

Abstract
Microglia are the tissue resident macrophages of the central nervous system (CNS) and they perform a variety of functions that support CNS homeostasis, including phagocytosis of damaged synapses or cells, debris, and/or invading pathogens. Impaired phagocytic function has been implicated in the pathogenesis of diseases such as Alzheimer's and age-related macular degeneration, where amyloid-β plaque and drusen accumulate, respectively. Despite its importance, microglial phagocytosis has been challenging to assess in vivo. Here, we describe a simple, yet robust, technique for precisely monitoring and quantifying the in vivo phagocytic potential of retinal microglia. Previous methods have relied on immunohistochemical staining and imaging techniques. Our method uses flow cytometry to measure microglial uptake of fluorescently labeled particles after intravitreal delivery to the eye in live rodents. This method replaces conventional practices that involve laborious tissue sectioning, immunostaining, and imaging, allowing for more precise quantification of microglia phagocytic function in just under six hours. This procedure can also be adapted to test how various compounds alter microglial phagocytosis in physiological settings. While this technique was developed in the eye, its use is not limited to vision research.
Keywords: Immunology, Issue 116, phagocytosis, microglia, macrophage, retina, in vivo, flow cytometry, myeloid cells
Introduction
The overall goal of this method is to accurately assess and quantify in vivo microglial phagocytosis. Microglia are the tissue resident macrophages of the central nervous system (CNS). They perform a variety of functions to ensure maintenance of tissue homeostasis. These include immune surveillance, secretion of neurotrophic factors and, of pivotal importance, phagocytosis1. Microglial phagocytosis is key in several important events during development of the brain and retina, such as phagocytosis of irrelevant synapses (synaptic pruning) and removal of apoptotic neurons2-4. Furthermore, microglial phagocytosis of damaged or apoptotic neurons, cellular debris, and invading microbes has been shown to be essential for maintaining CNS homeostasis through adulthood5. Finally, microglial phagocytosis has been implicated in the pathogenesis of several neurodegenerative diseases, including Alzheimer's disease and age-related macular degeneration, where it has been suggested that defective or insufficient phagocytic capacity may contribute to the build-up of amyloid-β (Aβ) plaques and drusen, respectively6,7.
Microglial function is tightly regulated by their microenvironment, notably by soluble factors such as tumor-growth factor β or cell-cell interactions. Neurons constitutively express several cell surface ligands, such as CD200 and CX3CL1, while microglia exclusively express the respective receptors CD200R and CX3CR1. These receptors contain immunoreceptor tyrosine-based inhibition motifs (ITIMs) in their intracellular portion. These inhibitor receptors are critical for preventing the over-stimulation of microglia, which can contribute to neuroinflammation. Thus, under normal physiological conditions, cell-cell interactions between neurons and microglia keep microglia in a quiescent state. During tissue injury, however, neurons can down-regulate expression of these ligands, removing their inhibitory effect on microglia activation. Microglial function (including phagocytosis) is thus tightly linked to their microenvironment8. Nevertheless, to date, there are no standardized assays to study microglia phagocytosis in a physiological context or in a way that fully replicates their CNS microenvironment.
Several assays have been developed to measure phagocytic activity of microglia in vitro, where primary microglia or microglia cell lines are cultured with target cells (e.g., apoptotic neurons) or fluorescently labeled beads. Target uptake is then assessed using fluorescent imaging microscopy or flow cytometry9-12. These assays allow testing of how pharmacological or genetic manipulation may affect microglial phagocytosis and, while informative, fail to fully replicate the complex in vivo environment. Indirect methods for examining microglial phagocytosis in vivo have been reported: these are accomplished by staining of molecules thought to be involved in phagocytosis (e.g., CD68), assessing physical proximity of microglia and targets for phagocytosis (e.g., compromised neurons or synaptic elements), or by immunohistochemical detection of phagocytic targets within microglial cells (e.g., Aβ)13-17. Two studies have used more direct approaches to assess microglia phagocytosis in vivo. Hughes and colleagues have used imaging techniques to measure microglial uptake of beads delivered via the intracranial route18. Sierra et al. developed a refined method to quantitatively assess microglia phagocytosis of apoptotic cells using complex imaging techniques4. However, these methods involve complicated protocols for tissue preparation, sectioning, imaging, and analysis. We have previously used flow cytometric analysis to assess phagocytosis of photoreceptor outer segments by retinal pigmented epithelium (RPE) cells in culture19. Here, we describe a protocol to rapidly assess uptake of fluorescently labeled particles by retinal microglia as a quantitative measure of in vivo microglia phagocytosis.
The protocol we describe here allows for reliable and quantitative measurement of retinal microglial phagocytosis in just under six hours in three critical steps: (1) intravitreal delivery of fluorescently labeled particles, (2) harvest and preparation of retinal tissue, and (3) flow cytometric analysis. The method we have developed is a robust method to assess microglial phagocytosis in the retina, and it can be successfully used to test how various compounds or genetic manipulation alter this key microglial function in physiological settings. As a specialized area of the CNS, the retina is an easily accessible model system to study microglia function20. While this method was developed in the eye, we believe it can be useful for all neuroscientists investigating microglia phagocytic function.
Protocol
All animals were treated in accordance with the ethical guidelines established by the Scripps Research Institute.
1. Preparation of Materials for Injection
Sterilize a 33 G needle and syringe: disassemble and autoclave at 115 οC. Prepare needles for injection by rising in sterile phosphate buffered saline (PBS).
Thaw fluorescently labeled particles at room temperature for 5 - 10 min. Prepare particle solution for injection by reconstituting to a 50 mg/ml solution in sterile PBS with Ca2+/Mg2+. NOTE: AF488-labeled particles of fungal origin that have been validated for phagocytosis assays are used in this protocol21-23. For optimal uptake, prepare particles fresh and immediately before injection. Note 2: Particle concentrations may be optimized for each specific experiment.
2. Intravitreal Injection of Bead Solution
NOTE: Two people are required to perform the injection, in a way such that the person performing the injection can hold the mouse and maintain the focus on the eyeball, while the other person passes the loaded syringe and pushes the plunger.
Anaesthetize the rodent by intraperitoneal injection of 100 mg/ml ketamine and 10 mg/ml xylazine at a dose of 20 µl/10 g of body weight. Before injection, use toe pinch to assess level of anesthesia. NOTE: Isoflurane has a profound effect on myeloid cell function, thus, its use throughout this assay should be avoided24,25.
Load needle with 0.5 µl of fluorescently labeled particle solution.
- Lay the mouse sideways under a surgical microscope (Figure 1A-B). Use a soft material, e.g., a gel pack, for better positioning of the mouse. For young mice in which the eyes are not yet open, gently open the eyelids with the help of fine 45o angled forceps by creating a fissure along the slit from which the eyelids will eventually open.
- With the help of fine 45o angled forceps carefully apply pressure around the eyelid, so that the eyeball pops slightly out of the socket. Hold the head with two fingers just above the ear and by its jaw and gently stretch the skin in parallel to the eyelids to keep the eye slightly out of the socket. Be careful not to grasp too close to the throat (Figure 1B).
To puncture the eyeball, insert the syringe's needle in the corneal limbus (where the cornea and sclera connect). This is visible as a gray circle in pigmented mice. Retract the needle slightly to expel a small volume of vitreous fluid and then inject. The person performing the injection should gently hold the mouse with one hand and the needle with the other; the second person should slowly push the plunger.
Retract the syringe slowly. Apply eye moisturizing drops to keep the eye hydrated.
Let the rodent recover in a cage over a heat pad and continue to monitor the animal. If working with pups, do not return the animal to a cage with other alert animals until it is breathing and capable of spontaneous movement. If working with adults, do not return the rodent to a cage with other alert animals until it regains sternal recumbence.
3. Harvesting of Retinal Tissue
NOTE: Retinal tissue from eyes not injected with fluorescently labeled particles should be collected as a control for flow cytometric analysis. Though the assay can be performed using a single retina, for best performance, two retinas should be pooled together.
Collect retinal tissue 3 hr after intravitreal injection of fluorescently labeled particles. NOTE: While time after injection of the particle solution may be optimized for each specific experiment, we found that 3 hr after injection, particle uptake could be seen throughout most layers of the retina (Figure 1C-D).
Sacrifice mice by cervical dislocation.
Collect the eyeballs by gently pressing against the eyelid with the help of fine 45o angled forceps to proptose the eyeball. Position the forceps behind the eyeball and pull.
Dissect the retina under a dissecting microscope. Transfer the eyeball to a Petri dish containing a small amount of PBS with Ca2+/Mg2+. In a dry area of the Petri dish, perforate the eyeball in the corneal limbus with the tip of superfine forceps.
Hold the eyeball with the help of fine 45o angled forceps and use spring scissors to cut around the corneal limbus, until roughly half of the corneal limbus circumference is cut.
Hold the eyeball with the fine 45o angled forceps and bring the eyeball into PBS. With a second pair of fine 45o angled forceps tear the cornea and sclera apart. The lens and retina will come out intact.
Separate the lens and retina. Collect the retina and transfer to a 5.4 ml polystyrene test tube containing 2 ml of PBS with Ca2+/Mg2+.
4. Preparing a Single Cell Suspension
Prepare single cell suspensions using a neuronal tissue dissociation kit with minor modifications to the manufacturer's instructions. Briefly, triturate retinas by pipetting up and down with a P1000 pipette and performing an enzymatic digestion at 37 oC without shaking.
5. Staining Single Cell Suspensions for Flow Cytometric Analysis
Resuspend cells in 200 µl of staining buffer (Dulbecco's phosphate-buffered saline with 0.2% bovine serum albumin (BSA) and 0.09% sodium azide) and transfer to a U-bottom 96-well plate. Centrifuge for 5 min at 130 x g. NOTE: Sodium azide is harmful to humans and the environment. Use appropriate personal protective equipment and discard waste in accordance with local regulations.
Invert the plate over a sink to discard supernatant. To block Fc receptors, resuspend cells in 25 µl of stain buffer containing 5 µg/ml of anti-mouse CD16/CD32 antibody per well. Incubate for 5 min at room temperature.
Add 25 µl of staining buffer containing 0.5 µg/ml of anti-mouse Ly6C-APC-Cy7, 0.5 µg/ml of anti-mouse Ly6G-Pe-Cy7 and 2.5 µg/ml of anti-mouse CD11b-AF650. Incubate for 15 min at room temperature in the dark.
Centrifuge for 5 min at 130 x g. Invert the plate to discard supernatant. Wash by resuspending cells in 200 µl of staining buffer.
Centrifuge for 5 min at 130 x g. Resuspend in 200 µl of stain buffer containing 0.5 µg/ml of propidium iodide (PI). Transfer to 1.2 ml microtiter tubes.
Wash wells with an additional 100 µl of staining buffer containing 0.5 µg/ml of PI and pool with the previous 200 µl in the 1.2 ml microtiter tubes. A total volume of 300 µl of stained cells is obtained.
6. Flow Cytometric Analysis
Using a conventional three laser (violet, blue, and red laser), avoid PE, PerCP-Cy5.5, BV510, and PI dyes due to significant spillover from the extremely bright fluorescent AF488-particles. Use a fourth (yellow) laser to optimize this assay, as it allows for a PE-labeled antibodies and PI (in the mCherry channel instead of the PerCP-Cy5.5 channel) to be used (Figure 2). NOTE: If a yellow laser is not available, use a dead cell exclusion dye in another channel.
Gate on PI negative cells (PI-) to exclude dead cells (Figure 2B).
After excluding dead cells, gate on CD11b positive cells (CD11b+), this will include all myeloid cells (Figure 2C).
Within the CD11b+ population, gate on Ly6C-/Ly6G- to exclude neutrophils (CD11b+/Ly6G+) and monocytes (CD11b+/Ly6C+; Figure 2D). After gating on microglia (here defined as CD11b+/Ly6C-/Ly6G- for simplicity), two clear populations should be visible; one negative and one positive for the fluorescently labeled particles. Phagocytic cells will have taken up particles and are therefore AF488+ (Figure 2E). NOTE: Ly6C positive or Ly6G positive populations can also be analyzed to measure particle uptake using this staining approach. The percentage of phagocytic CD11b+ cells correlates well with the percentage of phagocytic microglia (Figure 2F). For researchers without flow cytometry experience, a simpler analysis with CD11b staining alone can be performed, albeit this will include phagocytic neutrophils and monocytes, as well as microglia. Within this CD11b+/Ly6C-/Ly6G- population, these cells are > 99% microglia as judged by staining with the markers F4/80 and CX3CR1; these markers can be added but are not necessary in our hands.
Representative Results
Here we describe a method to rapidly and reliably quantify the number of phagocytic retinal microglia in a physiological setting using flow cytometric analysis (Figure 2). This method can be adapted to test the effect of compounds and/or genetic manipulation on the phagocytic capacity of microglia (Figures 3A, 3B). It can also be used in young (10 - 20 days postnatal) or adult mice (Figure 3C). Varying doses of lipopolysaccharide (LPS) were administered intraperitoneally. 24 hr after LPS challenge, the protocol described here was used to assess retinal microglia phagocytic function (Figure 3A). A dose of 1.42 mg/kg of LPS induced a statistically significant increase in the percentage of phagocytic microglia when compared to vehicle control (Figure 3B), as would be expected26.
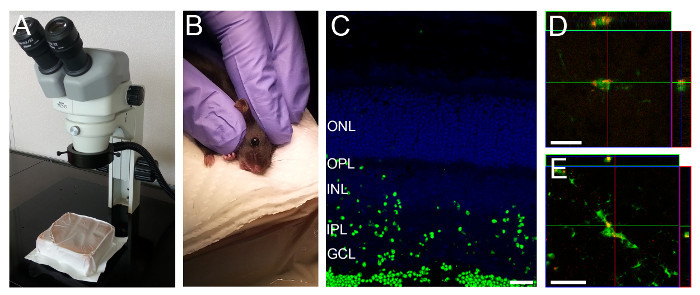 Figure 1:Intravitreal Injection Setup and Representative Retinal Section showing Fluorescently Labeled Particles in Retinal Layers. (A) A gel pack is positioned under the dissection microscope. (B) A rodent is held as shown to position the eye for intravitreal injection. (C) Three hours after injection fluorescently labeled particles can be seen throughout most retinal layers. (D,E) CX3CR1GFP/GFP mice and particles labeled with a red fluorophore were used to visualize particle uptake by microglia. Microglia in the deeper (D) and superficial (E) layers take up particles. Scale bars - 20 µm. Please click here to view a larger version of this figure.
Figure 1:Intravitreal Injection Setup and Representative Retinal Section showing Fluorescently Labeled Particles in Retinal Layers. (A) A gel pack is positioned under the dissection microscope. (B) A rodent is held as shown to position the eye for intravitreal injection. (C) Three hours after injection fluorescently labeled particles can be seen throughout most retinal layers. (D,E) CX3CR1GFP/GFP mice and particles labeled with a red fluorophore were used to visualize particle uptake by microglia. Microglia in the deeper (D) and superficial (E) layers take up particles. Scale bars - 20 µm. Please click here to view a larger version of this figure.
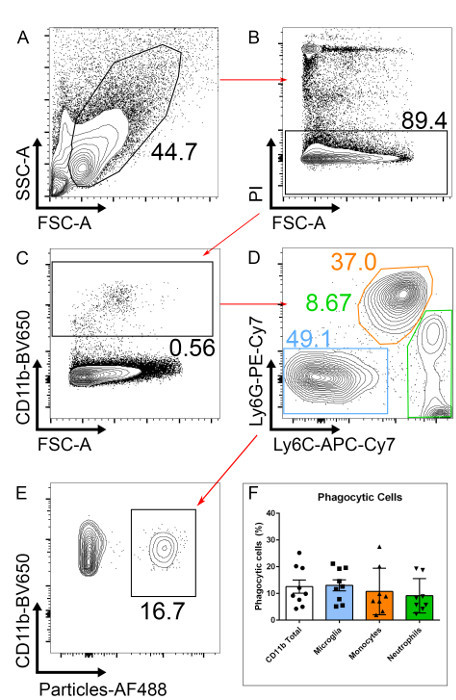 Figure 2: Gating Strategy to Select Phagocytic Microglia.(A) Single cells suspensions from retinal tissue from a p10 mouse are analyzed. (B) After excluding dead cells (PI+), (C) CD11b+ cells are selected. (D) To select only for microglia, CD11b+ neutrophils (Ly6G+) and monocytes (Ly6C+) are excluded. (E) Phagocytic microglia will have taken up fluorescently labeled particles. (F) The number of phagocytic CD11b+ cells is similar to that of microglia (CD11b+Ly6G-Ly6C-). n= 9, pooled from three independent experiments; error bars represent mean ± SEM. Please click here to view a larger version of this figure.
Figure 2: Gating Strategy to Select Phagocytic Microglia.(A) Single cells suspensions from retinal tissue from a p10 mouse are analyzed. (B) After excluding dead cells (PI+), (C) CD11b+ cells are selected. (D) To select only for microglia, CD11b+ neutrophils (Ly6G+) and monocytes (Ly6C+) are excluded. (E) Phagocytic microglia will have taken up fluorescently labeled particles. (F) The number of phagocytic CD11b+ cells is similar to that of microglia (CD11b+Ly6G-Ly6C-). n= 9, pooled from three independent experiments; error bars represent mean ± SEM. Please click here to view a larger version of this figure.
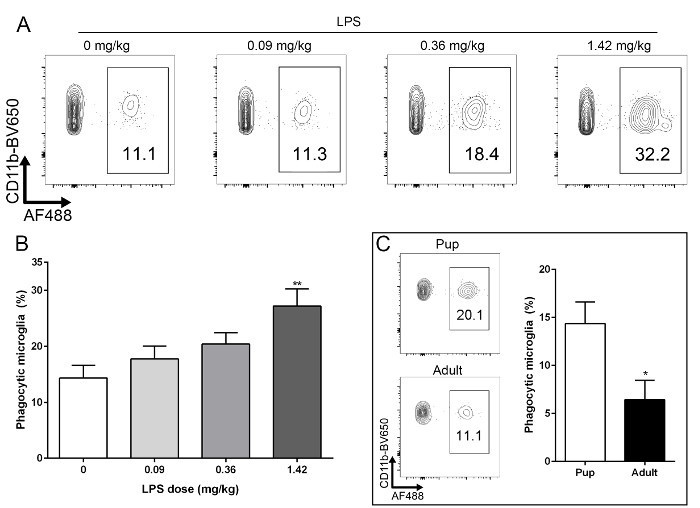 Figure 3:Microglia Phagocytic Function can be Assessed after LPS Challenge and in Pups and Adult Mice.(A) Young mice (p10) were challenged with intraperitoneal LPS at varying doses, 24 hr before assessing phagocytosis. (B) Challenge with 1.42 mg/kg resulted in a significantly increased percentage of phagocytic retinal microglia when compared to vehicle controls (p= 0.0021). (C) This protocol can be used to measure retinal microglial phagocytic function in pup and adult mice alike. There is a significantly smaller percentage of phagocytic microglia in adult mice when compared to young (p10) mice (p= 0.019) not challenged with LPS. n= 9 -12, pooled from three to four independent experiment; error bars represent mean ± SEM. Please click here to view a larger version of this figure.
Figure 3:Microglia Phagocytic Function can be Assessed after LPS Challenge and in Pups and Adult Mice.(A) Young mice (p10) were challenged with intraperitoneal LPS at varying doses, 24 hr before assessing phagocytosis. (B) Challenge with 1.42 mg/kg resulted in a significantly increased percentage of phagocytic retinal microglia when compared to vehicle controls (p= 0.0021). (C) This protocol can be used to measure retinal microglial phagocytic function in pup and adult mice alike. There is a significantly smaller percentage of phagocytic microglia in adult mice when compared to young (p10) mice (p= 0.019) not challenged with LPS. n= 9 -12, pooled from three to four independent experiment; error bars represent mean ± SEM. Please click here to view a larger version of this figure.
Discussion
There are three critical steps in this method: (1) intravitreal injection of fluorescently labeled particles; (2) harvesting and preparation of retinal tissue; and (3) flow cytometric analysis. We recommend that researchers practice intravitreal injections prior to performing the method we present here. Albino mice (e.g., BALB/c) and a colored solution (e.g., fluorescently labeled particles) can be used for easy visualization of the needle and injected solution. Intravitreal injections are challenging and if not performed correctly will lead to biased and variable results. Common problems associated with poor injection technique are perforation of the lens and/or damage to the retina, which may result in bleeding and inflammation. To minimize the risk of trauma to the eye, the rodent should be in a stable position with minimal head movement. To penetrate the eyeball, the syringe should be inserted slowly and not pushed too far to avoid hitting the lens. Another common complication is reflux of the injected material out of the eye. To avoid this, the plunger should be depressed slowly and, after injection, the eyeball should be allowed to retract into the eye socket and the syringe should be retracted slowly. The best intravitreal injections activate microglia at low levels, but clear differences between untreated and treated groups can be measured. It is essential that appropriate controls (e.g., non-challenged mice) are used in each experiment to establish baseline phagocytic levels. Another aspect of this method that requires practice is the retinal dissection. To ensure comparable single cell preparations across specimens, retinal tissue must be collected swiftly and with minimal possible tissue disruption. Performing the last steps of dissection in PBS (see 3.4 - 3.7) will facilitate collection. Finally, it is important to prepare the retinal tissue for flow cytometry correctly. Activated microglia upregulate Fc receptors and thus nonspecific binding to the Fc portion of antibodies may occur7. An Fc block must be performed to prevent nonspecific staining. Selection of appropriate fluorophores and compensation set up is also important27.
We routinely use mice aged 10 to 20 postnatal days for these experiments. However, this protocol can also be used to assess microglial phagocytic function in adult mice (Figure 3C). Of note, due to presence of the hyaloid vasculature, populations of exogenous myeloid cells (Ly6C+ or Ly6G+) are more prominent in young mice (approximately 40 - 50% of all CD11b+ cells) than in adults (approximately 5% of all CD11b+ cells). Researchers should determine the age most appropriate for their experiments. The protocol presented here allows for robust detection of the levels of phagocytic microglia in the retina. However, we recommend that three to four biological replicates be used in each experiment and that at least three independent experiments are performed. While the particles used in this protocol have been validated for phagocytic assays21-23, if additional validation is required, immunohistochemical co-localization of the particles with specific phagocytic markers could be performed.
One potential limitation of this method is differential access of microglia to the particles. In the normal retina, microglia reside in two layers, the deeper outer plexiform layer, and the more superficial inner plexiform layer, which is closer to the vitreous7. Upon stimulation, or during aging, microglia can migrate throughout all retinal layers. In this method, fluorescently labeled particles are injected into the vitreous. Three hours after injection this results in particle accumulation throughout the retina and particularly along the vitreal surface. Researchers may further optimize the concentration of particles or time after injection for their experiments. It is likely that microglia in closer proximity to the vitreous will have easier access to the particles. Nevertheless, we show that the particles spread across the retina and are taken up by microglia across all layers but not in retinal pigmented epithelium (RPE) cells, probably due to limited bead access (Figure 1C-E). We have yet to determine whether subretinal microglia (which can accumulate with age) would gain access to the particles. If this is important to the researcher, this protocol could be performed after careful subretinal injection of the particles. Another important consideration is that there may be inherently different behaviors between retinal microglia and microglia in other regions of the CNS. It is well known that specific brain regions have different susceptibilities to disease. For example, in Parkinson's disease, neurons in the substantia nigra are mostly affected, while in Alzheimer's disease, the hippocampal neurons are mostly affected28. Thus, when studying the pathology of a specific disease, brain region is likely to be an important factor. A similar protocol to the one described here could be performed after intracranial injection to explore potential differences in the phagocytic response between retinal and brain microglia. The retina has many common features with other regions of the brain, including the presence of a blood-retina-barrier20. Retina microglia and brain microglia originate from the same yolk-sac progenitors and appear very similar with regard to morphology and cell-surface marker expression7,29. Furthermore, several brain diseases (e.g., Alzheimer's disease, stroke, and multiple sclerosis) have ocular manifestations, indicating that the retina is also affected20. While researchers should be aware of potential regional differences, we believe this is an informative model for all microglia research, and suggest that all neuroscientists studying microglial phagocytosis could use this protocol.
A distinct advantage to performing this assay in the eye is that multiple techniques have been developed to induce diverse neurogenic stresses in the retina, and the stress responses can be monitored in vivo and quantified using standardized protocols30. By inducing neurogenic stress and then performing the assay described here, researchers can analyze microglia function across a broad spectrum of insults that vary in severity. Finally, when combined with flow cytometry sorting techniques, this protocol has the advantage over immunohistochemistry that it may allow for in depth analysis of phagocytic microglia (e.g., qPCR, cell-culture, proteomics).
Uptake of particles has previously been used to measure phagocytic function, both in in vitro and in vivo systems. The latter involved intracranial injection of particles, harvesting and freezing of brain tissue, cryosectioning, immunostaining, imaging and post-imaging analysis18. The protocol presented here allows for much faster quantification of retinal microglia phagocytic function. It has the advantage over in vitro systems that the highly sensitive microglia are not removed from their microenvironment, allowing for assessment of phagocytic function in a physiological context. Specifically, it should enable the testing of how defects in other cell types (e.g., genetic ablation of genes in neurons) affect microglia phagocytic function. It also has the advantage over previous in vivo assays that it uses flow cytometry, allowing for fast, quantitative, and accurate detection of phagocytic cell populations.
Disclosures
The authors have nothing to disclose.
Acknowledgments
Salome Murinello is supported by American Diabetes Association grant #1-16-PDF-072. This work was supported by grants to Martin Friedlander from the National Institutes of Health (National Eye Institute EY11254 and EY22025) and the Lowy Medical Research Institute.
References
- Gomez-Nicola D, Perry VH. Microglial dynamics and role in the healthy and diseased brain: a paradigm of functional plasticity. Neuroscientist. 2015;21:169–184. doi: 10.1177/1073858414530512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tremblay ME, Lowery RL, Majewska AK. Microglial interactions with synapses are modulated by visual experience. PLoS Biol. 2010;8:e1000527. doi: 10.1371/journal.pbio.1000527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schafer DP, et al. Microglia sculpt postnatal neural circuits in an activity and complement-dependent manner. Neuron. 2012;74:691–705. doi: 10.1016/j.neuron.2012.03.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sierra A, et al. Microglia shape adult hippocampal neurogenesis through apoptosis-coupled phagocytosis. Cell Stem Cell. 2010;7:483–495. doi: 10.1016/j.stem.2010.08.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sierra A, Abiega O, Shahraz A, Neumann H. Janus-faced microglia: beneficial and detrimental consequences of microglial phagocytosis. Front Cell Neurosci. 2013;7(6) doi: 10.3389/fncel.2013.00006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chan G, et al. CD33 modulates TREM2: convergence of Alzheimer loci. Nat Neurosci. 2015;18:1556–1558. doi: 10.1038/nn.4126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murinello S, Mullins RF, Lotery AJ, Perry VH, Teeling JL. Fcgamma receptor upregulation is associated with immune complex inflammation in the mouse retina and early age-related macular degeneration. Invest Ophthalmol Vis Sci. 2014;55:247–258. doi: 10.1167/iovs.13-11821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ransohoff RM, Perry VH. Microglial physiology: unique stimuli, specialized responses. Annu Rev Immunol. 2009;27:119–145. doi: 10.1146/annurev.immunol.021908.132528. [DOI] [PubMed] [Google Scholar]
- Derecki N, Cronk J, Kipnis J. Assay of phagocytic function in primary murine microglia. Protoc Exchange. 2012.
- Fricker M, Oliva-Martin MJ, Brown GC. Primary phagocytosis of viable neurons by microglia activated with LPS or Abeta is dependent on calreticulin/LRP phagocytic signalling. J Neuroinflammation. 2012;9:196. doi: 10.1186/1742-2094-9-196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koenigsknecht-Talboo J, Landreth GE. Microglial phagocytosis induced by fibrillar beta-amyloid and IgGs are differentially regulated by proinflammatory cytokines. J Neurosci. 2005;25:8240–8249. doi: 10.1523/JNEUROSCI.1808-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neher JJ, et al. Inhibition of microglial phagocytosis is sufficient to prevent inflammatory neuronal death. J Immunol. 2011;186:4973–4983. doi: 10.4049/jimmunol.1003600. [DOI] [PubMed] [Google Scholar]
- Fricker M, et al. MFG-E8 mediates primary phagocytosis of viable neurons during neuroinflammation. J Neurosci. 2012;32:2657–2666. doi: 10.1523/JNEUROSCI.4837-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mildner A, et al. Distinct and non-redundant roles of microglia and myeloid subsets in mouse models of Alzheimer's disease. J Neurosci. 2011;31:11159–11171. doi: 10.1523/JNEUROSCI.6209-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neher JJ, et al. Phagocytosis executes delayed neuronal death after focal brain ischemia. Proc Natl Acad Sci U S A. 2013;110:E4098–E4107. doi: 10.1073/pnas.1308679110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perego C, Fumagalli S, De Simoni MG. Temporal pattern of expression and colocalization of microglia/macrophage phenotype markers following brain ischemic injury in mice. J Neuroinflammation. 2011;8:174–174. doi: 10.1186/1742-2094-8-174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Preissler J, et al. Altered microglial phagocytosis in GPR34-deficient mice. Glia. 2015;63:206–215. doi: 10.1002/glia.22744. [DOI] [PubMed] [Google Scholar]
- Hughes MM, Field RH, Perry VH, Murray CL, Cunningham C. Microglia in the degenerating brain are capable of phagocytosis of beads and of apoptotic cells, but do not efficiently remove PrP(Sc), even upon LPS stimulation. Glia. 2010;58:2017–2030. doi: 10.1002/glia.21070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Westenskow PD, et al. Using flow cytometry to compare the dynamics of photoreceptor outer segment phagocytosis in iPS-derived RPE cells. Invest Ophthalmol Vis Sci. 2012;53:6282–6290. doi: 10.1167/iovs.12-9721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- London A, Benhar I, Schwartz M. The retina as a window to the brain-from eye research to CNS disorders. Nat Rev Neurol. 2013;9:44–53. doi: 10.1038/nrneurol.2012.227. [DOI] [PubMed] [Google Scholar]
- Fritzenwanger M, Jung C, Goebel B, Lauten A, Figulla HR. Impact of short-term systemic hypoxia on phagocytosis, cytokine production, and transcription factor activation in peripheral blood cells. Mediators Inflamm. 2011. p. 429501. [DOI] [PMC free article] [PubMed]
- Ragsdale RL, Grasso RJ. An improved spectrofluorometric assay for quantitating yeast phagocytosis in cultures of murine peritoneal macrophages. J Immunol Methods. 1989;123:259–267. doi: 10.1016/0022-1759(89)90230-5. [DOI] [PubMed] [Google Scholar]
- Stokes L, Surprenant A. Dynamic regulation of the P2X4 receptor in alveolar macrophages by phagocytosis and classical activation. Eur J Immunol. 2009;39:986–995. doi: 10.1002/eji.200838818. [DOI] [PubMed] [Google Scholar]
- Kotani N, et al. Intraoperative modulation of alveolar macrophage function during isoflurane and propofol anesthesia. Anesthesiology. 1998;89:1125–1132. doi: 10.1097/00000542-199811000-00012. [DOI] [PubMed] [Google Scholar]
- Xu X, Feng J, Zuo Z. Isoflurane preconditioning reduces the rat NR8383 macrophage injury induced by lipopolysaccharide and interferon gamma. Anesthesiology. 2008;108:643–650. doi: 10.1097/ALN.0b013e318167aeb4. [DOI] [PubMed] [Google Scholar]
- abd-el-Basset E, Fedoroff S. Effect of bacterial wall lipopolysaccharide (LPS) on morphology, motility, and cytoskeletal organization of microglia in cultures. J Neurosci Res. 1995;41:222–237. doi: 10.1002/jnr.490410210. [DOI] [PubMed] [Google Scholar]
- Menon V, Thomas R, Ghale AR, Reinhard C, Pruszak J. Flow cytometry protocols for surface and intracellular antigen analyses of neural cell types. J Vis Exp. 2014. [DOI] [PMC free article] [PubMed]
- Jackson WS. Selective vulnerability to neurodegenerative disease: the curious case of Prion Protein. Dis Model Mech. 2014;7:21–29. doi: 10.1242/dmm.012146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O'Koren EG, Mathew R, Saban DR. Fate mapping reveals that microglia and recruited monocyte-derived macrophages are definitively distinguishable by phenotype in the retina. Sci Rep. 2016;6:20636. doi: 10.1038/srep20636. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Usui Y, et al. Angiogenesis and Eye Disease. Annu Rev Vision. 2015;1:155–184. doi: 10.1146/annurev-vision-082114-035439. [DOI] [PubMed] [Google Scholar]