

Abstract
Axon regeneration is a chaotic process due largely to unorganized axon alignment. Therefore, in order for a sufficient number of regenerated axons to bridge the lesion site, properly organized axonal alignment is required. Since demyelination after nerve injury strongly impairs the conductive capacity of surviving axons, remyelination is critical for successful functioning of regenerated nerves. Previously, we demonstrated that mesenchymal stem cells (MSCs) aligned on a pre-stretch induced anisotropic surface because the cells can sense a larger effective stiffness in the stretched direction than in the perpendicular direction. We also showed that an anisotropic surface arising from a mechanical pre-stretched surface similarly affects alignment, as well as growth and myelination of axons. Here, we provide a detailed protocol for preparing a pre-stretched anisotropic surface, the isolation and culture of dorsal root ganglion (DRG) neurons on a pre-stretched surface, and show the myelination behavior of a co-culture of DRG neurons with Schwann cells (SCs) on a pre-stretched surface.
Keywords: Neuroscience, Issue 114, DRG isolation, pre-stretch, anisotropy, axon alignment, Schwann cell, myelination.
Introduction
In nerve injuries, the proximal and distal nerve stumps are often prevented from direct realignment of nerve fascicles due to the lesion site 1-2. Normally, axon tracts are composed of highly ordered and aligned bundles of axons, which form complex networks of connectivity. However, nerve regeneration is a chaotic process due to poorly organized axon alignment 3-4. Therefore, to generate a sufficient number of regenerating axons that bridge the lesion site, it is necessary to induce well organized axonal alignment. Additionally, demyelination accompanies nerve injuries due to death of the myelinating cells at the injury site. Since demyelination strongly impairs the conductive capacity of surviving axons, treatments targeting demyelination or promoting remyelination are significant for functional recovery after nerve injury 5. Thus the goal of this protocol is to illustrate an engineering approach that addresses these two issues of nerve regeneration.
Surface anisotropy, which is defined as a difference, when measured along different axes, in a material's physical or mechanical properties, has been applied to influence cell alignment, growth, and migration 6-7. In addition to topography, there are other methods to induce anisotropy. Previously, we investigated surface anisotropy induced by mechanical static pre-stretch of poly-dimethyl-siloxane (PDMS) membrane. The theory of "small deformation super imposed on large" predicted that the effective stiffness the cells sense in the stretched direction differs from the perpendicular direction, and this difference in effective stiffness is due to surface anisotropy 8. Mesenchymal stem cells (MSCs) cultured on a pre-stretched PDMS membrane are able to sense the anisotropy by actively pulling the surface and as a result, align in the pre-stretched direction 9. Similarly, an anisotropic surface arising from a mechanical pre-stretched surface affects the alignment, as well as growth and myelination of dorsal root ganglion (DRG) axons 10. Here we provide a protocol for inducing surface anisotropy on a static pre-stretched PDMS substrate to enhance axon regeneration 10.
To elicit axon alignment, topological features with desired patterns, reported to provide contact guidance through aligned fibers and channels 6,11-12, were demonstrated to facilitate axon alignment 11,13. However, reported techniques for inducing axon alignment through topological features, such as fibers, channels and patterning, were unable to lengthen and increase the thickness of the axons. In contrast, gradual mechanical stretching led to axon alignment in the stretch direction with longer and thicker axons that increased with the magnitude of the stretch 14. However, incorporating a powered motor device in vivo is not feasible. In contrast, static pre-stretched induced anisotropy is less complicated and can be more readily incorporated into future scaffold designs for in vivo applications.
In this protocol, a static pre-stretched cell culture system is used to induce surface anisotropy without topological features. The pre-stretched culture system is composed of a PDMS membrane, a stretchable frame and a stretching stage, whereupon the membrane is fixed onto the frame and a predetermined stretch magnitude is applied on the stretching stage. Freshly isolated DRG neurons cultured on the pre-stretched surface for up to 21 days are monitored for axon alignment and thickness. Subsequently, Schwann cells (SCs) co-cultured with the aligned axons are monitored for myelination. By incorporating pre-stretch induced surface anisotropy we were able to enhance cell alignment-differentiation and axon alignment-growth of MSCs and DRG neurons 9-10, respectively.
Protocol
All procedures for the isolation of the cells were approved by the Institutional Animal Care and Use Committee at Michigan State University.
1. Preparation of Pre-stretched Anisotropic Surface
Mix a 10:1 solution of base and curing agent and pour the mixture into a tissue culture dish (12 cm diameter). Use 4,900 mg base and 490 mg curing agent for the total crosslinking mixture.
Keep the gel mixture under vacuum for 20 min to remove air bubbles.
Place the gel mixture in the oven for overnight curing at 60 °C.
After the PDMS membrane cures, treat the surface with oxygen plasma using a plasma cleaning/etching system for 3 min at 165 mTorr and 65 sccm flow of O2.
Cut a piece of rectangular membrane (5 × 3.5 cm) from the dish, while being cognizant which side is oxygen-treated, make sure the oxygen-treated side is facing up and fix onto a stretching frame. Place the frame on a stretching stage and evenly stretch by turning the knob on the stage until it reaches 10% elongation (or some other pre-determined stretch; the elongation can be directly read from the stretching stage) in the longer axis 9. Fix the stretch by tightening the screws on the frame.
Remove the frame from the stage. Ensure the membrane surface is dry and free of dust, then place a silicone chamber onto the membrane allowing the sticky side of the chamber to attach tightly to the membrane. Note: The silicone chamber provides a well for retaining the cell medium on the PDMS membrane. The device design is shown in Figure 1.
Sterilize the surface with UV for 10 min.
Add 1 ml Poly-L-Lysine (PLL) (0.01% in Phosphate Buffered Saline) to the PDMS surface within the chamber, within 6 hr of plasma treatment and incubate for 2 hr at 37 °C prior to seeding the DRG neurons. This enhances cell attachment.
2. Isolation of DRG
Prepare standard growth medium for the DRG neurons.
Prior to the isolation, autoclave the isolation equipment and filter reagents. Add 5 ml of isolation buffer into one well of a 6-well plate, and place the plate on ice. Prepare 10 ml of dissociation medium by adding 1 ml of collagenase A (500 U/ml) to 9 ml of 0.05% Trypsin-EDTA (1 mg/ml), filter the solutions with a 0.22 µm filter and place on ice.
Use ten to twelve 5 - 7 days' old Sprague-Dawley rats for the isolation. Spray the pups with 75% isopropanol, and sacrifice by decapitation.
Cut away the skin overlying the spinal cord from the back and remove any excess tissue around the spine. Under a surgical magnifier with the surgical spot lights on, make the first incision from the neck and then one cut along the spine on both sides with scissors. Detach the spine from the body of the pups and remove the excess muscles. Then, use scissors to cut along the long axis of the spine and use tweezers to open the spine completely and extract the spinal cord.Remove the tip from a pipette to create a larger opening for transferring the DRGs with isolation buffer into a sterile 15 ml tube.
Using a fine pair of tweezers, remove the ganglion from the bone pocket and collect approximately 10 – 16 DRG from both sides. Trim the nerve roots and transfer the ganglia to the ice-cold isolation buffer.
Remove the tip from a pipette to create a larger opening for transferring the DRGs with isolation buffer into a sterile 15 ml tube. After the chemical dissociation, centrifuge the ganglia at 900 x g and 4 °C for 5 min.
Let the tissues settle to the bottom of the tube and gently remove the isolation buffer on top and add 10 ml of dissociation medium into the tube.
Incubate the dissected tissues in the dissociation medium in a 37 °C water bath for 1 hr while shaking the tube every 5 - 10 min.
After the chemical dissociation, centrifuge the ganglia at 900 x g and 4 °C for 5 min.
Remove the supernatant and re-suspend the pellet in 10 ml standard growth media and vortex.
Centrifuge the dissociated cells as indicated in section 2.9 once again.
Remove the supernatant, re-suspend the pellet in 12 ml of standard growth media and vortex.
3. Culture of DRG on Pre-stretched Surface
Prior to seeding the cells, remove the PLL solutions from the chamber and rinse the PDMS surface with sterile water and air dry.
After re-suspending the cells, let the suspension set for 1 - 2 min to allow the debris to settle to the bottom of the tube. Add 1.5 ml of cell suspension into each stretch chamber and incubate at 37 °C at 5% CO2.
To eliminate glial cells, at 1 day in vitro (DIV), add 10 µl (or 15 µl) mixture of fluoro-2 deoxy-uridine and uridine (FDU-U) stock solution (See Table of Materials) to each well. After 7 hr, replace this medium with fresh standard growth medium and place the pre-stretched culture device in a 37 °C incubator with 5% CO2.
During the cell culture period (2 - 3 weeks), change the media every two days by replacing half the spent medium with fresh medium. Note: After culturing on the stretched and unstretched surfaces for 2 weeks, purified SCs are added to the chamber and cultured for another week (step 4.7).
4. Co-culture of Schwann Cells (SCs) with DRG Neurons on Pre-stretched Surface
- Isolate SCs as described by Dr. Campana's group previously 15.
- Briefly, isolate the SCs from a 1 day old pup. Collect and dissociate the sciatic nerves both chemically (Trypsin-EDTA and Collagenase A) and mechanically (18 gauge needle/10 ml syringe) 15.
- Seed the dissociated cells into poly-D-lysine (PDL) coated T25 flask and culture at 37 °C at 5% CO2. On day 5, purify the cells through antibody selection using Anti-thy 1.1 antibody and Rabbit Complement and subculture the purified cells to passage 2 - 4. Use culture medium containing Dulbecco's Modified Eagle Medium (DMEM), 10% Fetal bovine serum (FBS), 1% Penicillin/Streptomycin (Penn/Strep), 21 µg/ml Bovine Pituitary Extract (BPE), and 4 µM Forskolin.
Remove the culture medium from the SCs, and add 5 ml 0.05% Trypsin-EDTA into the flask. Incubate the cells at 37 °C at 5% CO2 for 2 - 3 min.
Check cells under the optical microscope using a 10X objective to see if they lift up from the flask, then add 5 ml of culture medium and mix with the cells in the flask.
Add the cell suspension to a 15 ml centrifuge tube and centrifuge at 200 x g and 20 °C for 5 min.
Remove the supernatant and resuspend the pellet in 7 ml standard DRG growth media.
Take 10 µl of cell suspension into a 0.5 ml microcentrifuge tube, mix with 10 µl of trypan blue, and then count the cell number using a hemocytometer under optical microscopy using a 10X objective. Dilute the cell suspension to 5,000 cells per ml by adding DRG growth media.
Remove 0.5 ml of medium from the DRG culture in the stretched chamber and add 0.5 ml of SC suspension to the DRG culture.
Culture the cells for 1 week at 37 °C and 5% CO2. Change the media every two days by replacing half the spent medium with fresh standard growth medium for DRG. After 1 week co-culture, process the cells by immunohistochemistry 10.
Representative Results
The pre-stretched cell culture system promoted DRG axon alignment 10. DRG neurons were cultured onto pre-stretched and unstretched surfaces for 12 days. The axons were stained for β-III-tubulin to demonstrate their alignment. Figure 2 compares axon orientation on the pre-stretched and unstretched PDMS substrates after 12 days of culture. The DRG axons aligned parallel to the stretched direction, whereas they showed random alignment and formed an interconnected network on the unstretched PDMS substrate.
In addition to inducing axon alignment, pre-stretch induced anisotropy enhanced the fasciculation of axons. The aligned axons on the stretched surface formed axonal bundles, which differed from the connected axonal network observed on the unstretched surface. These fascicular bundles on the pre-stretched surface resembled nervous tissue tracts in vivo. Figure 3 shows axons of the DRG neurons on pre-stretched and unstretched surfaces after 21 days of culture. As Figure 3 illustrates, the axons on the pre-stretched substrate aggregated together to form bundles, which appeared similar to fascicular tracts. This suggested the pre-stretched surface was able to promote extensive axon growth in the direction of the pre-stretch, which are important in enhancing regeneration of nervous tissue.
To assess myelination, SCs were co-cultured with the aligned axons. After culturing the DRG neurons on the stretched and unstretched surfaces for 2 weeks, purified SCs were added to the chamber and cultured for another week. In Figure 4, the axons of the co-culture are stained with β-III-tubulin (green), and the SCs are stained with P0 (red). P0 is a marker of mature SCs and is also a component of the myelin sheath, as well as considered an indicator of myelination. There is significant co-localization of green and red on the pre-stretched surface, suggestive of SCs attaching to the axons on the stretched surface, but not on the unstretched surface where the green and red stains are randomly distributed. The pre-stretched surfaces enhanced axon alignment, fascicular-like tract formation, and attachment of SCs to the regenerated axons, all critical for myelination.
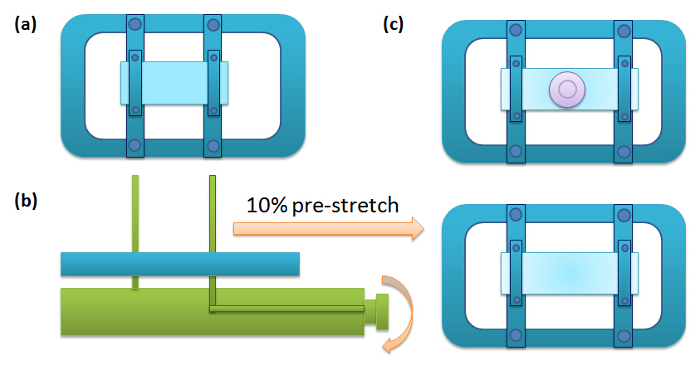 Figure 1: Schematic for the Pre-stretch Procedure. (a) A piece of PDMS membrane is clipped onto two strips of the frame that are slidable. The frame of the device is approximately 7" x 5"; the braces are about 5" x 1; the clamps on the braces are about 3" x .25", and the cup is about 1.5" diameter wide open top, 1" open bottom, and about 0.25" tall. (b) Place the frame onto the stretching stage, turn the roller to apply 10% elongation, and then fix the positions of the strips by tightening the screws. (c) Remove the frame from the stretching stage, and attach a silicon chamber onto the membrane. Please click here to view a larger version of this figure.
Figure 1: Schematic for the Pre-stretch Procedure. (a) A piece of PDMS membrane is clipped onto two strips of the frame that are slidable. The frame of the device is approximately 7" x 5"; the braces are about 5" x 1; the clamps on the braces are about 3" x .25", and the cup is about 1.5" diameter wide open top, 1" open bottom, and about 0.25" tall. (b) Place the frame onto the stretching stage, turn the roller to apply 10% elongation, and then fix the positions of the strips by tightening the screws. (c) Remove the frame from the stretching stage, and attach a silicon chamber onto the membrane. Please click here to view a larger version of this figure.
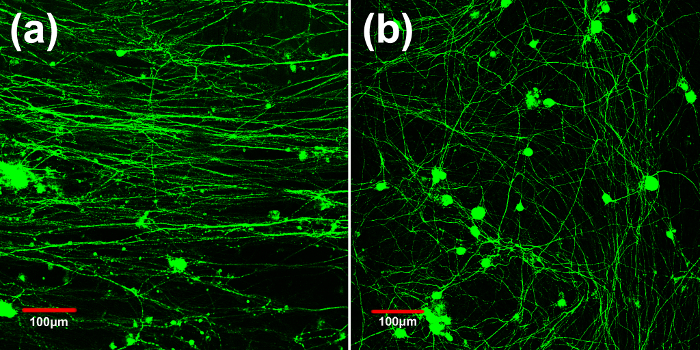 Figure 2:Axon Alignment on Pre-stretched and Unstretched Surface. Fluorescent images of DRG neurons seeded on the (a) static pre-stretched surface for 12 days, compared with DRG cells seeded on the (b) unstretched surface for 12 days. Axons aligned in the pre-stretched direction, while axons aligned randomly on the unstretched control surface. Axons were stained with anti-mouse β-III tubulin primary antibody, and imaged with confocal microscope using a 10X objective. Please click here to view a larger version of this figure.
Figure 2:Axon Alignment on Pre-stretched and Unstretched Surface. Fluorescent images of DRG neurons seeded on the (a) static pre-stretched surface for 12 days, compared with DRG cells seeded on the (b) unstretched surface for 12 days. Axons aligned in the pre-stretched direction, while axons aligned randomly on the unstretched control surface. Axons were stained with anti-mouse β-III tubulin primary antibody, and imaged with confocal microscope using a 10X objective. Please click here to view a larger version of this figure.
 Figure 3:Axon Thickness Growth on Stretched vs. Unstretched PDMS Substrate. Phase contrast images of DRG axons on (a) stretched and (b) unstretched surfaces after 21 days. Axons on the stretched surface formed thicker bundles, while on the unstretched surface the axons formed inter-connected axonal network. Images were taken using inversed phase contrast microscope using a 10X objective. Please click here to view a larger version of this figure.
Figure 3:Axon Thickness Growth on Stretched vs. Unstretched PDMS Substrate. Phase contrast images of DRG axons on (a) stretched and (b) unstretched surfaces after 21 days. Axons on the stretched surface formed thicker bundles, while on the unstretched surface the axons formed inter-connected axonal network. Images were taken using inversed phase contrast microscope using a 10X objective. Please click here to view a larger version of this figure.
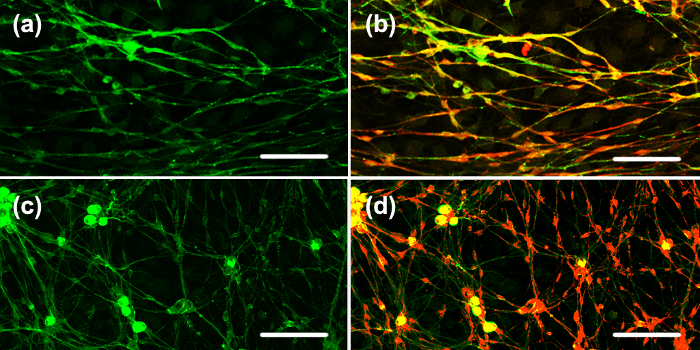 Figure 4:Co-cultured SCs with Aligned DRG Axons on Stretched vs. Unstretched PDMS Substrates. After culturing on the stretched and unstretched surfaces for 2 weeks, purified SCs were added to the chamber and cultured for another week. (a) β-III-tubulin (green) staining for axons on the stretched surface; (b) Overlay of β-III-tubulin (green), P0 (red) staining on the stretched surface; (c) β-III-tubulin (green) staining for axons on the unstretched surface; (d) Overlay of β-III-tubulin (green), P0 (red) staining on the unstretched surface. On the pre-stretched surface, the red and green stains colocalize, suggesting the SCs are attached to and likely myelinating the axons. On the unstretched surface, the red and green stains are randomly distributed, suggesting the SCs are not attached to the axons. Scale bar = 100 µm. Please click here to view a larger version of this figure.
Figure 4:Co-cultured SCs with Aligned DRG Axons on Stretched vs. Unstretched PDMS Substrates. After culturing on the stretched and unstretched surfaces for 2 weeks, purified SCs were added to the chamber and cultured for another week. (a) β-III-tubulin (green) staining for axons on the stretched surface; (b) Overlay of β-III-tubulin (green), P0 (red) staining on the stretched surface; (c) β-III-tubulin (green) staining for axons on the unstretched surface; (d) Overlay of β-III-tubulin (green), P0 (red) staining on the unstretched surface. On the pre-stretched surface, the red and green stains colocalize, suggesting the SCs are attached to and likely myelinating the axons. On the unstretched surface, the red and green stains are randomly distributed, suggesting the SCs are not attached to the axons. Scale bar = 100 µm. Please click here to view a larger version of this figure.
Discussion
To induce axon alignment on pre-stretched surface, there are two critical steps: 1) the PDMS membrane must be flat and of homogenous thickness; and 2) glial cells must be removed from the DRG. After mixing the PDMS and crosslinker and curing in an oven, the crosslinked PDMS gel should be kept on a flat bench top and handled carefully to avoid any tilting. The oxygen plasma treatment of the PDMS membrane should be followed within 6 hr by PLL coating, since the hydrophilicity of the surface (required for cell attachment) after plasma treatment attenuates over time. Since the plasma treatment is applied to one side of the membrane, care must be taken after peeling the PDMS off the dish to ensure the correct side is placed on the frame, that is, the plasma treated surface should be facing up for the cells to attach to.
The DRGs are located inside the bone pockets that align alongside the spine. Since the color of the vertebra is white in young pups, it is difficult to distinguish the DRGs from the vertebra. In this case, it is important to place the spine close to the spot light. Under the light, the DRGs turn transparent while the bone remains white and opaque. It is important to keep the dissection buffer on ice and at a physiological pH, since the pH influences the viability of the neural cells. The isolated tissues are very viscous and tend to stick to the pipette tip when transferring to a centrifuge tube. Thus cutting the very tip of the pipette to generate a larger opening facilitates the transfer.
In some protocols, there is one more centrifugation step required with the isolated tissues before adding the dissociation buffer. In lieu of this step, our protocol requires the tissues to settle in the dissection buffer for a couple of minutes, which reduces the extra centrifugation step that otherwise would impact the viability and density of the cells. The cell density is also dependent on the number of pups sacrificed, and thus is important to maintain a consistent number of pups each time. There are also glial cells that are isolated along with the DRG neurons, thus a mitotic inhibitor, FDU-U, is added to the culture medium at 1 DIV to suppress the growth of the glial cells. This is commonly used to inhibit glial cells during the early growth period of the neurons, although some protocols prefer a combination of arabinofuranosylcytidine (AraC) and FDU-U. The molar concentration of FDU-U added to the culture varies in the different protocols, depending on the density of the isolated cells. We found the mitotic inhibitor can be added multiple times (at the concentration specified in step 3.3), as needed (if glial cells are present), during the entire DRG culture. It is generally not recommended to add a large amount (more than 15 µl) at any given time since it impacts the attachment of the DRG neurons.
Plasma treatment enhances surface hydrophilicity and the PLL coating provides a charged surface for axons to initiate attachment. According to references in the literature, the plasma treatment induced hydrophilicity will attenuate over time, under dry conditions 16-17. However PDMS will undergo a slow reaction with water under liquid conditions and will gradually change from being hydrophobic to hydrophilic 18. Therefore even though the plasma treatment attenuates over time, it should not impact our results. For the PLL coating, according to the product information from the supplier, it is stable over years if placed under favorable conditions. Once the cells attach onto the PLL coating, they generally will not detach. Additionally, the PLL coating helps with the initial attachment, and as the cells grow they secrete Extracellular matrix (ECM) proteins which further help them to remain anchored to the surface 19. Future design improvements could include imbedding peptides into the PDMS network during crosslinking, which would eliminate the plasma treatment and PLL coating steps.
The protocol, herein, described an engineering approach that generated surface anisotropy to successfully induce axon alignment, growth and myelination. The approach can be experimentally expanded in two ways. First, the anisotropic surface could be applied to create nerve tissues in culture, and the in vitro nerve tracts could then be transplanted in vivo. Second, the concept of surface anisotropy could be incorporated into the design of transplantable scaffolds that could be used in vivo to enhance the alignment of the regenerating axons from the injury site and myelination from the host SCs. Compared to other approaches which require multiple and complicated devices 20-22, as well as the addition of growth factors, our platform is relatively simple and easy to apply, and relies only on pre-stretch to enhance both axon alignment and myelination without the need for growth factor addition. Furthermore we have tested this concept of pre-stretch in the experimental design on both MSCs and DRG neurons which could easily be applied to other cell types. The current design was used to investigate axon growth on 2-dimensional surfaces, however in future the device could be upgraded to enable the testing of 3-dimenstional cell cultures. Future applications in nerve repair and transplantation could incorporate pre-stretch into the transplantable scaffolds made from biodegradable natural or synthetic materials containing crosslinked peptides along with controlled-release drug properties.
Disclosures
The authors do not have any conflict of interest to disclose.
Acknowledgments
The authors would like to thank Eric Vasco for his assistance in the preparation of the PDMS substrates, Dr. Shiyong Wang in Dr. Marina Mata's lab at University of Michigan for helpful suggestions and training of the DRG isolation, and Dr. Mark Tuszynski and Dr. W. Marie Campana at UC San Diego for helpful suggestions and protocol for the SC isolation. This study was supported in part by the National Science Foundation (CBET 0941055 and CBET 1510895), the National Institute of Health (R21CA176854, R01GM089866, and R01EB014986).
References
- Liu C, et al. Layer-by-Layer Films for Biomedical Applications. Wiley-VCH Verlag GmbH & Co. KGaA; 2015. pp. 525–546. [Google Scholar]
- Faweett JW, Keynes RJ. Peripheral Nerve Regeneration. Annu Rev Neurosci. 1990;13:43–60. doi: 10.1146/annurev.ne.13.030190.000355. [DOI] [PubMed] [Google Scholar]
- Li Y, Field PM, Raisman G. Repair of adult rat corticospinal tract by transplants of olfactory ensheathing cells. Science. 1997;277:2000–2002. doi: 10.1126/science.277.5334.2000. [DOI] [PubMed] [Google Scholar]
- Geller HM, Fawcett JW. Building a bridge: Engineering spinal cord repair. Exp Neurol. 2002;174:125–136. doi: 10.1006/exnr.2002.7865. [DOI] [PubMed] [Google Scholar]
- Totoiu MO, Keirstead HS. Spinal cord injury is accompanied by chronic progressive demyelination. J Comp Neurol. 2005;486:373–383. doi: 10.1002/cne.20517. [DOI] [PubMed] [Google Scholar]
- Chua JS, et al. Extending neurites sense the depth of the underlying topography during neuronal differentiation and contact guidance. Biomaterials. 2014;35:7750–7761. doi: 10.1016/j.biomaterials.2014.06.008. [DOI] [PubMed] [Google Scholar]
- Dowell-Mesfin NM, et al. Topographically modified surfaces affect orientation and growth of hippocampal neurons. J Neural Eng. 2004;1:78–90. doi: 10.1088/1741-2560/1/2/003. [DOI] [PubMed] [Google Scholar]
- Baek S, Gleason RL, Rajagopal KR, Humphrey JD. Theory of small on large: Potential utility in computations of fluid-solid interactions in arteries. Comput Method Appl M. 2007;196:3070–3078. [Google Scholar]
- Liu C, et al. Effect of Static Pre-stretch Induced Surface Anisotropy on Orientation of Mesenchymal Stem Cells. Cell Mol Bioeng. 2014;7:106–121. doi: 10.1007/s12195-013-0300-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu C, et al. The impact of pre-stretch induced surface anisotropy on axon regeneration. Tissue Eng Part C Methods. 2015. [DOI] [PMC free article] [PubMed]
- Berns EJ, et al. Aligned neurite outgrowth and directed cell migration in self-assembled monodomain gels. Biomaterials. 2014;35:185–195. doi: 10.1016/j.biomaterials.2013.09.077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kidambi S, Lee I, Chan C. Primary neuron/astrocyte co-culture on polyelectrolyte multilayer films: A template for studying astrocyte-mediated oxidative stress in neurons. Adv Funct Mater. 2008;18:294–301. doi: 10.1002/adfm.200601237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xia H, et al. Directed neurite growth of rat dorsal root ganglion neurons and increased colocalization with Schwann cells on aligned poly(methyl methacrylate) electrospun nanofibers. Brain Research. 2014;1565:18–27. doi: 10.1016/j.brainres.2014.04.002. [DOI] [PubMed] [Google Scholar]
- Smith DH. Stretch growth of integrated axon tracts: Extremes and exploitations. Prog Neurobiol. 2009;89:231–239. doi: 10.1016/j.pneurobio.2009.07.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mantuano E, Jo M, Gonias SL, Campana WM. Low Density Lipoprotein Receptor-related Protein (LRP1) Regulates Rac1 and RhoA Reciprocally to Control Schwann Cell Adhesion and Migration. Journal of Biological Chemistry. 2010;285:14259–14266. doi: 10.1074/jbc.M109.085126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim B, ET KP, Papautsky I. Long-term stability of plasma oxidized PDMS surfaces. Conf Proc IEEE Eng Med Biol Soc. 2004;7:5013–5016. doi: 10.1109/IEMBS.2004.1404385. [DOI] [PubMed] [Google Scholar]
- Lopera S, Mansano RD. Plasma-Based Surface Modification of Polydimethylsiloxane for PDMS-PDMS Molding. ISRN Polymer Science. 2012;2012 [Google Scholar]
- Chandra G. Organosilicon Materials. Berlin Heidelberg: Springer; 2013. [Google Scholar]
- Wu MH. Simple poly(dimethylsiloxane) surface modification to control cell adhesion. Surf Interface Anal. 2009;41:11–16. [Google Scholar]
- Moore MJ, et al. Multiple-channel scaffolds to promote spinal cord axon regeneration. Biomaterials. 2006;27:419–429. doi: 10.1016/j.biomaterials.2005.07.045. [DOI] [PubMed] [Google Scholar]
- Clarke JC, et al. Micropatterned methacrylate polymers direct spiral ganglion neurite and Schwann cell growth. Hearing Res. 2011;278:96–105. doi: 10.1016/j.heares.2011.05.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pfister BJ, et al. Development of transplantable nervous tissue constructs comprised of stretch-grown axons. J Neurosci Methods. 2006;153:95–103. doi: 10.1016/j.jneumeth.2005.10.012. [DOI] [PubMed] [Google Scholar]