE, Ottet M-C, Cammoun L, Hagmann P, Meuli R, Eliez S, et al. 2011. Adaptive strategy for the statistical analysis of connectomes. PLoS One 6:e23009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Milham M, Banich M, Claus E, Cohen N. 2003a. Practice-related effects demonstrate complementary roles of anterior cingulate and prefrontal cortices in attentional control. Neuroimage 18:483–493 [DOI] [PubMed] [Google Scholar]
- Milham MP, Banich MT, Barad V. 2003b. Competition for priority in processing increases prefrontal cortex's involvement in top-down control: an event-related fMRI study of the stroop task. Brain Res Cogn Brain Res 17:212–222 [DOI] [PubMed] [Google Scholar]
- Müller H-G. 2011. Functional data analysis. In: Lovric M. (ed.) International Encyclopedia of Statistical Science. Heidelberg: Springer; p. 554.. [Extended version available in StatProb: the Encyclopedia Sponsored by Statistics and Probability Societies, id 242]. [Google Scholar]
- Nyberg L, McIntosh AR, Cabeza R, Habib R, Houle S, Tulving E. 1996. General and specific brain regions involved in encoding and retrieval of events: what, where, and when. Proc Natl Acad Sci U S A 93:11280–11285 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ragland JD, Gur RC, Valdez J, Turetsky BI, Elliott M, Kohler C, et al. 2004. Event-related fMRI of frontotemporal activity during word encoding and recognition in schizophrenia. Am J Psychiatry 161:1004–1015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramsay JO, Silverman BW. 2005. Functional Data Analysis, 2nd ed. Springer Series in Statistics, New York: Springer [Google Scholar]
- Rubinov M, Sporns O. 2009. Complex network measures of brain connectivity: uses and interpretations. Neuroimage 52:1059–1069 [DOI] [PubMed] [Google Scholar]
- Siebenhühner F, Weiss SA, Coppola R, Weinberger DR, Bassett DS. 2013. Intra-and inter-frequency brain network structure in health and schizophrenia. PLoS One 8:e72351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singh MK, Kesler SR, Hosseini SH, Kelley RG, Amatya D, Hamilton JP, et al. 2013. Anomalous gray matter structural networks in major depressive disorder. Biol Psychiatry 74:777–785 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spaniol J, Davidson PS, Kim AS, Han H, Moscovitch M, Grady CL. 2009. Event-related fMRI studies of episodic encoding and retrieval: meta-analyses using activation likelihood estimation. Neuropsychologia 47:1765–1779 [DOI] [PubMed] [Google Scholar]
- Sporns O. 2011. Networks of the Brain. Cambridge, MA: The MIT Press [Google Scholar]
- Tenenbaum JB, De Silva V, Langford JC. 2000. A global geometric framework for nonlinear dimensionality reduction. Science 290:2319–2323 [DOI] [PubMed] [Google Scholar]
- Tomasi D, Volkow ND. 2010. Functional connectivity density mapping. Proc Natl Acad Sci U S A 107:9885–9590 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tomasi D, Volkow ND. 2011. Association between functional connectivity hubs and brain networks. Cereb Cortex 21:2003–2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- van den Heuvel M, Mandl R, Pol HH. 2008. Normalized cut group clustering of resting-state fMRI data. PLoS One 3:e2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Veen V, Carter CS. 2005. Separating semantic conflict and response conflict in the stroop task: a functional MRI study. Neuroimage 27:497–504 [DOI] [PubMed] [Google Scholar]
- Van Wijk BC, Stam CJ, Daffertshofer A, et al. 2010. Comparing brain networks of different size and connectivity density using graph theory. PLoS One 5:e13701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Worsley KJ, Chen J-I, Lerch J, Evans AC. 2005. Comparing functional connectivity via thresholding correlations and singular value decomposition. Philos Trans R Soc B Biol Sci 360:913–920 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu Q, Sui J, Rachakonda S, He H, Pearlson G, Calhoun VD. 2011. Altered small-world brain networks in temporal lobe in patients with schizophrenia performing an auditory oddball task. Front Syst Neurosci 5:7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zalesky A, Fornito A, Bullmore E. 2012. On the use of correlation as a measure of network connectivity. Neuroimage 60:2096–2106 [DOI] [PubMed] [Google Scholar]
- Zang Y, Jiang T, Lu Y, He Y, Tian L. 2004. Regional homogeneity approach to fMRI data analysis. Neuroimage 22:394–400 [DOI] [PubMed] [Google Scholar]