

Abstract
There is increasing evidence suggesting the important role of inflammation and, subsequently, macrophages in the development and progression of renal disease. Macrophages are heterogeneous cells that have been implicated in kidney injury. Macrophages may be classified into two different phenotypes: classically activated macrophages (M1 macrophages), that release pro-inflammatory cytokines and promote fibrosis; and alternatively activated macrophages (M2 macrophages) that are associated with immunoregulatory and tissue-remodeling functions. These macrophage phenotypes need to be discriminated and analyzed to determine their contribution to renal injury. However, there are scarce studies reporting consistent phenotypic and functional information about macrophage subtypes in inflammatory renal disease models, especially in rats. This fact may be related to the limited macrophage markers used in rats, contrary to mice. Therefore, novel strategies are necessary to quantify and characterize the renal content of these infiltrating cells in a reliable way. This manuscript details a protocol for kidney digestion and further phenotypic and quantitative analysis of macrophages from rat kidneys by flow cytometry. Briefly, kidneys were incubated with collagenase and total macrophages were identified according to the dual presence of CD45 (leukocytes common antigen) and CD68 (PAN macrophage marker) in live cells.This was followed by surface staining of CD86 (M1 marker) and CD163 (M2 marker). Rat peritoneal macrophages were used as positive control for macrophage marker detection by flow cytometry. Our protocol resulted in low cellular mortality and allowed characterization of different intracellular and surface protein markers, thus limiting the loss of cellular integrity observed in other protocols. Moreover, this procedure allows the use of macrophages for further techniques, including cell sorting and mRNA or protein expression studies, among others.
Keywords: Immunology, Issue 116, Macrophages, Inflammation, M1, M2, Phenotypes, Polarization, Kidney, Rat
Introduction
Renal disease is a global health problem, with increased prevalence, and associated with elevated morbidity and mortality1. One of the most important mechanisms involved in the progression and development of renal injury is inflammation, mainly triggered by macrophages. Macrophages play a pivotal role in many inflammatory diseases, including renal disorders2. Thus, an elevated presence of infiltrating macrophages has been reported in biopsies from patients with acute kidney injury (AKI) or chronic kidney disease (CKD)3,4. Recent studies suggest that the long-term outcome of renal disease could be controlled by macrophages5,6. In response to the local microenvironment, macrophages may differentiate into different phenotypes that play diverse biological functions7. Two well differentiated macrophage phenotypes have been established: classically activated macrophages (M1 macrophages) and alternatively activated macrophages (M2)8. M1 macrophages promote inflammation, whereas M2 macrophages have an anti-inflammatory role and are involved in tissue repair9. Therefore, a better knowledge of macrophage heterogeneity is necessary to understand their regulation and contribution to renal pathology and develop novel therapeutic approaches.
Both, murine and rats models have been widely used to understand the molecular and cellular mechanism involved in renal injury10. However, there are substantial differences in the diverse markers used to identify macrophages phenotypes between these rodents. Hence, several murine markers, such as F4/80 or Ly6C are not used in rats, thus limiting the extrapolation of findings between these species. Moreover, there is a limited number of markers describing macrophage phenotypes in rats, explaining the scarce studies analyzing macrophage heterogeneity in these animals as compared with mice. Therefore, new strategies for macrophage subset characterization are necessary to understand the role of macrophages in renal disease models in rats.
This manuscript describes a protocol for the phenotypic and quantitative analysis of macrophages from rat kidneys by flow cytometry. This technique can be further followed by several assays, including cell sorting and mRNA or protein expression studies to allow in-depth characterization of the role of macrophages in renal disease.
Protocol
This protocol was approved by local Institutional Animal Care and Use Committees following the Directive 2010/63/EU of the European Parliament and the National Guideline 53/2013.
1. Preparation of Reagents and Solutions
Prepare all the reagents and solutions under sterile conditions and use under a laminar flow hood. Keep solutions at 4 °C.
Prepare staining buffer (2% Fetal Bovine Serum (FBS) in 1x Dulbecco's PBS).
Prepare collagenase solution by adding 0.5 mg of collagenase to each ml of saline.
Prepare anesthesia solution of ketamine/xylazine (2:1 v/v).
2. Kidney Perfusion and Extraction
Anesthetize rats by intraperitoneal injection of ketamine/xylazine (75 mg/12 mg/kg weight). Carefully pinch a small fold of skin, to check that the animal is sufficiently anesthetized. Then, cover eyes with vet ointment to prevent dryness while under anesthesia. Note: 4-month-old Wistar rats are used in this assay.
Once the rat is totally anesthetized, place it on a surgical table in a supine position.
Apply 70% ethanol to the abdomen.
Make a central incision through the abdominal skin and peritoneum, from the pubis to the rib cage, to expose the pleural and abdominal cavities.
Inject saline solution (0.9%) into the abdominal aorta to perfuse kidneys using a perfusion system until all blood is removed from kidneys. Cut the aorta at the abdominal level to help release the blood.
Remove both kidneys from the rat by cutting from the renal hilum (renal vein, artery and ureter). To decapsulate the kidney, press the edge of the kidney using fingers, carefully separating the capsule11.
Place the kidney into Hanks' balanced salt solution.
3. Kidney Digestion and Cell Suspension
Cut half of a kidney with scissors into small pieces and put the pieces into a 1.5 ml tube. Note: All the following concentrations are calculated for 1 sample (1/2 rat's kidney).
Add 1 ml of the collagenase solution and incubate at 37 °C for 30 min. Mix by inversion every 5 min to make sure that the collagenase solution accesses the entire tissue.
Collect the solution and pass it through a strainer (40 µm) with the aid of a plunger, and resuspend it in 10 ml of staining buffer.
Centrifuge at 400 x g for 15 min.
Re-suspend the pellet in 1 ml ACK (Ammonium-Chloride-Potassium) Lysing Buffer. After 1 min 30 sec at room temperature, add 10 ml of staining buffer to stop the reaction.
Centrifuge at 400 x g for 10 min.
Re-suspend the pellet in an adequate volume of staining buffer (1 ml) and filter the cell suspension using a strainer (30 µm).
Count the number of cells using trypan blue exclusion on a hemocytometer and add 2 million cells to a 1.5 ml tube for staining.
4. Cell Staining and Flow Cytometry Analysis
Centrifuge cells at 100 x g for 5 min.
Re-suspend the pellet in 100 µl of rat serum (diluted 1:100) for 10 min at 4 °C to block Fc receptors.
Wash by adding 1 ml of staining buffer and centrifuge 100 x g for 5 min.
Mix the antibodies for cell surface staining in 100 µl of staining buffer for each condition: CD45 APC-Cy7 (1:100), CD163 A647(4:100) and solution to detect live cells (3:1,000).
Re-suspend the cell pellet in the antibody mix for 20 min at 4 °C in the dark.
Wash by adding 1 ml of staining buffer and centrifuge at 100 x g for 5 min. Repeat this step.
Add 600 µl of Fixation/Permeabilization solution for 20 min at 4 °C in the dark.
Wash 2 times with 1 ml Permeabilization/Wash buffer and centrifuge at 170 x g for 5 min.
Add the CD68 FITC antibody (35:1,000) for the intracellular staining to 100 µl of Permeabilization/Wash buffer for each condition.
Re-suspend the pellet in the CD68-antibody solution and incubate 50 min at 4 °C in the dark.
Wash with 1 ml Permeabilization/Wash buffer and centrifuge at 170 x g for 5 min.
Re-suspend the pellet in 100 µl of Permeabilization/Wash buffer, add CD86PE antibody (35:1,000) and incubate for 20 min at 4 °C in the dark.
Wash by adding 1 ml of Permeabilization/Wash buffer and centrifuge at 100 x g for 5 min.
Re-suspend the pellet in 100 µl of Permeabilization/Wash buffer and pass it to a flow cytometry tube.
Analyze samples by flow cytometry. Determine CD45 staining in live cells gated in the Side-scattered light/forward-scattered light (SSC/FSC) window. In a second step, analyze CD86 and CD163 expression in CD68+ cells.
Representative Results
We analyzed macrophage heterogeneity in an inflammatory experimental model of renal injury associated with increased presence of infiltrating macrophages in the kidney. In this model, renal damage was induced by administration of aldosterone (1 mg-1kg-1day) plus salt (NaCl 1%) in drinking water for 3 weeks in Wistar rats, as previously reported12.
The schedule of our experiments is shown in Figure 1. First, kidney cells were isolated using mechanical methods and further enzymatic digestion with collagenase (Figure 1A). Thereafter, erythrocytes were lysed and renal macrophages were detected by using flow cytometry. Cells were incubated with antibodies for CD45, CD86 and CD163 for membrane detection and CD68 for intracellular staining (Figure 1B). Finally, the phenotype of the macrophage populations was analyzed according to the scheme showed in Figure 1C.
Renal macrophage content was analyzed by selecting cells with dual positivity for CD45 and CD68 (Figure 2A). Using the protocol described above, an increase in CD45+/CD68+ macrophages was observed in aldosterone-treated rats compared to the control group (0.57 ± 0.16 vs. 0.25 ± 0.02, %CD45+/CD68+ macrophages per total renal cells) (Figure 2, see table insert). Cell viability of CD45+/CD68+ macrophages, as determined by flow cytometry staining with violet dye, was routinely greater than 95% (data not shown).Thereafter, CD86 (M1 marker) and CD163 (M2 marker) expression was assessed among the CD45+/CD68+ cells. Aldosterone administration increased the content of CD45+/CD68+/CD86+ M1 macrophages (2.87 ± 2.3 vs. 1.36 ± 0.68; %CD45+/CD68+/CD86+ macrophages per total CD45+/CD68+ cells) (Figure 2). However, a low number of CD45+/CD68+/CD163+ M2 macrophages were observed in both the aldosterone-treated and control groups. To analyze whether the low CD163 expression was related to proteolytic shedding during the experimental protocol, we repeated this procedure, including collagenase digestion, in macrophages isolated from rat peritoneum, as previously described 13. As reported in Figure 2B, CD45+/CD68+ peritoneal macrophages were positive for both CD86 and CD163, validating the utility of the selected antibodies and confirming the effectiveness of our protocol for the characterization of M1/M2 macrophages in rat kidney.
To validate these results, the inflammatory response associated with renal damage in the above experimental models was studied by immunohistochemistry and real-time PCR. As reported in Figure 3, increased CD68+ macrophages were observed in aldosterone + salt treated rats, as compared to control. Then, macrophage phenotype markers were determined to characterize renal M1/M2 macrophage distribution. Increased mRNA expression of iNOS and IFN-γ (M1 markers) was observed after aldosterone + salt treatment, whereas Arg1 or IL-10 mRNA expression (M2 macrophage markers) did not change after aldosterone administration. Together, these results confirm that aldosterone + salt administration mediates the inflammatory M1 macrophage phenotype in vivo.
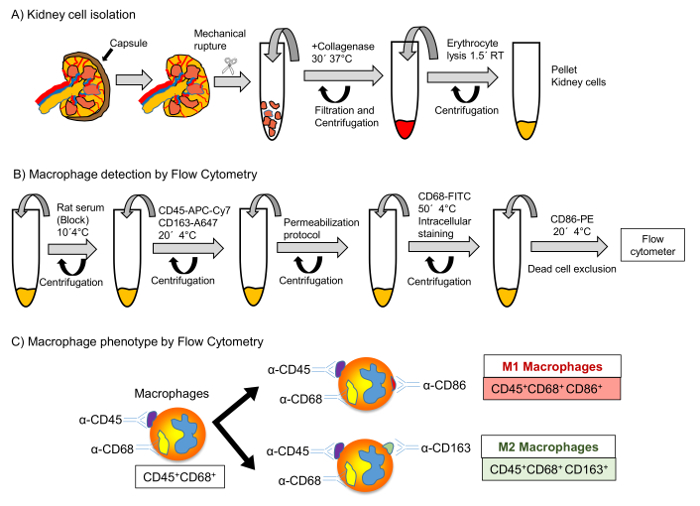 Figure 1: Protocol for Renal Cells Isolation and Macrophage Detection by Flow Cytometry. Schematic representation of renal cell isolation (A) and macrophage detection by flow cytometry (B). Representative scheme of the different macrophage phenotypes according to different distribution for CD45, CD68, CD86 and CD163 markers (C). Please click here to view a larger version of this figure.
Figure 1: Protocol for Renal Cells Isolation and Macrophage Detection by Flow Cytometry. Schematic representation of renal cell isolation (A) and macrophage detection by flow cytometry (B). Representative scheme of the different macrophage phenotypes according to different distribution for CD45, CD68, CD86 and CD163 markers (C). Please click here to view a larger version of this figure.
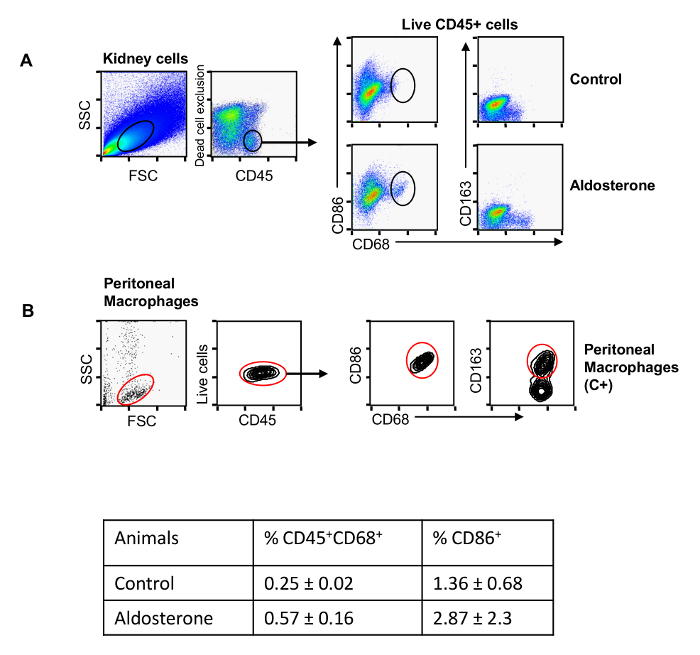 Figure 2: M1 and M2 Macrophage Markers in Kidneys from Aldosterone Treated Rats by Flow Cytometry. Representative dot-plots of CD45 staining in live cells gated in the Side-scattered light/forward-scattered light (SSC/FSC) window (A,
left) from kidney suspensions in control and aldosterone+salt-treated animals. The box inside the dot plots identifies live cells expressing CD45. Cells included in this box were analyzed to determine CD86 and CD163 expression in CD68+ cells (A, right). Dot plot for peritoneal macrophages as positive control for the detection of CD45, CD68, CD86 and CD163, applying the same strategy used previously. The data are presented as mean ± SEM (B). Please click here to view a larger version of this figure.
Figure 2: M1 and M2 Macrophage Markers in Kidneys from Aldosterone Treated Rats by Flow Cytometry. Representative dot-plots of CD45 staining in live cells gated in the Side-scattered light/forward-scattered light (SSC/FSC) window (A,
left) from kidney suspensions in control and aldosterone+salt-treated animals. The box inside the dot plots identifies live cells expressing CD45. Cells included in this box were analyzed to determine CD86 and CD163 expression in CD68+ cells (A, right). Dot plot for peritoneal macrophages as positive control for the detection of CD45, CD68, CD86 and CD163, applying the same strategy used previously. The data are presented as mean ± SEM (B). Please click here to view a larger version of this figure.
 Figure 3: Inflammatory Response after Aldosterone + Salt Administration. (A) Representative images of CD68 staining and (B) quantitative analyses of mRNA levels measured by RT-PCR for M1 markers (iNOS and INFγ) and M2 markers (Arg1 and IL-10) in control and aldosterone+salt-treated rats. Data are expressed as mean ± SEM. *p< 0.05 vs. control. Scale bar: 100 µm. Please click here to view a larger version of this figure.
Figure 3: Inflammatory Response after Aldosterone + Salt Administration. (A) Representative images of CD68 staining and (B) quantitative analyses of mRNA levels measured by RT-PCR for M1 markers (iNOS and INFγ) and M2 markers (Arg1 and IL-10) in control and aldosterone+salt-treated rats. Data are expressed as mean ± SEM. *p< 0.05 vs. control. Scale bar: 100 µm. Please click here to view a larger version of this figure.
Discussion
Macrophages are heterogeneous cells that play an important role in different inflammatory diseases, including renal disorders. There is increasing interest in the characterization of macrophages subsets in renal disease because each macrophage subpopulation contributes in a different way to the development of kidney injury, as reported in glomerulonephritis, diabetic nephropathy and kidney cancer14-16. In the early stages of acute renal injury, a predominance of M1 macrophages is observed, promoting tubular necrosis and inflammation. However, in later stages a higher content of M2 macrophages is observed to resolve inflammation and participate in tissue remodeling. In chronic kidney disease, both M1 and M2 macrophages co-exist simultaneously, although M1 may be predominant, thus increasing and perpetuating inflammatory renal damage. Therefore, is important to increase our knowledge of the pathophysiological role of macrophage subtypes in renal disease. Immunohistochemical studies cannot determine the percentage of macrophage subsets in a robust way17. Therefore, it is necessary to develop new techniques to quantitatively determine the different macrophage phenotypes and to understand the role of these cells on renal inflammatory response.
This manuscript describes a protocol to determine the number and phenotype of renal macrophages subsets by flow cytometry in rats. In our study, rats were subjected to an inflammatory model of renal damage by administration of aldosterone. In this model, we observed increased CD45+ and C68+ macrophage infiltration as determined by flow cytometry and confirmed by immunohistochemistry. Furthermore, an enrichment of M1 macrophages (CD68+/CD86+), but not M2 macrophage, were observed in aldosterone-treated mice. These results were further confirmed by determining mRNA expression of M1 (iNOS and IFN-γ) and M2 macrophage markers (Arg1 or IL-10). Flow cytometry analysis reported low cell mortality during the isolation protocol. There are several factors that may affect cell viability and surface markers detection, such as the level of tissue dissociation as well as duration and type of tissue digestion (enzymatic, mechanic, etc.). Excessive digestion increases cell mortality and macrophage activation and decreases surface marker expression, whereas the opposite effect may result from low kidney dissociation. However, when our protocol was conducted in macrophages isolated from rat peritoneum, an enhanced presence of CD86 and CD163 was observed in these cells.
Our protocol has been optimized to determine macrophage heterogeneity in kidneys from rats with aldosterone-mediated hypertension, and therefore, may be extrapolated to other inflammatory renal disease models. However, this procedure may be adapted to each experimental condition because inflammation modifies tissue integrity, thus affecting the rate of tissue digestion by collagenase.
It is important to note that for determination of total macrophage content, cells were fixed and permeabilized to allow intracellular labeling with CD68 antibody18. The protocol for fixation included the use of formaldehyde that may affect fluorochromes conjugated to primary antibodies, especially phycoerythrin. To resolve this issue, incubation with CD86-PE antibody was performed after addition of the Fixation/Permeabilization solution and further CD68-staining. On the other hand, it is important to note that formaldehyde may also interfere with other assays that may be performed after macrophages isolation, such as mRNA expression assays. However, there are commercial kits available to isolate mRNA from formaldehyde fixed cells.
In summary, this protocol describes a method for phenotypic characterization of renal macrophages in a quantitative manner using flow cytometry. Overall, the protocol described here is easy to use, reproducible, and useful to discriminate different macrophage populations from rat kidneys.
Disclosures
The authors have nothing to disclose.
Acknowledgments
This work was supported by grants from FIS/FEDER (Programa Miguel Servet: CP10/00479, PI13/00802 and PI14/00883), Spanish Society of Atherosclerosis, Spanish Society of Nephrology and Fundaciòn Renal Iñigo Alvarez de Toledo (FRIAT) to Juan Antonio Moreno. FIS/FEDER funds PI14/00386 and Instituto Reina Sofìa de Investigaciòn Nefrològica to Jesús Egido. Fundaciòn Conchita Rabago to Melania Guerrero Hue. Fundaciòn Renal Iñigo Alvarez de Toledo (FRIAT) to Alfonso Rubio Navarro.
References
- Gansevoort RT, et al. Chronic kidney disease and cardiovascular risk: epidemiology, mechanisms, and prevention. Lancet. 2013;382:339–352. doi: 10.1016/S0140-6736(13)60595-4. [DOI] [PubMed] [Google Scholar]
- Kon V, Linton MF, Fazio S. Atherosclerosis in chronic kidney disease: the role of macrophages. Nat. Rev. Nephrol. 2011;7:45–54. doi: 10.1038/nrneph.2010.157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim JH, et al. Macrophage depletion ameliorates glycerol-induced acute kidney injury in mice. Nephron Exp. Nephrol. 2014;128:21–29. doi: 10.1159/000365851. [DOI] [PubMed] [Google Scholar]
- Belliere J, et al. Specific macrophage subtypes influence the progression of rhabdomyolysis-induced kidney injury. J. Am. Soc. Nephrol. 2015;26:1363–1377. doi: 10.1681/ASN.2014040320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kinsey GR. Macrophage dynamics in AKI to CKD progression. J. Am. Soc. Nephrol. 2014;25:209–211. doi: 10.1681/ASN.2013101110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lech M, et al. Macrophage phenotype controls long-term AKI outcomes--kidney regeneration versus atrophy. J. Am. Soc. Nephrol. 2014;25:292–304. doi: 10.1681/ASN.2013020152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murray PJ, et al. Macrophage activation and polarization: nomenclature and experimental guidelines. Immunity. 2014;41:14–20. doi: 10.1016/j.immuni.2014.06.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gordon S. Alternative activation of macrophages. Nat. Rev. Immunol. 2003;3:23–35. doi: 10.1038/nri978. [DOI] [PubMed] [Google Scholar]
- Mosser DM, Edwards JP. Exploring the full spectrum of macrophage activation. Nat. Rev. Immunol. 2008;8:958–969. doi: 10.1038/nri2448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ortiz A, et al. Translational value of animal models of kidney failure. Eur. J. Pharmacol. 2015;759:205–220. doi: 10.1016/j.ejphar.2015.03.026. [DOI] [PubMed] [Google Scholar]
- Martina MN, Bandapalle S, Rabb H, Hamad AR. Isolation of double negative alphabeta T cells from the. J. Vis. Exp. 2014. [DOI] [PMC free article] [PubMed]
- Martin-Fernandez B, et al. Aldosterone Induces Renal Fibrosis and Inflammatory M1-Macrophage Subtype via Mineralocorticoid Receptor in Rats. PLoS. One. 2016;11:e0145946. doi: 10.1371/journal.pone.0145946. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Layoun A, Samba M, Santos MM. Isolation of murine peritoneal macrophages to carry out gene expression analysis upon Toll-like receptors stimulation. J. Vis. Exp. 2015. [DOI] [PMC free article] [PubMed]
- Komohara Y, et al. Macrophage infiltration and its prognostic relevance in clear cell renal cell carcinoma. Cancer Sci. 2011;102:1424–1431. doi: 10.1111/j.1349-7006.2011.01945.x. [DOI] [PubMed] [Google Scholar]
- Han Y, Ma FY, Tesch GH, Manthey CL, Nikolic-Paterson DJ. Role of macrophages in the fibrotic phase of rat crescentic glomerulonephritis. Am. J. Physiol Renal Physiol. 2013;304:F1043–F1053. doi: 10.1152/ajprenal.00389.2012. [DOI] [PubMed] [Google Scholar]
- Ndisang JF. Role of the heme oxygenase-adiponectin-atrial natriuretic peptide axis in renal function. Curr. Pharm. Des. 2015;21:4380–4391. doi: 10.2174/1381612821666150803145508. [DOI] [PubMed] [Google Scholar]
- Blackbeard J, et al. Quantification of the rat spinal microglial response to peripheral nerve injury as revealed by immunohistochemical image analysis and flow cytometry. J. Neurosci. Methods. 2007;164:207–217. doi: 10.1016/j.jneumeth.2007.04.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strobl H, Scheinecker C, Csmarits B, Majdic O, Knapp W. Flow cytometric analysis of intracellular CD68 molecule expression in normal and malignant haemopoiesis. Br. J. Haematol. 1995;90:774–782. doi: 10.1111/j.1365-2141.1995.tb05195.x. [DOI] [PubMed] [Google Scholar]