

Abstract
This study uses cellular c-fos activation to assess effects of novel ingestion of fat and sugar on brain dopamine (DA) pathways in rats. Intakes of sugars and fats are mediated by their innate attractions as well as learned preferences. Brain dopamine, especially meso-limbic and meso-cortical projections from the ventral tegmental area (VTA), has been implicated in both of these unlearned and learned responses. The concept of distributed brain networks, wherein several sites and transmitter/peptide systems interact, has been proposed to mediate palatable food intake, but there is limited evidence empirically demonstrating such actions. Thus, sugar intake elicits DA release and increases c-fos-like immunoreactivity (FLI) from individual VTA DA projection zones including the nucleus accumbens (NAC), amygdala (AMY) and medial prefrontal cortex (mPFC) as well as the dorsal striatum. Further, central administration of selective DA receptor antagonists into these sites differentially reduce acquisition and expression of conditioned flavor preferences elicited by sugars or fats. One approach by which to determine whether these sites interacted as a distributed brain network in response to sugar or fat intake would be to simultaneous evaluate whether the VTA and its major mesotelencephalic DA projection zones (prelimbic and infralimbic mPFC, core and shell of the NAc, basolateral and central-cortico-medial AMY) as well as the dorsal striatum would display coordinated and simultaneous FLI activation after oral, unconditioned intake of corn oil (3.5%), glucose (8%), fructose (8%) and saccharin (0.2%) solutions. This approach is a successful first step in identifying the feasibility of using cellular c-fos activation simultaneously across relevant brain sites to study reward-related learning in ingestion of palatable food in rodents.
Keywords: Neuroscience, Issue 114, Behavioral neuroscience, C-fos immunoreactivity, Sugar intake, Fat intake, Reward, Palatability, Amygdala, Medial prefrontal cortex, Nucleus accumbens, Caudate/putamen, Ventral tegmental area, Distributed brain network
Introduction
Brain dopamine (DA) has been implicated in central responses to intake of palatable sugars through proposed hedonic1,2, effort-related3 and habit-based4,5 mechanisms of action. The primary DA pathway implicated in these effects originates in the ventral tegmental area (VTA), and projects to the nucleus accumbens (NAC) core and shell, the basolateral and central-cortico-medial amygdala (AMY), and the prelimbic and infralimbic medial prefrontal cortex (mPFC) (see reviews6,7). The VTA has been implicated in sucrose intake8,9, and DA release is observed following sugar intake in the NAC10-15, AMY16,17 and mPFC18-20. Fat intake also stimulates DA NAC release21, and another DA-rich projection zone to the dorsal striatum (caudate-putamen) has been also associated with DA-mediated feeding22,23. Kelley24-27 proposed that these multiple projection zones of this DA-mediated system formed an integrated and interactive distributed brain network through extensive and intimate interconnections28-34.
In addition to the ability of DA D1 and D2 receptor antagonists to reduce intake of sugars35-37 and fats38-40, DA signaling has also been implicated in mediating the ability of sugars and fats to produce conditioned flavor preferences (CFP)41-46. Microinjections of a DA D1 receptor antagonist into the NAC, AMY or mPFC47-49 eliminate acquisition of CFP elicited by intragastric glucose. Whereas microinjections of either DA D1 or D2 receptor antagonists into the mPFC eliminates acquisition of fructose-CFP50, the acquisition and expression of fructose-CFP are differentially blocked by DA antagonists in the NAC and AMY51,52.
The c-fos technique53,54 has been employed to investigate neural activation induced by palatable intake and neural activation. The term "c-fos activation" will be used throughout the manuscript, and is operationally defined by increased transcription of c-Fos during neuronal depolarization. Sucrose intake increased fos-like immunoreactivity (FLI) in the central AMY nucleus, the VTA as well as the shell, but not core, of the NAC55-57. Whereas sucrose intake in sham-feeding rats significantly increased FLI in the AMY and the NAC, but not the VTA58, intragastric sucrose or glucose infusions significantly increased FLI in the NAC and central and basolateral nuclei of the AMY59,60. Repeated addition of sucrose to scheduled chow access increased FLI in the mPFC as well as the NAC shell and core61. A sucrose concentration downshift paradigm revealed that the greatest FLI increases occurred in the basolateral AMY and NAC, but not the VTA62. Following conditioning, extinction of sugar-related natural reward behaviors increased FLI in the basolateral AMY and the NAC63. Moreover, pairing sugar availability to a tone resulted in the tone subsequently increasing FLI levels in the basolateral AMY64. High-fat intake also increased FLI in NAC and mPFC sites65-67.
Most of the previously cited studies examined sugar and fat effects on c-fos activation in single sites that do not provide information about identification of reward-related distributed brain networks24-27. Further, many of the studies also did not delineate the relative contributions of sub-areas of the NAC (core and shell), AMY (basolateral and central-cortico-medial) and mPFC (prelimbic and infralimbic) that could potentially be examined by the advantage of excellent spatial, single-cell resolution in c-Fos mapping68. Our laboratory69 recently used c-fos activation and simultaneously measured alterations in the VTA DA pathway and its projection zones (NAC, AMY and mPFC) after novel ingestion of fats and sugars in rats. The present study describes the procedural and methodological steps to simultaneously analyze whether acute exposure to six different solutions (corn oil, glucose, fructose, saccharin, water and a fat emulsion control) would differentially activate FLI in sub-areas of the NAC, AMY, mPFC as well as the dorsal striatum. This simultaneous detection of differences allowed confirmation of significant effects on FLI in each site and determination as to whether changes in one particular site correlated with changes in related sites, thereby providing support for a distributed brain network24-27. These procedures tested whether the VTA, the prelimbic and infralimbic mPFC, the core and shell of the NAC, and the basolateral and central-cortico-medial AMY) as well as the dorsal striatum would display coordinated and simultaneous FLI activation after oral, unconditioned intake of glucose (8%), fructose (8%), corn oil (3.5%) and saccharin (0.2%) solutions.
Protocol
These experimental protocols have been approved by the Institutional Animal Care and Use Committee certifying that all subjects and procedures are in compliance with the National Institutes of Health Guide for Care and Use of Laboratory Animals.
1. Subjects
Purchase and/or breed male Sprague-Dawley rats (260 - 300 g).
House rats individually in wire mesh cages. Maintain them on a 12:12 hr light/dark cycle with rat chow and water available ad libitum.
Assign appropriate sample sizes (e.g., n ≈ 6 - 8) randomly into groups.
2. Testing Apparatus and Intake Procedures
Use calibrated centrifuge tubes with rubber stoppers and a 45° angle metal sipper tube to provide accurate measurement (± 0.1 ml) of the presented solutions. Secure them to the home cages by a taut metal spring to allow visibility of the calibrations.
Restrict food rations (~ 15/g/day) of the rats to reduce weight to 85% of their original body weight to increase motivation to consume the solutions. Note: The weight reduction should take between 3 - 5 days.
Provide pre-training solutions (10 ml) of 0.2% saccharin for four days over a 1 hr session to maximize the probability that rats will sample the subsequent test solutions with short (less than 1 min) latency.
Confirm flow through the centrifuge tube by spilling a few drops.
Weigh tubes before and after each session to obtain intake measurement.
Perform an intake test on the fifth day on subgroups receiving one of six solutions (10 ml, 1 hr): a) water, b) novel flavored (0.05% cherry flavor) 0.2% saccharin, c) 8% fructose, d) 8% glucose, e) 3.5% corn oil suspended in 0.3% xanthan gum, and f) 0.3% xanthan gum.
Ensure that the nutrient solutions are isocaloric; thus, the 3.5% corn-oil concentration is isocaloric to the 8% sugar solutions.
Ensure that rats sample solutions with short latency (less than 1 min). If this requirement is not met, then discard the subject from the study.
3. Tissue Preparation
Anesthetize each animal by an intraperitoneal injection of pentobarbital 90 min after initial exposure to each test solution. Confirm that animals are properly anesthetized by demonstrating that the animal is no longer responsive to such reflexes as withdrawal to foot pinch, blinking following direct corneal pressure or head shaking to deep pinna stimulation.
- Perfuse each animal transcardially as described previously69.
- Anesthetize rats with an overdose of sodium pentobarbital (65 mg/kg), remove the ribcage and expose the chest for free access to the heart69.
- Place the needle in the apex of the left heart valve, and cut the vena cava. Administer phosphate buffer solution (PBS, ~ 180 ml) followed by a phosphate-buffered fixative containing 4% paraformaldehyde (~ 180 ml).
- Ensure that the animal actually is being perfused correctly by examining whether liquid is leaving other cavities, such as the nose, mouth, and genital areas. Note: Proper fixation with paraformaldehyde will be accompanied by large muscle movements. If this does not occur, re-adjust the needle until this reaction occurs.
Remove the brain from the skull quickly by cutting fur and skin away from the skull. Use rongeurs to crack and remove the bone from the brain moving from rear to front. Work initially in the area below and behind the cerebellum, ensuring that the rongeur is between the bone and meningeal pia mater. Once the top and sides of the skull are removed, use a small spatula to lift the brain from the base, and snip cranial nerves with small scissors. Be careful not to damage the brain while attempting to remove the bone.
Fix the brains in a 4% paraformaldehyde solution overnight at 4 °C. Place the brains in a 30% sucrose/70% PBS solution at room temperature until they settle at the bottom of the container.
- Block the brain
- Remove the rostral part of the brain cutting transversely caudal to the olfactory bulb.
- Remove the caudal part of the brain cutting transversely at the level of the cerebellum and the pons.
Mount the brain coronally with the caudal part fixed to the stage of a sliding microtome, and cut coronal sections (40 µm) through the mPFC (+2.86 – +2.20 mm rostral to bregma), the NAC core and shell and dorsal striatum (+1.76 – +1.60 mm rostral to bregma), the AMY (-2.12 – -2.92 mm caudal to bregma), and the VTA (-5.20 – -5.60 mm caudal to bregma). Use a rat brain atlas70 for guidance.
Collect free-floating sections into individual wells of a 24-well plate filled with PBS for eventual immunohistochemical analysis71. Use Parafilm to seal the 24 well plate to make sure the PBS does not evaporate in the container and dry up the brain. Store the brain tissue in 4 °C.
4. c-fos Procedures (Adapted from 71)
Treat each section with 5 ml of 5% Normal Goat Serum and 0.2% Triton x-100 in PBS for 1 hr.
Incubate the treated sections with primary antibodies (rabbit anti-c-fos, 1:5,000) at 4 °C for 36 hr in wells containing 1 ml of PBS.
Rinse sections 3x with PBS (5 ml) for 10 min each.
Incubate with secondary antibodies (biotinylated goat anti-rabbit; 1:200) at RT for 2 hr in wells containing 1 ml of PBS.
Rinse each section 3x in PBS (5 ml) for 10 min each.
Incubate the rinsed sections for 2 hr in a commercially available avidin-horseradish peroxidase mixture that comes in a kit composed of Avadin DH (100 µl) and Biotinylated Horseradish Peroxidase H (100 µl) in 5 ml of PBS.
Re-rinse the sections 3x in PBS (5 ml) for 10 min each.
React the sections with 0.05% diaminobenzidine (DAB) in the presence of 0.0015% H2O2 for 5 - 10 min, depending on the reactivity of the tissue in wells containing 5 ml of the DAB solution.
Double-label the VTA sections. Incubate them with a tyrosine hydroxylase (TH) antibody (rabbit anti-rat TH, 1:2,000) in PBS (5 ml) overnight at 4 °C.
Rinse sections 3x in PBS (5 ml) for 10 min each.
Incubate with secondary antibodies (biotinylated goat anti-rabbit; 1:200) in PBS (5 ml) at room temperature for 2 hr.
Rinse sections 3x in PBS (5 ml) for 10 min each.
Visualize the antibodies by using a secondary antibody-peroxidase complex. React with a combination of 0.05% DAB and a 0.3% nickel sulfate solution, for 5 - 10 min, depending on the reaction of the tissue in wells containing 5 ml of the DAB/NiCl solution.
Ensure that the DAB solution is milky light green in color from its reaction with the 0.3% nickel sulfate. If the solution is too green, then the reaction will be too dark.
Mount all sections onto gelatin-coated slides. Let them dry overnight, and then cover-slip with a few drops of a Toluene-Based Solution (TBS).
Code slides so that the experimental condition is unknown to the observers.
5. Determination of c-fos Immunoreactive Counts
Assign pairs of unbiased observers to count Fos-positive neurons in these regions of interest (ROI): prelimbic mPFC, infralimbic mPFC, NAC core, NAC shell, basolateral AMY nuclei, central-cortico-medial AMY, dorsal striatum, and VTA. Delineate whether c-Fos immunoreactivity was present in TH+ and TH- cells in the VTA. Figure 1 provides a screen-captured picture of the NAC from the microscope.
 Figure 1. Representative Section of the Nucleus Accumbens (NAC) Displaying Regions of Interest Outlined by the Grid by which c-fos Counts are Made. The NAC shell is found medially and ventrally to the NAC core. The NAC core encircles the anterior commissure (Anterior Comm.). The ventral extent of the lateral ventricle (Lateral Vent.) is visible. Please click here to view a larger version of this figure.
Figure 1. Representative Section of the Nucleus Accumbens (NAC) Displaying Regions of Interest Outlined by the Grid by which c-fos Counts are Made. The NAC shell is found medially and ventrally to the NAC core. The NAC core encircles the anterior commissure (Anterior Comm.). The ventral extent of the lateral ventricle (Lateral Vent.) is visible. Please click here to view a larger version of this figure.
Analyze at least three representative slices per site common to all animals in all of the testing conditions.
- Use software and an optical microscope to analyze the entire region for each ROI by tracing an outline (Figure 1).
- For a given site, open the application and click on the acquisition drop-down menu and click "Live Image". Bring the ROI into focus and click the screen to establish a reference point. Then trace the chosen brain region using the grid as a guide. Once the trace is completed, count cells (steps 5.3.1.1 - 5.3.1.3).
- Double-click the software icon. Go to the menu bar, click on "Acquisition" and then "Live Image". Bring the ROI into focus and click the screen to establish a reference point.
- Go to the grid toolbar and click "Display Grid" and "Use grid labels". Outline the ROI with a predetermined trace.
- Count all cells in each ROI area, select a "+" in the left hand sidebar to keep count of c-fos cells. Click each cell singly to register counts. Consider a cell positive for c-fos when a defined dark red circle is observed (Figure 1).
- Repeat this process for each site.
- Record counts in a lab notebook and on the computer for future analysis. Go to the menu bar, click on "File", "Save Data File" to save the tracing and counts.
Ensure that inter-rater reliability (using correlation of counts) of the two uninformed raters for each section in each ROI always exceeds 0.8.
6. Statistics
Evaluate baseline saccharin intakes over the first four days using a repeated-measures 1-way analysis of variance (ANOVA) comparing the saccharin intakes of Days 1, 2, 3 and 469.
Compare saccharin intake (Day 4) with test intakes (Day 5) of the six groups using a randomized-block 2-way ANOVA69.
Use Tukey comparisons (p < 0.05) to determine individual significant effects69.
Determine inter-rater reliability, and then use a common observer's counts.
- Average c-fos counts for the three representative slices for each site69.
- Perform a 1-way ANOVA of c-fos activation induced by intake of the six solutions (3.5% corn oil, 8% glucose, 8% fructose, 0.2% flavored saccharin, xanthan gum control and water) for the perilimbic mPFC69.
- Repeat parallel analyses of the six groups for infralimbic mPFC, NAC core, NAC shell, basolateral AMY, central-cortico-medial AMY, VTA and dorsal striatum. Use Tukey comparisons (p < 0.05) to reveal individual significant effects69.
Compare corn oil intake with both water intake and intake of its suspension agent, xanthan gum. Compare fructose and glucose intakes with both water intake and intake of the non-nutritive sweetener, saccharin.
- Establish whether significant relationships between solution intakes and c-fos activation in each of the sites were observed using Bonferroni r correlations (p < 0.05).
- Systematically compare c-fos counts in the perilimbic and infralimbic pre-frontal cortex for each animal in the 3.5% corn oil group.
- Repeat systematically parallel analyses of each pair of the six sites (VTA, dorsal striatum, infralimbic mPFC, perilimbic mPFC, NAC core, NAC shell, basolateral AMY, central-cortico-medial AMY) for the 3.5% corn oil.
- Repeat systematically parallel analyses of these six sites for the other five experimental intake conditions (8% glucose, 8% fructose, 0.2% flavored saccharin, xanthan gum control and water).
Take advantage of the fact that the same animals within a solution condition were evaluated across all sites by determining significant relationships between c-fos activation across solutions and within each solution using Bonferroni r correlations (p < 0.05).
Representative Results
All representative results described below have been published previously69, and are re-presented here to support "proof of concept" in indicating the effectiveness of the technique.
Solution Intakes Significant differences in baseline saccharin intakes were observed over the first four days for all animals (F(3,108) = 57.27, p < 0.001) with intakes (Day 1: 1.3 (± 0.2) ml; Day 2: 3.9 (± 0.4) ml; Day 3: 5.9 (± 0.6) ml; Day 4: 7.1 (± 0.6) ml) significantly (p < 0.05, Tukey HSD test) and progressively increasing. Fructose and glucose intake, but not corn oil or saccharin intake on Day 5 significantly (p < 0.05, Tukey HSD test) increased relative to Day 4 saccharin intake (p < 0.05, Tukey HSD test) with fructose (9.6 (± 0.4) ml) and glucose (9.4 (± 0.6) ml) significantly higher than saccharin intake. Further, corn oil intake (7.4 (± 0.6) ml) was significantly (p < 0.05, Tukey HSD test) higher than xanthan gum intake.
These results raised the possibility that solution intake per se might account for any observed c-fos activation in any of the sites. To examine this, Bonferroni r correlations were performed in which intake of the five solutions was related to c-fos activation in each of the six sites. Significant correlations failed to be observed between solution intake and c-fos activation in the core (r(29) = 0.186), shell (r(29) = 0.029) or total (r(29) = 0.10) NAc, the prelimbic (r(29) = 0.23), infralimbic (r(29) = 0.30) or total (r(29) = 0.14) mPFC, the VTA (r(29) = 0.10), the dorsal striatum (r(29) = 0.14) or the basolateral (r(29) = 0.47), the centro-cortico-medial (r(29) = 0.48) or total (r(29) = 0.409) AMY. Given the higher correlation between intake and AMY c-fos activation, further correlations were performed for each individual solution. Significant (p < 0.05, Tukey HSD test) relationships failed to be observed between intake and AMY FLI for fructose (basolateral (r = 0.15), centro-cortico-medial (r = 0.13), total (r = 0.13)), glucose (basolateral (r = 0.17), centro-cortico-medial (r = 0.17), total r = 0.13)),saccharin (basolateral (r = 0.42), centro-cortico-medial (r = 0.42), total (r = 0.42)) or corn oil (basolateral (r = 0.54), centro-cortico-medial (r = 0.59), total (r = 0.64)). A significant (p < 0.05, Tukey HSD test) negative correlation was observed between xanthan gum intake and total AMY FLI(r = 0.94).
mPFC c-Fos Activation Corn oil significantly (p < 0.05, Tukey HSD test) increased total (Figure 2A), infralimbic (Figure 2B) and prelimbic (Figure 2C) mPFC c-Fos counts relative to water (*) or the xanthan gum control (#). Fructose significantly (p < 0.05, Tukey HSD test) increased c-Fos counts in the infralimbic mPFC relative to water (*) or saccharin (+) (Figure 2B), but not total or prelimbic mPFC counts. In contrast, glucose or saccharin failed to alter total, perilimbic or infralimbic mPFC c-Fos counts. Figure 3 displays representative mPFC sections of animals showing increased corn oil-induced FLI relative to water.
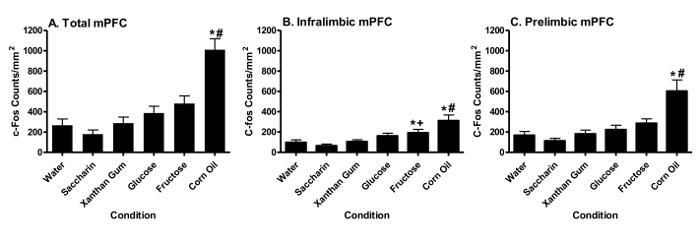 Figure 2. Fat or Sugar Intake Differentially Increases c-fos Activation in the Medial Prefrontal Cortex (mPFC). Changes in c-fos activation (mean ± S.E.M.) are noted in the entire mPFC (Panel A), the infralimbic mPFC area (Panel B), and the prelimbic mPFC area (Panel C) following consumption (1 hr) of water, saccharin (0.2%), xanthan gum (control for corn oil), glucose (8%), fructose (8%) or corn oil (3.5%). (Previously published69.) Please click here to view a larger version of this figure.
Figure 2. Fat or Sugar Intake Differentially Increases c-fos Activation in the Medial Prefrontal Cortex (mPFC). Changes in c-fos activation (mean ± S.E.M.) are noted in the entire mPFC (Panel A), the infralimbic mPFC area (Panel B), and the prelimbic mPFC area (Panel C) following consumption (1 hr) of water, saccharin (0.2%), xanthan gum (control for corn oil), glucose (8%), fructose (8%) or corn oil (3.5%). (Previously published69.) Please click here to view a larger version of this figure.
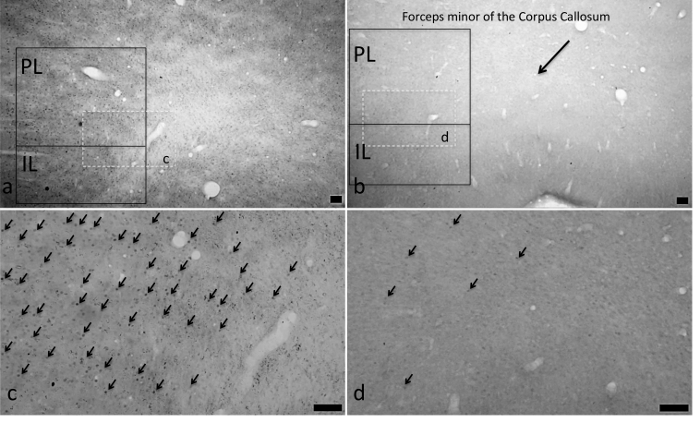 Figure 3.Actual mPFC c-fos Activation Following Fats and Sugars. C-fos activation was observed in animals exposed to intake of corn oil (Panels A (4-fold magnification) and C (10-fold magnification)) that was significantly greater than that of intake of water (Panels B (4-fold magnification) and D (10-fold magnification)). Representation of the delineated sub-areas of the Prelimbic (PL) and infralimbic (IL) mPFC are diagramed in Panels A (corn oil) and C (water) as are the part of those panels (A and C) magnified to 10-fold magnification in the corresponding panels (B and D). Arrows in Panels C and D indicate representative c-fos positive cells. All scale bars are 100 µm. (Previously published69.) Please click here to view a larger version of this figure.
Figure 3.Actual mPFC c-fos Activation Following Fats and Sugars. C-fos activation was observed in animals exposed to intake of corn oil (Panels A (4-fold magnification) and C (10-fold magnification)) that was significantly greater than that of intake of water (Panels B (4-fold magnification) and D (10-fold magnification)). Representation of the delineated sub-areas of the Prelimbic (PL) and infralimbic (IL) mPFC are diagramed in Panels A (corn oil) and C (water) as are the part of those panels (A and C) magnified to 10-fold magnification in the corresponding panels (B and D). Arrows in Panels C and D indicate representative c-fos positive cells. All scale bars are 100 µm. (Previously published69.) Please click here to view a larger version of this figure.
AMY c-Fos Activation Corn oil significantly (p < 0.05, Tukey HSD test) increased total AMY (Figure 4A), basolateral (Figure 4B) and central-cortico-medial (Figure 4C) sub-area AMY c-Fos counts relative to water (*) or the xanthan gum control (#). Glucose also significantly (p < 0.05, Tukey HSD test) increased total (Figure 4A), basolateral (Figure 4B) and central-cortico-medial (Figure 4C) sub-area AMY c-Fos counts relative to water (*) or saccharin (+).Glucose and fructose significantly (p < 0.05, Tukey HSD test) increased AMY FLI relative to saccharin or water. Fructose significantly (p < 0.05, Tukey HSD test) increased total (Figure 4A) and central-cortico-medial sub-area (Figure 4C) of the AMY c-Fos counts relative to water (*) or saccharin (+), but not in the basolateral AMY sub-area. Saccharin failed to alter total, basolateral or central-cortico-medial AMY c-Fos counts relative to water. Detailed analyses of individual nuclei within the AMY revealed that the significant changes noted in the basolateral area of the AMY were also noted in the individual basolateral and lateral AMY nuclei. The significant changes noted in the central-cortico-medial area of the AMY were also noted in the individual central, cortical and medial AMY nuclei. Figure 5 displays representative AMY sections of animals showing increased corn oil-, glucose-, and fructose-induced FLI relative to water. (Previously published69.)
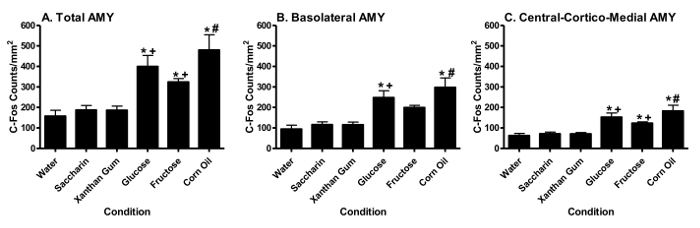 Figure 4. Fat or Sugar Intake Differentially Increases c-fos Activation in the Amygdala (AMY). Changes in c-fos activation (mean ± S.E.M.) are noted in the entire AMY (Panel A), the basolateral AMY area (Panel B), and the central-cortico-medial AMY area (Panel C) following consumption (1 hr) of water, saccharin, xanthan gum, glucose, fructose or corn oil. The significant changes noted in the basolateral area of the AMY were also noted in the individual basolateral and lateral AMY nuclei. The significant changes noted in the central-cortico-medial area of the AMY were also noted in the individual central, cortical and medial AMY nuclei. (Previously published69.) Please click here to view a larger version of this figure.
Figure 4. Fat or Sugar Intake Differentially Increases c-fos Activation in the Amygdala (AMY). Changes in c-fos activation (mean ± S.E.M.) are noted in the entire AMY (Panel A), the basolateral AMY area (Panel B), and the central-cortico-medial AMY area (Panel C) following consumption (1 hr) of water, saccharin, xanthan gum, glucose, fructose or corn oil. The significant changes noted in the basolateral area of the AMY were also noted in the individual basolateral and lateral AMY nuclei. The significant changes noted in the central-cortico-medial area of the AMY were also noted in the individual central, cortical and medial AMY nuclei. (Previously published69.) Please click here to view a larger version of this figure.
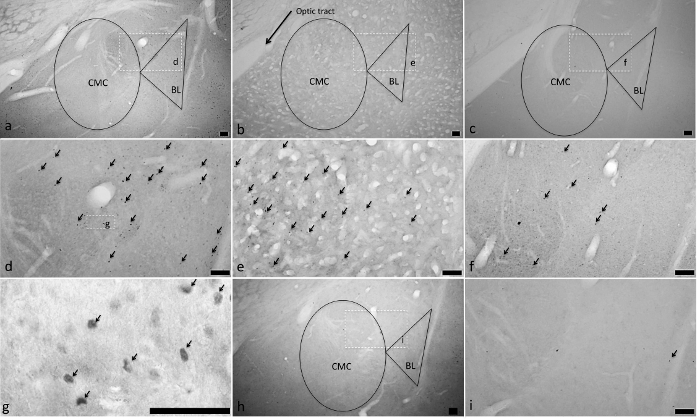 Figure 5. Actual AMY c-fos Activation Following Fats and Sugars. C-fos activation was observed in animals exposed to intakes of corn oil (Panels A (4-fold magnification), D (10-fold magnification) and G (60-fold magnification)), glucose (Panels B (4-fold magnification) and E (10-fold magnification)), and fructose (Panels C (4-fold magnification) and F (10-fold)) that were significantly greater than that of intake of water (Panels H (4-fold magnification) and I (10-fold magnification)). Representation of the delineated sub-areas of the central-cortico-medial (CMC) and the basolateral (BLA) AMY are diagramed in Panels A (corn oil), B (glucose), C (fructose) and H (water) as are the part of those panels (A, B, C and H) magnified to10-fold magnification in the corresponding panels (D, E, F and I). The delineated sub-area in Panel D (corn oil, 10-fold magnification) is magnified to 60-fold magnification in Panel D. Arrows in Panels D, E, F, G and I indicate representative c-fos positive cells. All scale bars are 100 µm, except for Panel G (50 µm). (Previously published69.) Please click here to view a larger version of this figure.
Figure 5. Actual AMY c-fos Activation Following Fats and Sugars. C-fos activation was observed in animals exposed to intakes of corn oil (Panels A (4-fold magnification), D (10-fold magnification) and G (60-fold magnification)), glucose (Panels B (4-fold magnification) and E (10-fold magnification)), and fructose (Panels C (4-fold magnification) and F (10-fold)) that were significantly greater than that of intake of water (Panels H (4-fold magnification) and I (10-fold magnification)). Representation of the delineated sub-areas of the central-cortico-medial (CMC) and the basolateral (BLA) AMY are diagramed in Panels A (corn oil), B (glucose), C (fructose) and H (water) as are the part of those panels (A, B, C and H) magnified to10-fold magnification in the corresponding panels (D, E, F and I). The delineated sub-area in Panel D (corn oil, 10-fold magnification) is magnified to 60-fold magnification in Panel D. Arrows in Panels D, E, F, G and I indicate representative c-fos positive cells. All scale bars are 100 µm, except for Panel G (50 µm). (Previously published69.) Please click here to view a larger version of this figure.
NAC c-Fos Activation Corn oil significantly (p < 0.05, Tukey HSD test) increased total (Figure 6A) and core (Figure 6B) NAc c-Fos counts relative to water (*), but not the NAc shell (Figure 6C). Glucose significantly (p < 0.05, Tukey HSD test) increased c-Fos counts in the NAc core (Figure 6B), but not in the total NAc or the NAc shell relative to saccharin (+) or water (*). In contrast, fructose and saccharin failed to differ from water in eliciting c-Fos activation in the NAc core and/or shell. Figure 7 displays representative sections in the NAc core of animals showing increased corn oil- or glucose-induced FLI relative to water.
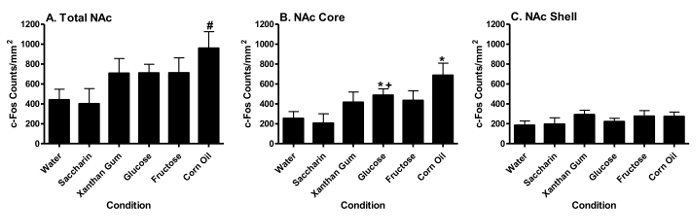 Figure 6. Fat or Sugar Intake Differentially Increases c-fos Activation in the NAC. Changes in c-fos activation (mean ± S.E.M.) in the entire NAC (Panel A), the NAC core (Panel B), and the NAC shell (Panel C) following consumption (1 hr) of water, saccharin, xanthan gum, glucose, fructose or corn oil. (Previously published69.) Please click here to view a larger version of this figure.
Figure 6. Fat or Sugar Intake Differentially Increases c-fos Activation in the NAC. Changes in c-fos activation (mean ± S.E.M.) in the entire NAC (Panel A), the NAC core (Panel B), and the NAC shell (Panel C) following consumption (1 hr) of water, saccharin, xanthan gum, glucose, fructose or corn oil. (Previously published69.) Please click here to view a larger version of this figure.
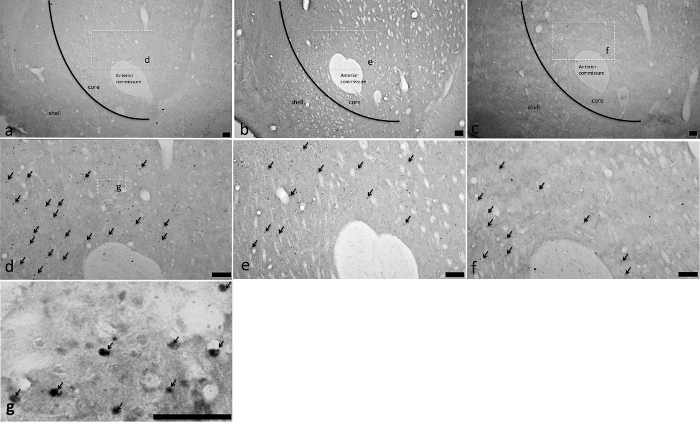 Figure 7. Actual NAC Core, but not NAC Shell c-fos Activation Following Fats and Sugars. C-fos activation was observed in animals exposed to intakes of corn oil (Panels A (4-fold magnification), D (10-fold magnification) and G (60-fold magnification)) and glucose (Panels B (4-fold magnification) and E (10-fold magnification)) that were significantly greater than that of intake of water (Panels C (4-fold magnification) and F (10-fold magnification)). Representation of the delineated sub-areas of the NAc core and the NAc shell are diagramed in Panels A (corn oil), B (glucose) and C (water) as are the part of those panels (A, B, C and H) magnified to10-fold magnification in the corresponding panels (D, E and F). The delineated sub-area in Panel D (corn oil, 10-fold magnification) is magnified to 60-fold magnification in Panel G. Arrows in Panels D, E, F and G indicate representative c-fos positive cells. All scale bars are 100 µm. (Previously published69.) Please click here to view a larger version of this figure.
Figure 7. Actual NAC Core, but not NAC Shell c-fos Activation Following Fats and Sugars. C-fos activation was observed in animals exposed to intakes of corn oil (Panels A (4-fold magnification), D (10-fold magnification) and G (60-fold magnification)) and glucose (Panels B (4-fold magnification) and E (10-fold magnification)) that were significantly greater than that of intake of water (Panels C (4-fold magnification) and F (10-fold magnification)). Representation of the delineated sub-areas of the NAc core and the NAc shell are diagramed in Panels A (corn oil), B (glucose) and C (water) as are the part of those panels (A, B, C and H) magnified to10-fold magnification in the corresponding panels (D, E and F). The delineated sub-area in Panel D (corn oil, 10-fold magnification) is magnified to 60-fold magnification in Panel G. Arrows in Panels D, E, F and G indicate representative c-fos positive cells. All scale bars are 100 µm. (Previously published69.) Please click here to view a larger version of this figure.
Dorsal Striatal c-Fos Activation Corn oil significantly (p < 0.05, Tukey HSD test) increased c-Fos counts in the dorsal striatum relative to water (*) or the xanthan gum (#) (Figure 8A). Glucose or fructose significantly (p < 0.05, Tukey HSD test) increased dorsal striatal FLI relative to saccharin (+) (Figure 8A). In contrast, saccharin failed to differ from water in eliciting dorsal striatal c-Fos activation. Figure 9 displays representative dorsal striatal sections of animals showing increased corn oil-, glucose- or fructose-induced FLI relative to water.
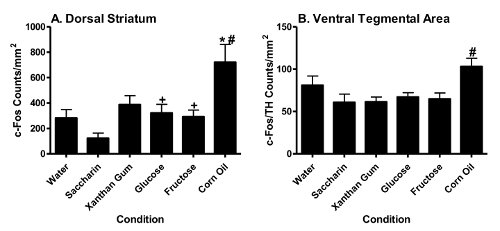 Figure 8. Fat or Sugar Intake Differentially Increases c-fos Activation in the Dorsal Striatum and Ventral Tegmental Area. Dorsal atriatal (Panel A) and ventral tegmental area (Panel B) alterations were noted for c-fos activation (mean ±S.E.M.) following consumption (1 hr) of water, saccharin, xanthan gum, glucose, fructose or corn oil. (Previously published69.) Please click here to view a larger version of this figure.
Figure 8. Fat or Sugar Intake Differentially Increases c-fos Activation in the Dorsal Striatum and Ventral Tegmental Area. Dorsal atriatal (Panel A) and ventral tegmental area (Panel B) alterations were noted for c-fos activation (mean ±S.E.M.) following consumption (1 hr) of water, saccharin, xanthan gum, glucose, fructose or corn oil. (Previously published69.) Please click here to view a larger version of this figure.
 Figure 9. Actual Dorsal Striatal c-fos Activation Following Fats and Sugars. C-fos activation was observed in animals exposed to intakes of corn oil (Panels A (4-fold magnification), D (10-fold magnification) and G (60-fold magnification)), glucose (Panels B (4-fold magnification) and E (10-fold magnification)), fructose(Panels C (4-fold magnification) and F (10-fold magnification)) that were significantly greater than that of intake of water (Panels H (4-fold magnification) and I (10-fold magnification)). Delineated sub-areas of the dorsal striatum in Panels A (corn oil), B (glucose), C (fructose) and H (water) are indicated that magnified to10-fold magnification in the corresponding panels (D, E, F and I). The delineated sub-area in Panel D (corn oil, 10-fold magnification) is magnified to 60-fold magnification in Panel G. Arrows in Panels D, E, F and G indicate representative c-fos positive cells. All scale bars are 100 µm, except for Panel G (50 µm). (Previously published69.) Please click here to view a larger version of this figure.
Figure 9. Actual Dorsal Striatal c-fos Activation Following Fats and Sugars. C-fos activation was observed in animals exposed to intakes of corn oil (Panels A (4-fold magnification), D (10-fold magnification) and G (60-fold magnification)), glucose (Panels B (4-fold magnification) and E (10-fold magnification)), fructose(Panels C (4-fold magnification) and F (10-fold magnification)) that were significantly greater than that of intake of water (Panels H (4-fold magnification) and I (10-fold magnification)). Delineated sub-areas of the dorsal striatum in Panels A (corn oil), B (glucose), C (fructose) and H (water) are indicated that magnified to10-fold magnification in the corresponding panels (D, E, F and I). The delineated sub-area in Panel D (corn oil, 10-fold magnification) is magnified to 60-fold magnification in Panel G. Arrows in Panels D, E, F and G indicate representative c-fos positive cells. All scale bars are 100 µm, except for Panel G (50 µm). (Previously published69.) Please click here to view a larger version of this figure.
VTA c-Fos Activation Corn oil significantly (p<0.05, Tukey HSD test) increased c-Fos counts in TH+ VTA cells relative to the xanthan gum control (#) (Figure 8B). In contrast, glucose, fructose or saccharin failed to alter c-Fos counts in the VTA relative to water. Figure 10 displays representative TH+ and TH- and c-Fos-activated VTA cells of animals showing increased corn oil-induced FLI relative to water.
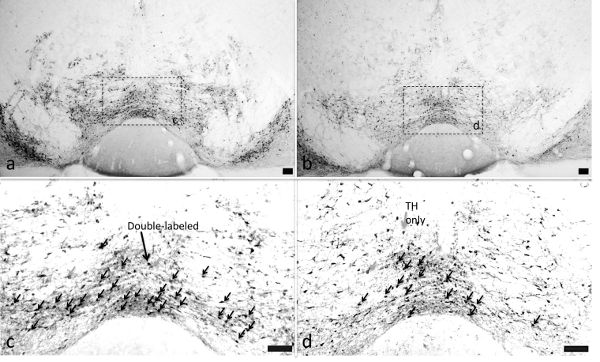 Figure 10. Actual Ventral Tegmental Area c-fos Activation Following Fats and Sugars. VTA c-fos activation was observed in animals exposed to corn oil (Panels A (4-fold) and C (10-fold)) and water (Panels B (4-fold) and D (10-fold)). Black arrows indicate representative double-labeled TH/c-fos positive cells, while gray arrows indicate representative c-fos only cells. All scale bars are 100 µm. (Previously published69.) Please click here to view a larger version of this figure.
Figure 10. Actual Ventral Tegmental Area c-fos Activation Following Fats and Sugars. VTA c-fos activation was observed in animals exposed to corn oil (Panels A (4-fold) and C (10-fold)) and water (Panels B (4-fold) and D (10-fold)). Black arrows indicate representative double-labeled TH/c-fos positive cells, while gray arrows indicate representative c-fos only cells. All scale bars are 100 µm. (Previously published69.) Please click here to view a larger version of this figure.
Relationships of c-Fos Activation Among Sites and Solutions The pattern of c-Fos counts in animals exposed to corn oil revealed significant (p < 0.05) positive correlations between the NAc core and either the NAc shell (r = 0.971) or entire mPFC (r = 0.670), between the prelimbic mPFC and either the infralimbic mPFC (r = 0.940) or dorsal striatum (r = 0.849), between the infralimbic mPFC and dorsal striatum (r = 0.749), between the basolateral and central-cortico-medial AMY (r = 0.999), and between the dorsal striatum and the VTA (r = 0.723). In contrast, the pattern of c-Fos counts in animals exposed to corn oil revealed significant (p < 0.05) negative correlations between the basolateral AMY and either the NAc core (r = -0.712) or shell (r = -0.708), and between the central-cortico-medial AMY and either the NAc core (r = -0.712) or shell (r = -0.710).The pattern of c-Fos counts in animals exposed to glucose revealed significant (p < 0.05) positive correlations between the prelimbic and infralimbic mPFC (r = 0.930), between the dorsal striatum and either the VTA (r = 0.821), basolateral (r = 0.910) or central-cortico-medial (r = 0.911) AMY, and between the basolateral and central-cortico-medial (r = 0.999) AMY. The pattern of c-Fos counts in animals exposed to fructose revealed significant (p < 0.05) positive correlations between the NAc core and either the NAc shell (r = 0.969) or prelimbic mPFC (r = 0.740),between the NAc shell and the prelimbic mPFC (r = 0.733),between the prelimbic and infralimbic mPFC (r = 0.959), and between the basolateral and central-cortico-medial AMY (r = 0.996).The pattern of c-Fos counts in animals exposed to saccharin revealed significant (p < 0.05) positive correlations between the NAc core and NAc shell (r = 0.792), between the NAc shell and dorsal striatum (r = 0.715), and between the prelimbic mPFC and infralimbic mPFC (r = 0.999).
Discussion
The goal of the study was to determine if the source (VTA) and forebrain projection targets (NAC, AMY, mPFC) of DA reward-related neurons were simultaneously activated after novel ingestion of fat and sugar in rats using the cellular c-fos technique. The present study is a detailed description of the protocols of a study published previously69. It was hypothesized that the VTA, its major projection zones to the prelimbic and infralimbic mPFC, the core and shell of the NAC and the basolateral and central-cortico-medial AMY, as well as the dorsal striatum would act as a distributed brain network24-27, and display coordinated and simultaneous FLI following novel intake of glucose (8%), fructose (8%) or corn oil (3.5%) solutions relative to saccharin (0.2%), water and other control solutions. Corn oil, glucose and fructose, but not saccharin intake produced significant and differential FLI activation of the VTA, the prelimbic and infralimbic mPFC, the core and shell of the NAC, the basolateral and central-cortico-medial AMY, and the dorsal striatum. In addition to the c-fos technique, behavioral measures of sugar, fat and artificial sweetener intake were employed.
One critical step included timely sampling of intake such that they would be comparatively equal, thereby ensuring that any differences in c-fos activation across sites were due to the solution being consumed rather the either the pattern or magnitude of intake. The four days of baseline saccharin intake ensured that the food-restricted animals sampled the solutions quickly, and thereby minimized non-specific effects. A second critical step was that the procedure caused minimal stress or novelty to the animals as changes in emotional valence independent of intake type could also produce c-fos activation. Therefore, the findings provide a convincing "proof of concept" for the effectiveness of this approach and protocols related to identify whether acute exposure to fat (e.g., corn oil), sugar (glucose and fructose) and non-nutritive sweetener (saccharin) solutions simultaneously activate DA-mediated ROI's in manner suggestive of a coordinated distributed brain system24-27.
Because optimal c-Fos activation requires time-sensitive responses prior to sacrifice51,52, previously validated procedures42,44 maximized solution sampling with short latency in the 1-hr test. Thus, food restricted rats were trained with 0.2% saccharin solutions (10 ml, 1 hr) for 4 days, and given the test solution on the fifth day. Baseline saccharin intakes significantly and progressively increased, and fructose and glucose, but not corn oil or saccharin intakes on the fifth day were significantly higher than fourth day saccharin intake. Hence, solutions associated with increased FLI significantly increased (glucose, fructose) or failed to affect (corn oil) intake relative to previous saccharin training, and appeared to be mediated through a reward-related behavioral incentive mechanism. Careful consideration needs to be taken to ensure sampling and equality of behavior. Other researchers can effectively use this procedure to study other types of novel solutions or introduce variations in the paradigm to understand mechanisms related to adaptation and learning.
The advantage of the present protocol is the ability to compare effects of well-studied sugars (fructose, glucose) and fats (corn oil) and compare their c-fos activating effects with than of important controls (the non-nutritive sweetener, saccharin, a control emulsifier, xanthan gum, and water), and then examine these effects across six related brain sites. Although this approach has obvious benefits in allowing simultaneous examination across brain sites of the different palatable substances, it has the drawback of producing a potentially astronomical data set of cells displaying neuronal expression. To make this more manageable, we took the approach of analyzing three representative coronal slices per site common to all animals in all of the testing conditions. This of course is accompanied by the caveat of choosing the appropriate levels of each ROI in these three sections. Given the broad rostro-caudal extent of the AMY, NAC, mPFC, dorsal striatum and VTA, this caveat should not be taken lightly. Further, it is then incumbent upon the investigators to be consistent in accurately selecting each of the three representative sections across all animals across all sites. Minor mistakes in this choice can lead to "false positives" and "false negatives". Efficiency of counting also is a relevant variable. Our solution for this potential confound was to assign two uninformed raters for each section in each ROI, and then ensure that inter-rater reliability (using correlation of counts) always exceeded 0.8. This approach, while duplicative, gave us far greater assurance about accuracy as the inter-rater reliability easily exceeded this minimum criterion. Sub-regions of the NAC (core vs. shell), AMY (baso-lateral vs. central-cortico-medial) and mPFC (perilimbic vs. infralimbic) were analyzed. These regions could be divided further, particularly the individual AMY nuclei, the patch and matrix compartments of the dorsal striatum, and the NAC shell (vertex, arch, cone, intermediate zone). Because the NAC shell failed to consistently display changes in FLI following corn oil, glucose or fructose, further sub-analyses of this structure was not performed. Definitive examination of the patch and matrix zones of the dorsal striatum required further immunohistochemical techniques that were not employed in the present study, but would be an important follow-up study. Analyses of individual AMY nuclei within each sub-region would also be an additional future study.
Previous studies showed that sucrose intake increased FLI in the central AMY nucleus, the VTA as well as the shell, but not core, of the NAC, yet oral or IG saccharin infusions are largely ineffective 55-57, 60-62. Glucose and fructose intake elicited sugar-specific effects upon FLI with both effective in the central-cortico-medial AMY and dorsal striatum, the former effective in the NAc core and the basolateral AMY, and the latter effective in the infralimbic mPFC. Saccharin intake failed to elicit any changes in FLI in any site relative to water. Fat intake also increased FLI in accumbal and mPFC sites in previous studies65-67, and produced simultaneous significant activation in the VTA, infralimbic and prelimbic mPFC, dorsal striatum, NAC core, and the basolateral and central-cortico-medial AMY.
Although previous studies demonstrated that sugar and fat intake induced FLI in forebrain meso-corticolimbic and nigro-striatal DA systems, the present study systematically evaluated simultaneous FLI activation in the VTA, basolateral and central-cortico-medial AMY, dorsal striatum, prelimbic and infralimbic mPFC, NAc core and shell following acute intake of corn oil, fructose, glucose or saccharin. Significant FLI increases were highly related to each other across forebrain sites, supporting the idea of distributed brain network activation mediating sugar and fat intake. Such protocols identifying simultaneous changes in multiple brain loci can be utilized under chronic and binging conditions as well as under conditioning and preferences. These studies show that a strong anatomical correlate (c-fos) can be effectively used in multiple brain sites simultaneously to identify candidates for mediating palatable intake and preferences in animals that may provide insights into human medical conditions related to obesity, diabetes and other eating disorders.
Disclosures
The authors have no competing financial interests.
Acknowledgments
Thanks to Diana Icaza-Culaki, Cristal Sampson and Theologia Karagiorgis for their hard work on this project.
References
- Koob GF. Neural mechanisms of drug reinforcement. Ann. N.Y. Acad. Sci. 1992;654:171–191. doi: 10.1111/j.1749-6632.1992.tb25966.x. [DOI] [PubMed] [Google Scholar]
- Wise RA. Role of brain dopamine in food reward. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2006;361:1149–1158. doi: 10.1098/rstb.2006.1854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salamone JD, Correa M. The mysterious motivational functions of mesolimbic dopamine. Neuron. 2012;76:470–485. doi: 10.1016/j.neuron.2012.10.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Horvitz JC, Choi WY, Morvan C, Eyny Y, Balsam PD. A "good parent" function of dopamine: transient modulation of learning and performance during early stages of training. Ann. N.Y. Acad. Sci. 2007;1104:270–288. doi: 10.1196/annals.1390.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wickens JR, Horvitz JC, Costa RM, Killcross S. Dopaminergic mechanisms in actions and habits. J. Neurosci. 2007;27:8181–8183. doi: 10.1523/JNEUROSCI.1671-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bjorklund A, Dunnett SB. Dopamine neuron systems in the brain: an update. Trends Neurosci. 2007;30:194–202. doi: 10.1016/j.tins.2007.03.006. [DOI] [PubMed] [Google Scholar]
- Swanson LW. The projections of the ventral tegmental area and adjacent regions: a combined fluorescent retrograde tracer and immunofluorescence study in the rat. Brain Res. Bull. 1982;9:321–353. doi: 10.1016/0361-9230(82)90145-9. [DOI] [PubMed] [Google Scholar]
- Cacciapaglia F, Wrightman RM, Careli RM. Rapid dopamine signaling differentially modulates distinct microcircuits within the nucleus accumbens during sucrose-directed behavior. J. Neurosci. 2011;31:13860–13869. doi: 10.1523/JNEUROSCI.1340-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martinez-Hernandez J, Lanuza E, Martinez-Garcia F. Selective dopaminergic lesions of the ventral tegmental area impair preference for sucrose but not for male sex pheromones in female mice. Eur. J. Neurosci. 2006;24:885–893. doi: 10.1111/j.1460-9568.2006.04944.x. [DOI] [PubMed] [Google Scholar]
- Bassareo V, Di Chiara G. Differential influence of associative and nonassociative learning mechanisms on the responsiveness of prefrontal and accumbal dopamine transmission to food stimuli in rats fed ad libitum. J. Neurosci. 1997;17:851–861. doi: 10.1523/JNEUROSCI.17-02-00851.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bassareo V, Di Chiara G. Differential responsiveness of dopamine transmission to food-stimuli in nucleus accumbens shell/core compartments. Neurosci. 1999;89:637–641. doi: 10.1016/s0306-4522(98)00583-1. [DOI] [PubMed] [Google Scholar]
- Cheng J, Feenstra MG. Individual differences in dopamine efflux in nucleus accumbens shell and core during instrumental conditioning. Learn. Mem. 2006;13:168–177. doi: 10.1101/lm.1806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Genn RF, Ahn S, Phillips AG. Attenuated dopamine efflux in the rat nucleus accumbens during successive negative contrast. Behav. Neurosci. 2004;118:869–873. doi: 10.1037/0735-7044.118.4.869. [DOI] [PubMed] [Google Scholar]
- Hajnal A, Norgren R. Accumbens dopamine mechanisms in sucrose intake. Brain Res. 2001;904:76–84. doi: 10.1016/s0006-8993(01)02451-9. [DOI] [PubMed] [Google Scholar]
- Hajnal A, Smith GP, Norgren R. Oral sucrose stimulation increases accumbens dopamine in the rat. Am. J. Physiol. 2003;286:R31–R37. doi: 10.1152/ajpregu.00282.2003. [DOI] [PubMed] [Google Scholar]
- Bassareo V, Di Chiara G. Modulation of feeding-induced activation of mesolimbic dopamine transmission by appetitive stimuli and its relation to motivational state. Eur. J. Neurosci. 1999;11:4389–4397. doi: 10.1046/j.1460-9568.1999.00843.x. [DOI] [PubMed] [Google Scholar]
- Hajnal A, Lenard L. Feeding-related dopamine in the amygdala of freely moving rats. Neuroreport. 1997;8:2817–2820. doi: 10.1097/00001756-199708180-00033. [DOI] [PubMed] [Google Scholar]
- Bassareo V, De Luca MA, Di Chiara G. Differential expression of motivational stimulus properties by dopamine in nucleus accumbens shell versus core and prefrontal cortex. J. Neurosci. 2002;22:4709–4719. doi: 10.1523/JNEUROSCI.22-11-04709.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feenstra M, Botterblom M. Rapid sampling of extracellular dopamine in the rat prefrontal cortex during food consumption, handling, and exposure to novelty. Brain Res. 1996;742:17–24. doi: 10.1016/s0006-8993(96)00945-6. [DOI] [PubMed] [Google Scholar]
- Hernandez L, Hoebel BG. Feeding can enhance dopamine turnover in the prefrontal cortex. Brain Res. Bull. 1990;25:975–979. doi: 10.1016/0361-9230(90)90197-8. [DOI] [PubMed] [Google Scholar]
- Liang NC, Hajnal A, Norgren R. Sham feeding corn oil increases accumbens dopamine in the rat. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006;291:R1236–R1239. doi: 10.1152/ajpregu.00226.2006. [DOI] [PubMed] [Google Scholar]
- Dunnett SB, Iversen SD. Regulatory impairments following selective kainic acid lesions of the neostriatum. Behav. Brain Res. 1980;1:497–506. doi: 10.1016/0166-4328(80)90004-2. [DOI] [PubMed] [Google Scholar]
- Salamone JD, Zigmond MJ, Stricker EM. Characterization of the impaired feeding behavior in rats given haloperidol or dopamine-depleting brain lesions. Neurosci. 1990;39:17–24. doi: 10.1016/0306-4522(90)90218-s. [DOI] [PubMed] [Google Scholar]
- Kelley AE. Ventral striatal control of appetitive motivation: role in ingestive behavior and reward-related learning. Neurosci. Biobehav. Rev. 2004;27:765–776. doi: 10.1016/j.neubiorev.2003.11.015. [DOI] [PubMed] [Google Scholar]
- Kelley AE. Memory and addiction: shared neural circuitry and molecular mechanisms. Neuron. 2004;44:161–179. doi: 10.1016/j.neuron.2004.09.016. [DOI] [PubMed] [Google Scholar]
- Kelley AE, Baldo BA, Pratt WE. A proposed hypothalamic-thalamic-striatal axis for the integration of energy balance, arousal and food reward. J. Comp. Neurol. 2005;493:72–85. doi: 10.1002/cne.20769. [DOI] [PubMed] [Google Scholar]
- Kelley AE, Baldo BA, Pratt WE, Will MJ. Corticostriatal-hypothalamic circuitry and food motivation: integration of energy, action and reward. Physiol. Behav. 2005;86:773–795. doi: 10.1016/j.physbeh.2005.08.066. [DOI] [PubMed] [Google Scholar]
- Berendse HW, Galis-de-Graaf Y, Groenewegen HJ. Topographical organization and relationship with ventral striatal compartments of prefrontal corticostriatal projections in the rat. J. Comp. Neurol. 1992;316:314–347. doi: 10.1002/cne.903160305. [DOI] [PubMed] [Google Scholar]
- Brog JS, Salyapongse A, Deutch AY, Zahm DS. The patterns of afferent innervation of the core and shell in the "accumbens" part of rat ventral striatum: immunohistochemical detection of retrogradely transported fluoro-gold. J. Comp. Neurol. 1993;338:255–278. doi: 10.1002/cne.903380209. [DOI] [PubMed] [Google Scholar]
- McDonald AJ. Organization of amygdaloid projections to the prefrontal cortex and associated stritum in the rat. Neurosci. 1991;44:1–14. doi: 10.1016/0306-4522(91)90247-l. [DOI] [PubMed] [Google Scholar]
- McGeorge AJ, Faull RL. The organization of the projection from the cerebral cortex to the striatum in the rat. Neurosci. 1989;29:503–537. doi: 10.1016/0306-4522(89)90128-0. [DOI] [PubMed] [Google Scholar]
- Sesack SR, Deutch AY, Roth RH, Bunney BS. Topographical organization of the efferent projections of the medial prefrontal cortex in the rat: an anterograde tract-tracing study with Phaseolus vulgaris leucoagglutinin. J. Comp. Neurol. 1989;290:213–242. doi: 10.1002/cne.902900205. [DOI] [PubMed] [Google Scholar]
- Wright CI, Beijer AV, Groenewegen HJ. Basal amygdaloid complex afferents to the rat nucleus accumbens are compartmentally organized. J. Neurosci. 1996;16:1877–1893. doi: 10.1523/JNEUROSCI.16-05-01877.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wright CI, Groenewegen HJ. Patterns of convergence and segregation in the medial nucleus accumbens of the rat: relationships of prefrontal cortical, midline thalamic and basal amygdaloid afferents. J. Comp. Neurol. 1995;361:383–403. doi: 10.1002/cne.903610304. [DOI] [PubMed] [Google Scholar]
- Geary N, Smith GP. Pimozide decreases the positive reinforcing effect of sham fed sucrose in the rat. Pharmacol. Biochem. Behav. 1985;22:787–790. doi: 10.1016/0091-3057(85)90528-3. [DOI] [PubMed] [Google Scholar]
- Muscat R, Willner P. Effects of selective dopamine receptor antagonists on sucrose consumption and preference. Psychopharmacol. 1989;99:98–102. doi: 10.1007/BF00634461. [DOI] [PubMed] [Google Scholar]
- Schneider LH, Gibbs J, Smith GP. D-2 selective receptor antagonists suppress sucrose sham feeding in the rat. Brain Res. Bull. 1986;17:605–611. doi: 10.1016/0361-9230(86)90231-5. [DOI] [PubMed] [Google Scholar]
- Baker RW, Osman J, Bodnar RJ. Differential actions of dopamine receptor antagonism in rats upon food intake elicited by mercaptoacetate or exposure to a palatable high-fat diet. Pharmacol. Biochem. Behav. 2001;69:201–208. doi: 10.1016/s0091-3057(01)00528-7. [DOI] [PubMed] [Google Scholar]
- Rao RE, Wojnicki FH, Coupland J, Ghosh S, Corwin RL. Baclofen, raclopride and naltrexone differentially reduce solid fat emulsion intake under limited access conditions. Pharmacol. Biochem. Behav. 2008;89:581–590. doi: 10.1016/j.pbb.2008.02.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weatherford SC, Smith GP, Melville LD. D-1 and D-2 receptor antagonists decrease corn oil sham feeding in rats. Physiol. Behav. 1988;44:569–572. doi: 10.1016/0031-9384(88)90320-4. [DOI] [PubMed] [Google Scholar]
- Azzara AV, Bodnar RJ, Delamater AR, Sclafani A. D1 but not D2 dopamine receptor antagonism blocks the acquisition of a flavor preference conditioned by intragastric carbohydrate infusions. Pharmacol. Biochem. Behav. 2001;68:709–720. doi: 10.1016/s0091-3057(01)00484-1. [DOI] [PubMed] [Google Scholar]
- Baker RM, Shah MJ, Sclafani A, Bodnar RJ. Dopamine D1 and D2 antagonists reduce the acquisition and expression of flavor-preferences conditioned by fructose in rats. Pharmacol. Biochem. Behav. 2003;75:55–65. doi: 10.1016/s0091-3057(03)00039-x. [DOI] [PubMed] [Google Scholar]
- Dela Cruz JA, Coke T, Icaza-Cukali D, Khalifa N, Bodnar RJ. Roles of NMDA and dopamine D1 and D2 receptors in the acquisition and expression of flavor preferences conditioned by oral glucose in rats. Neurobiol. Learn. Mem. 2014;114:223–230. doi: 10.1016/j.nlm.2014.07.004. [DOI] [PubMed] [Google Scholar]
- Dela Cruz JA, et al. Roles of dopamine D1 and D2 receptors in the acquisition and expression of fat-conditioned flavor preferences in rats. Neurobiol. Learn. Mem. 2012;97:332–337. doi: 10.1016/j.nlm.2012.01.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu WZ, Silva RM, Sclafani A, Delamater AR, Bodnar RJ. Pharmacology of flavor preference conditioning in sham-feeding rats: effects of dopamine receptor antagonists. Pharmacol. Biochem. Behav. 2000;65:635–647. doi: 10.1016/s0091-3057(99)00239-7. [DOI] [PubMed] [Google Scholar]
- Yu WZ, Silva RM, Sclafani A, Delamater AR, Bodnar RJ. Role of D(1) and D(2) dopamine receptors in the acquisition and expression of flavor-preference conditioning in sham-feeding rats. Pharmacol. Biochem. Behav. 2000;67(1):537–544. doi: 10.1016/s0091-3057(00)00396-8. [DOI] [PubMed] [Google Scholar]
- Touzani K, Bodnar RJ, Sclafani A. Activation of dopamine D1-like receptors in nucleus accumbens is critical for the acquisition, but not the expression, of nutrient-conditioned flavor preferences in rats. Eur. J. Neurosci. 2008;27:1525–1533. doi: 10.1111/j.1460-9568.2008.06127.x. [DOI] [PubMed] [Google Scholar]
- Touzani K, Bodnar RJ, Sclafani A. Dopamine D1-like receptor antagonism in amygdala impairs the acquisition of glucose-conditioned flavor preference in rats. Eur. J. Neurosci. 2009;30:289–298. doi: 10.1111/j.1460-9568.2009.06829.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Touzani K, Bodnar RJ, Sclafani A. Acquisition of glucose-conditioned flavor preference requires the activation of dopamine D1-like receptors within the medial prefrontal cortex in rats. Neurobiol. Learn. Mem. 2010;94:214–219. doi: 10.1016/j.nlm.2010.05.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Malkusz DC, et al. Dopamine signaling in the medial prefrontal cortex and amygdala is required for the acquisition of fructose-conditioned flavor preferences in rats. Behav. Brain Res. 2012;233:500–507. doi: 10.1016/j.bbr.2012.05.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bernal SY, et al. Role of dopamine D1 and D2 receptors in the nucleus accumbens shell on the acquisition and expression of fructose-conditioned flavor-flavor preferences in rats. Behav. Brain Res. 2008;190:59–66. doi: 10.1016/j.bbr.2008.02.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bernal SY, et al. Role of amygdala dopamine D1 and D2 receptors in the acquisition and expression of fructose-conditioned flavor preferences in rats. Behav. Brain Res. 2009;205:183–190. doi: 10.1016/j.bbr.2009.06.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dragunow M, Faull R. The use of c-fos as a metabolic marker in neuronal pathway tracing. J. Neurosci. Methods. 1989;29:261–265. doi: 10.1016/0165-0270(89)90150-7. [DOI] [PubMed] [Google Scholar]
- VanElzakker M, Fevurly RD, Breindel T, Spencer RL. Environmental novelty is associated with a selective increase in Fos expression in the output elements of the hippocampal formation and the perirhinal cortex. Learn. Mem. 2008;15:899–908. doi: 10.1101/lm.1196508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Norgren R, Hajnal A, Mungarndee SS. Gustatory reward and the nucleus accumbens. Physiol. Behav. 2006;89:531–535. doi: 10.1016/j.physbeh.2006.05.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park TH, Carr KD. Neuroanatomical patterns of fos-like immunoreactivity induced by a palatable meal and meal-paired environment in saline- and naltrexone-treated rats. Brain Res. 1998;805:169–180. doi: 10.1016/s0006-8993(98)00719-7. [DOI] [PubMed] [Google Scholar]
- Zhao XL, Yan JQ, Chen K, Yang XJ, Li JR, Zhang Y. Glutaminergic neurons expressing c-Fos in the brainstem and amygdala participate in signal transmission and integration of sweet taste. Nan.Fang Yi.Ke.Da.Xue.Xue.Bao. 2011;31:1138–1142. [PubMed] [Google Scholar]
- Mungarndee SS, Lundy RF, Norgren R. Expression of Fos during sham sucrose intake in rats with central gustatory lesions. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008;295:R751–R763. doi: 10.1152/ajpregu.90344.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Otsubo H, Kondoh T, Shibata M, Torii K, Ueta Y. Induction of Fos expression in the rat forebrain after intragastric administration of monosodium L-glutamate, glucose and NaCl. Neurosci. 2011;196:97–103. doi: 10.1016/j.neuroscience.2011.09.003. [DOI] [PubMed] [Google Scholar]
- Yamamoto T, Sako N, Sakai N, Iwafune A. Gustatory and visceral inputs to the amygdala of the rat: conditioned taste aversion and induction of c-fos-like immunoreactivity. Neurosci. Lett. 1997;226:127–130. doi: 10.1016/s0304-3940(97)00265-6. [DOI] [PubMed] [Google Scholar]
- Mitra A, Lenglos C, Martin J, Mbende N, Gagne A, Timofeeva E. Sucrose modifies c-fos mRNA expression in the brain of rats maintained on feeding schedules. Neurosci. 2011;192:459–474. doi: 10.1016/j.neuroscience.2011.06.033. [DOI] [PubMed] [Google Scholar]
- Pecoraro N, Dallman MF. c-Fos after incentive shifts: expectancy, incredulity, and recovery. Behav. Neurosci. 2005;119:366–387. doi: 10.1037/0735-7044.119.2.366. [DOI] [PubMed] [Google Scholar]
- Hamlin AS, Blatchford KE, McNally GP. Renewal of an extinguished instrumental response: Neural correlates and the role of D1 dopamine receptors. Neurosci. 2006;143:25–38. doi: 10.1016/j.neuroscience.2006.07.035. [DOI] [PubMed] [Google Scholar]
- Kerfoot EC, Agarwal I, Lee HJ, Holland PC. Control of appetitive and aversive taste-reactivity responses by an auditory conditioned stimulus in a devaluation task: A FOS and behavioral analysis. Learn. Mem. 2007;14:581–589. doi: 10.1101/lm.627007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang M, Kelley AE. Enhanced intake of high-fat food following striatal mu-opioid stimulation: microinjection mapping and fos expression. Neurosci. 2000;99:267–277. doi: 10.1016/s0306-4522(00)00198-6. [DOI] [PubMed] [Google Scholar]
- Teegarden SL, Scott AN, Bale TL. Early life exposure to a high fat diet promotes long-term changes in dietary preferences and central reward signaling. Neurosci. 2009;162:924–932. doi: 10.1016/j.neuroscience.2009.05.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Del Rio D, et al. Involvement of the dorsomedial prefrontal cortex in high-fat food conditioning in adolescent mice. Behav. Brain Res. 2015;283:227–232. doi: 10.1016/j.bbr.2015.01.039. [DOI] [PubMed] [Google Scholar]
- Knapska E, Radwanska K, Werka T, Kaczmarek L. Functional internal complexity of amygdala: focus on gene activity mapping after behavioral training and drugs of abuse. Physiol. Rev. 2007;87:1113–1173. doi: 10.1152/physrev.00037.2006. [DOI] [PubMed] [Google Scholar]
- Dela Cruz JAD, et al. c-Fos induction in mesotelencephalic dopamine pathway projection targets and dorsal striatum following oral intake of sugars and fats in rats. Brain Res. Bull. 2015;111:9–19. doi: 10.1016/j.brainresbull.2014.11.002. [DOI] [PubMed] [Google Scholar]
- Paxinos G, Watson C. The rat brain in stereotaxic coordinates. Elsevier; 2006. [DOI] [PubMed] [Google Scholar]
- Ranaldi R, et al. The effects of VTA NMDA receptor antagonism on reward-related learning and associated c-fos expression in forebrain. Behav. Brain Res. 2011;216:424–432. doi: 10.1016/j.bbr.2010.08.026. [DOI] [PubMed] [Google Scholar]