

Abstract
Inflammasomes are intracellular multiprotein complexes that comprise part of the innate immune response. Since their definition, inflammasome disorders have been linked to an increasing number of diseases. Autoinflammatory diseases refer to disorders in which local factors lead to the activation of innate immune cells, causing tissue damage when in the absence of autoantigens and autoantibodies. Skin symptoms include the main features of monogenic inflammasomopathies, such as Cryopyrin-Associated Periodic Syndromes (CAPS), Familial Mediterranean Fever (FMF), Schnitzler Syndrome, Hyper-IgD Syndrome (HIDS), PAPA Syndrome, and Deficiency of IL-1 Receptor Antagonist (DIRA). Concepts from other pathologies have also been reviewed in recent years, such as psoriasis, after the recognition of a combined contribution of innate and adaptive immunity in its pathogenesis. Inflammasomes are also involved in the response to various infections, malignancies, such as melanoma, autoimmune diseases, including vitiligo and lupus erythematosus, atopic and contact dermatitis, acne, hidradenitis suppurativa, among others. Inhibition of the inflammasome pathway may be a target for future therapies, as already occurs in the handling of CAPS, through the introduction of IL-1 inhibitors. This study presents a literature review focusing on the participation of inflammasomes in skin diseases.
Keywords: Inflammasomes; Immunity, Innate; Interleukin-1; Hereditary Autoinflammatory Diseases; Sweet's Syndrome
INTRODUCTION
1. THE INFLAMMASOME
Inflammasomes, first described in 2002, are intracellular multiprotein complexes consisting of three elements: a molecular pattern recognition receptor (PRR), an apoptosis-associated specklike protein containing a caspase-recruitment domain (ASC) adaptor protein, and a caspase-1 enzyme.1,2 Inflammasomes are constitutive of the innate immune system, and are responsible for regulating the immunological response to several stimuli, whether exogenous, such as bacterial agents, or endogenous, such as neoplasia, by means of cytokine production and secretion. Inflammasome assembly and activation are triggered by the recognition of stimuli by PPR receptors, which interact with the ASC adaptor molecule. ASC protein is then linked to the procaspase-1, which is then cleaved, activating caspase-1. Activation of caspase-1 culminates in the cleavage of proIL-1β and pro-IL18, resulting in their active forms IL-1β and IL18, respectively, which stimulate the inflammatory response.2,3,4 The activation of capase-1 through the inflammasome pathway is also capable of inducing pyroptosis, a type of inflammatory cell death.5,6
Inflammasomes are named in accordance with their intracellular receptor's nomenclature. Two receptor classes have already been identified: NLR-Nod like receptors, and ALR-AIM2-like receptors. At least 6 types of inflammasomes have been described: NLRP1, NLRP3, NLRP6, NLRP12, NLRC4/IPAF, and AIM-2. The first five are NLR receptors, whereas AIM-2 is an ALR receptor.4,7
Normal inflammasome function, with IL18 and especially IL-1β, synthesis, is required for the innate immune system to act effectively against pathogens (virus, fungi, and bacteria) and against other diseases, including some types of cancer. When its activity becomes excessive, deleterious effects may occur in the organism. Since their definition, inflammasome disorders have been implicated in several pathologies, becoming a possible target for therapeutic action.8 Hereditary fever syndromes represented the group of diseases initially associated with inflammasomes.9 In addition to these, other syndromes have recently had their physiopathogenesis associated with inflammasomes, such as: metabolic diseases (type 2 diabetes, obesity, arteriosclerosis),10 neurological disorders (Alzheimer's, multiple sclerosis),11,12 autoimmune diseases (vitiligo, rheumatoid arthritis, type 1 diabetes, Addison's disease, lupus erythematosus),13,14 infections (HIV, Francisella tularensis, Legionella pneumophila, Listeria monocytogenes, Mycobacterium tuberculosis, varicella zoster virus, S. pneumoniae),15,16,17,18,19,20 and neoplasia (gastric, hepatic, colorectal, breast, and melanoma).21,22
2. AUTOINFLAMMATION VS. AUTOIMMUNITY
When we speak of autoimmune diseases, we refer to adaptive immune disorders, involving inflammatory diseases with aberrant responses to autoantigens, controlled by B cells and T cells, with the presence of autoantibodies.23
The concept of autoinflammatory disease first appeared in 1999 and gained strength after the inflammasome was defined in 2002. Autoinflammatory diseases refer to disorders in which local factors lead to the activation of innate immune system cells, such as macrophages, neutrophils, mast cells and NK cells, resulting in tissue damage in the absence of autoantigens and autoantibodies. Since then, several diseases, previously described as autoimmune, or not fitting any classifications, have been classified as autoinflammatory, or, at least, as having an autoinflammatory component in their physiopathogenesis. Some pathologies share autoimmune and autoinflammatory characteristics.23,24,25
We will provide here a literature review on skin diseases associated with inflammasomes.
MONOGENIC AUTOINFLAMMATORY DISEASES
Monogenic autoinflammatory diseases constitute a group of rare hereditary syndromes that lead to exacerbated responses by the innate immune system and present some common characteristics among them: recurrent signs of systemic inflammation, such as fevers, with early onset in childhood; heredity, in many cases; and frequent skin manifestations. Among these diseases, inflammasomopathies represent the most important group, and will be described below.26,27
1- CRYOPYRIN-ASSOCIATED PERIODIC SYNDROMES (CAPS)
Cryopyrin-associated periodic syndromes (CAPS) include three rare entities that form a spectrum: familial cold autoinflammatory syndrome, Muckle-Wells syndrome, and neonatal onset multisystem inflammatory disease. These syndromes result from mutations in tbe NLRP3 gene and in the 1q44 chromosome, with dominant autosomal inheritance and variable penetrance, but with some new mutation cases, especially in the syndrome's more serious spectrum.26,28,29,30 Over 90 mutations have been reported.31 This disease's physiopathology is characterized by excessive IL-1β production by macrophages, monocytes, and chondrocytes.32 In an animal model, Nakamura and Kambe demonstrated that dermis mast cells produced histamine, as well as IL-1β, and are mainly responsible for the production of this cytokine.33 Manifestations tend to accompany patients throughout their lives, and prognosis depends mainly on the development of secondary amyloidosis, which predominantly affects the kidneys.34
Familial cold autoinflammatory syndrome, previously called familial cold urticaria, is the least severe form, beginning in the first months of life, with maculopapular or urticarial, usually not pruriginous, lesions that may be painful, associated with fever, chills, myalgia, headaches, arthralgia, and conjunctivitis.28,29,32 Histopathology of lesions evinces a sparse neutrophil infiltrate in the reticular dermis, which may be perivascular or perieccrine.35 Episodes, which tend to be less than 12 hours long, may be triggered by low ambient temperatures. However, local exposures to ice do not trigger the outbreak of lesions, which is different from that which occurs with urticaria and non-familial cold lesions. 28,29,32 The mechanism through which the cold triggers the lesions is unknown. Secondary amyloidosis is rare, and the prognosis regarding afterlife is usually good.28,29
Muckle Wells syndrome presents the same characteristics as the familial cold autoinflammatory syndrome, as well as sensorineural hearing loss during adolescence. In addition, outbreaks may last longer (between 12 and 36 hours). Secondary amyloidosis may occur in up to 25% of the cases.28,29,32 Hence, no association can be drawn between the ambient temperature and the manner in which the outbreaks are triggered.29
The most severe form of CAPS is called neonatal onset multisystem inflammatory disease (NOMID), also known as chronic infantile neurological, cutaneous, and articular syndrome (CINCA). It manifests itself as high fever episodes with maculopapular or urticarial lesions, with persistent exanthema, arthritis, lymphadenopathy, hepatosplenomegaly, and chronic meningitis. It is associated with late physical and mental development, sensorineural hearing loss, and a loss of vision. Exaggerated patella and distal femur osteocartiloginous growth is characteristic, and facial deformities, such as flattened nose tip, macrocephaly, frontal bossing, and protruding eyes, may also occur.26,28,29
The revolution in treatment took place with the introduction of the IL1 receptor antagonist, Anakinra. Response to treatment is usually good, with rapid improvements in the outbreaks. If started early, treatment improves the prognosis as regards the development of sequelae and its evolution into amyloidosis. Two other medications, Rilonacept, a dimeric fusion protein, and Canakinumab, a human monoclonal anti-IL-1β antibody, are other therapeutic options.29,32,36,38
2- FAMILIAL MEDITERRANEAN FEVER
Familial Mediterranean fever (FMF) is an autosomal recessive disease, with incomplete penetrance, due to MEFV mutation on chromosome 16p13.3, responsible for the synthesis of pyrins, leading to the defective inhibition of NLRP3, with increased IL-1 production.28,29,39
Symptoms start before 10 years of age in 65% of the patients, and before 20 years of age in 90% of the patients. It is characterized by high fever episodes for three days in a row, associated with serositis and synovitis. Skin lesions appear in up to 43% of the cases, and the most classical manifestation is erysipeloid erythema, which is characterized by well-defined erythematous-edematous plaques, which present growth on the borders, more frequently on legs and feet. Pruriginous, urticarial lesions, palmoplantar erythema, and alterations similar to Raynaud's phenomenon may also occur. Outbreaks recede spontaneously, and their frequency may range from weeks to months. Between outbreaks, the patient remains asymp-tomatic. Histopathology reveals edema on the dermis, sparse perivascular infiltrate with lymphocytes, neutrophils, and histiocytes. Acute or chronic renal failure, secondary to amyloidosis, may be a severe complication of the disease.28,29
Colchicine is the first line medication for treatment, leading to the reduction of inflammatory symptoms. Anakinra, thalidomide, and anti-TNF-α agents are therapeutic options.38,40
3- HYPER IGD SYNDROME (HIDS)
This syndrome is caused by autosomal recessive mutation in the MVK (mevalonate kinase) gene. Enzyme deficiency culminates in the increased caspase-1 activity and, consequently, in the synthesis of IL-1β.41
Symptoms usually begin in the first year of life, with episodes that last from 3 to 7 days, and that appear every 1 or 2 months, but which tend to become less frequent during adulthood.42 Vaccines, infections, mental stress, trauma, or surgeries may trigger crises.43 Episodes of fever of 40°C or more may occur, and may be associated with abdominal pain, diarrhea, vomiting, headache, lymphadenopathy, arthralgia, and splenomegaly. Up to 80% of patients present skin lesions, usually on the trunk and limbs. Undefined and painful macular, popular, or edematous, lesions are quite common. Erythema nodosum may also occur. Half of the patients develop aphthous ulcers in the mouth, sometimes with associated genital ulcers.28,30
Crises are treated with nonsteroidal anti-inflammatory drugs (NSAIDs) or corticosteroids in high doses. Dapsone is an option. IL-1 antagonists and anti-TNF may be useful in reducing symptoms.26,29
4- PAPA SYNDROME: PYOGENIC ARTHRITIS, PYODERMA GANGRENOSUM, AND ACNE
Pyogenic arthritis, pyoderma gangrenosum, and acne syndrome (PAPA syndrome) is a rare autosomal dominant disease, with mutation of the PSTPIP1/CD2BP1 gene on chromosome 15q. A deficiency in the inhibition of caspase-1 activation leads to an excessive production of IL-1β.29,44 A sporadic PAPA syndrome case, without mutation in the PSTPIP1/CD2BP1 gene has been described in prior literature.45
Clinically, these syndromes appear in recurrent sterile arthritis in early childhood, which may progress with deformities. Pyoderma gangrenosum lesions occur mainly on the distal portion of one's limbs, but cases with multiple disseminate lesions have also been reported. Skin abscesses may also occur. Cystic acne and hidradenitis occur during adolescence.29,30
The therapeutic response has varied in previosly reported cases, with IL-1 inhibitors being more effective to combat articular symptoms, and anti-TNF being more effective for skin conditions.30
5- SCHNITZLER SYNDROME
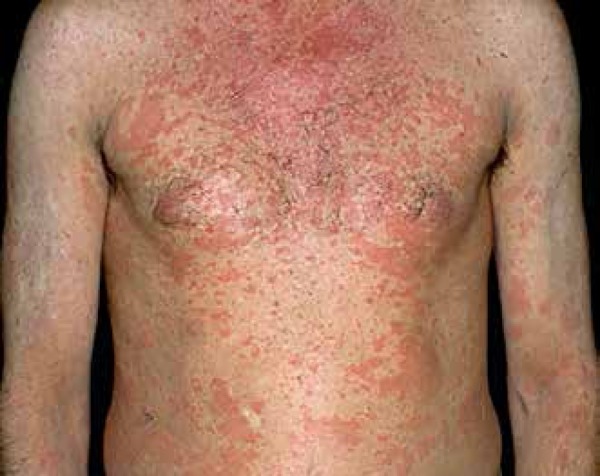
Classification of this syndrome as an autoinflammatory disease is still controversial. Some cases have been associated with autosomal dominant mutations on the NLRP3 gene, which could place it within the CAPS group, whereas, in other patients, this mutation has not been found.29,46,47
The presence of monoclonal gammopathy by IgM and the typical skin condition of a neutrophilic urticarial dermatosis are essential for the diagnosis, associated with at least two of the following criteria: arthralgia or arthritis, bone pain, palpable lymph nodes, hepatic or splenomegaly, increased erythrocyte sedimentation rate (ESR), leukocytosis, and abnormal findings in bone morphological investigations. Typical dermatological lesions (Figure 1) are usually the first symptom and are characterized by macules, papules, or erythematous plaques, with little or no prurigo, which disappear within hours and do not leave scars.48
Figure 1.
Schnitzler syndrome
6- DEFICIENCY OF THE IL-1 RECEPTOR ANTAGONIST (DIRA)
This disease was first described in 2009 in nine patients who had presented sterile multifocal osteomyelitis, periostitis, and pustulosis since the neonatal period, but without fever. The patients presented a homozygotic mutation of the IL1 receptor antagonist (IL1RA).49 The lack of opposition to IL-1 signaling leads to a severe condition of systemic inflammation, with a high mortality rate if not treated. Patients that received treatment with Anakinra witnessed a swift improvement in their symptoms. Cutaneous manifestations vary from discrete pustulous lesions to a condition of pustules covering the body, in addition to ichthiosiform lesions. Some patients present changes in their fingernails (pittings and onicomadesis) or ulcers in the oral mucosa. 49,51
DERMATOLOGICAL DISEASES ASSOCIATED WITH INFLAMMASOMES
1. NEUTROPHILIC DERMATOSES
Sweet's syndrome (Figure 2) and pyoderma gangrenosum are considered to be the prototypes of neutrophilic dermatoses, a group of diseases with the etiopathogenesis that has yet to be fully clarified. Their direct relation with inflammasomes has not yet been investigated. However, some evidence raises this hypothesis, such as the possible participation of the IL-1β and its similarities with autoinflammatory diseases. All monogenic autoinflammatory diseases present cutaneous symptoms, and these coincide with lesions stemming from neutrophilic dermatoses.52
Figure 2.
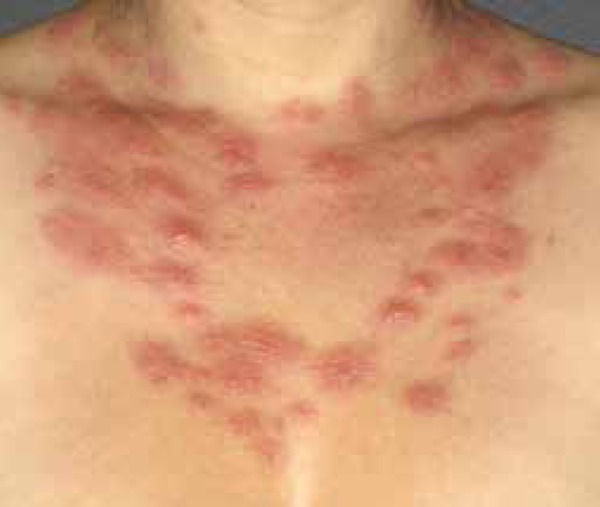
Sweet's syndrome
In a case series of 22 patients with autoinflammatory diseases associated with the mutation of the NLRP3 inflammasome, one of these patients was diagnosed with associated Sweet's syndrome.53 In two case reports of patients diagnosed with Sweet's syndrome, and who had been unaffected by various therapies, improvement was observed after treatment with Anakinra, an IL1RA, which is the medication of choice for CAPS and other autoinflammatory diseases.54,55
Mutations in the PSTPIP1 gene, the same associated with PAPA syndrome, were identified in patients with isolated pyoderma gangrenosum.56
Wallach, who in 1991 proposed criteria for the classification of neutrophilic dermatoses, in 2015 suggested the hypothesis that the neutrophilic dermatoses would represent the cutaneous expression of autoinflammation.52
2. PSORIASIS
The classification of psoriasis as an autoimmune disease itself presents limited evidence57 and has been questioned due to the absence of specific antibodies and defined autoantigens, to the non-activiation of B cells, and to the absence of genetic risk factors that are common in other autoimmune conditions. The presence of neutrophils in lesions and the participation of the innate immune response are characteristics that are similar between psoriasis and the group of autoinflammatory diseases.58
IL-1β and IL-18 play a key role in many inflammatory diseases, including psoriasis. The association of the inflammasome with psoriasis has been investigated due to its relationship with pro-inflammatory cytokines. Polymorphisms of NLRP1, NLRP3, and CARD8, negative regulators of the caspase-1 activity, were associated with the susceptibility of psoriasis.59,60 Johansen et al. reported a greater activity of caspase-1 in psoriasis lesions when compared with non-lesioned skin. 61 Later, Salskov-Iversen et al. observed an increased expression of caspase-5 in affected skin.62 Dombrowski et al. demonstrated an augmented expression of the AIM2-inflammasome in the keratinocytes of affected skin, associated with the presence of cytosolic DNA, this latter being able to act as a trigger for the AIM2-inflammasome, culminating in the synthesis of IL-1β.63
The current understanding favors a view of psoriasis as a systemic inflammatory disease, with participation in the innate and adaptive immune response, sharing common characteristics with autoinflammatory diseases.58
3. BEHCET'S DISEASE
Behcet's disease (BD) (Figure 3) is a systemic inflammatory pathology of still undefined etiology. Components of the innate and adaptive immunity seem to be a part of its physiopathogenesis. Some studies have evaluated the relation between BD and inflammasones. In one study conducted with 371 patients with BD, an association was found between the disease and the IFI16 single nucleotide polymorphism, a mediator of the AIM2-inflammasome pathway.64 Four mutations of the NLRP3/cryopyrin gene were identified in a study conducted with 50 cases of BD.65 However, another study was unable to demonstrate a greater activation of caspase-1 through the inflammasome pathway in dendritic cells and neutrophils of sick patients, as compared to the controls.66
Figure 3.
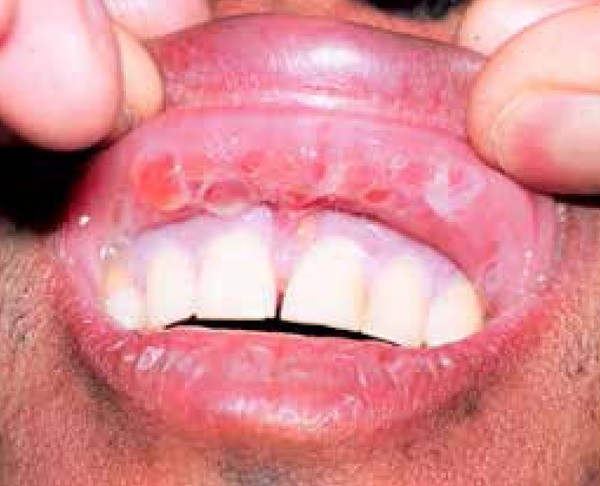
Behcet's disease
4. VITILIGO
In 2007, Jin et al. reported na association between single nucleotide polymorphisms in the NLRP1 gene of chromosome 17p13 and the risk for the development of generalized virtiligo, in addition to the association with other autoimmune and autoinflammatory diseases.67,68 In 2013, Levandowski et al. demonstrated that specific NLRP1 haplotypes presented an increased intrinsic activity of the NLRP1 inflammasome, leading to an increased production of IL-1β in an analogous mechanism, which occurred in autoinflammatory diseases stemming from the mutation of NLRP3 inflammasome. These authors suggest that these haplotypes of NLRP1 would be insuficiente to unleash the disease, but they could act as adjuvants. The increased production of IL-1β can contribute to the pathogenesis of autoimmunity to facilitate the presentation of autoantigens.69
In another work, Wang et al. illustrated Langerhans cells with NLRP1 inflammasome activated in vitiligo lesions.70 In 2014, Marie et al. showed statistically significant association between the disease and the positivity of NLRP1 and IL-1β in the perilesional area of vitiligo lesions, demonstrating that this is a better predictor than the mere presence of lymphocytic infiltrate.71
5. LUPUS ERYTHEMATOSUS
The participation of the inflammasome on the physiopathogenesis of systemic lupus erythematosus (SLE) has been a target of study in recent years. Two NLRP1 single nucleotide polymorphisms were associated with SLE, mainly with the development of nephritis, cutaneous lesions, and arthritis. NLRP3, AIM2, NLRC4, and caspase-1 polymorphisms showed not direct correlation.14 In another study, evaluating SLEs that begin in childhood and adolescence, a relationship was found with the IL-1β polymorphism but not with NLRP1.72 The relationship between IL-1β polymorphisms and the development of SLE has produced conflicting results in the literature.73 By contrast, IL-18 polymorphisms have proven to be associated with the risk of the development of the disease.74
The most reliable piece of evidence of the relationship between SLE and inflammasomes was found in lupus nephritis. As regards skin involvement, it is known that IL-18 seems to play an important role, as it presents an increased expression in the keratinocytes of patients with cutaneous lupus.75,76 Whether or not inflammasomes exert some sort of influence upon SLE's photosensitivity, however, is still under investigation, given that UVB radiation is capable of stimulating the synthesis of IL-1β in keratinocytes by activating the inflammasome.77
6. SYSTEMIC SCLEROSIS
Evidence has suggested a possible role played by inflammasomes in the physiopathology of systemic sclerosis, especially in the development of cutaneous and pulmonary fibrosis.78 Analyzing skin biopsies of 42 patients with systemic sclerosis, a greater expression of the NLRP3 inflammasome, when compared to the skin of normal individuals, was identified, including a correlation between the expression and the cutaneous thickness. However, no difference was found among the patients with limited cutaneous and diffuse cutaneous forms.79 One better expression of the NLRP3 and AIM2 genes was observed in another study in which the caspase-1 activity was also correlated with the synthesis of IL-1β and IL-18, coupled with the development of fibrosis.80
7. ACNE
Acne is a chronic inflammatory disease of the pilosebaceous unit, with four well-defined pathogenic factors: abnormal keratinization, excessive sebaceous production, colonization by Propionibacterium acnes, and inflammation. However, the sequence in which such events occur is still controversial. Some have postulated the possibility of an inflammation being the initial factor, by means of Toll-like 2 receptors and P. acnes inflammasomes, thus changing the hypothesis in which the intial point of the disease would be the abnormal keratinization with a microcomedone formation. All cases of acne would, therefore, be truly inflammatory.81 Qin et al. revealed the presence of caspase-1 and NLRP3 wrapped around the pilosebaceous follicules in acne lesions.82 Kistowska et al. demonstrated, in a study with myeloid cells, in vitro and in vivo, that P. acnes triggers the activation of the NLRP3 inflammasome, with a consequent production of IL-1β, suggesting a possible target for future therapies.83 Li et al. showed that the sebaceous cells can participate in the innate immune response, with the P. acnes activating the NLRP3 inflammasome, leading to the release of IL-1β.84
8. ROSACEA
One study, conducted with 98 patients and 48 controls, in which the relationship between rosacea and the colonization caused by Demodex folliculorum, found a greater expression of the NLRP3 inflammasome, IL-1β, and caspase-1 genes in patients with rosacea. A reduction in the expression of the ASC protein and of the IL-18 was observed, a finding that the authors suggest to be a possible explanation for the loss of microbial homeostasis.85 The participation of inflammasomes was also suggested due to the synthesis of IL-1β by the keratinocytes stimulated by UVB radiation.
9. HIDRADENITIS SUPPURATIVA
The participation of inflammasones in the physiopathology of hidradenitis suppurativa (HS) (Figure 4) has already been previously suggested due to the increase in IL-1β observed in the lesions.87 Corroborating this hypothesis, one recent study, analyzing 34 skin samples from 22 patients with HS, found a greater expression of NLPR3 and caspase-1 on the sick skin, as compared to normal skin. A greater caspase-1 activity in the keratinocytes of the affected skin was also observed.88 One therapy with colchicine, an effective medication in other diseases with inflammasomes, such as familial mediterranean fever and gout, was evaluated in eight patients with HS, but it proved not to be beneficial.89
Figure 4.
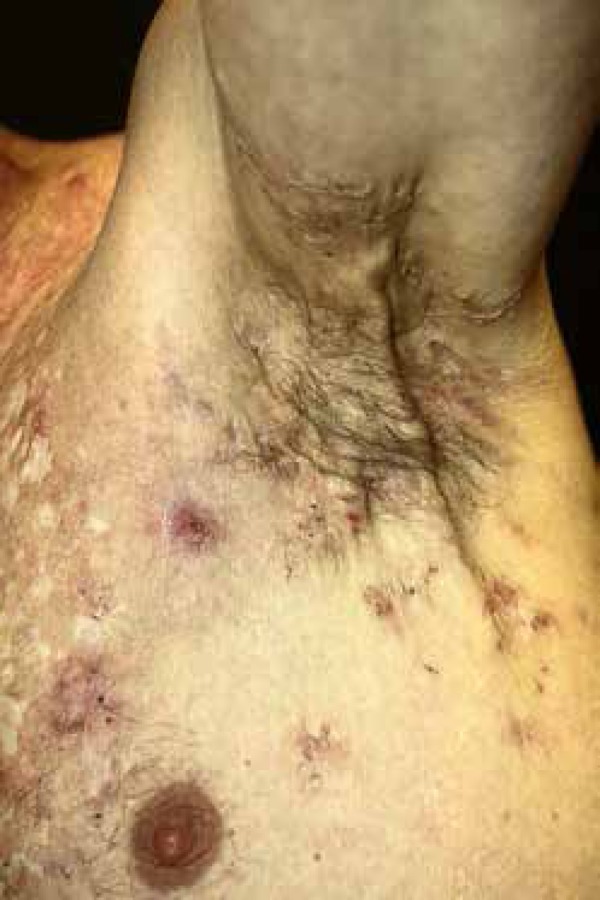
Hidradenitis suppurativa
10. CONTACT DERMATITIS
The role of inflammasones in contact dermatitis has been shown in a number of studies. Li and Zhong demonstrated that the inflammasome pathway is one of the possible paths through which nickel can be recognized by the innate immune system. Nickel is the most common allergen involved in contact sensitization; however, the exact physiopathological mechanism has yet to be fully elucidated. Nevertheless, most believe in the joint participation of the innate and adaptive immune systems. Nickel would be identified by the pattern recognition receptor NLRP3 in the epidermis, activating the inflammasome pathway, thus culminating in the production of the pro-inflammatory IL-1β cytokine.90 Watanabe et al., in an animal model, showed that the presence of inflammasomes would be essential for the development of contact hypersensitivity.91 Other studies have demonstrated the crucial role of IL-1β and IL-18 in contact dermatitis. Research with mice found that therapies with a IL1RAs can be useful in cases of contact dermatitis.
11. ATOPIC DERMATITIS
The role of inflammasomes has been debated in the etiopathogenesis of atopic dermatitis. It is believed that the activation of inflammasomes by allergens or pathogens can lead to an exacerbation of the disease.93,94 However, Niebuhr et al. reported lower NALP3 and caspase-1 expressions in atopic dermatitis lesions, when compared to normal skin and to psoriasis lesions, and suggest that this lesser function of the inflammasome may well explain, at least in part, the role of S. aureus as an aggravating factor in atopic patients.95 Evaluating NLRP1, NLRP3, and CARD8 single nucleotide polymorphisms, Bivik et al. found no association between atopic dermatitis and NLRP3 or CARD8. The association with NLRP1 was controversial, as it was significant in the case-control study, but it did not exist in the familial base study, suggesting only a modest role for NLRP1, which might be one of many risk factors for the development of atopic dermatitis.96
12. SKIN CANCER
The association between an inflammatory response and cancer continues to be controversial, and studies have shown the role of the activation of inflammasomes in tumorigenesis, both as a protective factor as will as a stimulating factor,
12.1. SQUAMOUS CELL CARCINOMA (SCC)
Drexler et al., in a study with mice, showed that the ASC protein would have a pro-inflammatory role in the tumor inflammatory infiltrate cells, in turn favoring the development of SCC, but with a contrary effect on the keratinocytes, acting as a tumor suppressor. In this same work, the loss of ASC expression in the SCC lesions in human tissues was observed.97 In one study conducted with SCC induced in mice, Gasparoto et al. demonstrated that the presence of the ASC and caspase-1 proteins play the role of protectors against both tumorigenesis and SCC progression.98 Opposing this supposed protective factor of the inflammasome, it is well-known that ultraviolet radiation, the main risk factor for the development of SCC, is a potent trigger for the release of IL-1β, through the activation of the inflammasomes within keratinocytes.99
12.2. MELANOMA
In a study with melonoma cells, Okamoto et al. found that melanomas in advanced stages exhibit characteristics of autoinflammatory diseases, with a constitutive activation of NLRP3 and the independent production of IL-1β, without the need for exogenous stimulus. In melanomas in intermediate stages, the activation occurred by triggering the IL-1 receptors (IL-1R). By contrast, in the initial stages, there was a clear need to stimulate the IL-1R and to produce a co-stimulation with muramyl dipeptide. Nevertheless, the mechanisms that would lead to this difference in the activation of the inflammasome still remain uncertain.100
Based on the understanding that the IL-1β participates in the invasion and tumor angiogenesis and that the synthesis of IL-1β is modulated by the NLRP1/NLRP3 inflammasome, Verma et al. investigated the association between the NLRP1 and NLRP3 polymorphisms and the susceptibility of the melanoma, and found a strong association between the nodular melanoma and a variant of NLRP1.101
Evaluating the human cell strains of primary and metastatic melanomas, Liu et al. showed a double role of the ASC protein. In primary melanomas, the ASC expression would be a tumorigenesis inhibitor, while in metastatic melanomas, it would be a promoting factor of tumor progression.102 Working with human cell strains, Gehrke et al. demonstrated that the metastatic melanoma cells are incapable of secreting IL-1β owing to the absence of at least one of the inflammasome components. Nonetheless, macrophages have proven to be capable of synthesizing large quantities of IL-1β, stimulated by factors stemming from necrotic cells of the melanoma. Therefore, though they are not IL-1β producers, it is still under debate whether or not cell tumors in fact benefit from the cytokines produced by the macrophages.103
Ahmad et al., in an in vitro model, demonstrated that the thymoquinone substance inhibits the cell migration of human and mice melanomas. These authors also showed the inhibitor effect of metastasis in an in vivo model in mice, through the inhibition of NLRP3, with a consequent reduction in the synthesis of IL-1β and IL18.104 Ellis et al. reported that physiological doses of epigallocatechin-3-gallate, a component of green tea, had an inhibitor effect on the proliferation of human cell strains of metastatic melanoma, by inhibiting NLRP1.105
13. INFECTIONS
The immediate response of the body against pathogens is given by the innate imune system, with the activation of the innate response regulating the prompting of the adaptive immune reponse. The understanding of the host response against infection has changed in recent years due to studies evaluating the role of inflammasomes.106
13.1. FUNGI
The fungi of the Malassezia genus are skin commensals, but are also involved in some skin diseases. Kistowska et al. showed that different species of Malassezia present the potential to activate the NLRP3 inflammasome in antigen-presenting cells, leading to a high synthesis of IL-1β.107
The NLRP3 and NLRC4 inflammasomes play key roles in the innate response against mucosal infections, as reported by Candida in two published studies.108,109 In addition, the activation of inflammasomes seems to depend on the transformation from a yeast form to a filamentous form.110,111 The relevant function of the NLRP3 inflammasome in the defense against infection caused by Microsporum canis and Trichophyton schoenleinii was also verified.112,113
In one recently published study conducted with mice, the participation of inflammasomes in the body's defense against sporotrichosis was postulated. The caspase-1 activity and the synthesis of IL-1β and IL-18 diminished during the period of greatest fungal proliferation and transitory immunosuppression prompted by the fungus.114
13.2. VIRUS
Nour et al. showed that the varicella-zoster virus is recognized by the innate immune system by means of the NLRP3, activating the inflammasome and culminating in the synthesis of IL-1β, both in the in vitro model as well as in the in vivo infected skin. Different from that observed with NLRP3 in the infected skin, no NLRP1 expression was observed. However, the study was unable to conclude if this activation is beneficial for the host or if the propagation of the virus is necessary.115
The role of the inflammasome in the response to infection by HPV-16 in keratinocytes was investigated, demonstrating the prompting of the formation of the AIM2 inflammasome, with a consequent synthesis of IL-1β. A possible pathway is being discussed in an attempt to understand the reason why the HPV infection persists in some individuals.116
Differently from the intracytoplasmatic receptors (NLRP1, NLRP3, NLRC4, and AIM2), the IFI16 is an intranuclear pattern recognition receptor that proved to be capable of recognizing the herpes virus associated with Kaposi's sarcoma (HVSK), leading to the activation of the IFI16 inflammasome, with the synthesis of IL-1β.117
Johnson et al. demonstrated that the type 1 herpes virus (HSV-1) activates the NLRP3 and IFI16 inflammasomes during the intial stages of the infection and that there is no participation of the AIM2 inflammasome. However, with the progression of the infection, the HSV-1 develops mechanisms to suppress this inflammatory response.118
13.3. BACTERIA
The relationship between the inflammasome and infection caused by Staphylococcus aureus has been studied in the literature. One mechanism to spark the neutrophilic response against cutaneous infection was proposed. In this model, in the epidermal infection (impetigo or seconday surface infection), the response would mainly be through the synthesis of IL-1α in keratinocytes. In dermal or subcutaneous infections (cellulitis, abcess), a synthesis of IL-1β would occur by means of the inflammasome pathway, both by resident cells and by recruited tissue cells.119 In one study conducted with mice, it was found that the neutrophilic response, essential to the formation of abscesses and the improvement of the infection, is mediated by the IL-1β and that the neutrophils themselves are the main cells responsible for the synthesis of this interleukin, through some given pathways, especially those of the NLRP3 inflammasome.120 Nonetheless, the response associated with the inflammasome did not prove to be beneficial. The α-hemolysin, known as a bacteria virulence fator, is capable of stimulating the NLRP3 inflammasome, causing cell death. This inflammatory response on the part of the host through tissue necrosis limits the capacity of infection control. The inflammasome appears to be a possible target for future therapies against severe infections caused by S. aureus.121
Haemophilus ducreyi is an etiological agente of a canker sore. Although it is an agent that is predominantly extracellular, some of its components can be recognized by the inflammasome. The mRNA expression of genes associated with the inflammasomes was investigated in the infected skin, showing an increase in the NLRP3, NLRC4, AIM2, caspase-1, caspase-4, caspase-5, and IL-1β expressions, illustrating that the inflammasome can participate in the innate immune response toward this pathogen. The synthesis of IL-1β in vitro in macrophages derived from monocytes, through the NLRP3 inflammasome pathway, was also observed.122
13.4. PROTOZOA
The true role of the inflammasone in American tegumentary leishmaniasis (ATL) is still uncertain and was recently presented for the first time in 25 patients from the city of Carte de Pedra, Bahia, Brazil, an endemic zone for Leishmania braziliensis. The profile of the genome from the lesional skin was compared to the normal skin of controls from a non-endemic area. This study found a high expression of the components of inflammasomes (IL-1β, AIM2, NLRP3, caspase-1, and caspase-5) in sick skin. This profile was observed in a similar manner both in the more recent, non-ulcerated lesions, as well as in the older, ulcerated lesions, which suggests that, shortly after the onset of the infection, the inflammatory response is triggered, which may well explain why even early treatment centers are unable to prevent this type of ulcer.123
14. PRESSURE ULCER
Using a biomechanical experimental model, in which young and elderly human skin were submitted to pressure and ischemia, Stojadinovic et al. observed that the elderly skin already presented a lesser expression of the components of the inflammasome and that the pressure did not result in any changes. The synthesis of IL-1β, which was already low in the elderly skin, did not change during the study. In the young skin, the expression of NLRP3 and caspase-1, but not of the ASC adaptor protein, increased significantly with pressure. By contrast, the synthesis of IL-1β diminished, suggesting that the pressure can undo the link between the components of the inflammasome, thus reducing the quantity of IL-1β produced, even with the increased presence of these components. It is also being debated whether or not the reduction in the innate immune response with age can be involved in the greater risk for the development of pressure ulcers .124
15. WOUND HEALING
The function of the inflammasome in the skin healing process is not well-defined in the literature, but some studies have treated the subject. In one in vitro model conducted with material from mice, genetic variations of NLRP1 showed the capacity to interfere in the production of cytokines that might be involved in the healing process.125
The NLRP3 inflammasome may well be involved in slow healing in diabetic patients, as seen in the findings from a work conducted with diabetic and obese mice in which the blocking of this inflammasome led to an improvement in the healing process.126 In another study with material from both humans and mice, it was postulated that the NLRP3 inflammasome may be continually ac-tivated within the macrophages present in the wounds of diabetic patients, and that this contributes to the difficulty of wound healing.127 By contrast, another study conducted with mice showed that the NLRP3 inflammasome would be important in the initial stage of the healing process.128
16. DRUG REACTIONS
Weston and Uetrecht debate the role of the inflammasome in the genesis of idiosyncratic medication reactions. The majority of these reactions occur due to medication that produces reactive metabolytes. Four medications were evaluated, two of which are associated with cutaneous reactions (teleprevir and dimethyl fumarate) and two that are not related to cutaneous lesions (boceprevir and ethacrynic acid). Only the former medications were able to activate the inflammasome in monocytes of human peripheral blood. The hypothesis raised is that the capacity of the reactive metabolyte to stimulate the inflammasome would be a determining factor in the potential of a given medication to cause idiosyncratic reactions.129 Another study proposed that the activation of the inflammasome may well be a fundamental part for the cutaneous rash that is caused by nevirapine.130 Further study is warranted to corroborate these hypotheses.
17. ANDROGENETIC ALOPECIA
Evaluating the biopsies from 18 male patients, Vaccari et al. identified an increased caspase-1 expression in the epidermis of patients with untreated androgenetic alopecia (AGA), as well as in cases of AGA in which finasteride was used, but with therapeutic failure. Patients with AGA who were using finasterie with a good therapeutic response and the normal controls presented a reduced caspase-1 expression. In addition, this same study demonstrated, in a keratinocyte culture of human skin, that finasteriade, in the presence of testosterone or of di-hydrotestosterone, leads to a reduction in the levels of caspase-1. These findings suggest that the caspase-1 activity is associated with AGA and shows a possible interaction between the steroid hormones and the innate immunity, which may represent a viable future therapy for AGA.131
CONCLUSION
Skin, the major organ of the human body, is continually exposed to a wide range of stimuli, which can be recognized by the innate immune system. The discovery of the inflammasome throughout the last decade has shed light on a necessary mechanism for the proper functioning of this system. The concept of the autoinflammatory disease, introduced a few years before the descriptions of inflammasomes, has been more well-understood and expanded since then. Periodic fever syndromes and other genetic disorders, such as those now called CAPS, were classified as monogenic autoinflammatory diseases, with a clear impact on the therapeutic response after the introduction of medications geared toward the inhibition of IL1. Diseases considered to be autoimmune, such as psoriasis, but that fail to fulfill the criteria of autoimmunity, have begun to be understood in a different manner. Diseases understood as autoimmune, such as SLE and vitiligo, have also shown that they in fact share the characteristics of innate immunity disorders. Neutrophilic dermatoses, especially Sweet's syndrome and pyoderma gangrenosum, represent a field of great expectations for new studies that can serve to associate these diseases once and for all, or not, with inflammasomopathies. Moreover, the best knowledge about the action of inflammasomes can open the door to new therapies for severe diseases, such as melanomas, or to those that heavily impact one's quality of life, such as acne and atopic dermatitis.
Questions.
-
About inflammasomes, it is incorrect to state that:
they are key pieces in adaptive immunity and are responsible for responses to autoantigens in autoimmune disorders.
described for the first time in 2002, they are constitutive of the innate immune response to exogenous and endogenous stimuli.
they are intracellular protein compounds, formed by a pattern recognition receptor, an adaptor protein, and a caspase-1 enzyme.
activation of inflammasome complex leads to the synthesis of cytokines (IL-1β and IL18).
-
Regarding the concept of autoinflammatory disease, it is correct to state that:
it is an old and obsolete concept, which included diseases currently better classified as autoimmune.
the presence of autoantigens and autoantibodies are essential to classify a pathology as autoinflammatory.
autoinflammation and autoimmunity may coexist in a single disease, as a result of innate and adaptive immune system disorders, respectively.
the activation of B and T cells is essential for the physiopathology of autoinflammatory diseases.
-
About cryopyrin-associated periodic syndromes (CAPS), it is incorrect to state that:
include a spectrum that encompasses familial cold autoinflammatory syndrome, Muckle-Wells syndrome, and neonatal onset multisystem inflammatory disease.
result from mutations in NLRP3 inflammasome, leading to the excessive IL-1β synthesis.
in familial cold autoinflammatory syndrome, previously called familial cold urticaria, episodes may be triggered by exposure to low ambient temperatures.
treatment with IL1 receptor antagonist (Anakinra) provides discouraging results, and does not interfere in the disease's natural history.
-
What is the wrong association between disease and its skin symptoms?
PAPA syndrome: ulcers, cystic acne, and hidradenitis.
familial Mediterranean fever: erysipeloid erythema, pruriginous, lesions, and palmoplantar erythema.
Schnitzler syndrome: macules, papules, and urticarial plaques that appear after exposure to cold.
Hyper IgD syndrome (HIDS): oral and genital aphthae, painful papules and macules, and panniculitis.
-
About pyoderma gangrenous and Sweet's syndrome, we may state that:
the hypothesis of these being autoinflammatory diseases is rejected due to clinical and histological differences in relation to hereditary autoinflammatory diseases.
the undefined pathogeny and the similarity to hereditary autoinflammatory diseases make way for the hypothesis that they are associated with inflammasome disorders.
experts on the subject have declared that they are against the association between pyoderma gangrenoum and autoinflammation.
they have definitely been associated with inflammasome gene mutations.
-
Which of the sequences below form an inflammasome complex?
NLRP1- ASC- aromatase
NLRP3- ASC- caspase-1
AIM2- NLRP1- ASC
IPAF- ASC- NLRP1
-
What is the correct inflammasome activation sequence?
NLRP3- caspase - ASC
caspase- ASC - AIM2
ASC- caspase - NLRP1
NLRP1 - ASC- caspase-1
-
Which of the diseases below represents a monogenic autoinflammatory fever syndrome?
Muckle-Wells syndrome
Psoriasis
Behcet's disease
Lambert-Eaton syndrome (LES)
-
The main complication and cause of mortality in cryopyrin- associated periodic syndromes (CAPS) is:
primary amyloidosis
hepatic failure
renal failure caused by secondary amyloidosis
cardiac failure
-
About vitiligo, it is possible to state that:
it is a monogenic autoinflammatory disease.
it is a typical autoimmune disease, with no evidence of innate immunity participation.
evidence suggests the participation of innate immunity by means of the inflammasome complex in its physiopathogeny.
Anakinra is currently the treatment of choice.
-
The following are factors suggesting the participation of inflammasomes in the physiopathology of hidradenitis, except:
increased IL-1β in the lesions.
favorable response to colchicine treatment.
greater expression of inflammasome components in the lesions.
a condition similar to hidradenitis in PAPA syndrome.
-
Regarding eczema and inflammasome, it is incorrect to state that
nickel may induce sensitization by means of NLRP3 inflammasome complex activation.
innate and adaptive immunity may be part of the contact dermatitis sensitization mechanism.
a contact allergen may stimulate IL-1β synthesis through the inflammasome pathway.
their relation to atopic dermatitis is well documented.
-
In the skin oncology field, inflammasomes:
may participate in tumor genesis by means of ultraviolet radiation stimulus, leading to IL-1β synthesis.
play a well-defined role as protector in squamous cell carcinoma.
inflammasome activation is associated with a better prognosis in melanoma.
it has been defined that inflammation acts as a protective factor in oncogenesis.
-
About the Anakinra drug, it is correct to state that:
it is an IL-1 receptor antagonist.
it is contraindicated in inflammasomepathies such as CAPS.
it acts by reducing IL-1 synthesis.
it acts by increasing IL-1 effect.
-
Pyroptosis is:
the inflammasome intracellular receptor activation mechanism.
apoptosis induced by inflammasome activation.
apoptosis induced by adaptive immunity,
apoptosis induced by inflammasome inhibition.
-
Differential diagnosis of infantile urticarial fever syndromes include the following, except:
familial cold autoinflammatory syndrome (familial cold urticaria)
Schnitzler syndrome
Muckle-Wells syndrome
deficiency of the interleukin-1–receptor antagonist (DIRA)
-
The following may be results of inflammasome pathway activation, except:
IL 18 synthesis
IL-1β synthesis
pyroptosis
caspase-1 inhibition
-
Inflammasome activation represents:
a mechanism that may be essential in fighting infections;
a pathological mechanism, inactive in healthy individuals.
a defense mechanism, inactive in hereditary diseases, such as familial Mediterranean fever.
a mechanism restricted to cutaneous-mucous diseases.
-
Which of the words below is not associated with autoinflammatory diseases:
autoantibodies
innate immunity
inflammasome
neutrophils
-
About psoriasis, it is correct to state that:
it is caused by NLRP3 gene mutation and is considered a monogenic inflammasomepathy.
it may be classified as a systemic inflammatory disease, with the participation of innate and adaptive immune response, and shares characteristics common to autoinflammatory diseases.
it is a classical autoimmune disease.
neutrophilic inflammatory infiltrates and the participation of adaptive immunity are similar characteristics to those in autoinflammatory diseases.
|
Answer key Geographic tongue and psoriasis: clinical, histopathological, immunohistochemical and genetic correlation - literature review 2016;91(4):410-21. | |||
| 1 - C | 6 - A | 11 - B | 16 - B |
| 2 - D | 7 - B | 12 - D | 17 - C |
| 3 - D | 8 - B | 13 - C | 18 - D |
| 4 - B | 9 - B | 14 - B | 19 - C |
| 5 - C | 10 - A | 15 - D | 20 - D |
| Papers Information for all members: The EMC-D questionnaire is now available at the homepage of the Brazilian Annals of Dermatology: www.anaisdedermatologia.org.br. The deadline for completing the questionnaire is 30 days from the date of online publication. |
Footnotes
Conflict of interest: none
Financial support: none
Study conducted at the Department of Dermatology of the Faculty of Medicine of the Universidade de São Paulo - São Paulo (SP), Brazil.
REFERENCES
- 1.Martinon F, Burns K, Tschopp J. The inflammasome: a molecular platform triggering activation of inflammatory caspases and processing of proIL-beta. Mol. Cell. 2002;10:417–426. doi: 10.1016/s1097-2765(02)00599-3. [DOI] [PubMed] [Google Scholar]
- 2.Dunn JH, Ellis LZ, Fujita M. Inflammasomes as molecular mediators of inflammation and cancer: Potential role in melanoma. Cancer Lett. 2012;314:24–33. doi: 10.1016/j.canlet.2011.10.001. [DOI] [PubMed] [Google Scholar]
- 3.Gross O, Thomas CJ, Guarda G, Tschopp J. The inflammasome: an integrated view. Immunol Rev. 2011;243:136–151. doi: 10.1111/j.1600-065X.2011.01046.x. [DOI] [PubMed] [Google Scholar]
- 4.de Zoete MR, Palm NW, Zhu S, Flavell RA. Inflammasomes. Cold Spring Harb Perspect Biol. 2014;6:a016287–a016287. doi: 10.1101/cshperspect.a016287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Guo H, Callaway JB, Ting JP. Inflammasomes: mechanism of action, role in disease, and therapeutics. Nat Med. 2015;21:677–687. doi: 10.1038/nm.3893. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Miao EA, Rajan JV, Aderem A. 2011. Caspase-1-induced pyroptotic cell death. Immunol Ver. 2011;243:206–214. doi: 10.1111/j.1600-065X.2011.01044.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Schroder K, Tschopp J. The inflammasomes. Cell. 2010;140:821–832. doi: 10.1016/j.cell.2010.01.040. [DOI] [PubMed] [Google Scholar]
- 8.Menu P, Vince JE. The NLRP3 inflammasome in health and disease: the good, the bad and the ugly. Clin Exp Immunol. 2011;166:1–15. doi: 10.1111/j.1365-2249.2011.04440.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Goldbach-Mansky R, Kastner DL. Autoinflammation: the prominent role of IL-1 in monogenic autoinflammatory diseases and implications for common illnesses. J Allergy Clin Immunol. 2009;124:1141–1149. doi: 10.1016/j.jaci.2009.11.016. quiz 1150-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Strowig T, Henao-Mejia J, Elinav E, Flavell R. Inflammasomes in health and disease. Nature. 2012;481:278–286. doi: 10.1038/nature10759. [DOI] [PubMed] [Google Scholar]
- 11.Pontillo A, Catamo E, Arosio B, Mari D, Crovella S. NALP1/NLRP1 GeneticVariants are Associated With Alzheimer Disease. Alzheimer Dis Assoc Disord. 2012;26:277–281. doi: 10.1097/WAD.0b013e318231a8ac. [DOI] [PubMed] [Google Scholar]
- 12.Freeman LC, Ting JP. The pathogenic role of the inflammasome in neurodegenerative diseases. J Neurochem. 2016;136(Suppl 1):29–38. doi: 10.1111/jnc.13217. [DOI] [PubMed] [Google Scholar]
- 13.Magitta NF, Bøe Wolff AS, Johansson S, Skinningsrud B, Lie BA, Myhr KM, et al. A codingpolymorphism in NALP1 confers risk for autoimmune Addison's disease and type 1 diabetes. Genes Immun. 2009;10:120–124. doi: 10.1038/gene.2008.85. [DOI] [PubMed] [Google Scholar]
- 14.Pontillo A, Girardelli M, Kamada AJ, Pancotto JA, Donadi EA, Crovella S, et al. Polimorphisms in inflammasome genes are involved in the predisposition to systemic lupus erythematosus. Autoimmunity. 2012;45:271–278. doi: 10.3109/08916934.2011.637532. [DOI] [PubMed] [Google Scholar]
- 15.Pontillo A, Oshiro TM, Girardelli M, Kamada AJ, Crovella S, Duarte AJ. Polymorphisms in inflammasome' genes and susceptibility to HIV-1 infection. J Acquir Immune Defic Syndr. 2012;59:121–125. doi: 10.1097/QAI.0b013e3182392ebe. [DOI] [PubMed] [Google Scholar]
- 16.Dorhoi A, Nouailles G, Jörg S, Hagens K, Heinemann E, Pradl L, et al. Activation of the NLRP3 inflammasome by Mycobacterium tuberculosis is uncoupled from susceptibility to active tuberculosis. Eur J Immunol. 42:374–384. doi: 10.1002/eji.201141548. [DOI] [PubMed] [Google Scholar]
- 17.Wu J, Fernandes-Alnemri T, Alnemri ES. Involvement of the AIM2, NLRC4, and NLRP3 inflammasomes in caspase-1 activation by Listeria monocytogenes. J Clin Immunol. 2010;30:693–702. doi: 10.1007/s10875-010-9425-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Gavrilin MA, Wewers MD. Francisella Recognition by Inflammasomes: Differences between Mice and Men. Front Microbiol. 2011;2:11–11. doi: 10.3389/fmicb.2011.00011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Case CL, Shin S, Roy CR. Asc and Ipaf Inflammasomes direct distinct pathways for caspase-1 activation in response to Legionella pneumophila. Infect Immun. 2009;77:1981–1991. doi: 10.1128/IAI.01382-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Witzenrath M, Pache F, Lorenz D, Koppe U, Gutbier B, Tabeling C, et al. The NLRP3 inflammasome is differentially activated by pneumolysin variants and contributes to host defense in pneumococcal pneumonia. J Immunol. 2011;187:434–440. doi: 10.4049/jimmunol.1003143. [DOI] [PubMed] [Google Scholar]
- 21.Kolb R, Liu GH, Janowski AM, Sutterwala FS, Zhang W. Inflammasomes in cancer: a double-edged sword. Protein Cell. 2014;5:12–20. doi: 10.1007/s13238-013-0001-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Liu R, Truax AD, Chen L, Hu P, Li Z, Chen J, et al. Expression profile of innate immune receptors, NLRs and AIM2, in human colorectal cancer: correlation with cancer stages and inflammasome components. Oncotarget. 2015;6:33456–33469. doi: 10.18632/oncotarget.5587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.McGonagle D, McDermott MF. A Proposed Classification of the Immunological Diseases. PLoS Med. 2006;3: doi: 10.1371/journal.pmed.0030297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Yang CA, Chiang BL. Inflammasomes and human autoimmunity: A comprehensive review. J Autoimmun. 2015;61:1–8. doi: 10.1016/j.jaut.2015.05.001. [DOI] [PubMed] [Google Scholar]
- 25.McDermott MF, Aksentijevich I, Galon J, McDermott EM, Ogunkolade BW, Centola M, et al. Germline Mutations in the Extracellular Domains of the 55 kDa TNF Receptor, TNFR1, Define a Family of Dominantly Inherited Autoinflammatory Syndromes. Cell. 1999;97:133–144. doi: 10.1016/s0092-8674(00)80721-7. [DOI] [PubMed] [Google Scholar]
- 26.Dávila-Seijo P, Hernández-Martín A, Torrelo A. Autoinflammatory syndromes for the dermatologist. Clin Dermatol. 2014;32:488–501. doi: 10.1016/j.clindermatol.2014.02.004. [DOI] [PubMed] [Google Scholar]
- 27.Moghaddas F, Masters SL. Monogenic autoinflammatory diseases: Cytokinopathies. Cytokine. 2015;74:237–246. doi: 10.1016/j.cyto.2015.02.012. [DOI] [PubMed] [Google Scholar]
- 28.Nguyen TV, Cowen EW, Leslie KS. Autoinflammation: From monogenic syndromes to common skin diseases. J Am Acad Dermatol. 2013;68:834–853. doi: 10.1016/j.jaad.2012.11.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Braun-Falco M, Ruzicka T. Skin manifestations in autoinflammatory syndromes. J Dtsch Dermatol Ges. 2011;9:232–246. doi: 10.1111/j.1610-0387.2010.07580.x. [DOI] [PubMed] [Google Scholar]
- 30.Sanchez GA, de Jesus AA, Goldbach-Mansky R. Monogenic autoinflammatory diseases: disorders of amplified danger sensing and cytokine dysregulation. Rheum Dis Clin North Am. 2013;39:701–734. doi: 10.1016/j.rdc.2013.08.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Contassot E, Beer HD, French LE. Interleukin-1, inflammasomes, autoinflammation and the skin. Swiss Med Wkly. 2012;142:w13590–w13590. doi: 10.4414/smw.2012.13590. [DOI] [PubMed] [Google Scholar]
- 32.Miyamae T. Cryopyrin-associated periodic syndromes: diagnosis and management. Paediatr Drugs. 2012;14:109–117. doi: 10.2165/11595040-000000000-00000. [DOI] [PubMed] [Google Scholar]
- 33.Nakamura Y, Kambe N. Linkage of bacterial colonization of skin and the urticaria-like rash of NLRP3-mediated autoinflammatory syndromes through mast cell-derived TNF-a. J Dermatol Sci. 2013;71:83–88. doi: 10.1016/j.jdermsci.2013.04.009. [DOI] [PubMed] [Google Scholar]
- 34.Aksentijevich I, Putnam CD, Remmers EF, Mueller JL, Le J, Kolodner RD, et al. The Clinical Continuum of Cryopyrinopathies: Novel CIAS1 Mutations in North American Patients and a New Cryopyrin Model. Arthritis Rheum. 2007;56:1273–1285. doi: 10.1002/art.22491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Yu JR, Leslie KS. Cryopyrin-associated periodic syndrome: an update on diagnosis and treatment response. Curr Allergy Asthma Rep. 2011;11:12–20. doi: 10.1007/s11882-010-0160-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Jesus AA, Goldbach-Mansky R. IL-1 blockade in autoinflammatory syndromes. Annu Rev Med. 2014;65:223–244. doi: 10.1146/annurev-med-061512-150641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Abramovits W, Oquendo M. Introduction to autoinflammatory syndromes and diseases. Dermatol Clin. 2013;31:363–385. doi: 10.1016/j.det.2013.04.010. [DOI] [PubMed] [Google Scholar]
- 38.ter Haar NM, Oswald M, Jeyaratnam J, Anton J, Barron KS, Brogan PA, et al. Recommendations for the management of autoinflammatory diseases. Ann Rheum Dis. 2015;74:1636–1644. doi: 10.1136/annrheumdis-2015-207546. [DOI] [PubMed] [Google Scholar]
- 39.Sohar E., Gafni J., Pras M., Heller H. Familial Mediterranean fever. A survey of 470 cases and review of the literature. Am J Med. 1967;43:227–253. doi: 10.1016/0002-9343(67)90167-2. [DOI] [PubMed] [Google Scholar]
- 40.Caso F, Rigante D, Vitale A, Lucherini OM, Costa L, Atteno M, et al. Monogenic autoinflammatory syndromes: state of the art on genetic, clinical, and therapeutic issues. Int J Rheumatol. 2013;2013:513782–513782. doi: 10.1155/2013/513782. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Kuijk LM, Beekman JM, Koster J, Waterham HR, Frenkel J, Coffer PJ. HMG-CoA reductase inhibition induces IL-1beta release through Rac1/PI3K/PKB-dependent caspase-1 activation. Blood. 2008;112:3563–3573. doi: 10.1182/blood-2008-03-144667. [DOI] [PubMed] [Google Scholar]
- 42.Drenth JP, Haagsma CJ, van der Meer JW. Hyperimmunoglobulinemia D and periodic feversyndrome: The clinical spectrum in a series of 50 patients. Medicine (Baltimore) 1994;73:133–144. [PubMed] [Google Scholar]
- 43.Yoram C., Padeh A.R. Hyperimmunoglobulin-D syndrome: pathophysiology. © 2013. 2012. [UpToDate] [Google Scholar]
- 44.Smith EJ, Allantaz F, Bennett L, Zhang D, Gao X, Wood G, et al. Clinical, Molecular, and Genetic Characteristics of PAPA Syndrome: A Review. Curr Genomics. 2010;11:519–527. doi: 10.2174/138920210793175921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Hong JB, Su YN, Chiu HC. Pyogenic arthritis, pyoderma gangrenosum, and acne syndrome (PAPA syndrome): report of a sporadic case without an identifiable mutation in the CD2 BP1 gene. J Am Acad Dermatol. 2009;61:533–535. doi: 10.1016/j.jaad.2008.11.017. [DOI] [PubMed] [Google Scholar]
- 46.Look J, Lamprecht P, Timmann C, Mrowietz U, Csernok E, Gross WL. Genetic predisposition (NLRP3 V198m mutation) for IL-1-mediated inflammation in a patient with Schnitzler syndrome. J Allergy Clin Immunol. 2010;125:500–502. doi: 10.1016/j.jaci.2009.10.066. [DOI] [PubMed] [Google Scholar]
- 47.Pizzirani C, Falzoni S, Govoni M, La Corte R, Donadei S, Di Virgilio F, et al. Dysfunctional inflammasome in Schnitzler's syndrome. Rheumatology (Oxford) 2009;48:1304–1308. doi: 10.1093/rheumatology/kep222. [DOI] [PubMed] [Google Scholar]
- 48.Lipsker D. The Schnitzler syndrome. Orphanet J Rare Dis. 2010;5:38–38. doi: 10.1186/1750-1172-5-38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Aksentijevich I, Masters SL, Ferguson PJ, Dancey P, Frenkel J, van Royen-Kerkhoff A, et al. An autoinflammatory disease with deficiency of the interleukin-1-Receptor antagonist. N Engl J Med. 2009;360:2426–2437. doi: 10.1056/NEJMoa0807865. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Jesus AA, Osman M, Silva CA, Kim PW, Pham TH, Gadina M, et al. A Novel Mutation of IL1RN in the deficiency of interleukin-1 receptor antagonist syndrome: description of two unrelated cases from Brazil. Arthritis Rheum. 2011;63:4007–4017. doi: 10.1002/art.30588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Naik HB, Cowen EW. Autoinflammatory pustular neutrophilic diseases. Dermatol Clin. 2013;31:405–425. doi: 10.1016/j.det.2013.04.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Wallach D, Vignon-Pennamen MD. Pyoderma gangrenosum and Sweet syndrome: the prototypic neutrophilic dermatoses. Br J Dermatol. 2015 Jul 22; doi: 10.1111/bjd.13955. [DOI] [PubMed] [Google Scholar]
- 53.Leslie KS, Lachmann HJ, Bruning E, McGrath JA, Bybee A, Gallimore JR, et al. Phenotype, genotype, and sustained response to anakinra in 22 patients with autoinflammatory disease associated with CIAS-1/NALP3 mutations. Arch Dermatol. 2006;142:1591–1597. doi: 10.1001/archderm.142.12.1591. [DOI] [PubMed] [Google Scholar]
- 54.Kluger N, Gil-Bistes D, Guillot B, Bessis D. Efficacy of Anti-Interleukin-1 Receptor Antagonist Anakinra (Kineret®) in a Case of Refractory Sweet's Syndrome. Dermatology. 2011;222:123–127. doi: 10.1159/000326112. [DOI] [PubMed] [Google Scholar]
- 55.Delluc A, Limal N, Puéchal X, Francès C, Piette JC. Cacoub P: Efficacy of anakinra, an IL1 receptor antagonist, in refractory Sweet syndrome. Ann Rheum Dis. 2008;67:278–279. doi: 10.1136/ard.2006.068254. [DOI] [PubMed] [Google Scholar]
- 56.Nesterovitch AB, Hoffman MD, Simon M, Petukhov PA, Tharp MD, Glant TT. Mutations in the PSTPIP1 gene and aberrant splicing variants in patients with pyoderma gangrenosum. Clin Exp Dermatol. 2011;36:889–895. doi: 10.1111/j.1365-2230.2011.04137.x. [DOI] [PubMed] [Google Scholar]
- 57.Nestle FO, Kaplan DH, Barker J. Psoriasis. N Engl J Med. 2009;361:496–509. doi: 10.1056/NEJMra0804595. [DOI] [PubMed] [Google Scholar]
- 58.Rivas Bejarano JJ, Valdecantos WC. Psoriasis as autoinflammatory disease. Dermatol Clin. 2013;31:445–460. doi: 10.1016/j.det.2013.04.009. [DOI] [PubMed] [Google Scholar]
- 59.Carlström M, Ekman AK, Petersson S, Söderkvist P, Enerbäck C. Genetic support for the role of the NLRP3 inflammasome in psoriasis susceptibility. Exp Dermatol. 2012;21:932–937. doi: 10.1111/exd.12049. [DOI] [PubMed] [Google Scholar]
- 60.Ekman AK, Verma D, Fredrikson M, Bivik C, Enerbäck C. Genetic variations of NLRP1: susceptibility in psoriasis. Br J Dermatol. 2014;171:1517–1520. doi: 10.1111/bjd.13178. [DOI] [PubMed] [Google Scholar]
- 61.Johansen C, Moeller K, Kragballe K, Iversen L. The activity of caspase-1 is increased in lesional psoriatic epidermis. J Invest Dermatol. 2007;127:2857–2864. doi: 10.1038/sj.jid.5700922. [DOI] [PubMed] [Google Scholar]
- 62.Salskov-Iversen ML, Johansen C, Kragballe K, Iversen L. Caspase-5 Expression Is Upregulated in Lesional Psoriatic Skin. J Invest Dermatol. 2011;131:670–676. doi: 10.1038/jid.2010.370. [DOI] [PubMed] [Google Scholar]
- 63.Dombrowski Y, Peric M, Koglin S, Kammerbauer C, Göss C, Anz D. Cytosolic DNA triggers inflammasome activation in keratinocytes in psoriatic lesions. Sci Transl Med. 2011;3:82ra38–82ra38. doi: 10.1126/scitranslmed.3002001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Ortiz-Fernández L, García-Lozano JR, Montes-Cano MA, Conde-Jaldón M, Ortego-Centeno N, García-Hernández FJ. Variants of the IFI16 gene affecting the levels of expression of mRNA are associated with susceptibility to Behçet Disease. J Rheumatol. 2015;42:695–701. doi: 10.3899/jrheum.140949. [DOI] [PubMed] [Google Scholar]
- 65.Yüksel S, Eren E, Hatemi G, Sahillioglu AC, Gültekin Y, Demiröz D, et al. Novel NLRP3/cryopyrin mutations and pro-inflammatory cytokine profiles in Behçet's syndrome patients. Int Immunol. 2014;26:71–81. doi: 10.1093/intimm/dxt046. [DOI] [PubMed] [Google Scholar]
- 66.Türe-Özdemir F, Tulunay A, Elbasi MO, Tatli I, Maurer AM, Mumcu G, et al. Pro-inflammatory cytokine and caspase-1 responses to pattern recognition receptor activation of neutrophils and dendritic cells in Behcet's disease. Rheumatology (Oxford) 2013;52:800–805. doi: 10.1093/rheumatology/kes399. [DOI] [PubMed] [Google Scholar]
- 67.Jin Y, Mailloux CM, Gowan K, Riccardi SL, LaBerge G, Bennett DC, et al. NALP1 in vitiligo-associated multiple autoimmune disease. N Engl J Med. 2007;356:1216–1225. doi: 10.1056/NEJMoa061592. [DOI] [PubMed] [Google Scholar]
- 68.Jin Y, Birlea SA, Fain PR, Spritz RA. Genetic variations in NALP1 are associated with generalized vitiligo in a Romanian population. J Invest Dermatol. 2007;127:2558–2562. doi: 10.1038/sj.jid.5700953. [DOI] [PubMed] [Google Scholar]
- 69.Levandowski CB, Mailloux CM, Ferrara TM, Gowan K, Ben S, Jin Y, et al. NLRP1 haplotypes associated with vitiligo and autoimmunity increase interleukin-1ß processing via the NLRP1 inflammasome. Proc Natl Acad Sci U.S.A. 2013;110:2952–2956. doi: 10.1073/pnas.1222808110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Wang CQF, Cruz-Inigo AE, Fuentes-Duculan J, Moussai D, Gulati N, Sullivan-Whalen M, et al. Th17 Cells and Activated Dendritic Cells Are Increased in Vitiligo Lesions. PLoS ONE. 2011;6: doi: 10.1371/journal.pone.0018907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Marie J, Kovacs D, Pain C, Jouary T, Cota C, Vergier B, et al. Inflammasome activation and vitiligo/nonsegmental vitiligo progression. Br J Dermatol. 2014;170:816–823. doi: 10.1111/bjd.12691. [DOI] [PubMed] [Google Scholar]
- 72.Pontillo A, Reis EC, Liphaus BL, Silva CA, Carneiro-Sampaio M. Inflammasome polymorphisms in juvenile systemic lupus erythematosus. Autoimmunity. 2015;48:434–437. doi: 10.3109/08916934.2015.1064399. [DOI] [PubMed] [Google Scholar]
- 73.Wang B, Zhu JM, Fan YG, Feng CC, Chen GM, Chen H, et al. The association of IL1a and IL1ß polymorphisms with susceptibility to systemic lupus erythematosus: a meta-analysis. Gene. 2013;527:95–101. doi: 10.1016/j.gene.2013.05.059. [DOI] [PubMed] [Google Scholar]
- 74.Wen D, Liu J, Du X, Dong JZ, Ma CS. Association of interleukin-18 (-137G/C) polymorphism with rheumatoid arthritis and systemic lupus erythematosus: a meta-analysis. Int Rev Immunol. 2014;33:34–44. doi: 10.3109/08830185.2013.816699. [DOI] [PubMed] [Google Scholar]
- 75.Wittmann M, Macdonald A, Renne J. IL-18 and skin inflammation. Autoimmun Rev. 2009;9:45–48. doi: 10.1016/j.autrev.2009.03.003. [DOI] [PubMed] [Google Scholar]
- 76.Wang D, Drenker M, Eiz-Vesper B, Werfel T, Wittmann M. Evidence for a pathogenetic role of interleukin-18 in cutaneous lupus erythematosus. Arthritis Rheum. 2008;58:3205–3215. doi: 10.1002/art.23868. [DOI] [PubMed] [Google Scholar]
- 77.Kahlenberg JM, Kaplan MJ. The Inflammasome and lupus- another innate immune mechanism contributing to disease pathogenesis? Curr Opin Rheumatol. 2014;26:475–481. doi: 10.1097/BOR.0000000000000088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.van Bon L, Cossu M, Radstake TRDJ. An update on an immune system that goes awry in systemic sclerosis. Curr Opin in Rheumatol. 2011;23:505–510. doi: 10.1097/BOR.0b013e32834b0dac. [DOI] [PubMed] [Google Scholar]
- 79.Martínez-Godínez MA, Cruz-Domínguez MP, Jara LJ, Domínguez-López A, Jarillo-Luna RA, Vera-Lastra O, et al. Expression of NLRP3 Inflammasome, Cytokines and Vascular Mediators in the Skin of Systemic Sclerosis Patients. Isr Med Assoc J. 2015;17:5–10. [PubMed] [Google Scholar]
- 80.Artlett CM, Sassi-Gaha S, Rieger JL, Boesteanu AC, Feghali-Bostwick CA, Katsikis PD. The inflammasome activating caspase-1 mediates fibrosis and myofibroblast differentiation in systemic sclerosis. Arthritis Rheum. 2011;63:3563–3574. doi: 10.1002/art.30568. [DOI] [PubMed] [Google Scholar]
- 81.Rosen J, Friedman AJ. Inflammatory acne: new developments in pathogenesis and treatment. Cutis. 2014;94:266–267. [PubMed] [Google Scholar]
- 82.Qin M, Pirouz A, Kim MH, Krutzik SR, Garbán HJ, Kim J. Propionibacterium acnes induces IL-1ß secretion via the NLRP3 inflammasome in human monocytes. J Invest Dermatol. 2014;134:381–388. doi: 10.1038/jid.2013.309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Kistowska M, Gehrke S, Jankovic D, Kerl K, Fettelschoss A, Feldmeyer L, et al. IL-1ß Drives Inflammatory Responses to Propionibacterium acnes In Vitro and In Vivo. J Invest Dermatol. 2014;134:677–685. doi: 10.1038/jid.2013.438. [DOI] [PubMed] [Google Scholar]
- 84.Li ZJ, Choi DK, Sohn KC, Seo MS, Lee HE, Lee Y, et al. Propionibacterium acnes activates the NLRP3 inflammasome in human sebocytes. J Invest Dermatol. 2014;134:2747–2756. doi: 10.1038/jid.2014.221. [DOI] [PubMed] [Google Scholar]
- 85.Casas C, Paul C, Lahfa M, Livideanu B, Lejeune O, Alvarez-Georges S, et al. Quantification of Demodex folliculorum by PCR in rosacea and its relationship to skin innate immune activation. Exp Dermatol. 2012;21:906–910. doi: 10.1111/exd.12030. [DOI] [PubMed] [Google Scholar]
- 86.Salzer S, Kresse S, Hirai Y, Koglin S, Reinholz M, Ruzicka T, et al. Cathelicidin peptide LL-37 increases UVB-triggered inflammasome activation: Possible implications for rosacea. J Dermatol Sci. 2014;76:173–179. doi: 10.1016/j.jdermsci.2014.09.002. [DOI] [PubMed] [Google Scholar]
- 87.van der Zee HH, van den Broecke DG, Dik WA, Laman JD, Prens EP. Elevated levels of tumour necrosis factor (TNF)-a, interleukin (IL)-1ß and IL-10 in hidradenitis suppurativa skin: a rationale for targeting TNF-a and IL-1ß. Br. J Dermatol. 2011;164:1292–1298. doi: 10.1111/j.1365-2133.2011.10254.x. [DOI] [PubMed] [Google Scholar]
- 88.Lima AL, Karl I, Giner T, Poppe H, Schmidt M, Presser D, et al. Keratinocytes and neutrophils are important sources of proinflammatory molecules in hidradenitis suppurativa. Br J Dermatol. 2016;174:514–521. doi: 10.1111/bjd.14214. [DOI] [PubMed] [Google Scholar]
- 89.van der Zee HH, Prens EP. The Anti-Inflammatory Drug Colchicine Lacks Efficacy in Hidradenitis Suppurativa. Dermatology. 2011;223:169–173. doi: 10.1159/000332846. [DOI] [PubMed] [Google Scholar]
- 90.Li X, Zhong F. Nickel induces interleukin-1ß secretion via the NLRP3-ASC-caspase-1 pathway. Inflammation. 2014;37:457–466. doi: 10.1007/s10753-013-9759-z. [DOI] [PubMed] [Google Scholar]
- 91.Watanabe H, Gaide O, Petrilli V, Martinon F, Contassot E, Roques S, et al. Activation of the IL-1beta-processing inflammasome is involved in contact hypersensitivity. J Invest Dermatol. 2007;127:1956–1963. doi: 10.1038/sj.jid.5700819. [DOI] [PubMed] [Google Scholar]
- 92.Martin SF, Esser PR, Weber FC, Jakob T, Freudenberg MA, Schmidt M, et al. Mechanisms of chemical induced innate immunity in allergic contact dermatitis. Allergy. 2011;66:1152–1163. doi: 10.1111/j.1398-9995.2011.02652.x. [DOI] [PubMed] [Google Scholar]
- 93.Abramovits W, Rivas Bejarano JJ, Valdecantos WC. Role of Interleukin 1 in Atopic Dermatitis. Dermatol Clin. 2013;31:437–444. doi: 10.1016/j.det.2013.04.008. [DOI] [PubMed] [Google Scholar]
- 94.Krause K, Metz M, Makris M, Zuberbier T, Maurer M. The role of interleukin-1 in allergy-related disorders. Curr Opin Allergy Clin Immunol. 2012;12:477–484. doi: 10.1097/ACI.0b013e3283574d0c. [DOI] [PubMed] [Google Scholar]
- 95.Niebuhr M, Baumert K, Heratizadeh A, Satzger I, Werfel T. Impaired NLRP3 inflammasome expression and function in atopic dermatitis due to Th2 milieu. Allergy. 2014;69:1058–1067. doi: 10.1111/all.12428. [DOI] [PubMed] [Google Scholar]
- 96.Bivik C, Verma D, Winge MC, Lieden A, Bradley M, Rosdahl I, et al. Genetic Variation in the Inflammasome and Atopic Dermatitis Susceptibility. J Invest Dermatol. 2013;133:2486–2489. doi: 10.1038/jid.2013.168. [DOI] [PubMed] [Google Scholar]
- 97.Drexler SK, Bonsignore L, Masin M, Tardivel A, Jackstadt R, Hermeking H, et al. Tissue-specific opposing functions of the inflammasome adaptor ASC in the regulation of epithelial skin carcinogenesis. Proc Natl Acad Sci USA. 2012;109:18384–18389. doi: 10.1073/pnas.1209171109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Gasparoto TH, de Oliveira CE, de Freitas LT, Pinheiro CR, Hori JI, Garlet GP, et al. Inflammasome Activation Is Critical to the Protective Immune Response during Chemically Induced Squamous Cell Carcinoma. PLoS One. 2014;9: doi: 10.1371/journal.pone.0107170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Feldmeyer L, Keller M, Niklaus G, Hohl D, Werner S, Beer HD. The inflammasome mediates UVB-induced activation and secretion of interleukin-1beta by keratinocytes. Curr Biol. 2007;17:1140–1145. doi: 10.1016/j.cub.2007.05.074. [DOI] [PubMed] [Google Scholar]
- 100.Okamoto M, Liu W, Luo Y, Tanaka A, Cai X, Norris DA, et al. Constitutively active inflammasome in human melanoma cells mediating autoinflammation via caspase-1 processing and secretion of interleukin-1beta. J Biol Chem. 2010;285:6477–6488. doi: 10.1074/jbc.M109.064907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Verma D, Bivik C, Farahani E, Synnerstad I, Fredrikson M, Enerbäck C, et al. Inflammasome polymorphisms confer susceptibility to sporadic malignant melanoma. Pigment Cell Melanoma Res. 2012;25:506–513. doi: 10.1111/j.1755-148X.2012.01008.x. [DOI] [PubMed] [Google Scholar]
- 102.Liu W, Luo Y, Dunn JH, Norris DA, Dinarello CA, Fujita M. Dual role of apoptosis-associated speck-like protein containing a CARD (ASC) in tumorigenesis of human melanoma. J Invest Dermatol. 2013;133:518–527. doi: 10.1038/jid.2012.317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Gehrke S, Otsuka A, Huber R, Meier B, Kistowska M, Fenini G, et al. Metastatic melanoma cell lines do not secrete IL-1ß but promote IL-1ß production from macrophages. J Dermatol Sci. 2014;74:167–169. doi: 10.1016/j.jdermsci.2014.01.006. [DOI] [PubMed] [Google Scholar]
- 104.Ahmad I, Muneer KM, Tamimi IA, Chang ME, Ata MO, Yusuf N. Thymoquinone suppresses metastasis of melanoma cells by inhibition of NLRP3 inflammasome. Toxicol Appl Pharmacol. 2013;270:70–76. doi: 10.1016/j.taap.2013.03.027. [DOI] [PubMed] [Google Scholar]
- 105.Ellis LZ, Liu W, Luo Y, Okamoto M, Qu D, Dunn JH, et al. Green tea polyphenol epigallocatechin-3-gallate suppresses melanoma growth by inhibiting inflammasome and IL-1ß secretion. Biochem Biophys Res Commun. 2011;414:551–556. doi: 10.1016/j.bbrc.2011.09.115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.van de Veerdonk FL, Joosten LA, Netea MG. The interplay between inflammasome activation and antifungal host defense. Immunol Rev. 2015;265:172–180. doi: 10.1111/imr.12280. [DOI] [PubMed] [Google Scholar]
- 107.Kistowska M, Fenini G, Jankovic D, Feldmeyer L, Kerl K, Bosshard P, et al. Malassezia yeasts activate the NLRP3 inflammasome in antigen-presenting cells via Syk-kinase signalling. Exp Dermatol. 2014;23:884–889. doi: 10.1111/exd.12552. [DOI] [PubMed] [Google Scholar]
- 108.Hise AG, Tomalka J, Ganesan S, Patel K, Hall BA, Brown GD, et al. An essential role for the NLRP3 inflammasome in host defense against the human fungal pathogen Candida albicans. Cell Host Microbe. 2009;5:487–497. doi: 10.1016/j.chom.2009.05.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Tomalka J, Ganesan S, Azodi , Patel K, Majmudar P, Hall BA, et al. A Novel Role for the NLRC4 Inflammasome in Mucosal Defenses against the Fungal Pathogen Candida albicans. PLoS Pathog. 2011;7: doi: 10.1371/journal.ppat.1002379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Joly S, Ma N, Sadler JJ, Soll DR, Cassel SL, Sutterwala FS. Cutting edge: Candida albicans hyphae formation triggers activation of the Nlrp3 inflammasome. J Immunol. 2009;183:3578–3581. doi: 10.4049/jimmunol.0901323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.van de Veerdonk FL, Lenardon M, Stoffels M, Plantinga T, Smeekens S, et al. The dectin-1/inflammasome pathway is responsible for the induction of protective T-helper 17 responses that discriminate between yeasts and hyphae of Candida albicans. J Leukoc Biol. 2011;90:357–366. doi: 10.1189/jlb.1210702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Mao L, Zhang L, Li H, Chen W, Wang H, Wu S, et al. Pathogenic fungus Microsporum canis activates the NLRP3 inflammasome. Infect Immun. 2014;82:882–892. doi: 10.1128/IAI.01097-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Li H, Wu S, Mao L, Lei G, Zhang L, Lu A, et al. Human pathogenic fungus Trichophyton schoenleinii activates the NLRP3 inflammasome. Protein Cell. 2013;4:529–538. doi: 10.1007/s13238-013-2127-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Gonçalves AC, Maia DC, Ferreira LS, Monnazzi LG, Alegranci P, Placeres MC, et al. Involvement of major components from Sporothrix schenckii cell wall in the caspase-1 activation, nitric oxide and cytokines production during experimental sporotrichosis. Mycopathologia. 2015;179:21–30. doi: 10.1007/s11046-014-9810-0. [DOI] [PubMed] [Google Scholar]
- 115.Nour AM, Reichelt M, Ku CC, Ho MY, Heineman TC, Arvin AM. Varicella-Zoster Virus Infection Triggers Formation of an Interleukin-1ß (IL-1ß)-processing Inflammasome Complex. J Biol Chem. 2011;286:17921–17933. doi: 10.1074/jbc.M110.210575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Reinholz M, Kawakami Y, Salzer S, Kreuter A, Dombrowski Y, Koglin S, et al. HPV16 activates the AIM2 inflammasome in keratinocytes. Arch Dermatol Res. 2013;305:723–732. doi: 10.1007/s00403-013-1375-0. [DOI] [PubMed] [Google Scholar]
- 117.Kerur N, Veettil MV, Sharma-Walia N, Bottero V, Sadagopan S, Otageri P, et al. IFI16 acts as a nuclear pathogen sensor to induce the inflammasome in response to Kaposi sarcoma associated herpesvirus infection. Cell Host Microbe. 2011;9:363–375. doi: 10.1016/j.chom.2011.04.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Johnson K.E., Chikoti L., Chandran B. Herpes simplex virus 1 infection induces activation and subsequent inhibition of the IFI16 and NLRP3 inflammasomes. J Virol. 2013;87:5005–5018. doi: 10.1128/JVI.00082-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Krishna S, Miller LS. Innate and adaptive immune responses against Staphylococcus aureus skin infections. Semin Immunopathol. 2012;34:261–280. doi: 10.1007/s00281-011-0292-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Cho JS, Guo Y, Ramos RI, Hebroni F, Plaisier SB, Xuan C, et al. Neutrophil-derived IL-1ß Is Sufficient for Abscess Formation in Immunity against Staphylococcus aureus in Mice. PLoS Pathog. 2012;8: doi: 10.1371/journal.ppat.1003047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Craven RR, Gao X, Allen IC, Gris D, Bubeck Wardenburg J, McElvania-Tekippe E, et al. Staphylococcus aureus a-Hemolysin Activates the NLRP3-Inflammasome in Human and Mouse Monocytic Cells. PLoS One. 2009;4: doi: 10.1371/journal.pone.0007446. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Li W, Katz BP, Bauer ME, Spinola SM. Haemophilus ducreyi Infection Induces Activation of the NLRP3 Inflammasome in Nonpolarized but Not in Polarized Human Macrophages. Infect Immun. 2013;81:2997–3008. doi: 10.1128/IAI.00354-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Novais FO, Carvalho LP, Passos S, Roos DS, Carvalho EM, Scott P, et al. Genomic profiling of human Leishmania braziliensis lesions identifies transcriptional modules associated with cutaneous immunopathology. J Invest Dermatol. 2015;135:94–101. doi: 10.1038/jid.2014.305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Stojadinovic O, Minkiewicz J, Sawaya A, Bourne JW, Torzilli P, de Rivero Vaccari JP, et al. Deep tissue injury in development of pressure ulcers: a decrease of inflammasome activation and changes in human skin morphology in response to aging and mechanical load. PLoS One. 2013;8: doi: 10.1371/journal.pone.0069223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Hu Y, Liang D, Li X, Liu HH, Zhang X, Zheng M, et al. The Role of Interleukin-1 in Wound Biology: Part I: Murine In Silico and In Vitro Experimental Analysis. Anesth Analg. 2010;111:1525–1533. doi: 10.1213/ANE.0b013e3181f5ef5a. [DOI] [PubMed] [Google Scholar]
- 126.Bitto A, Altavilla D, Pizzino G, Irrera N, Pallio G, Colonna MR, et al. Inhibition of inflammasome activation improves the impaired pattern of healing in genetically diabetic mice. Br J Pharmacol. 2014;171:2300–2307. doi: 10.1111/bph.12557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Mirza RE, Fang MM, Weinheimer-Haus EM, Ennis WJ, Koh TJ. Sustained Inflammasome Activity in Macrophages Impairs Wound Healing in Type 2 Diabetic Humans and Mice. Diabetes. 2014;63:1103–1114. doi: 10.2337/db13-0927. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Weinheimer-Haus EM, Mirza RE, Koh TJ. Nod-Like Receptor Protein-3 Inflammasome Plays an Important Role during Early Stages of Wound Healing. PLoS One. 2015;10: doi: 10.1371/journal.pone.0119106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Weston JK, Uetrecht J. Activation of inflammasomes by agents causing idiosyncratic skin reactions: a possible biomarker. Chem Res Toxicol. 2014;27:949–951. doi: 10.1021/tx5001333. [DOI] [PubMed] [Google Scholar]
- 130.Zhang X, Sharma AM, Uetrecht J. Identification of danger signals in nevirapine-induced skin rash. Chem Res Toxicol. 2013;26:1378–1383. doi: 10.1021/tx400232s. [DOI] [PubMed] [Google Scholar]
- 131.de Rivero Vaccari JP, Sawaya ME, Brand F 3rd, Bauman AJ, Bramlett HM. Caspase-1 Level Is Higher in the Scalp in Androgenetic Alopecia. Pt 1Dermatol Surg. 2012;38(7):1033–1039. doi: 10.1111/j.1524-4725.2012.02378.x. [DOI] [PubMed] [Google Scholar]