

Abstract
Background
Gram-negative bacteria of the genus Serratia are potential producers of many useful secondary metabolites, such as prodigiosin and serrawettins, which have potential applications in environmental bioremediation or in the pharmaceutical industry. Several Serratia strains produce prodigiosin and serrawettin W1 as the main bioactive compounds, and the biosynthetic pathways are co-regulated by quorum sensing (QS). In contrast, the Serratia strain, which can simultaneously produce prodigiosin and serrawettin W2, has not been reported. This study focused on analyzing the genomic sequence of Serratia sp. strain YD25T isolated from rhizosphere soil under continuously planted burley tobacco collected from Yongding, Fujian province, China, which is unique in producing both prodigiosin and serrawettin W2.
Results
A hybrid polyketide synthases (PKS)-non-ribosomal peptide synthetases (NRPS) gene cluster putatively involved in biosynthesis of antimicrobial serrawettin W2 was identified in the genome of YD25T, and its biosynthesis pathway was proposed. We found potent antimicrobial activity of serrawettin W2 purified from YD25T against various pathogenic bacteria and fungi as well as antitumor activity against Hela cells. Subsequently, comparative genomic analyses were performed among a total of 133 Serratia species. The prodigiosin biosynthesis gene cluster in YD25T belongs to the type I pig cluster, which is the main form of pig-encoding genes existing in most of the pigmented Serratia species. In addition, a complete autoinducer-2 (AI-2) system (including luxS, lsrBACDEF, lsrGK, and lsrR) as a conserved bacterial operator is found in the genome of Serratia sp. strain YD25T. Phylogenetic analysis based on concatenated Lsr and LuxS proteins revealed that YD25T formed an independent branch and was clearly distant from the strains that solely produce either prodigiosin or serrawettin W2. The Fe (III) ion reduction assay confirmed that strain YD25T could produce an AI-2 signal molecule. Phylogenetic analysis using the genomic sequence of YD25T combined with phylogenetic and phenotypic analyses support this strain as a member of a novel and previously uncharacterized Serratia species.
Conclusion
Genomic sequence and metabolite analysis of Serratia surfactantfaciens YD25T indicate that this strain can be further explored for the production of useful metabolites. Unveiling the genomic sequence of S. surfactantfaciens YD25T benefits the usage of this unique strain as a model system for studying the biosynthesis regulation of both prodigiosin and serrawettin W2 by the QS system.
Electronic supplementary material
The online version of this article (doi:10.1186/s12864-016-3171-7) contains supplementary material, which is available to authorized users.
Keywords: Serratia, Antimicrobial activity, Genome comparisons, Serrawettin W2, Non-ribosomal peptide synthetases, Quorum sensing, Polyphasic taxonomy
Background
Gram-negative bacteria of the genus Serratia have been isolated from water, air, soil, plants, and animals and are members of the Enterobacteriaceae [1]. The ubiquity of Serratia is largely attributed to the variety of compounds that are released into the environment [2, 3]. Some species of Serratia such as S. plymuthica, S. rubidaea, S. marcescens and S. nematodiphila produce a non-diffusible red pigment identified as prodigiosin, which is an alkaloid secondary metabolite with a unique tripyrrole chemical structure [4]. In addition, some species of Serratia also produce various useful secondary metabolites including oocydin A, carbapenem, althiomycin, bacteriocins, and serrawettins [5–7]. These useful secondary metabolites have potential applications in the environmental bioremediation and pharmaceutical industry.
Prodigiosin has been shown to have antimicrobial (antifungal, antibacterial, antiprotozoal), antimalarial, antitumor, and immunosuppressant activities at nontoxic levels [8–10]. Other important secondary metabolites are serrawettins, which are useful biosurfactants produced by Serratia [11]. Three molecular species, serrawettin W1, W2, and W3, have been reported [12]. Serrawettin W1 is a symmetric dilactone structure composed of two serine residues connected with two 3-hydroxydecanoic acids [13]. It has been regarded as a good anti-cancer drug, which could inhibit cell growth and induce apoptosis of several cell lines derived from T-cell leukemia or Burkitt lymphoma [14, 15]. Serrawettin W2 contains a fatty acid connected with five amino acid residues, which was first isolated from S. marcescens in 1986 [16]. Serrawettin W2 is a biosurfactant that can disperse Caenorhabditis elegans [17], and antimicrobial activity against Staphylococcus aureus has been reported [18]. Moreover, there are fewer reports about the bioactivity of cyclic lipopeptides serrawettin W2 and W3. It was found that several Serratia strains, including S. marcescens ATCC 274 [19], S. marcescens 2170 [20], S. marcescens CH-1 [21], and S. marcescens NS-38 [12], could produce prodigiosin and serrawettin W1 at the same time. However, the strains that could parallel-produce prodigiosin and serrawettin W2 as the main bioactive compounds have not previously been published.
Many active metabolites produced by Serratia strains are regulated by quorum sensing (QS), including butanediol fermentation; production of exoenzymes; nuclease and secondary metabolites such as biosurfactant, carbapenem, oocydin A; and prodigiosin [22–26]. Furthermore, a wide spectrum of important processes, such as bioluminescence, motility, sporulation, virulence, and biofilm formation, are also regulated by the QS system, which influences bacteria community gene regulation by cell–cell communication via the production and detection of diffusible auto-inducer signaling molecules [27]. Generally, the most extensively described QS system in Gram-negative bacteria is the autoinducer-1 (AI-1)-mediated system, which employs N-acyl-L-homoserine lactone (AHL) as the autoinducer signaling molecules [28, 29]. Another type of diffusible autoinducer molecules is autoinducer-2 (AI-2), whose synthesis is dependent on LuxS [30, 31]. In Enterobacteriaceae, the AI-2 is internalized into the cells by means of an ABC transporter encoded by lsr operon, and intracellular AI-2 is phosphorylated by LsrK. Subsequently, the phosphorylated form of the signal (AI-2-P) binds LsrR, the repressor of the lsr operon, and induces lsr transcription simultaneously. The further processing of intracellular AI-2-P is then required by LsrF and LsrG proteins [32, 33]. At present, increasingly more AI-2-mediated systems are continuously discovered and explored in Serratia, which is also taxonomically classified as Enterobacteriaceae [34–36]. Nevertheless, the genetic characteristics and molecular mechanisms about the AI-2 systems of Serratia sp. are still less well understood.
In recent years, increasingly more new Serratia species have been identified [37]. To date, eighteen species have been known to belong in the genus Serratia. The availability of complete bacterial genomes has provided new possibilities for bacterial species classification [38–40]. The most relevant comparative parameter for ascertaining the identity of a strain is the calculation of the average nucleotide identity (ANI), a highly accurate technique that measures the genetic and evolutionary distance between two genomes [41]. OrthoANI, using the recently improved new ANI algorithm, is used as an alternative to DNA–DNA hybridization (DDH) for species delineation, and the 95-96 % OrthoANI value is equivalent to the 70 % DDH threshold that is frequently used for species demarcation [42, 43]. Genome-to-genome distance calculator (GGDC) analysis is another parameter based on genome sequences, which have been proposed to discriminate species [44, 45]. Furthermore, multi-locus sequence analysis (MLSA) has been recommended as a replacement for DDH in species delineation for taxonomic studies [46].
In this work, analysis of genomic sequence and metabolites from a novel pigment-producing strain YD25T isolated from rhizosphere soil was reported. The antagonistic capability of YD25T against vital fungal and bacterial pathogens was explored. The main antibacterial compounds simultaneously produced by YD25T were prodigiosin, serrawettin W2, and seven other putative serrawettin W2 analogues. With the completion of the genome sequence of YD25T, taxonomic comparative analysis of the genome and phenotypic analysis data support this strain as a member of a novel and previously uncharacterized Serratia species. Additionally, the comprehensive comparative-genomic analysis and mining of the biosynthetic gene cluster and regulation mechanism of secondary metabolites were also performed.
Methods
Strain isolation and cultivation
The rhizosphere soils under continuously planted burley tobacco were collected from Yongding, Fujian province, China. The site belongs to a subtropical maritime monsoon climate zone and is characterized by hill-gully. The rhizosphere soil was sandy soil, and the plants were infected by the tobacco mosaic virus and pathogen Ralstonia solanacearum due to continuous planting. One gram of the sample was diluted with sterile deionized distilled water serially and spread onto a King’s B medium plate (KB) supplemented with ampicillin (200 mg/mL), chloramphenicol (34 mg/mL) and cycloheximide (80 mg/mL). The agar plates were incubated at 30 °C, and the colonies that appeared were picked up and streaked onto fresh agar plates to obtain pure cultures. In our laboratory, the strain was maintained at −80 °C in 20 % (w/v) glycerol and at 4 °C in culture medium. Morphological features were examined by light microscopy and scanning electron microscopy (S3400, Hitachi) at 30000× magnification.
Antagonistic bioassays of pathogenic bacteria and fungi
Exserohilum turcicum, Fusarium oxysporum, Alternaria alternata and Cochliobolus sativus, all of which are common plant pathogens, were investigated for antagonistic activities of YD25T against fungus. The fungal antagonistic assays were performed following the methods in previous reports [47]. Micrococcus luteus and Ralstonia solanacearum were used as the indicator microorganisms to investigate the antagonistic activities of YD25T against bacterial pathogens. The biofilm and planktonic culture were used to assay antibacterial competition as described previously [48]. Samples containing only YD25T or bacterial pathogens were set up as controls. Three independent experiments were performed for each assay.
Extraction, purification and structure identification of the active compounds
YD25T was inoculated in 50 mL of LB medium at 30 °C for 24 h. The seed culture was transferred to 200 mL of KB medium and incubated for another 72 h. The culture was centrifuged to obtain cells, and the cells were washed with deionized H2O twice and then extracted with 10 volumes of ethanol acidified with HCl (pH 2.0). The sediments and ethanol were removed by centrifugation and evaporation, respectively. The dry material was then further extracted with an excess of ethyl acetate. The crude extract was further purified by flash chromatography employing 200-300 mesh silica gel, while collected fractions were appraised with thin layer chromatography and UV-vis spectral analysis. Oil displacement activity and antibacterial activity of the collected fractions were also checked at the same time. The solvent used for the purification of the crude was ethyl acetate: petroleum ether (1:1, v/v) followed by ethanol.
The fraction eluted with ethanol was further purified using a semi-preparative HPLC system (Shimadzu LC-8A, Japan) equipped with a Sinochrome ODS BP C18 column (10 μm, 20 × 250 mm, Dalian Elite, Dalian, China). Eluent A was composed of purified water containing 0.05 % trifluoroacetic acid, and methanol was selected as eluent B. The following gradient of eluent B was used to run the column: 87-92 % for 0-15 min, 92-97 % for 15-35 min. The flowrate was 10 mL/min. UV detection was performed at 215 nm. The purity of each component was evaluated by an analytical HPLC system (Shimadzu LC-20AT, Japan) equipped with a Sinochrome ODS BP C18 column (5 μm, 4.6 × 250 mm, Dalian Elite). The solvent system and the timetable were the same as the semi-preparative HPLC. The flowrate was 0.6 mL/min.
Electrospray ionization (ESI) mass spectra were acquired on a Bruker ion trap mass spectrometer (Esquire 6000, Bruker, Karlsruhe, Germany) coupled with an Agilent 1100 series HPLC. The purified components were infused to the mass spectrometer directly. One component sw-5 was further identified by nuclear magnetic resonance (NMR). Sw-5 was dissolved in 500 μL of dimethyl sulfoxide-d6. NMR spectra were then recorded at 600 and 153 MHz (Bruker AM 600) for 1H- and 13C-NMR, respectively. Complete chemical shift assignments of sw-5 were supported by 2D NMR (HMQC, HMBC), and the amino acid sequencing was determined by HMBC and ROESY experiments.
Biological activity assays
Production of YD25T antibiotic compounds was tested against bacterial pathogens Bacillus subtilis A47 and M. luteus CGMCC 1.2299 using the paper disc method. To further examine the antibacterial spectrum of sw-5, an expanded indicator panel composed of Gram-negative and Gram-positive bacteria were tested, including Escherichia coli 44102, Pseudomonas aeruginosa A62, Rhodococcus rhodochrous CGMCC 4.1815, Enterococcus faecium CGMCC 1.2025, Klebsiella pneumoniae CGMCC 1.10617, Psychrobacter faecalis CGMCC 1.10869, Acinetobacter baumannii CGMCC 1.6769, Shigella dysenteriae CGMCC 1.1869, and the drug-resistant S. aureus clinical isolates. E. coli 44102, P. aeruginosa A62, B. subtilis A47 and the clinically relevant pathogens S. aureus were purchased from Shaanxi Institute of Microbiology. The other tested strains were purchased from China General Microbiological Culture Collection Center (CGMCC). The culture of indicator strain was diluted to 108 colony forming units (CFU)/mL, and 100 μL of the dilutions was spread on an LB agar plate. A paper disc (6 mm in diameter) impregnated with 10 μL of YD25T compounds (300 μg/mL) dissolved in methanol was placed on the surface of the agar plate. Meanwhile, the paper disc impregnated with 10 μL of methanol was used as a control. The diameter of the inhibition zone was measured after incubation for 24 h.
Cancer cell lines (human cervical cancer cells, HeLa; human epithelial colorectal adenocarcinoma cells, Caco2) and nonmalignant cell lines (human embryonic kidney cell line 293, HEK293; African green monkey kidney cell line, Vero) were incubated with increasing concentrations of sw-5 (3.25-40 μM) to examine the anti-tumor activity of YD25T compounds. After 24 h, 10 μL of MTT solution (Sigma) at 5 mg/mL in PBS was added to each well and incubated for 4 h. The blue MTT formazan precipitate was dissolved in 100 μL of DMSO, and the absorbance at 550 nm was determined.
Genome sequencing, assembly and annotation
Paired-end (PE) strategies with Illumina technology were performed at life sequencing SL (Valencia, Spain) using next-generation sequencing (NGS) to obtain the almost complete genome sequence of YD25T. A strategy using short (approximately 500 nts sequenced by Miseq PE300) and long (consensus sequences of up to 3 kb sequenced by Hiseq2500 PE125) fragments was used to improve the assembly. De novo assembly was performed by the sequencing company using Velvet v 1.2.10. Genome annotation was performed using the life sequencing annotation pipeline using Blast. Contigs were assembled using Velvet software (version v 1.2.10). Sequences were annotated using the NCBI Gene Locator and Interpolated Markov ModelER (GLIMMER; http://www.ncbi.nlm.nih.gov/genomes/MICROBES/glimmer_3.cgi) (locus tag VIB2010). BLASTp [49] was applied to align the amino acid sequences against the COG/SwissProt databases [50, 51].
Genome mining of YD25T
To identify the potential NRPS/PKS secondary metabolite biosynthesis gene clusters encoded within the YD25T genome, the bioinformatics tool anti-SMASH was employed [52]. To investigate the likely function of prodigiosin biosynthesis gene clusters, manual analysis using BLASTp was performed. Open reading frames (ORFs) in the serrawettin W2 and prodigiosin gene cluster were automatically predicted using Glimmer 3.0, available on the National Center for Biotechnology Information (NCBI) website [53]. Multiple alignments were performed using the program Clustal W [54]. Phylogenetic analysis was performed by using MEGA version 6.0 [55] and the neighbor-joining method [56] based on the Kimura two-parameter model of evolution [57].
We retrieved all complete genomic sequences of Serratia sp. including 133 strains present in Genbank (http://www.ncbi.nlm.nih.gov/genome/, last accessed January 20, 2016). For QS analysis, 43 strains are representatives of bacteria with a complete AI-2 system that were chosen from 133 strains to perform the phylogenetic analysis of AI-2 in Serratia sp. The gene content of the lsr (luxS regulated) clusters was manually identified in each strain. Multiple sequence alignments of the lsr clusters were carried out with Clustal W, and then an NJ tree was constructed consisting of ten concatenated genes-lsrA, lsrB, lsrC, lsrD, lsrE, lsrF, lsrG, lsrK, lsrR and luxS, based on the Kimura two-parameter model.
AI-2 bioassay
The detection of AI-2 using the Fe (III) ion reduction assay was performed as previously reported [58]. A working solution of 10 mM 1,10-phenanthroline/3.32 mM Fe (III) was prepared by dissolving 1,10-phenanthroline and ferric ammonium sulfate in deionized distilled water and was adjusted to pH 2 using 1 M HCl. A fresh 1 mg/mL stock solution of ascorbic acid in deionized distilled water was used for standardization and as a positive control for the reduction of Fe (III) ion.
YD25T was grown in KB medium overnight at 30 °C, and then the resulting seed culture was 1/1000 inoculated into the fresh medium and cultured at 30 °C, 200 rpm. Samples were taken from flasks every 2 h over a period of 24 h, and the total number of viable cells was determined by the spreading plate method. At the same time, cell-free supernatant sample (CFS) was prepared by centrifuging cultures at 12,000 rpm for 10 min and passing the medium through 0.22 μm Millipore membrane filters. For detection of AI-2 in CFS, 100 μL of CFS was mixed with 100 μL of Fe (III)-1,10-phenanthroline reagent and left to stand for 1 min to develop the full color. The solution was then diluted to 500 μL using KB medium and scanned for the absorption spectrum against a blank solution within 3 min using a Microplate Luminometer (Epoch, USA). The negative and positive controls used were KB medium without and with ascorbic acid (60 μg/mL), respectively. All assays were performed in triplicate.
Sequence analyses and genome-wide comparative analysis
For strain identification, genomic DNA was extracted and purified by using a MiniBEST bacterial genomic DNA purification kit (TaKaRa). The primers used for PCR amplification of the 16S rRNA gene were the universal primers: 27 F (5’-AGAGTTTGATCCTGGCTCAG-3’) and 1429R (5’-AAGGAGGTGATCCAAGCCGCA-3’) [59]. The amplified 16S rRNA gene was sequenced by TaKaRa. The calculation of pairwise gene sequence identities used the Web-based EzTaxon-e program (http://eztaxon-e.ezbiocloud.net/) [60]. The ANI calculations were determined in this study using the complete genome sequences of Serratia strains available at NCBI and whole-genome sequences from YD25T, S. nematodiphila DSM 21420T, S. marcescens LMG 2792T, S. ureilytica Lr5/4 LG59, S. proteamaculans LMG 8751T, S. liquefaciens LMG 7884T, S. plymuthica S13, S. symbiotica DSM 23270T, S. odorifera LMG 7885T, S. fonticola LMG 7882T. The ten genomes were uploaded into the software package (http://www.ezbiocloud.net/sw/oat) to perform pairwise genome calculations of OrthoANI using the recently improved new ANI algorithm [61]. DNA-DNA hybridization values between strain YD25T and strains of phylogenetically related species were calculated using the GGDC available online (http://ggdc.dsmz.de/distcalc2.php) with the formula 2 method. In addition, the rpoB, gyrB, infB and atpD genes were amplified and sequenced. The sequences of type strains were available from the Institut Pasteur multi-locus sequence typing (MLST) Web site (www.pasteur.fr/mlst). Each gene was blasted against the nucleotide collection (nr/nt) using the BlastN algorithm (http://blast.ncbi.nlm.nih.gov/).
Phenotypic and fatty acid analyses
The physiological and biochemical characteristics were examined according to standard methods described by Grimont et al. [62]. The strain was characterized phenotypically using BiOLOG GEN III microplate (Biolog) and VITEK 2 GN microplate (bioMérieux VITEK-2 Compact) identification systems. Antibiotic sensitivity tests were performed on KB agar at 30 °C containing the following antibiotics: streptomycin, kanamycin, ampicillin, chloramphenicol, tetracycline, carbenicillin, gentamicin, and apramycin. For quantitative analysis of the cellular fatty acid composition, a loopful of cell mass was harvested, and whole-cell fatty acids were analyzed by fatty acid methyl esters (FAMEs).
Results
Antagonistic effects of YD25T on pathogenic bacterial and fungal strains
Strain YD25T was isolated from rhizosphere soils under continuously planted burley tobacco in Fujian Province, China. YD25T produced pigments at 30 °C, which resulted in red colonies on KB medium but could not produce pigments at 37 °C. When it was point-inoculated onto the surface of a KB plate for 24 h, circular irregular margin morphology was observed. Cells are motile, non-spore-forming short rods, 1.2-1.4 × 0.6-0.7 μm and with one weak flagellum (Additional file 1: Figure S1).
In the bacterial–fungal confrontation assays, all tested fungi were inhibited by the YD25T colony as early as the second day and completely inhibited on the fourth day of inoculation (Fig. 1a). All fungi that did not grow past the YD25T colony streaked across the agar plates, though the fungal mycelia also turned a darker color at the later stages of bacteria–fungi interaction (Additional file 2: Figure S2A). The noncontact inhibition seemed to be more evident in C. sativus than others.
Fig. 1.

Antagonistic activities of YD25T against plant pathogenic fungi and Ralstonia solanacearum. a Growth of E. turcicum, F. oxysporum, A. alternate and C. sativus with and without YD25T were closely monitored. b The planktonic culture assay of the quantities of R. solanacearum and YD25T recorded at 0, 2, 4, 6, 8, 10, 12 and 24 h after incubation. Numbers show an average of three replications, and error bars show standard errors of the means
The biofilm culture assay showed that M. luteus and R. solanacearum were dramatically inhibited when they were co-cultured with YD25T at an initial cell ratio of 1:1 after an 8-h incubation. A comparison of the culture without and with YD25T revealed a 104 times increase of M. luteus and 105 times increase of R. solanacearum in the number of viable cells recovered (Additional file 2: Figure S2BC). The planktonic culture assay showed a similar result when YD25T and R. solanacearum were co-cultured at an initial ratio of 1:1 (Fig. 1b). Based on these experiments, it was concluded that YD25T was capable of inhibiting the growth of plant fungal pathogens and bacterial pathogens.
Purification and structure analysis of the active compounds of YD25T
Nine compounds named sw-0 to sw-8 were identified (Fig. 2a). The isolated sw-0 had the maximum absorption spectrum at 532 nm under acidic conditions and at 470 nm under alkaline conditions. ESI–MS analysis indicated that sw-0 had a molecular mass of 323.2 Da. Induced dissociation of the “parent” ion (324.3 Da) revealed the following fragments of the molecular ion: 149.1, 161.0, 238.1, 252.1, 266.1, 292.1 and 309.2 Da (Fig. 2b). The molecular weight of the “parent” ion of sw-0 and the weights of the fragments exactly corresponded to the prodigiosin with elemental composition of C20H25N3O (Fig. 2c).
Fig. 2.

HPLC analysis and structure analyses of the main compounds produced by YD25T. a HPLC analysis of the fraction extracted from YD25T; b The ESI-MS/MS analyses of sw-0 extracted from YD25T; c The structure of sw-0 has been constructed with the positions of bonds whose breakage resulted in the formation of the respective fragments; d The ESI-MS/MS analyses of component sw-5; e The structure of sw-5 has been constructed, and key ROESY and HMBC correlations for sw-5 are shown
Sw-5 was the most abundant compound. Based on ion peaks at m/z 732.2 [M + H]+, m/z 754.6 [M + Na]+, and m/z 770.6 [M + K]+, the molecular mass of sw-5 was determined to be 731.2 Da. Some amino acid residues and their connecting relationship could be determined by comparison of the differences between peaks in MS spectra. Thus, the [M + Na]+ ion was chosen as the precursor ion for further collision-induced dissociation analysis. As shown in the MS/MS spectrum (Fig. 2d), sets of fragment ions were observed. Because Leu and Ile shared the same molecular weight, peaks 754.6 → 641.4 → 494.3 → 306.5 suggested the connection of amino acid residues in the form of Leu-Phe-Thr-Ser or Ile-Phe-Thr-Ser. Peak 561.4 indicated an Ile/Leu linked to the fatty acid chain. Another series fragment formed as an ester bond was cleaved following the double hydrogen transfer mechanism [20]. The peaks of 376.3, 476.3, 579.3 and 623.4 were due to the losses of Thr-Phe-Leu/Ile-H2O (378.3 Da), Phe-Leu/Ile-H2O (278.3 Da), C10H14-Na-H2O (175.2 Da) and Leu/Ile-H2O (131.2 Da). These fragments gave the same structure information in accordance with the above analysis. To further confirm the structure of sw-5, NMR was carried out to identify the amino sequence and the positions of linkages within the carbohydrate and lipid molecules. The complete 13C NMR chemical shift and partly 1H chemical shift assignments are shown in Additional file 3: Table S1. The NMR results demonstrated that the fatty acid residue was β-hydroxydecanoic acid, and five different amino acids constituted the peptide chain. In addition, the chemical shift (δc, 71.7; δH, 4.92) of the third position of fatty acid was consistent with the shift due to lactone ring formation. The sequence of the residues of amino acids was Leu-Ser-Thr-Phe-Ile, determined by interpretation of HMBC data, which was corroborated by NOESY evidence (Fig. 2e). According to comparison with literature data [21, 22], sw-5 was identical to serrawettin W2.
The molecular weights of sw-1, sw-2, sw-6, sw-7, and sw-8 are 703.3, 717.3, 745.4, 757.2 and 759.3 Da, respectively. Sw-3 and sw-4 have the same molecular weight (731 Da) as sw-5, although their retention times are 16.809 and 18.101, respectively. The similar analyses of MS/MS spectra were carried out for the putative serrawettin W2 analogues (Table 1). The results indicated that sw-1 and sw-2 varied at the first or fifth amino acid positions, while sw-6, sw-7 and sw-8 varied at the first, second or fifth amino acid positions. For sw-3 and sw-4, different structures of oligopeptides or the fatty acid chain from serrawettin W2 were observed.
Table 1.
The characteristics and the putative structures of the putative serrawettin W2 analogues
| Compound | [M + H]+ (m/z) | [M + Na]+ (m/z) | [M + K]+ (m/z) | MWb (m/z) | MS fragment (ESI-MS/MS) |
Structure predicatingc |
|---|---|---|---|---|---|---|
| sw-1 | 704.3 | 726.5 | 742.4 | 703.3 | 613.3,595.3a,551.4,533.3a,466.3,448.3a, | C10OH-Abu/Aib-Ser-Thr-Phe-Ile/Leu |
| 365.2, 347.3a | ||||||
| sw-2 | 718.3 | 740.6 | 756.5 | 717.3 | 641.3,627.4a,565.3,547.3a,494.3,480.3a, | C10OH-Val-Ser-Thr-Phe-Ile/Leu |
| 448.3, 379.3, 361.2a | ||||||
| sw-3 | 732.2 | 754.6 | 770.4 | 731.2 | 641.4,623.5a,579.8,551.4a,494.3,476.3a, | C10OH-Leu/Ile-Ser-Thr-Phe-Ile/Leu |
| 448.4, 393.3, 375.1a | ||||||
| sw-4 | 732.3 | 754.6 | 770.6 | 731.3 | 641.3,623.4a,579.3,551.1a,494.3,476.3a, | C10OH-Leu/Ile-Ser-Thr-Phe-Ile/Leu |
| 448.3, 393.1, 375.2a | ||||||
| sw-6 | 746.4 | 768.6 | 784.5 | 745.4 | 655.4,637.2a,593.4,575.2a,508.3,490.3a, | C10OH-Leu/Ile-Thr-Thr-Phe-Ile/Leu |
| 462.3, 407.2, 389.1a | ||||||
| sw-7 | 758.2 | 780.5 | 796.5 | 757.2 | 667.4, 649.5a, 605.7, 587.4a, 520.2, | C10OH-Leu/Ile-Leu/Ile-Thr-Phe-Ile/Leu |
| 502.3a, 474.3, 419.2, 402.1a | ||||||
| sw-8 | 760.3 | 782.5 | 798.3 | 759.3 | 669.3, 651.2a, 625.5, 607.4a, 522.3, | C10OH-Leu/Ile-Asp-Thr-Phe-Ile/Leu |
| 504.5a, 476.5, 421.3, 403.3a |
aThese peaks were formed as an ester bond was cleaved following double hydrogen transfer (DHT) mechanism. bThe molecular weight (MW) was based on the peaks of [M + H]+, [M + Na]+ and [M + K]+. cMass spectrometry couldn’t distinguish between isomers like leucine and isoleucine
Biological activities of the active compounds of YD25T
Among the eight compounds (sw-1, 2, 3, 4, 5, 6, 7, 8) of YD25T, sw-5 (serrawettin W2) inhibited M. luteus CGMCC 1.2299 growth highly and B. subtilis A47 minimally, but sw-6 showed the opposite results. Sw-7 and sw-8 had a slight inhibition to M. luteus CGMCC 1.2299 and no activity to B. subtilis A47. The other compounds had no antibacterial activities (Fig. 3a, b). To further examine the antibacterial spectrum of serrawettin W2, an expanded indicator panel composed of Gram-negative and Gram-positive bacteria was tested. The result indicated that serrawettin W2 had activities to R. rhodochrous CGMCC 4.1815, S. dysenteriae CGMCC 1.1869, and P. aeruginosa A62. Furthermore, it was noteworthy that among thirty drug-resistant S. aureus clinical isolates, serrawettin W2 exhibited evident inhibitory activities to nine strains and slight inhibition to twelve strains (Table 2).
Fig. 3.
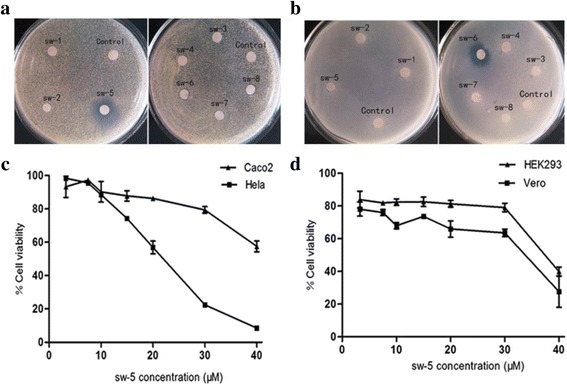
The biological activities of the compounds from YD25T. a The antibacterial activities of the components against M. luteus CGMCC 1.2299; b The antibacterial activities of the components against B. subtilis A47; c The cytotoxicity on Hela and Caco2 cells of sw-5 (serrawettin W2); d The cytotoxicity on nonmalignant cells of sw-5 (serrawettin W2). Numbers show an average of three replications, and error bars show standard errors of the means
Table 2.
Antibacterial activity of sw-5 (serrawettin W2) produced by YD25T
| Bacteria | Drug resistance | Inhibitory zone | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | diameter (mm) | |
| Staphylococcus aureu A1 | R | R | R | R | R | R | R | - | - | NI |
| Staphylococcus aureu A2 | R | S | S | R | R | R | R | - | - | SI |
| Staphylococcus aureu A3 | S | R | S | S | R | S | S | - | - | NI |
| Staphylococcus aureu A4 | R | R | R | R | R | R | R | - | - | NI |
| Staphylococcus aureu A5 | R | R | R | R | R | R | R | - | - | SI |
| Staphylococcus aureu A6 | R | R | R | R | R | R | R | - | - | SI |
| Staphylococcus aureu B1 | S | R | S | R | R | S | S | - | - | NI |
| Staphylococcus aureu B2 | R | R | R | R | R | R | R | - | - | SI |
| Staphylococcus aureu B3 | R | R | R | R | R | R | R | - | - | NI |
| Staphylococcus aureu B4 | S | R | S | S | R | S | S | - | - | NI |
| Staphylococcus aureu B5 | S | S | S | S | R | S | S | - | - | SI |
| Staphylococcus aureu B6 | S | S | S | S | S | S | S | - | - | SI |
| Staphylococcus aureu C1 | R | R | R | R | R | R | R | - | - | NI |
| Staphylococcus aureu C2 | R | R | R | R | R | R | R | - | - | SI |
| Staphylococcus aureu C3 | S | R | S | S | R | I | S | - | - | 11.2 ± 0.4 |
| Staphylococcus aureu C4 | R | R | R | R | R | R | R | - | - | SI |
| Staphylococcus aureu C5 | R | R | R | R | R | R | R | - | - | SI |
| Staphylococcus aureu C6 | S | R | S | S | R | S | S | - | - | 9.2 ± 0.3 |
| Staphylococcus aureu D1 | R | S | R | R | R | R | R | - | - | NI |
| Staphylococcus aureu D2 | S | R | S | S | R | S | S | - | - | 7.6 ± 0.4 |
| Staphylococcus aureu D3 | R | R | R | R | R | R | R | - | - | SI |
| Staphylococcus aureu D4 | R | R | R | R | R | R | R | - | - | NI |
| Staphylococcus aureu D5 | R | R | S | R | R | R | R | - | - | 8.5 ± 0.5 |
| Staphylococcus aureu D6 | R | S | S | R | R | R | R | - | - | SI |
| Staphylococcus aureu E1 | S | R | S | R | R | S | S | - | - | 11.2 ± 0.3 |
| Staphylococcus aureu E2 | R | S | R | R | R | R | R | - | - | SI |
| Staphylococcus aureu E3 | - | - | - | S | - | S | S | R | R | 8.3 ± 0.2 |
| Staphylococcus aureu E4 | - | - | - | - | - | S | S | R | R | 11.4 ± 0.4 |
| Staphylococcus aureu E5 | S | I | S | S | R | S | S | - | - | 10.1 ± 0.3 |
| Staphylococcus aureu E6 | S | R | S | S | R | S | S | - | - | 8.1 ± 0.4 |
| Escherichia coli 44102 | - | - | - | - | - | - | - | - | - | NI |
| Pseudomonas aeruginosa A62 | - | - | - | - | - | - | - | - | - | 14.4 ± 0.4 |
| Bacillus subtilis A47 | - | - | - | - | - | - | - | - | - | SI |
| Rhodococcus rhodochrous | - | - | - | - | - | - | - | - | - | 12.5 ± 0.3 |
| CGMCC 4.1815 | ||||||||||
| Enterococcus faecium | - | - | - | - | - | - | - | - | - | NI |
| CGMCC 1.2025 | ||||||||||
| Klebsiella pneumoniae | - | - | - | - | - | - | - | - | - | NI |
| CGMCC 1.10617 | ||||||||||
| Psychrobacter faecalis | - | - | - | - | - | - | - | - | - | NI |
| CGMCC 1.10869 | ||||||||||
| Acinetobacter baumannii | - | - | - | - | - | - | - | - | - | NI |
| CGMCC 1.6769 | ||||||||||
| Shigella dysenteriae | - | - | - | - | - | - | - | - | - | 10.5 ± 0.5 |
| CGMCC 1.1869 | ||||||||||
| Micrococcus luteus | - | - | - | - | - | - | - | - | - | 19.4 ± 0.5 |
| CGMCC 1.2299 | ||||||||||
1, prostaphlin (10 units); 2, clindamycin (2 μg); 3, rifampicin (5 μg); 4, levofloxacin (5 μg); 5, erythromycin (15 μg); 6, amikacin (30 μg); 7, cefazolin (30 μg); 8, paediatric compound sulfamethoxazole tablets (23.75 μg); 9, ampicillin (10 μg). R, resistance to the antibiotic; S, sensitive to the antibiotic; I, intermediate; “-”, not determined. NI, no inhibition; SI, slight inhibition. SI indicated the diameter of the inhibition zone was less than 7 mm. The diameter of the control was 6 mm. Numbers show an average of three replications
In addition to the excellent antibacterial activity, serrawettin W2 also exhibited anticancer activity. Serrawettin W2 showed cytotoxicity on Hela and Caco2 cells at concentrations from 3.25 μM to 30 μM. The IC50 of serrawettin W2 was 20.9 μM for the Hela cell line and 54.1 μM for Caco2 (Fig. 3c). Thus, serrawettin W2 inhibited growth of Hela cells more significantly than Caco2 cells. For the normal cell lines, serrawettin W2 did not show a marked decrease in viability of Vero cells and HEK293 cells at concentrations of up to 30 μM. Furthermore, a higher concentration (40 μM) of serrawettin W2 resulted in an approximate 60 % cell death (Fig. 3d). These results indicate that serrawettin W2 can significantly suppress the growth of cancer cell lines and has slight effects on the viability of nonmalignant cells.
Analysis of the genomic sequence to identify putative serrawettin W2 gene cluster
The new genomic sequence for YD25T had 17 contigs with a total length of approximately 5,115,690 bp (Additional file 4: Figure S3). The mean contig size was 300,923 bp, with the N50 (50 % of the genome is contained in contigs of size N or greater) contig size being 1,200,118 bp. Analysis of the genome sequence of YD25T resulted in the identification of a putative cyclic lipopeptide antibiotics serrawettin W2 biosynthesis gene cluster, which is a more than 21 kb interrupted DNA sequence characterized by a hybrid PKS-NRPS system. The PKS gene consists of an acyltransferase (AT), ketosynthase (KS), and a keto reductase (KR) domain. In the putative typical features of the NRPS gene, there are a total of five modules, and each module consists of specific condensation (C), adenylation (A), and thiolation (T) domains. The five modules harbor one thioesterase (TE) domain. The distance between the PKS and NRPS gene is 9478 bp on a linear map of the genome sequence of YD25T.
Speculation of the domain functions is based on the sequence homology to known PKS and NRPS domains. The putative peptide sequence of serrawettin W2 is Leu-Ser-Thr-Phe/Val-Ile/Val. According to the results of structural analysis of serrawettin W2, the fourth and fifth amino acids are Phe and Ile, and the prediction is in full agreement with the peptide sequence of serrawettin W2 determined by the results of MS. Based on the presence of PKS and NRPS encoding genes, the pathway for serrawettin W2 biosynthesis is deduced (Fig. 4). Firstly, the fatty acid (FA) of the C10 unit, which is used as the precursor material, is synthesized by PKS SwrEFG and other undetermined proteins and is released as a fatty acyl-CoA. Secondly, a 17.7 kb swrA encodes the core W2-peptide chain (the structure of the CLP with a 5-amino acid peptide moiety), which contains a total of five modules. The N-terminal C-domain of the SwrA initiation module is predicted to catalyze the condensation of the fatty acyl-CoA and leucine. After initiation, chain elongation is mediated by the other three linear domains correlated with the serine, threonine, and phenylalanine, respectively. Finally, the oligopeptide is transferred onto the active-site isoleucine of the C-terminal TE domain of the last module. This organization is consistent with a role in catalyzing the last step of chain elongation, lactonization, and the subsequent cyclorelease of the cyclic lipopeptide. The genome-based identification and structure prediction of serrawettin W2 provides a basis for further improving the production of serrawettin W2 as a useful metabolite by YD25T.
Fig. 4.

Genetic organization of swr biosynthetic gene cluster and proposed biosynthetic pathway for serrawettin W2 identified in YD25T. Model for the biosynthesis of serrawettin W2 showing the structures of some of the proposed intermediates. SwrEFG, and SwrA form a hybrid PKS-NRPS system, which consists of five modules as indicated. Underneath the genes are the relative size, module and domain organization, and their predicted products of the PKS-NRPS. The amino acids are predicted to be incorporated into serrawettin W2 peptide moiety based on specific signature sequences in each A-domain. Domains within the PKS-NRPS are as follows. AT: acyltransferase; KS, ketosynthase; KR: ketoreductase; A: adenylation; C: condensation; T: thiolation; TE: thioesterase
Identification of the prodigiosin biosynthetic gene cluster
A putative prodigiosin gene cluster was found in the genomic sequence of YD25T, and putative functions of pig protein were assigned based on the results of BLASTp searches and compared with the other Serratia pig proteins in S. marcescens ATCC 274, S. plymuthica AS 13, and Serratia sp. ATCC 39006 (Additional file 5: Table S2). Subsequently, comparative sequence analysis of the pig clusters among the pigmented Serratia species was shown (Fig. 5). The pig clusters were observed in the complete genomic sequences of 34 strains among the total of 133 Serratia species present in Genbank (last accessed January 20, 2016). They could be divided into three types: type I-pig cluster is flanked by cueR and copA in 29 strains represented by S. marcescens ATCC 274; type II-cueR and copA are adjacent to each other and in the downstream of the pig cluster in 4 strains including S. plymuthica AS9, S. plymuthica AS12, S. plymuthica AS13 and S. rubidaea CIP103234; type III-pigO is present immediately downstream of the pig cluster only in Serratia sp. ATCC39006. Furthermore, pig cluster genes of YD25T showed 98, 77 and 74 % identity at the amino acid level to the three types of Serratia pig-encoding genes, respectively. Consequently, the prodigiosin biosynthesis gene cluster in YD25T belongs to the type I pig cluster, which is the main form of pig-encoding genes existing in the most of pigmented Serratia species.
Fig. 5.

Comparison of prodigiosin biosynthetic gene clusters among pigmented Serratia sp. The applicable gene sequence data are from a variety of sources: S. marcescens ATCC274, S. nematodiphila DSM 21420T, S. plymuthica AS13, S. rubidaea CIP101234T, Serratia sp. ATCC 39006. The genetic organizations of three types of prodigiosin biosynthetic gene clusters are symbolized by arrows. The positions and intergenic regions of each pig gene along a linear representation have been indicated. The identities of the homologue gene clusters between YD25T and the three types are 98 %, 77 %, and 74 %, respectively
Analysis of quorum sensing (QS) system and detection of AI-2 signal molecule
AI-1 and AI-2 are the most extensively described QS system in Gram-negative bacteria. For the AI-1 system, two luxR homologue genes-sfcR and sfsR, both with the size of 771 bps-were identified within the YD25T genome. However, no luxI homologue gene was identified within the upstream and downstream of sfcR and sfsR, so we hypothesized that these are solo luxR. For the AI-2 system, there are luxS and a complete lsr gene clusters including lsrBACDEF, lsrGK, and lsrR within the YD25T genome (Fig. 6a). The predicted functions were assigned (Additional file 6: Table S3), and the result of comparison with the lsr cluster in Salmonella typhimurium LT2, E. coli K-12, and E. fergusonii ATCC 35469 was shown that the ten proteins in YD25T were similar in size to the homologues encoded in the lsr cluster. LuxS and LsrGK exhibited a high degree of sequence similarity at the amino acid level, though LsrBACDEF and LsrR show a lower identity to their homologues from S. typhimurium LT2, E. coli K-12, and E. fergusonii ATCC 35469. To search for orthologs of LuxS and Lsr proteins in Serratia sp., we carried out an analysis against all 133 completely sequenced genomes present in the NCBI database as of January 2016. The isolates with Lsr proteins (LsrBACDEF and LsrGK) identified as being orthologs total 80 strains, in which lsrR was not detected in the genome sequences of 37 organisms. Therefore, there are 43 strains whose genome sequences include luxS and the complete lsr operon (lsrBACDEF, lsrGK, and lsrR). Subsequently, the result of phylogenetic analysis using concatenated amino acid sequences of 10 proteins (LuxS, LsrBACDEF, LsrGK and LsrR) among the 43 strains indicated that YD25T formed an independent branch from the other organisms (Fig. 6b). Furthermore, analyzing the gene clusters of prodigiosin and serrawettin W2 within these 43 organisms clearly revealed two distinct groups (group I and group II). Group I includes Serratia strains containing only the prodigiosin gene cluster, while group II includes Serratia strains containing only the serrawettin W2 gene cluster. Importantly, YD25T producing prodigiosin and serrawettin W2 simultaneously formed an independent branch and was clearly distant from the two groups. These results are the same as the phylogenetic analysis results using the ten-gene (lsr operon and luxS) sequences (Additional file 7: Figure S4).
Fig. 6.

AI-2-mediated quorum sensing system in YD25T. a Distribution and comparison of gene organization in the complete lsr operon between the Serratia sp. and the S. typhimurium/E. coli. The orientation of the arrows indicates the direction of transcription; b Phylogenetic relationships based on the concatenated amino acid sequences of complete Lsr protein and LuxS. Boxes represent the strains in which the biosynthetic gene cluster of prodigiosin (red) or serrawettin W2 (blue) are observed in the genome. Bootstrap analysis (1000 resamplings) was used to evaluate the topology of the NJ tree. Bar, 0.005 substitutions per nucleotide position. All sequences were retrieved from the NCBI database or in published genome projects. c AI-2 bioassay of YD25T by the Fe (III) ion reduction assay. Representative absorption spectra of 10 mM Fe(III)-1,10-phenanthroline with 60 μg/mL ascorbic acid, a cell-free supernatant sample of YD25T or KB medium; d Cell growth of YD25T cultures in KB and the production of extracellular AI-2 levels in the culture media. Numbers show an average of three replications and are derived from one independent sample
In order to examine whether an AI-2 signal molecule is indeed produced in YD25T, the Fe (III) ion reduction assay was used as a bioassay method to identify the diffusible autoinducer signaling molecules and detect the level in different bacterial growth phases. In the Fe (III) ion reduction assay, the spectra following the addition of ascorbic acid (as the positive control) or AI-2 to the Fe(III)-1,10-phenanthroline had the maximum absorption wavelength at 510 nm due to the formation of the [(o-phen)3Fe]SO4 ferroin complex. The AI-2 production of YD25T is shown in Fig. 6c. Representative absorption spectra of 10 mM Fe (III)-1, 10-phenanthroline in the presence of 60 μg/mL ascorbic acid or CFS of YD25T were similar. In contrast, the spectrum of Fe(III)-1,10-phenanthroline alone or with KB medium showed no maximum absorption wavelength at 510 nm. The assay of the AI-2 level in different bacterial growth phases is shown in Fig. 6d. These results clearly indicate that AI-2 was maximally produced by YD25T during the mid-to late-log phase and achieved the maximum at the stationary phase, and then the extracellular AI-2 gradually decreased.
Taxonomic comparative analysis of the genome of YD25T with other similar species
The resulting 16S rRNA gene sequence (1534 bp) of YD25T was initially deposited in the GenBank database with the accession number KM093865. Comparisons of 16S rRNA gene sequences revealed that the YD25T strain was related most closely to S. nematodiphila DSM 21420T (99.7 % similarity) found in Heterorhabditidoides chongmingensis. It was also closely related to S. marcescens LMG 2792T (99.4 % similarity). Strain YD25T shared similarity less than 98.7 % with other type strains (Additional file 8: Table S4). Phylogenetic analysis based on the neighbor-joining algorithms revealed that strain YD25T was included in the cluster of species of the genus Serratia. The phylogenetic tree showed that strain YD25T was related most closely to S. nematodiphila and S. marcescens. However, the sequence divergence between strain YD25T and the type strains of phylogenetically related species suggested that strain YD25T represents a novel species within the genus Serratia (Additional file 9: Figure S5). The mean DNA G + C content of strain YD25T was calculated to be 59.62 mol%. This value is similar to S. marcescens LMG 2792T (57.5-60.0 mol%) [2], S. nematodiphila DSM 21420T (59.52 mol%) [63], and S. ureilytica LMG 22860T (60 mol%) [64].
The ten genomes were analyzed by pairwise genome calculations using OrthoANI, and the results are shown in Fig. 7a. In all cases, these OrthoANI values clearly indicate that YD25T has OrthoANI less than 95 % of other closely related Serratia species, representing a different genomo-species from that currently classified within the group and again confirming species-level distinctiveness. Although the genome sequence of the S. ureilytica and S. plymuthica type strains is unavailable, the two species may belong to the group represented by Lr5/4 LG59 and S13. Although the genome sequences of the remaining species Serratia is not publicly available, the comparison results are unaffected because these species give the low 16S rRNA identity compared with YD25T. DDH values between strain YD25T and S. nematodiphila DSM 21420T, S. marcescens LMG 2792T, S. ureilytica Lr5/4 LG59, S. proteamaculans LMG 8751T, S. liquefaciens LMG 7884T, S. plymuthica S13, S. symbiotica DSM 23270T, S. odorifera LMG 7885T, S. fonticola LMG 7882T were < 58 % (Table 3), confirming that strain YD25T represents a novel species of the genus Serratia.
Fig. 7.

Taxonomic classification by genome-wide comparative analysis of YD25T. a Heatmap generated with OrthoANI values calculated from the OAT software. Pairwise OrthoANI calculations among the selected Serratia genomes. A comparison with genome sequences of the strains of other established Serratia species showed that strain YD25T shared < 95 % similarity: 94.5 % with S. nematodiphila, 94.2 % with S. marcescens, 93.9 % with S. ureilytica, 84.4 % with S. plymuthica, 83.5 % with S. proteamaculans, 83.4 % with S. liquefaciens, 82.7 % with S. symbiotica, 82.0 % with S. odorifera and 80.8 % with S. fonticola. b Neighbor-joining tree showing the phylogenetic relationships of strains YD25T and phylogenetically related reference strains based on concatenated partial rpoB, atpD, gyrB and infB gene sequences. Bootstrap analysis (1000 resamplings) was used to evaluate the topology of the NJ tree, and the bootstrap values >70 % are displayed at branch points. Bar, 0.02 substitutions per nucleotide position
Table 3.
DNA-DNA hybridization values between strain YD25T and strains of phylogenetically related species. The parameter is calculated using the genome-to-genome distance calculator (GGDC) available online with formula 2 method
| Strains | Identities/HSP length | Probability > 70 % | Probability > 79 % |
|---|---|---|---|
| S. nematodiphila DSM 21420T | 57.90 % ± 2.77 | 45.06 % | 35.51 % |
| S. marcescens LMG 2792T | 56.40 % ± 2.75 | 39.98 % | 8.95 % |
| S. ureilytica Lr5-4 LG59 | 55.30 % ± 2.72 | 35.95 % | 7.86 % |
| S. plymuthica S13 | 28.30 % ± 2.43 | 0.05 % | 0.02 % |
| S. proteamaculans LMG 8751T | 27.20 % ± 2.43 | 0.03 % | 0.02 % |
| S. symbiotica DSM 23270T | 27.00 % ± 2.42 | 0.03 % | 0.01 % |
| S. liquefaciens LMG 7884T | 27.00 % ± 2.42 | 0.03 % | 0.01 % |
| S. odorifera LMG 7885T | 25.10 % ± 2.40 | 0.01 % | 0.01 % |
| S. fonticola LMG 7882T | 24.60 % ± 2.40 | 0.01 % | 0 % |
In addition, once the gene sequences were retrieved from the genomes, an MLSA approach was performed using the concatenated sequences of the four housekeeping genes (rpoB, gyrB, infB and atpD; total alignment length 2636 bp). Strain YD25T showed 98 % concatenated partial rpoB, gyrB, infB and atpD gene sequence similarity to the types of the most phylogenetically related species, S. nematodiphila and S. marcescens (Additional file 10: Table S5). For the analysis, strain YD25T formed an independent branch and was clearly distant from the core species of the group (S. nematodiphila, S. marcescens and S. ureilytica) (Fig. 7b). The bootstrap values indicated a very stable branching order and mirrored the results observed with the phylogenetic reconstruction analyses based on 16S rRNA. Individual rpoB gene sequences were also analyzed, and the tree agreed in the topology and showed a consistent branching order with the tree of the remaining four housekeeping genes (Additional file 11: Figure S6). These results reinforced the observation that YD25T represented a novel species of the group.
Biochemical and physiological features of YD25T
The biochemical, physiological, and morphological characteristics of strain YD25T are summarized in the species description (Additional file 12), and a comparison of these features with the closely related species is presented in Table 4. In contrast to S. nematodiphila DSM 21420T or S. marcescens LMG 2792T, strain YD25T can use adonitol, D-arabinose, D-xylose and melibiose but cannot utilize D-lactose or raffinose and is methyl red test-positive. Importantly, three highly specific features of the YD25T taxon are the presence of urease activity, absence of lipase activity and failure of lactate assimilation. The presence of urease activity may be explained by the presence of gene coding for α, β, and γ subunits of urease, and the absence of lipase activity may be explained by the absence of gene coding for lipase, as observed in the genome of YD25T. Concerning the absence of lactate assimilation, we observed that the genome of YD25T does not contain the lactate oxidase encoding gene. These biochemical and physiological features distinguish YD25T from its close relatives.
Table 4.
Biochemical and physiological characteristics of strain YD 25T and phylogenetically related reference type strains in the genus Serratia
| Characteristic | 1 | 2 | 3 | 4 | 5 | 6 | 7 |
|---|---|---|---|---|---|---|---|
| Fluorescence | + | - | - | - | - | NA | - |
| Pigment production | + | + | - | - | + | - | + |
| Methyl red test | - | - | + | + | - | + | + |
| Urease | - | - | + | - | - | - | + |
| Lipase (Tween 80) | + | + | + | - | + | + | - |
| Adonitol | + | - | + | - | + | + | + |
| D-Lactose | + | - | - | + | + | + | + |
| D-Arabinose | + | - | - | + | + | - | + |
| Raffinose | + | - | - | + | + | + | - |
| D-Xylose | + | - | + | + | + | + | + |
| Melibiose | + | - | + | + | + | + | + |
| Lactate | + | + | + | + | + | + | - |
| D-Serine | + | + | + | - | - | NA | + |
| L-Ornithine | + | + | + | + | - | + | + |
| Alanine | + | + | + | + | - | + | + |
| L-Proline | + | + | + | + | + | + | + |
| Phenylalanine | - | + | + | - | - | - | - |
| DNA G + C content (mol%) | 59.52 | 57.5-60 | 60 | NA | NA | 59.6 | 59.62 |
1, S. nematodiphila DSM 21420T; 2, S. marcescens LMG 2792T; 3, S. ureilytica LMG 22860T; 4, S. odorifera LMG 7885T; 5, S. rubidaea LMG 5019 T; 6, S. ficaria LMG 7881T; 7, strain YD25T. +, positive; -, negative; NA, data not available
The comparative fatty acid compositions of YD25T and the type strains of phylogenetically related reference strains are given in Table 5. The major fatty acid in strain YD25T is C16:0, which was in agreement with the profiles of most Serratia species. More importantly, YD25T contained the major fatty acid C16:0 (30.73 %), which was the same as those of S. nematodiphila DSM 21420T, S. marcescens LMG 2792T, and S. ureilytica LMG 22860T. However, the relative proportions of the major fatty acids of YD25T were significantly different from the three close relative strains. In particular, the predominant major fatty acid of YD25T was C16:0, C18:1 ω7c and C17:0 cyclo, while those of the other three strains were C16:0, C17:0 cyclo and C19:0 cyclo ω8c; C16:0, C18:1 ω7c and C17:0 cyclo; and C16:0, C18:1 ω7c and summed feature 1 (comprises C14:0 3-OH/C16:1 iso I), respectively. In addition, there were significant differences in the percentages of other fatty acids between YD25T and each of the latter three phylogenetically related reference type strains. The most marked differences between YD25T and the abovementioned three reference type strains tested concerned the fatty acids summed feature 2 (comprises C16:1 ω7c/C16:1 ω6c) and C19:0 cyclo ω8c, which were present in YD25T and absent from large amounts in most tested strains. However, the YD25T presented significantly higher amounts of C14:0 2-OH and C12:0 2-OH and significantly lower amounts of C14:0 compared to the latter strains.
Table 5.
Cellular fatty acid compositions (%) of strain YD25T and phylogenetically related reference type strains in the genus Serratia
| Fatty acid | 1 | 2 | 3 | 4 | 5 | 6 | 7 |
|---|---|---|---|---|---|---|---|
| C10:0 3-OH | tr | ND | ND | ND | ND | ND | tr |
| C12:0 | 2.47 | 1.46 | 1.4 | 2.95 | 4.28 | 5.80 | 1.62 |
| C12:0 2-OH | tr | tr | tr | tr | ND | 1.51 | 1.73 |
| C12:0 3-OH | tr | ND | ND | ND | ND | ND | tr |
| C14:0 | 8.4 | 6.94 | 6.28 | 7.4 | 6.52 | 8.14 | 3.39 |
| C14:0 2-OH | 1.02 | 2.05 | 2.21 | ND | ND | ND | 2.89 |
| C15:0 | tr | ND | tr | ND | 1.31 | ND | ND |
| C16:0 | 34.76 | 31.86 | 31.01 | 30.74 | 17.98 | 24.51 | 30.73 |
| C16:0 ω5c | ND | ND | ND | ND | ND | ND | tr |
| C17:0 cyclo | 20.03 | 11.68 | 2.69 | 4.96 | ND | 12.45 | 14.56 |
| C17:0 | ND | tr | tr | ND | tr | 2.26 | tr |
| C18:1 ω7c | 1.67 | 16.88 | 17.89 | 14.4 | 11.39 | 9.56 | 15.29 |
| C19:0 iso | tr | ND | ND | ND | ND | ND | tr |
| C19:0 cyclo ω7c | 17.24 | 1.24 | ND | tr | ND | ND | ND |
| C19:0 cyclo ω8c | ND | ND | tr | ND | ND | 1.68 | 2.42 |
| Summed featuresa | |||||||
| 1 | 7.56 | 8.44 | 8.01 | 8.98 | 4.11 | 14.54 | 8.36 |
| 2 | ND | ND | ND | ND | ND | ND | 15.78 |
aSummed feature 1 comprises C14:0 3-OH/C16:1 iso I; summed feature 2 comprises C16:1 ω7c/C16:1 ω6c
Strains: 1, S. nematodiphila DSM 21420T; 2, S. marcescens LMG 2792T; 3, S. ureilytica LMG 22860T; 4, S. odorifera LMG 7885T; 5, S. rubidaea LMG 5019 T; 6, S. ficaria LMG 7881T; 7, strain YD25T. tr, Trace amount (<1.0 %); ND, not detected
In combination with the unique features of its genomic sequence, the biochemical, physiological, and morphological characteristics of strain YD25T confirmed that it represents a distinct and separate species within the genus Serratia.
Discussion
Serratia belongs to Enterobacteriaceae and can produce various extracellular products, including lipases, chitinases, nucleases, proteases, and various antimicrobial secondary metabolites [65]. This has been linked to the ability of Serratia strains to colonize a wide range of ecological niches [4]. Prodigiosin is the most characterized antimicrobial compound in Serratia strains [66]. Serrawettin W2 acts as a biosurfactant to reduce the surface tension of the environment to allow bacterial spreading across surfaces and has been shown to possess antimicrobial activity [67]. Furthermore, Serratia species produce a wealth of volatile organic compounds, such as sodorifen, dimethyl trisulfide, methanethiol, and terpenoids, which might influence the growth of phytopathogenic fungi or bacteria [68, 69]. From these aspects, it was established that YD25T is an unusual antagonistic bacterium through secretion of a variety of different antimicrobial substances.
YD25T has a special trait of simultaneous production of both serrawettin W2 and prodigiosin. Such a co-production feature has not been reported in other Serratia sp. strains. For serrawettins, serrawettin W1 was produced by pigmented S. marcescens ATCC 274 [70], S. marcescens CH-1 [71], S. marcescens NS-38 [72], S. marcescens WW4 [73] and so on, while serrawettin W2 and W3 were produced by nonpigmented Serratia strains [11, 73]. Moreover, Serratia strains that could produce more than one type of serrawettin have not been recognized. Serrawettin W2 was originally isolated from nonpigmented S. marcescens NS 25 [16]. S. marcescens Db10 is an insect pathogen that could produce serrawettin W2 but lacks the genes to produce prodigiosin [6]. There are pigment genes, and a 2631 bp sequence of swrA was found in S. marcescens W2.3 isolated from diseased tilapia fish; nevertheless, the related products have not been isolated and identified [74]. To date, YD25T is the first reported Serratia strain that could concurrently produce the antibiotic secondary metabolite, prodigiosin and serrawettin W2 and also produce a spectrum of putative serrawettin W2 analogues simultaneously.
Cyclic lipopeptides exhibiting antibacterial activities, such as massetolide, viscosin, syringomycin, arthrofactin, and orfamide, have emerged as promising candidates for the development of new antibiotics [75, 76]. Serrawettin W2 is a broad spectrum lipopeptide that could suppress both Gram-positive and Gram-negative bacteria in our research. Furthermore, serrawettin W2 has cytotoxicity on cancer cells and shows selectivity for different cancer cell lines. Therefore, serrawettin W2 has attracted interest as a potentially valuable antibiotic. At present, the available biosynthetic gene cluster for serrawettin W2 has not been identified, unless the 2.8 kb incomplete swrA is involved in the biosynthesis of the serrawettin W2 NRPS gene in S. marcescens MG1 [73] and S. marcescens A88copa13 [74]. In our study, a gene cluster of serrawettin W2 biosynthesis in YD25T was directed by a hybrid PKS-NRPS system, and domain functions and a biosynthesis pathway were proposed. Although the formation of the C10 unit fatty acid chain by the PKS remains unclear; Serratia, a genetically tractable bacterium, has allowed a detailed dissection of how the secondary metabolite is biosynthesized. Furthermore, modules and domains of PKS and NRPS enzymes within the serrawettin W2 biosynthetic gene cluster offers potential future exploitation for the generation and synthesis of useful and modified natural products.
In a complete AI-1 system, the luxI/R homologs interact with each other [28]. The LuxI-type proteins synthesize various AHL signals, and LuxR-type transcriptional regulatory proteins bind their cognate signal and in turn induce/repress multiple gene expression accordingly [27]. However, luxI and luxR genes do not occur in pairs in the YD25T genome. The unpaired luxR gene has also been reported in the S. marcescens W2.3 genome [74]. CarR as a solo luxR, regulates the production of two secondary metabolite antibiotics, prodigiosin and carbapenem, in S. marcescens ATCC 39006 [77]. However, the sequence similarities at the amino acid level of sfcR and sfsR with carR are only 37 % and 40 %, respectively; therefore, the two solo luxR may be classified as different transcriptional protein families from carR. Moreover, a complete AI-2-mediated system is found in the YD25T genome. Interestingly, comparative sequence analysis of the lsr clusters in 43 other Serratia isolates, including the luxS and complete lsr gene cluster in the genome sequence, also revealed the consistent arrangement order of ORF with the one in YD25T. Specifically, the lsr genes are all in one polycistron in other Enterobacteriaceae bacteria, such as S. typhimurium [78], E. coli [79], and E. fergusonii [33], while lsrBACDEF, lsrGK, and lsrR are dispersed in the Serratia genome sequence. In addition, due to the differences among genera, the sequence similarity of LsrBACDEF and LsrR of Serratia show a lower degree with their homologues in other Enterobacteriaceae bacteria. Based on phylogenetic analysis with concatenated amino acid sequences of 10 proteins (LsrABCDEFGKR and LuxS), we observed a significant correlation between the lsr operon and secondary metabolites, prodigiosin and serrawettin W2, of Serratia isolates in general. Therefore, we conclude that AI-2 regulates the biosynthesis of the two compounds. AI-2-mediated systems have been shown to regulate various secondary metabolites produced by strains of Serratia [80], and it will be interesting to study whether AI-2 is directly involved in regulating the expression of the pig and swr gene clusters in Serratia.
The 16S rRNA gene sequence of YD25T was related most closely to S. nematodiphila DSM 21420T [63] and S. marcescens LMG 2792T [2, 81]. The phylogenetic reconstruction based on 16S rRNA was of very low resolution due to the high interspecies similarity of the 16S rRNA gene sequences. However, the biochemical, physiological and morphological features of these three strains distinguishes YD25 T from the other two strains. In addition, the results of OrthoANI, GGDC, and MLSA analysis based on the analysis of genome sequences strongly support that YD25T is different from other species of Serratia. Moreover, phylogenetic analysis based on concatenated Lsr and LuxS proteins revealed that YD25T forms an independent branch and is clearly distant from the strains that solely produce either prodigiosin or serrawettin W2. Combining both classical taxonomic analysis and genomic sequence analysis, we propose that YD25T is a novel species of the genus Serratia, to which the name Serratia surfactantensis sp. nov. is proposed.
Conclusions
The phylogenetic, genotypic and phenotypic data support that strain Serratia sp. YD25T (=CCTCC AB 2015384; =KCTC 42987) is a member of a novel and previously uncharacterized Serratia species; the name Serratia surfactantensis sp. nov. is proposed. The genomic sequence and metabolite analysis of S. surfactantfaciens YD25T indicates that this strain can be further explored for the production of useful metabolites. S. surfactantfaciens YD25T can simultaneously produce both prodigiosin and serrawettin W2, therefore unveiling its genomic sequence benefits using this novel species as a model system for studying the biosynthesis regulation of these two useful compounds by the QS system.
Acknowledgements
The authors are grateful for the helpful discussions with Professor Qianli An, Institute of Biotechnology, Zhejiang University, China.
Funding
This work was supported by grants from the National Natural Science Foundation of China (No. 31400057, 31600038), the Postdoctoral Science Foundation of China (No. 2016 M592745), the Natural Science Basic Research Plan in Shaanxi Province of China (No. 2014JM3067), the Postdoctoral Funds in Shaanxi Province of China, the Innovative Experiment Projects of Educational Ministry of China for Undergraduate (No. 201610718019, 201610718021), and the Projects of Science and Technology Program of China National Tobacco Corporation, Fujian Branch (No.2012049).
Availability of data and materials
This whole genome project has been deposited at DDBJ/EMBL/GenBank under accession number LNZI00000000. The S. surfactantfaciens YD25T has been preserved in the Korean Collection for Type Cultures and China Center for Type Culture Collection; the strain numbers are CCTCC AB 2015384 and KCTC 42987, respectively.
Authors’ contributions
Conceived and designed the experiments: YS, CS, and ZL. Collected the soil sample: ZJX and LJL. Performed the experiments: CS, ZJX, TJC, FC, XRW, YDZ, and FRZ. Analyzed the data: ZJX, YBL, and CS. Wrote the paper: CS, YS and XQZ. All authors edited, read and approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Consent for publication
Not applicable.
Ethics approval and consent to participate
Not applicable.
Abbreviations
- A
Adenylation domain
- AHL
N-acyl-L-homoserine lactone
- AI-1
Autoinducer-1
- AI-2
Autoinducer-2
- ANI
Average nucleotide identity
- AT
Acyltransferase
- C
Condensation domain
- CFS
Cell-free supernatant sample
- DDH
DNA-DNA hybridization
- ESI
Electrospray ionization
- FA
Fatty acid
- GGDC
Genome to genome distance calculator
- KR
Keto reductase
- KS
Ketosynthase
- MLSA
Multi-locus sequence analysis
- MLST
Multi-locus sequence typing
- NCBI
National Center for Biotechnology Information
- NRPS
Non-ribosomal peptide synthetases
- ORF
Open reading frame
- PKS
Hybrid polyketide synthases
- QS
Quorum sensing
- T
Thiolation domain
- TE
Thioesterase domain
Additional files
The morphological characteristics of YD25T. (A) A circular irregular margin morphology of YD25T after incubation for 48 h at 30 °C on KB agar; (B) Scanning electron micrograph of YD25T, Bar, 100 nm; (C) Swimming colony of YD25T after incubation for 48 h at 30 °C on 0.3 % agar KB swimming plate; (D) Swarming colony of YD25T after incubation for 48 h at 30 °C on 0.5 % agar LB swarming plate. (DOCX 434 kb)
Antagonistic activities of YD25T against fungi. (A) Visualization of the YD25T-E. turcicum, YD25T-F. oxysporum, YD25T-A. alternata and YD25T-C. sativus confrontation assays, 3-7 days after inoculation of fungi in the presence of YD25T or in its absence. (B) The biofilm culture assay of M. luteus co-cultured with and without YD25T at 30 °C; (C) The biofilm culture assay of R. solanacearum co-cultured with and without YD25T at 30 °C. Numbers show an average of three replications, and error bars show standard errors of the means. (DOCX 826 kb)
1H and 13C NMR chemical shift assignments of sw-5 (serrawettin W2). (DOCX 16 kb)
Circular map of the YD25T chromosome. From outside to the center: genes on forward strand, genes on reverse strand, GC content, GC skew. (DOCX 297 kb)
Deduced functions and homologues of the gene products from the pig clusters. The relatedness of each YD25T pig protein to its homologue in S. marcescens ATCC 274, S. plymuthica AS 13, and Serratia sp. ATCC 39006 is shown in the comparison of Serratia pig protein percentage sequence identity column. (DOCX 15 kb)
Deduced functions and homologues of the gene products from the lsr clusters. The relatedness of each YD25T Lsr protein to its homologue in Salmonella typhimurium LT2, E. coli K-12, and E. fergusonii ATCC 35469 is shown in the comparison of Lsr protein percentage sequence identity column. (DOCX 15 kb)
Phylogenetic analyses of concatenated gene sequences in the lsr operon and luxS. Bootstrap analysis (1000 resamplings) was used to evaluate the topology of the NJ tree. Bar, 0.01 substitutions per nucleotide position. All sequences were retrieved from the NCBI database or in published genome projects. Boxes represent the strains in which the biosynthetic gene cluster of prodigiosin (red) or serrawettin W2 (blue) are observed in the genome. (DOCX 241 kb)
The less than 98.7 % 16S rRNA gene sequence similarities between YD25T and type strains of phylogenetically related species. (DOCX 12 kb)
Neighbor-joining tree showing the phylogenetic relationships of YD25T and phylogenetically related reference strains based on 16S rRNA gene sequences. Bootstrap analysis (1000 resamplings) was used to evaluate the topology of the NJ tree, and the bootstrap values > 70 % are displayed at branch points. Bar, 0.005 substitutions per nucleotide position. (DOCX 155 kb)
16S rRNA, rpoB, atpD, gyrB and infB gene sequence similarities between YD25T and type strains of phylogenetically related species. (DOCX 13 kb)
Neighbor-joining tree showing the phylogenetic relationships of YD25T and phylogenetically related reference strains based on rpoB gene sequences. Bootstrap analysis (1000 resamplings) was used to evaluate the topology of the NJ tree, and the bootstrap values > 50 % are displayed at branch points. Bar, 0.01 substitutions per nucleotide position. (DOCX 148 kb)
Description of Serratia surfactantfaciens sp. nov. (DOCX 16 kb)
Contributor Information
Yan Sun, Phone: 86-29-85310546, Email: sunyan@snnu.edu.cn.
Zhi Li, Phone: 86-29-85310581, Email: lizhi@snnu.edu.cn.
References
- 1.Forsythe SJ, Abbott SL, Pitout J. Klebsiella, Enterobacter, Citrobacter, Cronobacter, Serratia, Plesiomonas, and other Enterobacteriaceae. 2015. [Google Scholar]
- 2.Grimont F, Grimont PA: The genus serratia. In: The prokaryotes. Springer; 2006:219-244. http://www.annualreviews.org/doi/abs/10.1146/annurev.mi.32.100178.001253?journalCode=micro.
- 3.Domik D, Thürmer A, Weise T, Brandt W, Daniel R, Piechulla B: A terpene synthase is involved in the synthesis of the volatile organic compound sodorifen of Serratia plymuthica 4Rx13. Front. Microbiol. 2016;7:737. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4872519/. [DOI] [PMC free article] [PubMed]
- 4.Petersen LM, Tisa LS. Friend or foe? A review of the mechanisms that drive Serratia towards diverse lifestyles. Can J Microbiol. 2013;59(9):627–640. doi: 10.1139/cjm-2013-0343. [DOI] [PubMed] [Google Scholar]
- 5.Matsuyama T, Tanikawa T, Nakagawa Y: Serrawettins and other surfactants produced by Serratia. In: Biosurfactants. Springer; 2011:93-120. http://link.springer.com/chapter/10.1007%2F978-3-642-14490-5_4.
- 6.Gerc AJ, Song L, Challis GL, Stanley-Wall NR, Coulthurst SJ. The insect pathogen Serratia marcescens Db10 uses a hybrid non-ribosomal peptide synthetase-polyketide synthase to produce the antibiotic althiomycin. PLoS One. 2012;7(9):e44673. doi: 10.1371/journal.pone.0044673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Wilf NM, Salmond GP. The stationary phase sigma factor, RpoS, regulates the production of a carbapenem antibiotic, a bioactive prodigiosin and virulence in the enterobacterial pathogen Serratia sp. ATCC 39006. Microbiology. 2012;158(3):648–658. doi: 10.1099/mic.0.055780-0. [DOI] [PubMed] [Google Scholar]
- 8.Stankovic N, Senerovic L, Ilic-Tomic T, Vasiljevic B, Nikodinovic-Runic J. Properties and applications of undecylprodigiosin and other bacterial prodigiosins. Appl Microbiol Biotechnol. 2014;98(9):3841–3858. doi: 10.1007/s00253-014-5590-1. [DOI] [PubMed] [Google Scholar]
- 9.Hong B, Prabhu VV, Zhang S, van den Heuvel APJ, Dicker DT, Kopelovich L, El-Deiry WS. Prodigiosin rescues deficient p53 signaling and antitumor effects via upregulating p73 and disrupting its interaction with mutant p53. Cancer Res. 2014;74(4):1153–1165. doi: 10.1158/0008-5472.CAN-13-0955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Genes C, Baquero E, Echeverri F, Maya JD, Triana O: Mitochondrial dysfunction in Trypanosoma cruzi: the role of Serratia marcescens prodigiosin in the alternative treatment of Chagas disease. Parasit Vectors 2011;4(8). https://parasitesandvectors.biomedcentral.com/articles/10.1186/1756-3305-4-66. [DOI] [PMC free article] [PubMed]
- 11.Soberón-Chávez G, Maier RM: Biosurfactants: a general overview. In: Biosurfactants. Springer; 2011:1-11. http://link.springer.com/chapter/10.1007/978-3-642-14490-5_1.
- 12.Matsuyama T, Murakami T, Fujita M, Fujita S, Yano I. Extracellular vesicle formation and biosurfactant production by Serratia marcescens. Microbiology. 1986;132(4):865–875. doi: 10.1099/00221287-132-4-865. [DOI] [Google Scholar]
- 13.Thies S, Santiago-Schübel B, Kovačić F, Rosenau F, Hausmann R, Jaeger K-E. Heterologous production of the lipopeptide biosurfactant serrawettin W1 in Escherichia coli. J Biotechnol. 2014;181:27–30. doi: 10.1016/j.jbiotec.2014.03.037. [DOI] [PubMed] [Google Scholar]
- 14.Garding A, Bhattacharya N, Claus R, Ruppel M, Tschuch C, Filarsky K, Idler I, Zucknick M, Caudron-Herger M, Oakes C. Epigenetic upregulation of lncRNAs at 13q14. 3 in leukemia is linked to the in Cis downregulation of a gene cluster that targets NF-kB. PLoS Genet. 2013;9(4):e1003373. doi: 10.1371/journal.pgen.1003373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Sun X, Shen X, Jain R, Lin Y, Wang J, Sun J, Wang J, Yan Y, Yuan Q. Synthesis of chemicals by metabolic engineering of microbes. Chem Soc Rev. 2015;44(11):3760–3785. doi: 10.1039/C5CS00159E. [DOI] [PubMed] [Google Scholar]
- 16.Matsuyama T, Kaneda K, Nakagawa Y, Isa K, Hara-Hotta H, Yano I. A novel extracellular cyclic lipopeptide which promotes flagellum-dependent and-independent spreading growth of Serratia marcescens. J Bacteriol. 1992;174(6):1769–1776. doi: 10.1128/jb.174.6.1769-1776.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Pradel E, Zhang Y, Pujol N, Matsuyama T, Bargmann CI, Ewbank JJ. Detection and avoidance of a natural product from the pathogenic bacterium Serratia marcescens by Caenorhabditis elegans. Proc Natl Acad Sci. 2007;104(7):2295–2300. doi: 10.1073/pnas.0610281104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Gerc AJ, Stanley-Wall NR, Coulthurst SJ. Role of the phosphopantetheinyltransferase enzyme, PswP, in the biosynthesis of antimicrobial secondary metabolites by Serratia marcescens Db10. Microbiology. 2014;160(8):1609–1617. doi: 10.1099/mic.0.078576-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Tanikawa T, Nakagawa Y, Matsuyama T. Transcriptional downregulator hexS controlling prodigiosin and serrawettin W1 biosynthesis in Serratia marcescens. Microbiol Immunol. 2006;50(8):587–596. doi: 10.1111/j.1348-0421.2006.tb03833.x. [DOI] [PubMed] [Google Scholar]
- 20.Soto-Cerrato V, Montaner B, Martinell M, Vilaseca M, Giralt E, Pérez-Tomás R. Cell cycle arrest and proapoptotic effects of the anticancer cyclodepsipeptide serratamolide (AT514) are independent of p53 status in breast cancer cells. Biochem Pharmacol. 2005;71(1):32–41. doi: 10.1016/j.bcp.2005.10.020. [DOI] [PubMed] [Google Scholar]
- 21.Soo P-C, Horng Y-T, Chang Y-L, Tsai W-W, Jeng W-Y, Lu C-C, Lai H-C. ManA is regulated by RssAB signaling and promotes motility in Serratia marcescens. Res Microbiol. 2014;165(1):21–29. doi: 10.1016/j.resmic.2013.10.005. [DOI] [PubMed] [Google Scholar]
- 22.Slater H, Crow M, Everson L, Salmond GP. Phosphate availability regulates biosynthesis of two antibiotics, prodigiosin and carbapenem, in Serratia via both quorum-sensing -dependent and -independent pathways. Mol Microbiol. 2003;47(2):303–320. doi: 10.1046/j.1365-2958.2003.03295.x. [DOI] [PubMed] [Google Scholar]
- 23.Rao B, Zhang LY, Sun JA, Su G, Wei D, Chu J, Zhu J, Shen Y. Characterization and regulation of the 2, 3-butanediol pathway in Serratia marcescens. Appl Microbiol Biotechnol. 2012;93(5):2147–2159. doi: 10.1007/s00253-011-3608-5. [DOI] [PubMed] [Google Scholar]
- 24.Matilla MA, Leeper FJ, Salmond GP. Biosynthesis of the antifungal haterumalide, oocydin A, in Serratia, and its regulation by quorum sensing, RpoS and Hfq. Environ Microbiol. 2015;17(8):2993–3008. doi: 10.1111/1462-2920.12839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Wilf NM, Reid AJ, Ramsay JP, Williamson NR, Croucher NJ, Gatto L, Hester SS, Goulding D, Barquist L, Lilley KS. RNA-seq reveals the RNA binding proteins, Hfq and RsmA, play various roles in virulence, antibiotic production and genomic flux in Serratia sp. ATCC 39006. BMC Genomics. 2013;14(1):1. doi: 10.1186/1471-2164-14-822. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Gristwood T, Fineran PC, Everson L, Williamson NR, Salmond GP. The PhoBR two-component system regulates antibiotic biosynthesis in Serratia in response to phosphate. BMC Microbiol. 2009;9(1):112. doi: 10.1186/1471-2180-9-112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Liu X, Jia J, Popat R, Ortori CA, Li J, Diggle SP, Gao K, Cámara M. Characterisation of two quorum sensing systems in the endophytic Serratia plymuthica strain G3: differential control of motility and biofilm formation according to life-style. BMC Microbiol. 2011;11(1):1. doi: 10.1186/1471-2180-11-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Bassler BL, Miller MB: Quorum sensing. In: The prokaryotes. Springer; 2013:495-509. http://link.springer.com/referenceworkentry/10.1007%2F978-3-642-30123-0_60.
- 29.Cude WN, Buchan A. Acyl-homoserine lactone-based quorum sensing in the Roseobacter clade: complex cell-to-cell communication controls multiple physiologies. 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Pereira CS, Thompson JA, Xavier KB. AI-2-mediated signalling in bacteria. FEMS Microbiol Rev. 2013;37(2):156–181. doi: 10.1111/j.1574-6976.2012.00345.x. [DOI] [PubMed] [Google Scholar]
- 31.Galloway WR, Hodgkinson JT, Bowden SD, Welch M, Spring DR. Quorum sensing in Gram-negative bacteria: small-molecule modulation of AHL and AI-2 quorum sensing pathways. Chem Rev. 2010;111(1):28–67. doi: 10.1021/cr100109t. [DOI] [PubMed] [Google Scholar]
- 32.Marques JC, Oh IK, Ly DC, Lamosa P, Ventura MR, Miller ST, Xavier KB. LsrF, a coenzyme A-dependent thiolase, catalyzes the terminal step in processing the quorum sensing signal autoinducer-2. Proc Natl Acad Sci. 2014;111(39):14235–14240. doi: 10.1073/pnas.1408691111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Brito PH, Rocha EP, Xavier KB, Gordo I. Natural genome diversity of AI-2 quorum sensing in Escherichia coli: conserved signal production but labile signal reception. Genome Biol Evol. 2013;5(1):16–30. doi: 10.1093/gbe/evs122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Sun S-J, Liu Y-C, Sun J, Zhu H. The dependence of quorum sensing in Serratia marcescens JG on the transcription of luxS gene. Arch Microbiol. 2015;197(5):715–721. doi: 10.1007/s00203-015-1099-y. [DOI] [PubMed] [Google Scholar]
- 35.Joyner J, Wanless D, Sinigalliano CD, Lipp EK: Direct Detection of Serratia marcescens in marine and other aquatic environments using quantitative real time PCR. Appl Environ Microbiol 2013:AEM. 02755-02713. http://aem.asm.org/content/early/2013/12/23/AEM.02755-13.short. [DOI] [PMC free article] [PubMed]
- 36.Ee R, Lim Y-L, Tee K-K, Yin W-F, Chan K-G. Quorum sensing activity of Serratia fonticola strain RB-25 isolated from an ex-landfill site. Sensors. 2014;14(3):5136–5146. doi: 10.3390/s140305136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Gastmeier P: Serratia marcescens: an outbreak experience. Front. Microbiol. 2014;5. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3944479/. [DOI] [PMC free article] [PubMed]
- 38.Ramasamy D, Mishra AK, Lagier J-C, Padhmanabhan R, Rossi M, Sentausa E, Raoult D, Fournier P-E. A polyphasic strategy incorporating genomic data for the taxonomic description of novel bacterial species. Int J Syst Evol Microbiol. 2014;64(2):384–391. doi: 10.1099/ijs.0.057091-0. [DOI] [PubMed] [Google Scholar]
- 39.Chan JZ, Halachev MR, Loman NJ, Constantinidou C, Pallen MJ. Defining bacterial species in the genomic era: insights from the genus Acinetobacter. BMC Microbiol. 2012;12(1):302. doi: 10.1186/1471-2180-12-302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Bennett JS, Jolley KA, Earle SG, Corton C, Bentley SD, Parkhill J, Maiden MC. A genomic approach to bacterial taxonomy: an examination and proposed reclassification of species within the genus Neisseria. Microbiology. 2012;158(6):1570–1580. doi: 10.1099/mic.0.056077-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Chun J, Rainey FA. Integrating genomics into the taxonomy and systematics of the bacteria and archaea. Int J Syst Evol Microbiol. 2014;64(2):316–324. doi: 10.1099/ijs.0.054171-0. [DOI] [PubMed] [Google Scholar]
- 42.Kim M, Oh H-S, Park S-C, Chun J. Towards a taxonomic coherence between average nucleotide identity and 16S rRNA gene sequence similarity for species demarcation of prokaryotes. Int J Syst Evol Microbiol. 2014;64(2):346–351. doi: 10.1099/ijs.0.059774-0. [DOI] [PubMed] [Google Scholar]
- 43.Richter M. Rosselló-Mo ra R. 2009. Shifting the genomic gold standard for the prokaryotic species definition. Proc Natl Acad Sci U S A, 2009;106(45):19126-19131. http://www.pnas.org/content/106/45/19126.short. [DOI] [PMC free article] [PubMed]
- 44.Auch AF, Klenk H-P, Göker M. Standard operating procedure for calculating genome-to-genome distances based on high-scoring segment pairs. Stand Genomic Sci. 2010;2(1):142–148. doi: 10.4056/sigs.541628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Meier-Kolthoff JP, Auch AF, Klenk H-P, Göker M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinformatics. 2013;14(1):1. doi: 10.1186/1471-2105-14-60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Glaeser SP, Kämpfer P. Multilocus sequence analysis (MLSA) in prokaryotic taxonomy. Syst Appl Microbiol. 2015;38(4):237–245. doi: 10.1016/j.syapm.2015.03.007. [DOI] [PubMed] [Google Scholar]
- 47.Mela F, Fritsche K, de Boer W, van Veen JA, de Graaff LH, van den Berg M, Leveau JH. Dual transcriptional profiling of a bacterial/fungal confrontation: Collimonas fungivorans versus Aspergillus niger. ISME J. 2011;5(9):1494–1504. doi: 10.1038/ismej.2011.29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Li P, Kwok AH, Jiang J, Ran T, Xu D, Wang W, Leung FC. Comparative genome analyses of serratia marcescens FS14 reveals its high antagonistic potential. PLoS One. 2015;10(4):e0123061. doi: 10.1371/journal.pone.0123061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Altschul SF, Madden TL, Schäffer AA, Zhang J, Zhang Z, Miller W, Lipman DJ. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 1997;25(17):3389–3402. doi: 10.1093/nar/25.17.3389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Boeckmann B, Bairoch A, Apweiler R, Blatter M-C, Estreicher A, Gasteiger E, Martin MJ, Michoud K, O'Donovan C, Phan I. The SWISS-PROT protein knowledgebase and its supplement TrEMBL in 2003. Nucleic Acids Res. 2003;31(1):365–370. doi: 10.1093/nar/gkg095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Tatusov RL, Natale DA, Garkavtsev IV, Tatusova TA, Shankavaram UT, Rao BS, Kiryutin B, Galperin MY, Fedorova ND, Koonin EV. The COG database: new developments in phylogenetic classification of proteins from complete genomes. Nucleic Acids Res. 2001;29(1):22–28. doi: 10.1093/nar/29.1.22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Blin K, Medema MH, Kazempour D, Fischbach MA, Breitling R, Takano E, Weber T: antiSMASH 2.0-a versatile platform for genome mining of secondary metabolite producers. Nucleic Acids Res. 2013;3:gkt449. http://nar.oxfordjournals.org/content/early/2013/06/03/nar.gkt449.short. [DOI] [PMC free article] [PubMed]
- 53.Delcher AL, Bratke KA, Powers EC, Salzberg SL. Identifying bacterial genes and endosymbiont DNA with Glimmer. Bioinformatics. 2007;23(6):673–679. doi: 10.1093/bioinformatics/btm009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Larkin MA, Blackshields G, Brown N, Chenna R, McGettigan PA, McWilliam H, Valentin F, Wallace IM, Wilm A, Lopez R. Clustal W and Clustal X version 2.0. Bioinformatics. 2007;23(21):2947–2948. doi: 10.1093/bioinformatics/btm404. [DOI] [PubMed] [Google Scholar]
- 55.Tamura K, Stecher G, Peterson D, Filipski A, Kumar S: MEGA6: molecular evolutionary genetics analysis version 6.0. Mol Biol Evol. 2013;30(12):2725–2729. http://mbe.oxfordjournals.org/content/30/12/2725.short. [DOI] [PMC free article] [PubMed]
- 56.Saitou N, Nei M. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol. 1987;4(4):406–425. doi: 10.1093/oxfordjournals.molbev.a040454. [DOI] [PubMed] [Google Scholar]
- 57.Guindon S, Gascuel O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst Biol. 2003;52(5):696–704. doi: 10.1080/10635150390235520. [DOI] [PubMed] [Google Scholar]
- 58.Brosius J, Palmer ML, Kennedy PJ, Noller HF. Complete nucleotide sequence of a 16S ribosomal RNA gene from Escherichia coli. Proc Natl Acad Sci. 1978;75(10):4801–4805. doi: 10.1073/pnas.75.10.4801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Kim O-S, Cho Y-J, Lee K, Yoon S-H, Kim M, Na H, Park S-C, Jeon YS, Lee J-H, Yi H. Introducing EzTaxon-e: a prokaryotic 16S rRNA gene sequence database with phylotypes that represent uncultured species. Int J Syst Evol Microbiol. 2012;62(3):716–721. doi: 10.1099/ijs.0.038075-0. [DOI] [PubMed] [Google Scholar]
- 60.Lee I, Kim YO, Park S-C, Chun J: OrthoANI: An improved algorithm and software for calculating average nucleotide identity. Int J Syst Evol Microbiol. 2015;66:1100-1103. http://ijs.microbiologyresearch.org/content/journal/ijsem/10.1099/ijsem.0.000760. [DOI] [PubMed]
- 61.Widdel F, Bak F, Balows A, Trüper H, Dworkin M, Harder W, Schleifer K. The Prokaryotes. A handbook on the biology of bacteria: ecophysiology, isolation, identification, applications. 1992. [Google Scholar]
- 62.Wattanavanitchakorn S, Prakitchaiwattana C, Thamyongkit P. Rapid and simple colorimetric method for the quantification of AI-2 produced from Salmonella Typhimurium. J Microbiol Methods. 2014;99:15–21. doi: 10.1016/j.mimet.2014.01.014. [DOI] [PubMed] [Google Scholar]
- 63.Zhang C-X, Yang S-Y, Xu M-X, Sun J, Liu H, Liu J-R, Liu H, Kan F, Sun J, Lai R. Serratia nematodiphila sp. nov., associated symbiotically with the entomopathogenic nematode Heterorhabditidoides chongmingensis (Rhabditida: Rhabditidae) Int J Syst Evol Microbiol. 2009;59(7):1603–1608. doi: 10.1099/ijs.0.003871-0. [DOI] [PubMed] [Google Scholar]
- 64.Bhadra B, Roy P, Chakraborty R. Serratia ureilytica sp. nov., a novel urea-utilizing species. Int J Syst Evol Microbiol. 2005;55(5):2155–2158. doi: 10.1099/ijs.0.63674-0. [DOI] [PubMed] [Google Scholar]
- 65.Vaaje‐Kolstad G, Horn SJ, Sørlie M, Eijsink VG. The chitinolytic machinery of Serratia marcescens-a model system for enzymatic degradation of recalcitrant polysaccharides. FEBS J. 2013;280(13):3028–3049. doi: 10.1111/febs.12181. [DOI] [PubMed] [Google Scholar]
- 66.Perez-Tomas R, Vinas M. New insights on the antitumoral properties of prodiginines. Curr Med Chem. 2010;17(21):2222–2231. doi: 10.2174/092986710791331103. [DOI] [PubMed] [Google Scholar]
- 67.Soberón-Chávez G: Biosurfactants: from genes to applications. vol. 20: Springer Science & Business Media; 2010.
- 68.Kai M, Haustein M, Molina F, Petri A, Scholz B, Piechulla B. Bacterial volatiles and their action potential. Appl Microbiol Biotechnol. 2009;81(6):1001–1012. doi: 10.1007/s00253-008-1760-3. [DOI] [PubMed] [Google Scholar]
- 69.Kai M, Crespo E, Cristescu SM, Harren FJM, Francke W, Piechulla B. Serratia odorifera: analysis of volatile emission and biological impact of volatile compounds on Arabidopsis thaliana. Appl Microbiol Biotechnol. 2010;88(4):965–976. doi: 10.1007/s00253-010-2810-1. [DOI] [PubMed] [Google Scholar]
- 70.Li H, Tanikawa T, Sato Y, Nakagawa Y, Matsuyama T. Serratia marcescens gene required for surfactant serrawettin W1 production encodes putative aminolipid synthetase belonging to nonribosomal peptide synthetase family. Microbiol Immunol. 2005;49(4):303–310. doi: 10.1111/j.1348-0421.2005.tb03734.x. [DOI] [PubMed] [Google Scholar]
- 71.Lai H-C, Soo P-C, Wei J-R, Yi W-C, Liaw S-J, Horng Y-T, Lin S-M, Ho S-W, Swift S, Williams P. The RssAB two-component signal transduction system in Serratia marcescens regulates swarming motility and cell envelope architecture in response to exogenous saturated fatty acids. J Bacteriol. 2005;187(10):3407–3414. doi: 10.1128/JB.187.10.3407-3414.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Chung W-C, Chen L-L, Lo W-S, Kuo P-A, Tu J, Kuo C-H. Complete genome sequence of Serratia marcescens WW4. Genome Announc. 2013;1(2):e00126–00113. doi: 10.1128/genomeA.00126-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Labbate M, Zhu H, Thung L, Bandara R, Larsen MR, Willcox MD, Givskov M, Rice SA, Kjelleberg S. Quorum-sensing regulation of adhesion in Serratia marcescens MG1 is surface dependent. J Bacteriol. 2007;189(7):2702–2711. doi: 10.1128/JB.01582-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Chan XY, Chang CY, Hong KW, Tee KK, Yin WF, Chan KG. Insights of biosurfactant producing Serratia marcescens strain W2. 3 isolated from diseased tilapia fish: a draft genome analysis. Gut Pathogens. 2013;5(1):1. doi: 10.1186/1757-4749-5-29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Bionda N, Pitteloud J-P, Cudic P. Cyclic lipodepsipeptides: a new class of antibacterial agents in the battle against resistant bacteria. Future Med Chem. 2013;5(11):1311–1330. doi: 10.4155/fmc.13.86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Bionda N, Cudic P. Cyclic lipodepsipeptides in novel antimicrobial drug discovery. Croat Chem Acta. 2011;84(2):315–329. doi: 10.5562/cca1819. [DOI] [Google Scholar]
- 77.Wilf NM, Williamson NR, Ramsay JP, Poulter S, Bandyra KJ, Salmond GP. The RNA chaperone, Hfq, controls two luxR-type regulators and plays a key role in pathogenesis and production of antibiotics in Serratia sp. ATCC 39006. Environ Microbiol. 2011;13(10):2649–2666. doi: 10.1111/j.1462-2920.2011.02532.x. [DOI] [PubMed] [Google Scholar]
- 78.Taga ME, Semmelhack JL, Bassler BL. The LuxS-dependent autoinducer AI-2 controls the expression of an ABC transporter that functions in AI-2 uptake in Salmonella typhimurium. Mol Microbiol. 2001;42(3):777–793. doi: 10.1046/j.1365-2958.2001.02669.x. [DOI] [PubMed] [Google Scholar]
- 79.Blattner FR, Plunkett G, Bloch CA, Perna NT, Burland V, Riley M, Collado-Vides J, Glasner JD, Rode CK, Mayhew GF. The complete genome sequence of Escherichia coli K-12. Science. 1997;277(5331):1453–1462. doi: 10.1126/science.277.5331.1453. [DOI] [PubMed] [Google Scholar]
- 80.Van Houdt R, Givskov M, Michiels CW. Quorum sensing in Serratia. FEMS Microbiol Rev. 2007;31(4):407–424. doi: 10.1111/j.1574-6976.2007.00071.x. [DOI] [PubMed] [Google Scholar]
- 81.Vos P, Garrity G, Jones D, Krieg NR, Ludwig W, Rainey FA, Schleifer K-H, Whitman W: Bergey's manual of systematic bacteriology: volume 3: the firmicutes, vol. 3. USA: Springer Science & Business Media; 2011. https://books.glgoo.com/books?hl=zh-CN&lr=&id=0-VqgLiCPFcC&oi=fnd&pg=PR1&dq=Bergey's+manual+of+systematic+bacteriology:+volume+3:+the+firmicutes&ots=kGDeu-Kv7y&sig=4OWubZmnLe3C2mke9Rh6nJzhaUU#v=onepage&q=Bergey's%20manual%20of%20systematic%20bacteriology%3A%20volume%203%3A%20the%20firmicutes&f=false.
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
This whole genome project has been deposited at DDBJ/EMBL/GenBank under accession number LNZI00000000. The S. surfactantfaciens YD25T has been preserved in the Korean Collection for Type Cultures and China Center for Type Culture Collection; the strain numbers are CCTCC AB 2015384 and KCTC 42987, respectively.