

Abstract
Manganese (Mn) is an essential heavy metal. However, Mn’s nutritional aspects are paralleled by its role as a neurotoxicant upon excessive exposure. In this review, we covered recent advances in identifying mechanisms of Mn uptake and its molecular actions in the brain as well as promising neuroprotective strategies. The authors focused on reporting findings regarding Mn transport mechanisms, Mn effects on cholinergic system, behavioral alterations induced by Mn exposure and studies of neuroprotective strategies against Mn intoxication. We report that exposure to Mn may arise from environmental sources, occupational settings, food, total parenteral nutrition (TPN), methcathinone drug abuse or even genetic factors, such as mutation in the transporter SLC30A10. Accumulation of Mn occurs mainly in the basal ganglia and leads to a syndrome called manganism, whose symptoms of cognitive dysfunction and motor impairment resemble Parkinson’s disease (PD). Various neurotransmitter systems may be impaired due to Mn, especially dopaminergic, but also cholinergic and GABAergic. Several proteins have been identified to transport Mn, including divalent metal tranporter-1 (DMT-1), SLC30A10, transferrin and ferroportin and allow its accumulation in the central nervous system. Parallel to identification of Mn neurotoxic properties, neuroprotective strategies have been reported, and these include endogenous antioxidants (for instance, vitamin E), plant extracts (complex mixtures containing polyphenols and non-characterized components), iron chelating agents, precursors of glutathione (GSH), and synthetic compounds that can experimentally afford protection against Mn-induced neurotoxicity.
Keywords: Manganese, Manganese-transporters, Acetylcholine, Neuroprotection
Background
Manganese (Mn) is a naturally occurring heavy metal present as the fifth most abundant metal in the environment and twelfth most abundant element as a whole. Mn is essential for humans and animals and daily requirements are commonly met by an adequate diet. Legumes, rice, nuts and whole grains contain the highest levels of the metal. Mn is also found in seafood, seeds, chocolate, tea, leafy green vegetables, spices, soybean, and some fruits such as pineapple and acai. An overview of Mn content in common Mn-rich foods can be found in Table 1. The recommended daily intake of Mn for adult men is 2.3 and 1.8 mg/day for adult women [1]. For children, these values vary with age and are shown in Table 2. For ages 0 to 6 months the Institute of Medicine’s Dietary Reference Intake for Mn cites an adequate intake (AI) that reflects the observed mean Mn intake from human milk. In an earlier study, total Mn secretion in human milk was estimated to be 1.9 μg/day over the first 3 months and 1.6 μg/day over the second 3 months [2]. Based on these values, the AI is set according to average milk volume consumption (0.78 L/day). At ages 7 to 12 months, with the introduction of complementary foods, AI is increased. For ages 1 through 18 years, the AI is based on median Mn intake data obtained from the Food and Drug Administration Total Diet Study. The Dietary Reference Intake also lists 9–11 mg/day Mn as the upper tolerable limit likely to pose no risk of adverse health effects for adults, and 2–6 mg/day Mn for children, depending on the age. Only a small percentage of these amounts are absorbed from the intestine, since the gut tightly controls body Mn load and the metal is rapidly and efficiently excreted via bile as long as no hepatic disease takes place [3, 4].
Table 1.
An overview of manganese (Mn) content in food and drinks
| Food | Country of origin | Mn content (±SD) | Reference |
|---|---|---|---|
| Apple | Italy and Bucovat Nigeria |
0.95 μg/g 0.53 ppm |
[212] [213] |
| Beef meat | Xangai, China | 0.848 ± 1.10 μg/g | [214] |
| Cabbage | Egypt | 37.80 ± 1.20 μg/g | [215] |
| Cherry | Bulgaria Bucovat |
0.65 μg/g 0.80 μg/g |
[212] |
| Chicken meat | Xangai, China | 0.561 ± 4.18 μg/g | [214] |
| Chinese chive | South Korea | 1.17 ± 0.04 μg/g | [216] |
| Coffee | Bosnia and Herzegovina Brazil Lebanon Poland |
5.02 ± 0.52 μg/g 4.97 ± 0.10 μg/g 6.19 ± 0.02 μg/g 6.28 ± 1.01 μg/g |
[217] |
| Duck meat | Xangai, China | 0.54 ± 0.23 μg/g | [214] |
| Freshwater fish | Xangai, China | 0.47 to 0.96 μg/g | [214] |
| Garlic | Kosovo | 0.012 to 1.14 μg/g | [218] |
| Grape | Egypt Nigeria |
0.75 μg/g 0.08 ppm |
[212] [213] |
| Infant formula | |||
| Cow-based Soy-based |
USA | 30–50 μg/L 200–300 μg/L |
[220] |
| Lemon | Nigeria | 0.23 ppm | [213] |
| Lettuce | Egypt | 20.0 ± 1.0 μg/g | [215] |
| Marine mussels Mytilus edulis and Perna viridis |
England, USA, Sweden, Scotland, Canada, China.Hong Kong, China, Malasya, | 10 to 100 μg/g 10 to 150 μg/g |
[221] |
| Melon | Periam Turkey |
0.65 μg/g 0.85 μg/g |
[212] |
| Milk | [222] | ||
| Fresh or processed Human milk |
Pakistan – |
0.0215 ± 0.01 or 0.0166 ± 0.01 ppm 3 to 10 μg/L |
[220] |
| Mineral water | Egypt | 2.35 ± 0.16 ɳg/mL | [215] |
| Onion | Kosovo | 0.002 to 1.29 μg/g | [219] |
| Orange | Nigeria | 0.45 ppm | [213] |
| Pears | Italy Bucovat |
0.40 μg/g 0.90 μg/g |
[212] |
| Pineapple | Nigeria | 15.00 ppm | [213] |
| Plums | Chile Romania |
0.95 μg/g 1.05 μg/g |
[212] |
| Pork meat | Xangai, China | 0.602 ± 0.697 μg/g | [214] |
| Potatoes (white, red, orange-fleshed) |
Canary Islands, Spain | 2.71 ± 2.22, 2.57 ± 1.69, 1.74 ± 0.14 μg/g |
[223] |
| Radish | Kosovo | 0.038 to 1.33 μg/g | [219] |
| Rice | Australia | 24.4 μg/g | [224] |
| River water | Egypt | 2.16 ± 0.16 ɳg/mL | [215] |
| Seawater fish | Xangai, China | 0.437 to 0.953 μg/g | [214] |
| Shellfish | Xangai, China | 0.437 to 18.15 μg/g | [214] |
| Soybean | |||
| Non-transgenic samples | Brazil | 16.4 to 24.7 μg/g | [224] |
| Transgenic samples Soy extracts |
Japan Canada |
18.2 to 44.3 μg/g 16.5 ± 8.6 μg/g |
[225] [226] |
| Spinach | Egypt Kosovo South Korea |
24.60 ± 1.10 μg/g 0.006 to 1.25 μg/g 3.69 ± 0.0001 μg/g |
[215] [218] [216] |
| Strawberry | Belgium Romania |
3.65 μg/g 2.15 μg/g |
[212] |
| Tap water | Egypt | 1.74 ± 0.10 ɳg/mL | [215] |
| Tomatoes | Basque Country | 6.8 to 23.0 μg/g | [227] |
| Watermelon | Greece Roamania |
0.95 μg/g 0.90 μg/g |
[212] |
| White bread | Egypt | 3.20 ± 0.36 μg/g | [215] |
| Wild chive | South Korea | 3.06 ± 0.04 μg/g | [216] |
| Wild parsley | South Korea | 3.45 ± 0.05 μg/g | [216] |
| Wine | Greece | 2.2 to 10 mg/L | [228] |
Content expressed as interval, standard deviation (SD) or parts per million (ppm)
Table 2.
Summary of Mn adequate intake ages 0 through 18 years
| Gender | Age group | AI (mg/day of Mn) |
|---|---|---|
| – | 0–6 months | 0.003 |
| 7–12 months | 0.6 | |
| 1–3 years | 1.2 | |
| 4–8 years | 1.5 | |
| Girls | 9–13 years | 1.6 |
| 14–18 years | 1.6 | |
| Boys | 9–13 years | 1.9 |
| 14–18 years | 2.2 |
Source: Institute of Medicine’s Dietary Reference Intake for Mn
Abbreviations: AI adequate intake
The physiological concentration of Mn in the human brain is estimated to lie between 5.32 and 14.03 ng Mn/mg protein (20.0–52.8 μM Mn), whereas 15.96–42.09 ng Mn/mg protein (60.1–158.4 μM Mn) is the estimated pathophysiological threshold [5]. Mn is essential for several physiological processes participating in enzymatic reactions as a cofactor. Mn acts in gluconeogenesis as an activator of pyruvate carboxylase and in the Krebs Cycle as a cofactor for isocitrate dehydrogenase. In the antioxidant defense system, Mn is part of superoxide dismutase (SOD). Moreover, Mn is present in the central nervous system (CNS) as a cofactor for glutamine synthetase (GS), which is preferentially localized in astrocytes [6]. Mn deficiency is a rare concern. Few reports of experimental Mn deficiency have cited poor bone growth, skeletal abnormalities, ataxia, skin alterations and hypocholesterolemia [4, 7].
Mn overload may arise from an impaired or not fully developed excretion system, transporter malfunction or exposure to excessive levels of Mn by air, water, food or total parenteral nutrition (TPN). Given the similarities between Mn and iron (Fe), homeostasis of both metals is interdependent, thus the Fe status also influences Mn accumulation. This is noted in cases of anemia, for example, when low levels of Fe facilitate Mn uptake [8]. Occupational exposure is one of the main concerns for Mn intoxication and it occurs in activities involving mining, welding, battery manufacture and with the use of fungicides containing the metal in its composition, such as maneb and mancozeb [9–12]. Periods of occupational exposure of 6 months to 2 years may lead to the development of manganism. The motor and neuropsychiatric symptoms may remain even 14 years after the end of exposure to Mn [13].
The risk of Mn exposure is not limited to miners or welders. The availability of the metal in the environment, water or food containing high levels of Mn represents a source of contamination for the general population [14]. Furthermore, the levels of Mn in the atmosphere may increase secondary to the use of the gasoline additive methylcyclopentadienyl manganese tricarbonyl (MMT) [15]. Drug abuse has recently become a concern for Mn poisoning, since abusers of the injectable drug methcathinone may be exposed to contaminating Mn due to the use of potassium permanganate in the synthesis process [16]. Patients with hepatic impairment and those receiving TPN, especially newborns, are susceptible to Mn accumulation [9, 17–19]. Infants and children are particularly vulnerable to inappropriate supplementation of Mn, which in some cases may lead to hypermanganesemia, dependent upon the duration of the treatment [17, 18, 20, 21]. Additionally, Mn is present at levels considered excessive in children’s formula [17].
Mutations in the SLC30A10 gene have been reported to induce a genetic Mn overload syndrome. SLC30A10 is a Mn transporter and a recessive loss-of-function mutation in its gene causes a syndrome of movement disorder and chronic liver disease. Magnetic resonance imaging (MRI) of patients with this mutation shows Mn accumulation in the basal ganglia and white matter, even in the absence of previous exposure to high Mn levels [3, 22, 23].
The central nervous system (CNS) is the main target of Mn. Excess Mn preferentially accumulates in the basal ganglia, especially in the striatum (caudate nucleus, putamen and nucleus accumbens), globus pallidus (GP) and the substantia nigra (SN) [24, 25]. Recently, the SN pars compacta (SNpc) was identified as a site of Mn accumulation in rats exposed intraperitoneally (ip) [26]. The neurodegenerative process induced by accumulation of Mn is called manganism. Manganism is a syndrome similar to Parkinson’s disease (PD), characterized by psychiatric and cognitive deficits and motor impairment [27, 28]. Mn is also a putative environmental modifier of Huntington’s disease (HD) [29–31]. The symptoms caused by the accumulation of Mn include dystonia, bradykinesia and rigidity due to damage to dopaminergic (DAergic) neurons and gliosis [12, 32]. Manganism and PD affect different areas of the brain, which allowing for a distinction between the two syndromes. SNpc DAergic neurons are progressively lost in PD, whereas the GP is predominantly affected in manganism. Lewy bodies formation is a hallmark of PD, which is not observed in manganism. In addition, manganism is not responsive to treatment with DA precursor levodopa, a drug used in the early stages of PD. Furthermore, manganism presents with a lack of resting tremor but consistent presence of dystonia [33–35].
Mn exposure alters intracellular signaling pathways in mouse and rat striatum, as well as cell culture models. These include alterations in Akt, ERK, p38, DARPP-32 and tyrosine hydroxylase (TH) phosphorylation [36–42]. Transcription factors’ localization, such as NF-κB and NF-E2-related factor 2 (Nrf2), is affected [43, 44]. Of particular interest, Mn-induced p53 phosphorylation, as well as upregulation of p53 levels, have been shown to be important events in cellular response to Mn exposure both in vivo and in vitro, possibly contributing to neuronal apoptosis [31, 45–47]. Endoplasmic reticulum (ER) stress is another factor that may lead to Mn-induced apoptosis [48].
A proper balance of Mn levels is essential for maintenance of health and avoidance of neurotoxicity. It is thus imperative to study the regulatory mechanisms of Mn uptake as well as its molecular mechanism of toxicity. The main topics of this review will focus on Mn effects in the brain, especially mechanisms of Mn transport and disruption of neurotransmitter signaling. We will discuss the behavioral aspects of Mn intoxication and possible neuroprotective strategies.
Main text
Mechanisms of Mn uptake into the CNS
As Mn is required for multiple cellular events but becomes toxic at high levels, the intracellular Mn concentration has to be under strict control. Several mechanisms regulate Mn homeostasis in the CNS, which mainly relies on different Mn transporters. Given the similar physical properties of Fe and Mn, most transporters are able to transport both metals, which compete for binding at the plasma membrane. To date, no proteins are identified as Mn-specific transporters. The brain is protected by the blood-brain barrier (BBB) and there are primarily two ways for Mn to cross the BBB and reach the brain for its function, discussed below.
Membrane localized Mn importers
Membrane importers are the primary route of Mn transport into the CNS. These transporters include the divalent metal transporter 1 (DMT1), Zrt-like, Irt-like proteins ZIP8 (SLC39A8) and ZIP14 (SLC39A14), dopamine transporter (DAT), voltage-regulated, store-operated and ionotropic glutamate receptor Ca channels, choline transporters and the citrate transporter [49, 50]. These proteins are localized on cell membranes and are able to form a membrane pore to take up divalent Mn from the extracellular matrix. Moreover, Mn may block transient receptor potential channel (TRPC3), a receptor-operated plasma membrane channel of astrocytes that responds to ATP-induced Ca signaling, thus decreasing purinergic signaling [51].
DMT1 is the most representative and best studied one. It is also known as divalent cation transporter 1 (DCT1), natural resistance-associated macrophage protein 2 (NRAMP 2) or solute carrier family 11 member 2 (SLC11A2). Gunshin et al. (1997), first cloned and characterized DMT1 with a wide range of substrates, including Fe2+, Zn2+, Mn2+, Cu2+, Co2+, Cd2+, Ni2+ and Pb2+ [52]. Garrick et al. (2006), showed that Mn is DMT1 preferred substrate, with the following transport affinity (reflecting transport efficacy): Mn > Cd > Fe > Pb ~ Co ~ Ni > Zn [53]. Thus, although Fe has also been linked to PD pathology, Mn might play a more prominent role in this disease given its higher affinity for DMT1. In the brain, DMT1 is highly expressed in the basal ganglia, including SN, GP, hypothalamic nucleus and striatum [54–56], rendering these regions more susceptible to Mn accumulation and toxicity. DMT1 regulates Mn influx into neurons by two ways. One is via a direct transport mechanism whereby the membrane-localized DMT1 opens up a pore and allows the extracellular divalent Mn to enter neurons. The other way is via a transferrin (Tf)-dependent process, which will be discussed next.
Transferrin (Tf) and transferrin receptor (TfR)
While the majority of Mn in the body is in the divalent oxidation state, there is a small amount of trivalent Mn, which is not a substrate for the above referenced importers. Tf/TfR facilitates Mn3+ influx into the CNS from the blood stream [57]. Tf is synthesized in the liver and then released into the blood [58]. Mn exposure increases the expression of TfR by enhancing the binding of iron regulatory proteins (IRPs) to iron responsive element-containing RNA in vitro [59]. TfR is a membrane protein with high affinity for Mn, which is expressed in neurons, microglia, astrocytes and the endothelial cells of the BBB [60]. When TfR recognizes and binds to Tf, the cell membrane expands inwardly and forms an endocytic vesicle, which brings in the Mn [67, 74]. Mn3+ is a stronger oxidizing agent than Mn2+ and it may cause severe oxidative stress. Ferrireductase reduces Mn3+ into Mn2+, which is released into the cytosol by DMT1 localized on the endosomal membrane [50].
Mn export in the CNS
Efflux plays a fundamental role in regulating intracellular concentrations of Mn in the CNS. Compared with Mn import, efflux of Mn is less studied, partially due to limited proteins identified in Mn export. However, with the recent discovery of four proteins facilitating Mn export, the role of Mn export has begun to be elucidated. These four proteins include ferroportin (Fpn), SLC30A10 (solute carrier family 30 member 10), secretory pathway Ca2+-ATPase 1 (SPCA1) and ATPase 13A2 (ATP13A2 or PARK9). Among them, Fpn and SLC30A10 are able to directly export cytosolic Mn out of neurons, while SPCA1 and ATP13A2 indirectly regulate Mn efflux through the Golgi apparatus and lysosome, respectively. Together, these proteins maintain Mn homeostasis in the CNS and mutations in them have been associated with certain diseases.
Membrane localized Mn exporters
Currently, these exporters include Fpn and SCL30A10. Fpn was the first known Mn exporter, however, it was first identified as a Fe exporter. And that is why it is also known as iron-regulated transporter 1 or solute carrier family 40 member 1 (SLC40A1). In the brain, Fpn has been found in neurons, astrocytes, the endothelial cells of the BBB, oligodendrocytes, the choroid plexus and ependymal cells [61]. Fpn expression levels are increased in mice and human embryonic kidney cells in the presence of Mn [62]. Xenopus laevis oocytes expressing human Fpn showed lower intracellular Mn and higher extracellular Mn [63]. Although these results indicate Fpn may play an important role on Mn homeostasis in the CNS, a direct study to investigate brain Mn levels in human or animal models carrying Fpn mutations has not been reported yet.
Interestingly, the recently identified SLC30A10 has been well known to play a critical role in regulating CNS Mn homeostasis. Currently, it is the only known protein associated with the first hereditary or familial form of Mn-induced parkinsonism. People carrying mutations in SLC30A10 suffer from hypermanganesemia with dystonia, polycythemia and hepatic cirrhosis [22, 64, 65]. The patients have ~10-fold increase in blood Mn levels and magnetic resonance imaging (MRI) studies show high levels of Mn accumulated in the basal ganglia without a history of exposure to elevated Mn from environmental or occupational sources [66]. The mechanisms by which mutations in SLC30A10 mediate Mn accumulation were recently characterized in rat-derived differentiated γ-aminobutyric acid (GABA) ergic AF5 cells, primary mice midbrain neurons and C. elegans. Leyva-Illades, Chen et al. (2014), found that wild type (WT) SLC30A10 is localized on the cell membrane, while 5 mutant transporters are all trapped in the endoplasmic reticulum (ER) or in the cytoplasm [67]. While the WT protein is able to protect from Mn induced DAergic neurodegeneration and cell toxicity, the mislocalization deprives these mutants of this essential efflux with ensuing retention of high Mn concentrations in the plasma.
Mn efflux mediated by SPCA1 and ATP13A2
SPCA1 is a Golgi-localized Ca/Mn ion pump, which belongs to the P-type ATPase family, with highest expression in keratinocytes but also in other tissues including liver and brain [68]. In HeLa cells, SPCA1 is required for transport of Mn into the Golgi, followed by secretion via exocytosis as a bona-fide Mn efflux pathway [69].
ATP13A2 (PARK9) is a transmembrane cation transporting ATPase localized on the membrane of vacuoles and lysosomes [67]. ATP13A2 has been associated with early-onset parkinsonism and Kufor-Rakeb syndrome [70–72]. In primary rat neurons, ATP13A2 levels were increased in the presence of excess Mn, while expression of wild-type ATP13A2 lowered intracellular Mn levels and prevented Mn-induced neuronal death [73].
Despite the evidence in cell culture studies, the role of SPCA1 and ATP13A2 in mediating Mn efflux in the CNS remains unclear. MRI studies to investigate Mn accumulation in the brain of patients or animal models carrying mutations in SPCA1 or ATP13A2 are needed to validate the results from the cell culture studies. The most studied Mn importers and exporters are summarized in Table 3.
Table 3.
Transporters and their roles in Mn uptake and efflux
| Transporter | Localization | Role in Mn transport | Associated pathologies | Reference | |
|---|---|---|---|---|---|
| Mn importers | DMT-1 | Highly expressed in the basal ganglia | Transports both Mn and Fe and a range of other cations. | Increased expression of DMT1 has been found in the SNpc of PD patients. Alterations in DMT1 are associated with spinal onset amyotrophic lateral sclerosis, AD onset in males, iron anaemia and restless legs syndrome | [3, 50] |
| ZIP8 and ZIP14 | Apical surface of various cell types | Regulation of Mn homeostasis (in duodenum, liver, brain, lungs, and kidney) and transfer of Mn, Fe, Zn, and Cd into the cells | ZIP 8 and −14 facilitate Cd accumulation, a non-essential toxic metal. MT-null Cd-resistant cells have been found to exhibit suppressed expression of both ZIP8 and ZIP14, suggesting that the down-regulation of both contributes to the decrease in Cd and Mn uptake | [229–231] | |
| DAT | Neurons of SNpc, GP and striatum | Reuptake of dopamine into presynaptic vesicles. Also shown to transport Mn | Patients chronically exposed to Mn display decreased DAT density and activity. DAT knockout mice exposed to Mn accumulate significantly less Mn in the striatum compared to WT | [50, 232] | |
| Ca channels | Plasma membrane | Voltage-regulated, store-operated Ca2+ channels as well as ionotropic glutamate receptors also facilitate Mn uptake into the brain | The number of known ion channel diseases (channelopathies) has increased dramatically and include cystic fibrosis, Bartter syndrome and epilepsy | [233, 234] | |
| Choline transporter | Plasma membrane | Choline uptake was found to be significantly inhibited in the presence of Cd and Mn, but not Cu or Al | Prenatal choline deficiency is associated with increased choline transporter mRNA expression in the septum and hippocampus of rats as a compensatory mechanism for acetylcholine synthesis | [235, 236] | |
| Citrate transporter | Plasma membrane | Mn citrate represents the major non-protein-bound species of Mn to enter the brain at the BBB. The influx transfer coefficient for Mn citrate was shown to be greater than that of Mn2 + alone and Tf–Mn3+ | Defects in SLC25A1, a mitochondrial citrate carrier, were identified to cause combined D-2- and L-2-hydroxyglutaric aciduria | [237, 238] | |
| Tf/TfR | Tf in plasma and TfR in the membrane of neurons, microglia, astrocytes and the endothelial cells of the BBB | Tf/TfR facilitates Mn3+ influx into the CNS from the blood stream | Polymorphisms in TfR gene have been correlated with increased risk of age related macular degeneration (AMD) | [57, 237, 239, 240] | |
| Mn exporters | Fpn | Transmembrane, expressed in the duodenum, liver, spleen, intestine, endothelial cells of the BBB, neurons, oligodendrocytes, astrocytes, choroid plexus and ependymal cells | Increased Fpn expression in HEK293 cells is associated with decreased intracellular Mn concentration and attenuated cytotoxicity | Mutations in Fpn cause type IV hemochromatosis, commonly known as Fpn disease, characterized by Fe accumulation in reticuloendothelial macrophages | [50, 62, 63] |
| SLC30A10 | Cell surface-localized. Present in basal ganglia and liver | Mediates Mn efflux from cells | Mutations in SLC30A10 that impair Mn export induce hypermanganesemia, dystonia, and polycythemia with a variable degree of hepatic dysfunction | [22, 23] | |
| SPCA1 | Mainly in Golgi membrane of keratinocytes, liver and brain | Imports Mn2+ from the cytosol to the Golgi lumen | Monoallelic mutations in SPCA1 are known to cause Hailey-Hailey disease, a blistering skin disorder. Complete loss of function is thought to be unviable | [3, 241] | |
| ATP13A2 or PARK9 | Transmembrane, localized on the membrane of vacuoles and lysosomes | Cation transporting ATPase. Shuttles cations across lysosomal membrane | ATP13A2 mutations have been associated with early-onset parkinsonism and Kufor-Rakeb syndrome. | [242–244] |
Abbreviations: AD Alzheimer’s disease, ATP13A2 ATPase type 13A2, DAT dopamine transporter, DMT1 divalent metal transporter 1, Fpn ferroportin, MT, metalothionein, SLC solute carrier, SPCA1 secretory pathway Ca2 + −ATPase isoform 1, Tf transferrin, TfR transferrin receptor
Recently, a high throughput screening approach was carried out to identify small molecules responsible for intracellular regulation of Mn homeostasis at physiologically relevant levels. It’s suggested that intracellular Mn levels are actively controlled by the cell and not exclusively by the BBB or blood-cerebrospinal fluid barrier. Furthermore, mechanisms regulating Mn content might be developmentally regulated in DAergic neurons reflecting the changing physiological demand [74].
Mn and the cholinergic system
Mn-induced alterations in behavioral patterns, namely motor incoordination or emotional and cognitive dysfunction, which observed in both patients and/or animal models, are associated with neurotransmitter metabolism disruption. Impaired neurotransmitter signaling may occur via diverse mechanisms, such as neurotransmitter release inhibition, alterations in neurotransmitter clearance from the synaptic cleft, or modulation of receptor levels or activity. The main neurotransmitter system studied in Mn neurotoxicity is the dopaminergic (DAergic) system [24, 75]; several studies have also described Mn’s effects on the GABAergic [76] and glutamatergic systems [77–82].
Mn at neurotoxic levels also affects the cholinergic system. Acetylcholine (ACh) is an important excitatory neurotransmitter both in the central and peripheral nervous system, modulating essential cognitive functions, such as learning, memory and locomotion. Given the scarcity of attention devoted to this system, we will focus next on Mn’s effects and cholinergic dysfunction [83–86].
The cholinergic system encompasses the neurotransmitter ACh, the enzyme that synthesizes ACh named Choline Acetyltransferase (ChaT; E.C. 2.3.1.6), the enzymes that hydrolyze ACh called cholinesterases (acetylcholinesterase-AChE; E.C. 3.1.1.7 and butyrylcholinesterase-BuChE; E.C. 3.1.1.8), by the cholinergic receptors (muscarinic and nicotinic) and by the system that reuptakes choline. Dysfunction of the cholinergic system is associated with several diseases, such as Alzheimer’s disease (AD) and myasthenia gravis. Mn effects on the cholinergic system may contribute to impairments in learning, memory and locomotion [87]. Although several symptoms of PD and manganism are largely related to effects on the DAergic system, studies suggest that the cholinergic system might play an important role in such diseases [83, 87]. Furthermore, Mn’s toxic effects might be related to an imbalance between the DAergic and cholinergic systems, predominantly in the basal ganglia [83].
ChAT is a marker of cholinergic function. A decrease in its activity leads to diminished storage and release of ACh affecting directly its function. Several reports have addressed the ability of Mn to alter ChAT activity. Numerous factors may contribute to this effect, including the age of the animals and the duration of treatment, since cholinergic neurons are exquisitely vulnerable in the developing brains [83, 84].
AChE is an important regulatory enzyme that rapidly hydrolyzes ACh at brain cholinergic synapses as well as at the neuromuscular junction [88, 89]. AChE possesses unique characteristics not found in any other enzyme, such as its active site organization and its catalytic mechanism [90–92]. AChE is extremely important in regulating brain function, development, neurite outgrowth, neuronal survival, and calcium levels [83, 93]. Various toxicological conditions that generate oxidative stress alter AChE activity, mainly its membrane bound form. Such changes in activity are commonly accompanied by clear signs of neurobehavioral alterations [83, 94, 95]. For example, an increase in the enzyme activity was observed by [95] and [96] correlating positively with thiobarbituric acid reactive substances (TBARS) production, possibly due to lipid peroxidation.
Several studies have addressed Mn’s influence on AChE activity. Table 4 summarizes the source of the enzyme and Mn’s effect on its activity. It is important to emphasize that Mn effects in biological systems depend on the routes of exposure, dose, age, period of exposure, environmental factors and nutritional state [83, 87, 94, 97–100].
Table 4.
Effects of Manganese (Mn) exposure on AChE activity in different experimental protocols
| Model | Tissue | Administration route and dose | Effect on AChE activity | Reference |
|---|---|---|---|---|
| Adult rats | Whole Brain | Intraperitoneal-acute (10–15 mg/kg) and chronic (mg/kg) | Increase | [100] |
| Adult rats | Cerebellum | Oral via by drinking water, 30 days (20 mg/ml) | Increase | [189] |
| Rats from 21–74 days | Brain | Oral via by food, for 53 days, chronic exposure (500 mg/kg) | Increase | [94] |
| Adult mice | Hypothalamus, pons, cerebellum, striatum, medulla, cerebral cortex and hippocampus. | Chronic treatment by oral via from conception to 60 days old | No alterations | [98] |
| 0-adult rats | Cerebellum and striatum | Oral via by drinking water, 60 days (20 mg/ml) | Increase | [97] |
| Adult rats | Brain | Intraperitoneal, 7 days (50 mg/kg) | Increase | [99] |
| Adult rats | Whole Brain | Intraperitoneal-(25 mg/kg) in 4 and 8 doses | Decrease | [84] |
ACh binds to two types of cholinergic receptors: the ionotropic family of nicotinic receptors and the metabotropic family of muscarinic receptors. The nicotinic acetylcholine receptor (nAChR), at the nerve/muscle synapse, is one of the best-characterized transmitter-gated ion channels [101, 102]. The muscarinic receptors belong to the large superfamily of plasma membrane-bound G protein coupled receptors (GPCRs) [103]. The muscarinic receptor family has five known members designated M1–M5. Mn exposure can affect the binding of ACh to cholinergic receptors. For example, intranasal Mn treatment in adult mice down-regulates nicotinic acetylcholine receptors (nAChR) in the prefrontal cortex in wild-type (high Fe accumulation) Hfe+/+ and Hfe-knockout Hfe−/−animals [85]. However, in other studies no alterations were found in the binding or density of cholinergic receptors. Chronic administration of MnCl2 (5 mg Mn/kg body weight/day) for 9 weeks, did not affect the [3H]-quinuclidinyl benzilate binding to muscarinic cholinergic receptors in mouse brain [104]. No changes in the muscarinic receptor density (Bmax) and the dissociation constant (Kd) of 3H-QNB in the different mouse brain regions was observed after daily ip injections of MnCl2 (5 mg Mn/kg) for 9 weeks [105]. Finally, the density of muscarinic receptors in monkeys remained unchanged after Mn exposure for 26 months at a dose comparable to what workers might inhale in dusty environments [106]. An overview of the potential effects of Mn on cholinergic function is depicted in Fig. 1.
Fig. 1.
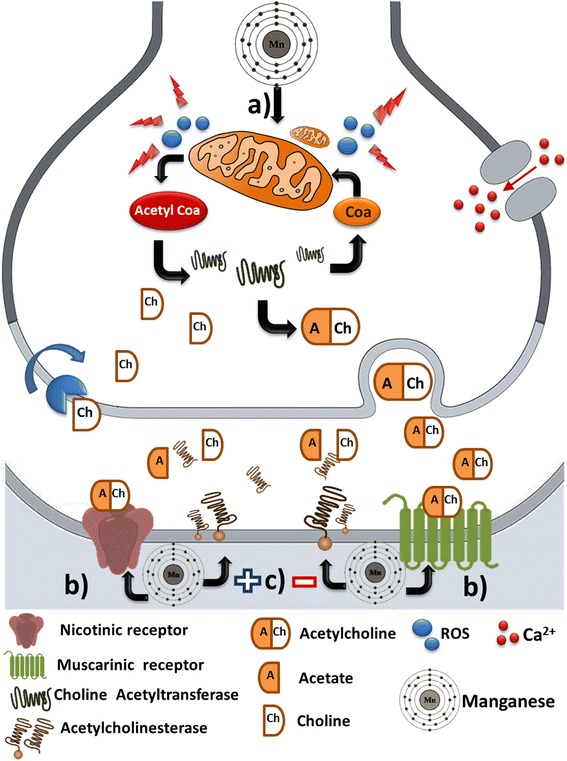
Overview of Manganese (Mn) effects on cholinergic signaling. a Mn promotes an increase in reactive oxygen species production through of mitochondrial dysfunction. In addition, Mn impairs the synthesis of precursors for acetylcholine neurotransmitter production. b Mn induces up-regulation of nicotinic and muscarinic receptors. c Mn has a controversial effect on acetylcholinesterase since it is able to increase, reduce or not alter the activity of this enzyme across diferent models of Mn exposure
Cholinergic signaling is involved in anti-inflammatory reactions. ACh is the main vagus neurotransmitter [107–109] and the efferent arm of the inflammatory reflex, now termed the cholinergic anti-inflammatory pathway. It is a highly robust mechanism for cytokine control [110]. The vagus nerve releases ACh when stimulated (either electrically or pharmacologically), inhibiting macrophage activation and release of pro-inflammatory cytokines, e.g. interleukin-6 (IL-6), tumor necrosis factor alpha (TNF-α), IL-1 and IL-18. One of the molecular mechanisms for cytokine synthesis inhibition is attributable to ACh [107, 108, 111, 112]. Accordingly, the cholinergic system controls inflammatory process and is recognized as a possible marker of low-level systemic inflammation [113–115].
Behavioral consequences of Mn exposure in humans and experimental models
Mn exposure by inhalation in occupational settings
It is estimated that over one million workers in the USA perform welding as part of their job. The pipes used in heating and ventilation systems as well as industrial process piping often require welding, which is also essential for ductwork, laboratory hoods, tanks, boilers, and process vessels. Welding produces respirable fumes that may contain Mn as well as other chemicals, such as chromium, arsenic, iron and nickel. The level of Mn exposure varies dependent upon the type of welding activity performed, ranging from 0.01 to 2.0 mg/m3 [116]. In contrast, the world health organization (WHO) recommends that levels of Mn do not exceed 30 μg/m3. It has been demonstrated that the use of ventilation systems reduces these values and could be an effective approach to minimize Mn exposure [116].
Using rats to model Mn exposure via inhalation, it has been demonstrated that the inhalation route is more efficient than ingestion at delivering Mn to the brain [117]. Mn is taken up via the olfactory tract and transferred along olfactory neurons processes through the cribriform plate to synaptic junctions with olfactory bulb neurons, thus bypassing the BBB. Once in the brain, Mn can continue to traverse synapses and be transported along neuronal tracts to other sites of the brain [118, 119]. Furthermore, the accumulation of Mn in the blood after intranasal instillation is much greater than via the oral route because Mn bypasses the biliary excretion [120]. DMT-1 is important for Mn transport across the olfactory epithelium into the brain of rats and can be influenced by Fe status [121]. Other transporters may regulate Mn uptake from the olfactory epithelium. Candidates are SLC30A10 or Mn binding proteins [120]. DMT-1 also plays a role in lung uptake of inhaled Mn [122].
Several studies point to a strong correlation between occupational Mn exposure and an increased risk of PD [123]. Parkinsonian symptoms in welders attributed to Mn exposure have been reported in numerous studies. A statistically significant difference in the age of PD onset between welders (46 years of age) and a control group (63 years) [124] has been noted. Alpha-synuclein (α-Syn), the major component of Lewy bodies and the hallmark of PD, contains metal binding sites, and its activity is not yet fully understood. It has been proposed that α-Syn attenuates Mn-induced DAergic degeneration in the early stage, but after prolonged exposure, Mn promotes α-Syn aggregation [125]. In C. elegans, α-Syn attenuates Mn-induced toxicity in the background of PD-associated genes [126]. Recently, α-Syn has been proposed to act as a intracellular Mn store [127].
Because of its paramagnetic properties, Mn accumulation can be visualized using T1-weighted magnetic resonance imaging (MRI) [128]. In a study of 193 subjects exposed to welding activities from the Midwestern USA it was shown that Mn accumulates throughout the basal ganglia, with a diffused T1 signal as well as elevated blood Mn levels when compared to age and gender matched controls. However, it was found that the MRI data not always correlated with clinical symptomatology [129, 130]. This may occur because modern occupational exposure to Mn occurs at much lower levels than reported in the past, resulting in a less distinguishable clinical phonotype. Even asymptomatic welder apprentices display increased T1 signal in the basal ganglia, but when evaluated in the Grooved Pegboard (for dexterity and fine motor control) or the unified PD rating scale motor subsection 3 (UPDRS3-for parkinsonian signs such as rest and postural tremor, bradykinesia and gait disturbance), the subjects performed within the reference range [131]. Nevertheless important neuropathological alterations have been observed even in the absence of motor symptoms [129, 132, 133]. It is not clear from the clinical studies, however, whether Mn facilitates the development of PD or induces a distinct parkinsonian syndrome. Future studies should address this issue by clearly diagnosing either PD or manganism based on the known distinctions between the two diseases.
To better understand the significance of MRI findings, an ex vivo study correlated imaging with neuropathology in 19 mine workers and 10 race- and sex-matched controls from South Africa (where 80 % of the world’s Mn reserves are located). It was found an inverse relationship between the T1 intensity indices and neuronal density in the caudate and putamen, suggesting neuronal loss. The authors also noted increased microglial cell density in the basal ganglia. Based on this and their previous study [133] they propose that the pre-clinical stage of Mn-induced neurotoxicity is marked by an initial inflammatory response that may progress to astrocyte isruption and neuronal injury [132]. This would be in agreement with in vitro findings that report a 50 fold higher accumulation of Mn in astrocytes, which may alter their neurotrophic actions and contribute no neuronal injury [134–137]. Astrocytes are initially affected in manganism showing changes in the expression of glial fibrilary acidic protein (GFAP) preceding neuronal death [138]. Increased GFAP expression is observed in the striatum of rats, which indicates glial activation in response to Mn [139, 140]. Microglial cells are also affected by Mn with increased release of proinflammatory cytokines [134] and may activate astrocytes to release inflammatory mediators such as prostaglandin E2 and nitric oxide [141].
Environmental Mn exposure
Contaminated air or water pose a risk of Mn intoxication to the general population. Mn exposure from environmental sources has also been associated with a higher prevalence of Parkinsonian disturbances [142]. For instance, near foundries, Mn concentrations may reach 200–300 ng/m3, contrasting to normal levels of Mn in the air that are around 10–30 ng/m3 according to the WHO. Recently, a study by Bowler et al. (2015) was performed to assess cognitive function in adults environmentally exposed to Mn in Ohio, USA, in two towns identified as having high levels of air-Mn from industrial sources. The authors report that non-occupational environmental Mn exposure appears to be associated with lower performance on neuropsychological tests measuring a variety of cognitive functions [143].
North America’s longest operating ferromanganese refinery is located in Marietta, Ohio, USA. To address the population leading environmental public health concern, a study was conducted to evaluate children’s cognitive function. It was found that both high and low blood and hair levels of Mn could negatively impact children’s IQ, consistent with the notion that Mn is both a nutrient and a neurotoxicant. Of note, lead (Pb) and cotinine (a nicotine metabolite) were also measured in children’s blood, serum or hair since environmental exposures to toxic chemicals rarely occur isolated. Pb levels in blood of that study population were similar to mean blood Pb of children in the USA and did not influence IQ scores. Cotinine levels were significantly associated with IQ scores, demonstrating that secondhand tobacco smoke can negatively impact child cognitive function [144]. Airborne Mn also detrimentally influenced children’s postural stability in this population [145]. Mn has been identified as a developmental neurotoxicant associated with hyperactivity, lower intellectual function, impaired motor skills and reduced olfactory function in children [146, 147]. In animal models, the immature CNS is more susceptible to Mn neurotoxicity compared to the adult [148] and experimental evidence suggests that exposure to this metal during development can affect neurological function in adulthood [139, 140, 149, 150].
The presence of excessive Mn levels in drinking water has been associated with poorer memory and attention [14], and hyperactive behavior [151] in school-aged children. Consumption of water containing elevated Mn levels had adverse effects on 10-year-old children’s cognitive function [152]. Children exposed to elevated airborne Mn in an area close to a ferromanganese alloy plant in Brazil presented lower I.Q., impairment of verbal skills [153] and lower neuropsychological performance in tests of executive function of inhibition responses, strategic visual formation and verbal working memory [154].
Mn and parenteral nutrition
Mn is present in parenteral nutrition formulations both as an essential element but also as a contaminant, thus posing as an important source of excessive exposure to Mn. The content of Mn in TPN varies from 0.18 μmol/d (0.01 mg/d) to 40 μmol/d (2.2 mg/d) [21]. Toxicity to Mn has been observed in adults receiving >500 μg/d and in pediatric patients receiving >40 μg/kg/d. Furthermore, duration of TPN treatment is associated with increased blood and brain concentrations of Mn [155–157]. Thus, current guidelines recommend monitoring patients for Mn toxicity if they receive TPN longer than 30 days [158].
Parenteral administration bypasses the regulatory mechanisms of the gastrointestinal tract. The bioavailability of Mn in parenteral fluid is 100 %, compared to only 5 % for enteral dietary Mn. For newborns, the Mn burden derived from parenteral nutrition can be 100 times greater than human milk. Of particular importance, the hepatic mechanisms responsible for Mn excretion are not completely developed in newborns. This factor combined with the high bioavailability of the metal in TPN increases the risk of Mn overload. That is also true for patients with hepatic dysfunction [17, 18, 21, 157].
Behavioral studies of Mn intoxication
Several reports address the effects of Mn exposure on behavioral tasks [67, 139, 149, 159–170]. Some of these effects are described on Table 5. As for ChAT and AChE activity, it can be observed that the animal model, the duration of exposure and the administration route are important variables when studying behavioral parameters. Briefly, the most common tasks analyzed in the references below are: Morris water maze task (MWM) an hippocampal-dependent learning test, including acquisition of spatial memory and long-term spatial memory [171]; 8-arm radial maze paradigms to evaluate reference and working memory performance simultaneously [172]; active avoidance paradigms that utilizes the passive avoidance and active avoidance test paradigms, which assay different forms of fear-based conditioned avoidance considered to be an escape response [173]; variable delayed response (VDR) task where monkeys are trained to perform cognitive tasks while seated in a restraining chair. VDR analyzes both attentional and spatial working memory components [165]; self-ordered spatial search (SOSS) task and Five Choice serial reaction time (5-CSRT) task. The SOSS task requires animals to touch identical squares located in different spatial locations in a self-ordered sequence without returning to a previously touched square. The 5-choice serial reaction time (5-CSRT) task is a widely used test to measure multiple aspects of cognition including attention, impulsivity and perseveration [167]; The object recognition task utilizes the exploration time spent in the new and familiar objects are used as parameters to asses memory and finally the social recognition test to observe short-term memory impairments [139].
Table 5.
Effects of Manganese (Mn) on different behavioral tasks
| Experimental model | Treatment | Behavioral task | Results | Reference |
|---|---|---|---|---|
| Male Sprague-Dawley rats | Neonate rats were orally exposed to MnCl2(0, 25, 50 mg/kg/day) between PND1-21 | 8-arm radial maze paradigm | Mn-exposed males showed working memory impairment | [149] |
| Young adult male Wistar rats | One group was treated with 14.84 (low dose group), and another one with 59.36 (high dose group), mg/kg Mn given by gavage, 5 days a week for 10 weeks. | 8-arm radial maze task | MnCl2 treated groups showed, compared to control animals, a decrease in the average memory performance | [170] |
| Male Wistar rats | Single oral doses of MnCl2 (50 mg/kg) or chronic oral MnCl2 (20 or 50 mg/kg/ day) for 1 month | Active avoidance paradigm |
Single dose induced decline of the memory acquisition of an avoidance reaction in response to unconditioned and conditioned stimuli. Chronic manganese poisoning also led to significant impairment of learning processes | [168] |
| C. elegans | 1 h Mn (10 or 25 mM) exposure on L1 larval stage | Basal slowing response | Mn-exposed worms had an impaired basal slowing response, indicating DAergic damage. This was reversed by SLC30A10 (a cell surface-localized Mn efflux transporter) expression in DAergic neurons | [67] |
| C. elegans | 30 min Mn (50 mM) exposure on L1 larval stage | Dauer movement | In WT dauer worms, the locomotion was increased in the presence of Mn, indicating DA signaling impairment | [161] |
| Male and female Sprague-Dawley rats | Pregnant females treated with Mn (2 mg/ml) in drinking water from the first day of pregnancy until PND20. | MWM | PND 21–25: Mn-exposed females displayed memory deficits in the probe trial PND 56–60: Mn-exposed males performed significantly worse in the acquisition trial. Females exposed to Mn displayed deficits in learning and memory |
[159] |
| Three-month-old male Wistar rats | Intranasal 2-week-long MnCl2 (0.8 mg/kg body weight) | MWM | Spatial memory deficits | [160] |
| Male Sprague-Dawley rats | Intraperitoneal injection of MnCl2 15 mg/kg for 8, 12 or 18 weeks | MWM | Escaping latency and swimming distance of rats in the model groups increased, suggesting spatial learning and memory impairment | [162] |
| Male Sprague Dawley rats | Intraperitoneal injections of 0, 5, 10 and 20 mg/kg MnCl2 once daily, 5 days/ week for 18 weeks. | MWM | Mn impaired learning and memory as follow: In 6 weeks at dose 20 mg/kg. In 12 weeks at doses 10 and 20 mg/kg. In 18 weeks at doses 5, 10 and 20 mg/kg |
[163] |
| Male Sprague-Dawley rats (6 weeks of age) | Intraperitoneal injections 15 mg/kg MnCl2 daily for 8 weeks. | MWM | The escape latency in the Morris water maze test was significantly longer in the rats injected with Mn indicating worsening in spatial memory | [164] |
| Sprague-Dawley rats, 3-week- old | Intraperitoneal injections (5, 10, 20 mg/kg MnSO4) 5 days a week for 24 weeks | MWM | Mn exposure decreased the spatial learning ability in a dose- and time- dependent manner | [169] |
| Male Wistar rats | Exposed intraperitoneally to MnCl2 at doses 5, 10 or 20 mg/kg/day from PND 8–12 | Object and social recognition tasks | PND 60–65: Rats exposed to the highest Mn dose failed to recognize a familiar object when replaced by a novel object as well as to recognize a familiar juvenile rat after a short period of time, indicating cognitive impairment | [139] |
| Adult male M. fascicularis macaques, 5 to 6 years old | Mn was administered as MnSO4 for 15 mg/kg/week for 5 weeks and then 20 mg /kg/week for the remainder of the study period (12 months) | SOSS and 5-CSRT tasks |
Deficits in performance of the SOSS task began to appear by the fourth month of Mn exposure but only became consistently significantly impaired beginning at the ninth month of Mn exposure. Performance on the 5-CSRT became significantly affected by the third month of Mn exposure | [167] |
| Adult male M. fascicularis monkeys with 5 to 6 years old | MnSO4 at doses 10–15 mg/kg/week over an exposure period lasting 272 ± 17 days |
Variable delayed response task | Animals developed subtle deficits in spatial working memory and had modest decreases in spontaneous activity and manual dexterity | [166] |
| Adult male M. fascicularis macaques, 5 to 6 years old | MnSO4 at doses 15–20 mg/kg/week over an exposure period lasting 227.5 ± 17.3 days | Variable delayed response task | Animals developed mild deficits in spatial working memory, more significant deficits in non-spatial working memory and no deficits in reference memory | [165] |
Abbreviations: 5-CSRT five choice serial reaction time, DA dopamine, MWM Morris water maze, PND post-natal day, SOSS self-ordered spatial search
In C. elegans, Mn exposure has been shown to result specifically in DAergic neurodegeneration [174]. In C. elegans DAergic neurons are considered mechanosensory and any condition impairing DA signaling will affect the ability to sense or respond to changes in its environment. DA signaling plays an important role in learning and regulation of locomotor behavior, including basal slowing response, ethanol preference, area-restricted searching, habituation task/tap withdrawal response, egg laying, dauer movement, pharyngeal pumping and thrashing behaviors [175, 176]. Among these behaviors, basal slowing response is DA-specific, and other behaviors are usually controlled by DA along with other neurotransmitters, such as serotonin, glutamate or GABA, etc. To date, basal slowing response and dauer movement have been studied with Mn exposure [175, 177, 178]. Levya-Illades, Chen et al. (2014), have shown that Mn exposure resulted in decreased basal slowing response, while expression of Mn exporter SLC30A10 exclusively in DAergic neurons rescued this behavioral defect together with decreased DAergic neurodegeneration [67]. In WT dauer worms, the locomotion was increased in the presence of Mn, indicating DA signaling is damaged by Mn exposure [176]. Similarly, the movement in djr-1.2 (homolog of mammalian DJ-1) worms was increased, indicating that loss of DJ-1 function resulted in abnormal DAergic neurons.
Neuroprotective strategies against Mn
Mn-induced neurotoxicity may present in different animal models with distinct damage, depending on time of exposure, dose and route of exposure [179, 180]. In this regard, different therapeutic approaches have been studied in different models. Originally, Mn-induced parkinsonism patients were treated with levodopa, however they were unresponsive to the treatment [181, 182] possibly due to the relatively intact nigrostriatal pathway in the latter phase of the disorder [9]. Hence, other treatments have been tested. We will briefly discuss in vitro and in vivo investigations on the properties of endogenous antioxidants (for instance, vitamin E), plant extracts (complex mixtures containing polyphenols and non-characterized components), Fe chelating agents, precursors of glutathione (GSH), and synthetic compounds that can experimentally afford protection against Mn-induced neurotoxicity.
Vitamin E and GSH
Vitamin E and trolox (a hydrophilic analog of vitamin E) have been reported to protect the CNS of rodents and cultured cells from the toxic effects of Mn [183–185]. I.p. exposure of lactating rats to Mn caused striatal and hippocampal oxidative stress and motor impairments, which were prevented by trolox co-administration [183]. GSH and N-Acetylcysteine (NAC), a precursor of GSH, can also decrease the toxicity of Mn in vitro [186]; however, the protective mechanism involved in NAC and GSH has yet to be fully studied. It is likely that these compounds serve as indirect antioxidants since GSH is a substrate of glutathione peroxidase (GPx) enzymes.
Plant extracts
Plant extracts have been demonstrated to confer protection against Mn neurotoxicity after in vitro [81] and in vivo exposure in mice [187]. Acai (Euterpe oleracea) methanolic extract protected astrocytes from Mn-induced oxidative stress. The protective effects may be associated with the antioxidant and anti-inflammatory effects of its anthocyanin components [81]. Similarly, crude aqueous extracts of Melissa officinalis blunted the Mn-induced striatal and hippocampal lipid peroxidation [187]. Purified flavonoids, such as, silymarin (obtained from Silybum marianum, a plant with hepatoprotecive properties) protected neuroblastoma cells [188] and prevented Mn-induced oxidative stress in brain, liver, and kidney of rats [189–191]. Lycopene has also been reported to decrease the neurotoxicity of Mn in rats [192].
Chelating agents
Because of the chemical similarities between Mn and Fe, it is possible that the neurotoxic effects of Mn might be associated with competition with Fe for “non-redox” domains in proteins [193]. Consequently, compounds with Fe chelating properties or those interfering with the Fenton’s reaction, such as polyphenol compounds, can be of potential pharmacological importance in the treatment of Mn toxicity [194–196]. Indeed, the treatment with a calcium disodium salt of the chelator EDTA (CaNa2EDTA) reduced Mn-induced DA autooxidation in vitro [197], enhanced urinary excretion of Mn in humans [198] and reduced Mn levels in the brain and liver of Mn-exposed rats [199]. However, there is still controversy regarding the amelioration provided by this chelating therapy [200, 201].
Synthetic compounds
Synthetic molecules have also been reported to reduce Mn toxicity. For instance, several organochalcogens (i.e. organocompounds containing selenium or tellurium atoms bound to carbon) have been reported to possess antioxidant and anti-inflammatory properties [202]. The protective effects of organoselenide and telluride compounds against Mn-induced neurotoxicity, including ebselen, have been reported [184]. One proposed mechanism might be related to a direct scavenger activity against ROS produced by Mn as most of these compounds have thiol-peroxidase activity catalyzed by glutathione-peroxidase isoforms [202]. Using the complementary animal model C. elegans, it was shown that these compounds could modulate the transcription factor DAF-16 (FOXO in mammals), increasing its translocation to the nucleus. In turn, the expression of antioxidant enzymes such as superoxide dismutase increased, thus protecting the worms from Mn-induced toxicity [203, 204]. An additional proposed mechanism is the anti-inflammatory action of some of these compounds, e.g. ebselen. Consequently, in addition to counteracting free radicals and modulating gene expression, ebselen and related compounds could decrease Mn toxicity via anti-inflammatory properties. Of note, anti-inflammatory agents have been reported to decrease Mn neurotoxicity in vitro and after in vivo exposure. For instance, Santos et al. (2013) demonstrated in vitro that 5- aminosalicylic acid (5-ASA) and para-aminosalicylic acid (4-PAS) increased mitochondrial and cell viability following Mn exposure [205]. Ibuprofen, a nonsteroidal anti-inflammatory drug, protected striatal neurons from dendritic atrophy and spine loss in rats treated for 2 weeks with the drug prior to Mn exposure [184].
The indirect pro-oxidative effects of Mn have been linked to disruption of synaptic glutamate homeostasis by interfering with glutamate uptake in astrocytes [206]. The increase in extracellular glutamate can cause excitotoxicity, which is associated with oxidative stress in neurons [206]. Furthermore, Mn decreases astrocytic glutamate uptake and expression of the astrocytic glutamate/aspartate transporter (GLAST) via disruption of intracellular signaling [207]. Of potential clinical significance, estrogen and tamoxifen have been reported to increase the expression of glutamate transporters (both GLAST and GLT-1) in astrocytes, potentially decreasing Mn toxicity [77, 207–210]. Raloxifene, which is a selective estrogen receptor modulator, also attenuates the reduction of GLT-1 and GLAST expression and the glutamate uptake induced by Mn in astrocytes [211], thus confirming how promising this class of molecules might be.
Finally, preventing or reducing Mn exposure is essential. For instance, methodologies by which welding fumes generation rate and/or welding practices can be modified to reduce toxic workplace exposures should be sought. In this context, a recent study of Sriram et al. (2015) demonstrated that rats exposed by whole body inhalation to an altered welding process (parameters: voltage, current and shielding gas) showed absence of neurotoxicity when compared to the rats exposed to regular welding process [11]. Reducing Mn levels in infant milk formulas and on parenteral nutrition should also be a strategy as safety policy.
Conclusions
The interest in researching Mn toxicity has grown in the past few decades. Recent clinical studies in populations exposed to the metal via occupational or environmental sources demonstrate Mn accumulation in the brain with T1-weighted MRI. Evidence for cognitive and motor impairment, especially in children has also been presented. Furthermore, it is evidenced by the work mentioned above that the use of rodent and other complimentary models is an important tool for the study of mechanisms of Mn toxicity, focusing on Mn transport, metal homeostasis, behavioral outcomes and neuroprotective strategies. Animal models facilitate the use of different routes of exposure to Mn, as well as the use of different chemical forms of Mn, that can mimic environmental or occupational exposure. C. elegans is also an excellent tool for genetic analysis and manipulations. The availability of mutants and green fluorescent protein (GFP)-tagging makes it easy to explore a wide range of chemicals and their effects. Several effects in response to exposure to metals, especially those involving gene expression and behavior have been reported using the nematode as a model.
One of the particularities of Mn mechanism of action is that it accumulates preferentially in the basal ganglia and targets DAergic neurons. However, various studies show that Mn may also affect other neurotransmitter systems. In this context, it is important to emphasize that to better understand Mn neurotoxic effects a cross talk between DAergic and cholinergic systems seems to be important, specially when regarding the brain regions related with PD and manganism, such as striatum, where cholinergic interneurons are present. Moreover, the neurotransmission at the neuromuscular junction and how it can lead to the motor impairment observed in manganism is an area that needs further exploration.
Acknowledgments
The authors would like to thank Jassié Gutierres and Gustavo Thomé for the figure design.
Funding
This work was supported in part by grants from the National Institute of Environmental Health Sciences R01 ES10563 and R01 ES020852. MRCS received a fellowship from CNPq (Brazil) 200783/2014-9.
Availability of data and materials
Not applicable.
Authors’ contributions
TVP reviewed the literature, wrote sections of the manuscript, designed the tables, and revised the work. MRCS reviewed the literature, wrote sections of the manuscript. PC reviewed the literature, wrote sections of the manuscript. FC reviewed the literature, designed the tables and figure. DSA reviewed the literature, wrote sections of the manuscript. AB revised critically for important intellectual content. MA revised critically for important intellectual content, gave final approval of the version to be published. All authors read and approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Consent for publication
Not applicable.
Ethics approval and consent to participate
Not applicable.
Abbreviations
- ACh
Acetylcholine
- AChE
Acetylcholinesterase
- AD
Alzheimer’s disease
- AI
Adequate intake
- BBB
Blood-brain barrier
- ChaT
Choline acetyltransferase
- CNS
Central nervous system
- DA
Dopamine
- DAT
Dopamine transporter
- DMT1
Divalent metal transporter 1
- FPN
Ferroportin
- GABA
γ-aminobutyric acid
- GFAP
Glial fibrilary acidic protein
- GP
Globus pallidus
- GPx
Glutathione peroxidase
- GS
Glutamine synthetase
- GSH
Glutathione
- HD
Huntington’s disease
- MMT
Methylcyclopentadienyl manganese tricarbonyl
- NAC
N-Acetylcysteine
- nAChR
Nicotinic acetylcholine receptor
- PD
Parkinson’s disease
- SN
Substantia nigra
- SOD
Superoxide dismutase
- Tf
Transferrin
- TfR
Transferrin receptor
- TH
Tyrosine hydroxylase
- TPN
Total parenteral nutrition
- WT
Wild type
Contributor Information
Tanara V. Peres, Phone: +1 718 430 2317, Email: tanara.peres-vieira@einstein.yu.edu
Maria Rosa C. Schettinger, Email: mariachitolina@gmail.com
Pan Chen, Email: pan.chen@einstein.yu.edu.
Fabiano Carvalho, Email: fabiseco@yahoo.com.br.
Daiana S. Avila, Email: avilads1@gmail.com
Aaron B. Bowman, Email: aaron.bowman@vanderbilt.edu
Michael Aschner, Phone: +1 718 430 2317, Email: michael.aschner@einstein.yu.edu.
References
- 1.Williams M, Todd GD, Roney N, Crawford J, Coles C, McClure PR, Garey JD, Zaccaria K, Citra M: In Toxicological Profile for Manganese. Atlanta (GA); 2012: Agency for Toxic Substances and Disease Registry (ATSDR) Toxicological Profiles]. [PubMed]
- 2.Casey CE, Neville MC, Hambidge KM. Studies in human lactation: secretion of zinc, copper, and manganese in human milk. Am J Clin Nutr. 1989;49:773–785. doi: 10.1093/ajcn/49.5.773. [DOI] [PubMed] [Google Scholar]
- 3.Tuschl K, Mills PB, Clayton PT: Chapter Twelve-Manganese and the Brain. In Int Rev Neurobiol. Volume Volume 110. Edited by Kailash PB, Susanne AS: Academic Press; 2013: 277–312 [DOI] [PubMed]
- 4.Trumbo P, Yates AA, Schlicker S, Poos M. Dietary reference intakes: vitamin A, vitamin K, arsenic, boron, chromium, copper, iodine, iron, manganese, molybdenum, nickel, silicon, vanadium, and zinc. J Am Diet Assoc. 2001;101:294–301. doi: 10.1016/S0002-8223(01)00078-5. [DOI] [PubMed] [Google Scholar]
- 5.Bowman AB, Aschner M. Considerations on manganese (Mn) treatments for in vitro studies. Neurotoxicology. 2014;41:141–142. doi: 10.1016/j.neuro.2014.01.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Santamaria AB. Manganese exposure, essentiality & toxicity. Indian J Med Res. 2008;128:484–500. [PubMed] [Google Scholar]
- 7.Keen CL, Ensunsa JL, Watson MH, Baly DL, Donovan SM, Monaco MH, et al. Nutritional aspects of manganese from experimental studies. Neurotoxicology. 1999;20:213–223. [PubMed] [Google Scholar]
- 8.Fitsanakis V, Zhang N, Garcia S, Aschner M. Manganese (Mn) and Iron (Fe): Interdependency of Transport and Regulation. Neurotox Res. 2010;18:124–131. doi: 10.1007/s12640-009-9130-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Aschner M, Erikson K, Hernández E, Tjalkens R. Manganese and its Role in Parkinson’s Disease: From Transport to Neuropathology. Neuro Mol Med. 2009;11:252–266. doi: 10.1007/s12017-009-8083-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Josephs KA, Ahlskog JE, Klos KJ, Kumar N, Fealey RD, Trenerry MR, et al. Neurologic manifestations in welders with pallidal MRI T1 hyperintensity. Neurology. 2005;64:2033–2039. doi: 10.1212/01.WNL.0000167411.93483.A1. [DOI] [PubMed] [Google Scholar]
- 11.Sriram K, Lin GX, Jefferson AM, Stone S, Afshari A, Keane MJ, et al. Modifying welding process parameters can reduce the neurotoxic potential of manganese-containing welding fumes. Toxicology. 2015;328:168–178. doi: 10.1016/j.tox.2014.12.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Cersosimo MG, Koller WC. The diagnosis of manganese-induced parkinsonism. Neuro Toxicol. 2006;27:340–346. doi: 10.1016/j.neuro.2005.10.006. [DOI] [PubMed] [Google Scholar]
- 13.Bouchard M, Mergler D, Baldwin M, Panisset M, Bowler R, Roels HA. Neurobehavioral functioning after cessation of manganese exposure: A follow-up after 14 years. Am J Ind Med. 2007;50:831–840. doi: 10.1002/ajim.20407. [DOI] [PubMed] [Google Scholar]
- 14.Oulhote Y, Mergler D, Barbeau B, Bellinger DC, Bouffard T, Brodeur ME, Saint-Amour D, Legrand M, Sauve S, Bouchard MF: Neurobehavioral Function in School-Age Children Exposed to Manganese in Drinking Water. Environ Health Perspect 2014. [DOI] [PMC free article] [PubMed]
- 15.Gulson B, Mizon K, Taylor A, Korsch M, Stauber J, Davis JM, et al. Changes in manganese and lead in the environment and young children associated with the introduction of methylcyclopentadienyl manganese tricarbonyl in gasoline--preliminary results. Environ Res. 2006;100:100–114. doi: 10.1016/j.envres.2005.03.013. [DOI] [PubMed] [Google Scholar]
- 16.Stepens A, Logina I, Liguts V, Aldiņš P, Ekšteina I, Platkājis A, et al. A Parkinsonian Syndrome in Methcathinone Users and the Role of Manganese. N Engl J Med. 2008;358:1009–1017. doi: 10.1056/NEJMoa072488. [DOI] [PubMed] [Google Scholar]
- 17.Aschner JL, Aschner M. Nutritional aspects of manganese homeostasis. Mol Asp Med. 2005;26:353–362. doi: 10.1016/j.mam.2005.07.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Boggio Bertinet D, Tinivella M, Alessandro Balzola F, de Francesco A, Davini O, Rizzo L, et al. Brain Manganese Deposition and Blood Levels in Patients Undergoing Home Parenteral Nutrition. J Parenter Enteral Nutr. 2000;24:223–227. doi: 10.1177/0148607100024004223. [DOI] [PubMed] [Google Scholar]
- 19.Nagatomo S, Umehara F, Hanada K, Nobuhara Y, Takenaga S, Arimura K, et al. Manganese intoxication during total parenteral nutrition: report of two cases and review of the literature. J Neurol Sci. 1999;162:102–105. doi: 10.1016/S0022-510X(98)00289-5. [DOI] [PubMed] [Google Scholar]
- 20.Dobson AW, Erikson KM, Aschner M: Manganese Neurotoxicity. Ann N Y Acad Sci 2004, 1012:115–128. [DOI] [PubMed]
- 21.Santos D, Batoreu C, Mateus L, Marreilha Dos Santos AP, Aschner M. Manganese in human parenteral nutrition: considerations for toxicity and biomonitoring. Neurotoxicology. 2014;43:36–45. doi: 10.1016/j.neuro.2013.10.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Quadri M, Federico A, Zhao T, Breedveld Guido J, Battisti C, Delnooz C, et al. Mutations in SLC30A10 Cause Parkinsonism and Dystonia with Hypermanganesemia, Polycythemia, and Chronic Liver Disease. Am J Hum Genet. 2012;90:467–477. doi: 10.1016/j.ajhg.2012.01.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Tuschl K, Clayton Peter T, Gospe Jr Sidney M, Gulab S, Ibrahim S, Singhi P, et al. Syndrome of Hepatic Cirrhosis, Dystonia, Polycythemia, and Hypermanganesemia Caused by Mutations in SLC30A10, a Manganese Transporter in Man. Am J Hum Genet. 2012;90:457–466. doi: 10.1016/j.ajhg.2012.01.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Guilarte TR, Chen M-K, Mcglothan JL, Verina T, Wong DF, Zhou Y, et al. Nigrostriatal dopamine system dysfunction and subtle motor deficits in manganese-exposed non-human primates. Exp Neurol. 2006;202:381–390. doi: 10.1016/j.expneurol.2006.06.015. [DOI] [PubMed] [Google Scholar]
- 25.Guilarte TR, Mcglothan JL, Degaonkar M, Chen M-K, Barker PB, Syversen T, et al. Evidence for Cortical Dysfunction and Widespread Manganese Accumulation in the Nonhuman Primate Brain following Chronic Manganese Exposure: A 1H-MRS and MRI Study. Toxicol Sci. 2006;94:351–358. doi: 10.1093/toxsci/kfl106. [DOI] [PubMed] [Google Scholar]
- 26.Robison G, Sullivan B, Cannon JR, Pushkar Y: Identification of dopaminergic neurons of the substantia nigra pars compacta as a target of manganese accumulation. Metallomics 2015. [DOI] [PMC free article] [PubMed]
- 27.Bowler RM, Gysens S, Diamond E, Nakagawa S, Drezgic M, Roels HA. Manganese exposure: Neuropsychological and neurological symptoms and effects in welders. Neuro Toxicol. 2006;27:315–326. doi: 10.1016/j.neuro.2005.10.007. [DOI] [PubMed] [Google Scholar]
- 28.Bowler RM, Koller W, Schulz PE. Parkinsonism due to manganism in a welder: Neurological and neuropsychological sequelae. Neuro Toxicol. 2006;27:327–332. doi: 10.1016/j.neuro.2005.10.011. [DOI] [PubMed] [Google Scholar]
- 29.Stansfield KH, Bichell TJ, Bowman AB, Guilarte TR. BDNF and Huntingtin protein modifications by manganese: implications for striatal medium spiny neuron pathology in manganese neurotoxicity. J Neurochem. 2014;131:655–666. doi: 10.1111/jnc.12926. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Kumar KK, Goodwin CR, Uhouse MA, Bornhorst J, Schwerdtle T, Aschner M, et al. Untargeted metabolic profiling identifies interactions between Huntington’s disease and neuronal manganese status. Metallomics. 2015;7:363–370. doi: 10.1039/C4MT00223G. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Tidball AM, Bryan MR, Uhouse MA, Kumar KK, Aboud AA, Feist JE, et al. A novel manganese-dependent ATM-p53 signaling pathway is selectively impaired in patient-based neuroprogenitor and murine striatal models of Huntington’s disease. Hum Mol Genet. 2015;24:1929–1944. doi: 10.1093/hmg/ddu609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Gunter TE, Gavin CE, Aschner M, Gunter KK. Speciation of manganese in cells and mitochondria: A search for the proximal cause of manganese neurotoxicity. Neuro Toxicol. 2006;27:765–776. doi: 10.1016/j.neuro.2006.05.002. [DOI] [PubMed] [Google Scholar]
- 33.Perl DP, Olanow CW. The neuropathology of manganese-induced Parkinsonism. J Neuropathol Exp Neurol. 2007;66:675–682. doi: 10.1097/nen.0b013e31812503cf. [DOI] [PubMed] [Google Scholar]
- 34.Kwakye GF, Paoliello MM, Mukhopadhyay S, Bowman AB, Aschner M. Manganese-Induced Parkinsonism and Parkinson’s Disease: Shared and Distinguishable Features. Int J Environ Res Public Health. 2015;12:7519–7540. doi: 10.3390/ijerph120707519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Guilarte TR, Gonzales KK. Manganese-Induced Parkinsonism Is Not Idiopathic Parkinson’s Disease: Environmental and Genetic Evidence. Toxicol Sci. 2015;146:204–212. doi: 10.1093/toxsci/kfv099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Cordova FM, Aguiar AS, Jr, Peres TV, Lopes MW, Gonçalves FM, Remor AP, et al. In Vivo Manganese Exposure Modulates Erk, Akt and Darpp-32 in the Striatum of Developing Rats, and Impairs Their Motor Function. PLoS One. 2012;7:e33057. doi: 10.1371/journal.pone.0033057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Williams BB, Li D, Wegrzynowicz M, Vadodaria BK, Anderson JG, Kwakye GF, et al. Disease-toxicant screen reveals a neuroprotective interaction between Huntington’s disease and manganese exposure. J Neurochem. 2010;112:227–237. doi: 10.1111/j.1471-4159.2009.06445.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Exil V, Ping L, Yu Y, Chakraborty S, Caito SW, Wells KS, et al. Activation of MAPK and FoxO by manganese (Mn) in rat neonatal primary astrocyte cultures. PLoS One. 2014;9:e94753. doi: 10.1371/journal.pone.0094753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Posser T, Franco JL, Bobrovskaya L, Leal RB, Dickson PW, Dunkley PR. Manganese induces sustained Ser40 phosphorylation and activation of tyrosine hydroxylase in PC12 cells. J Neurochem. 2009;110:848–856. doi: 10.1111/j.1471-4159.2009.06185.x. [DOI] [PubMed] [Google Scholar]
- 40.Zhang D, Kanthasamy A, Anantharam V, Kanthasamy A. Effects of manganese on tyrosine hydroxylase (TH) activity and TH-phosphorylation in a dopaminergic neural cell line. Toxicol Appl Pharmacol. 2011;254:65–71. doi: 10.1016/j.taap.2010.03.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.McDougall SA, Der-Ghazarian T, Britt CE, Varela FA, Crawford CA. Postnatal manganese exposure alters the expression of D2L and D2S receptor isoforms: Relationship to PKA activity and Akt levels. Synapse. 2011;65:583–591. doi: 10.1002/syn.20877. [DOI] [PubMed] [Google Scholar]
- 42.Crittenden PL, Filipov NM. Manganese modulation of MAPK pathways: effects on upstream mitogen activated protein kinase kinases and mitogen activated kinase phosphatase-1 in microglial cells. J Appl Toxicol. 2011;31:1–10. doi: 10.1002/jat.1552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Filipov NM, Seegal RF, Lawrence DA. Manganese Potentiates In Vitro Production of Proinflammatory Cytokines and Nitric Oxide by Microglia Through a Nuclear Factor kappa B–Dependent Mechanism. Toxicol Sci. 2005;84:139–148. doi: 10.1093/toxsci/kfi055. [DOI] [PubMed] [Google Scholar]
- 44.Li H, Wu S, Shi N, Lian S, Lin W. Nrf2/HO-1 pathway activation by manganese is associated with reactive oxygen species and ubiquitin–proteasome pathway, not MAPKs signaling. J Appl Toxicol. 2011;31:690–697. doi: 10.1002/jat.1654. [DOI] [PubMed] [Google Scholar]
- 45.Ma X, Han J, Wu Q, Liu H, Shi S, Wang C, et al. Involvement of dysregulated Wip1 in manganese-induced p53 signaling and neuronal apoptosis. Toxicol Lett. 2015;235:17–27. doi: 10.1016/j.toxlet.2014.12.019. [DOI] [PubMed] [Google Scholar]
- 46.Shi S, Zhao J, Yang L, Nie X, Han J, Ma X, et al. KHSRP Participates in Manganese-Induced Neurotoxicity in Rat Striatum and PC12 Cells. J Mol Neurosci. 2015;55:454–465. doi: 10.1007/s12031-014-0367-7. [DOI] [PubMed] [Google Scholar]
- 47.Wan C, Ma X, Shi S, Zhao J, Nie X, Han J, et al. Pivotal roles of p53 transcription-dependent and -independent pathways in manganese-induced mitochondrial dysfunction and neuronal apoptosis. Toxicol Appl Pharmacol. 2014;281:294–302. doi: 10.1016/j.taap.2014.10.013. [DOI] [PubMed] [Google Scholar]
- 48.Wang T, Li X, Yang D, Zhang H, Zhao P, Fu J, et al. ER stress and ER stress-mediated apoptosis are involved in manganese-induced neurotoxicity in the rat striatum in vivo. Neurotoxicology. 2015;48:109–119. doi: 10.1016/j.neuro.2015.02.007. [DOI] [PubMed] [Google Scholar]
- 49.Tuschl K, Mills PB, Clayton PT. Manganese and the brain. Int Rev Neurobiol. 2013;110:277–312. doi: 10.1016/B978-0-12-410502-7.00013-2. [DOI] [PubMed] [Google Scholar]
- 50.Chen P, Chakraborty S, Mukhopadhyay S, Lee E, Paoliello MMB, Bowman AB, et al. Manganese homeostasis in the nervous system. J Neurochem. 2015;134:601–610. doi: 10.1111/jnc.13170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Streifel KM, Miller J, Mouneimne R, Tjalkens RB. Manganese inhibits ATP-induced calcium entry through the transient receptor potential channel TRPC3 in astrocytes. Neuro Toxicol. 2013;34:160–166. doi: 10.1016/j.neuro.2012.10.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Gunshin H, Mackenzie B, Berger UV, Gunshin Y, Romero MF, Boron WF, et al. Cloning and characterization of a mammalian proton-coupled metal-ion transporter. Nature. 1997;388:482–488. doi: 10.1038/41343. [DOI] [PubMed] [Google Scholar]
- 53.Garrick MD, Singelton ST, Vargas F, Kuo H-C, Zhao L, Knöpfel M, et al. DMT1: Which metals does it transport? Biol Res. 2006;39:79–85. doi: 10.4067/S0716-97602006000100009. [DOI] [PubMed] [Google Scholar]
- 54.Huang E, Ong WY, Connor JR. Distribution of divalent metal transporter-1 in the monkey basal ganglia. Neuroscience. 2004;128:487–496. doi: 10.1016/j.neuroscience.2004.06.055. [DOI] [PubMed] [Google Scholar]
- 55.Williams K, Wilson MA, Bressler J. Regulation and developmental expression of the divalent metal-ion transporter in the rat brain. Cell mol biol (Noisy-le-Grand, France. 2000;46:563–571. [PubMed] [Google Scholar]
- 56.Burdo JR, Menzies SL, Simpson IA, Garrick LM, Garrick MD, Dolan KG, et al. Distribution of divalent metal transporter 1 and metal transport protein 1 in the normal and Belgrade rat. J Neurosci Res. 2001;66:1198–1207. doi: 10.1002/jnr.1256. [DOI] [PubMed] [Google Scholar]
- 57.Gunter TE, Gerstner B, Gunter KK, Malecki J, Gelein R, Valentine WM, et al. Manganese transport via the transferrin mechanism. Neuro Toxicol. 2013;34:118–127. doi: 10.1016/j.neuro.2012.10.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Aisen P, Leibman A, Zweier J. Stoichiometric and site characteristics of the binding of iron to human transferrin. J Biol Chem. 1978;253:1930–1937. [PubMed] [Google Scholar]
- 59.Li GJ, Zhao Q, Zheng W. Alteration at translational but not transcriptional level of transferrin receptor expression following manganese exposure at the blood–CSF barrier in vitro. Toxicol Appl Pharmacol. 2005;205:188–200. doi: 10.1016/j.taap.2004.10.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Moos T, Morgan E. Transferrin and Transferrin Receptor Function in Brain Barrier Systems. Cell Mol Neurobiol. 2000;20:77–95. doi: 10.1023/A:1006948027674. [DOI] [PubMed] [Google Scholar]
- 61.Wu LJ-c, Leenders AGM, Cooperman S, Meyron-Holtz E, Smith S, Land W, Tsai RYL, Berger UV, Sheng Z-H, Rouault TA: Expression of the iron transporter ferroportin in synaptic vesicles and the blood–brain barrier. Brain Res 2004, 1001:108–117. [DOI] [PubMed]
- 62.Yin Z, Jiang H, Lee E-SY, Ni M, Erikson KM, Milatovic D, et al. Ferroportin is a manganese-responsive protein that decreases manganese cytotoxicity and accumulation. J Neurochem. 2010;112:1190–1198. doi: 10.1111/j.1471-4159.2009.06534.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Madejczyk MS, Ballatori N: The iron transporter ferroportin can also function as a manganese exporter. Biochimica et Biophysica Acta (BBA) - Biomembranes 2012, 1818:651–657. [DOI] [PMC free article] [PubMed]
- 64.Tuschl K, Clayton PT, Gospe SM, Jr, Gulab S, Ibrahim S, Singhi P, et al. Syndrome of hepatic cirrhosis, dystonia, polycythemia, and hypermanganesemia caused by mutations in SLC30A10, a manganese transporter in man. Am J Hum Genet. 2012;90:457–466. doi: 10.1016/j.ajhg.2012.01.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Stamelou M, Tuschl K, Chong WK, Burroughs AK, Mills PB, Bhatia KP, et al. Dystonia with brain manganese accumulation resulting from SLC30A10 mutations: A new treatable disorder. Mov Disord. 2012;27:1317–1322. doi: 10.1002/mds.25138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Tuschl K, Mills PB, Parsons H, Malone M, Fowler D, Bitner-Glindzicz M, et al. Hepatic cirrhosis, dystonia, polycythaemia and hypermanganesaemia--a new metabolic disorder. J Inherit Metab Dis. 2008;31:151–163. doi: 10.1007/s10545-008-0813-1. [DOI] [PubMed] [Google Scholar]
- 67.Leyva-Illades D, Chen P, Zogzas CE, Hutchens S, Mercado JM, Swaim CD, et al. SLC30A10 Is a Cell Surface-Localized Manganese Efflux Transporter, and Parkinsonism-Causing Mutations Block Its Intracellular Trafficking and Efflux Activity. J Neurosci. 2014;34:14079–14095. doi: 10.1523/JNEUROSCI.2329-14.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Hu Z, Bonifas JM, Beech J, Bench G, Shigihara T, Ogawa H, et al. Mutations in ATP2C1, encoding a calcium pump, cause Hailey-Hailey disease. Nat Genet. 2000;24:61–65. doi: 10.1038/71701. [DOI] [PubMed] [Google Scholar]
- 69.Mukhopadhyay S, Linstedt AD. Identification of a gain-of-function mutation in a Golgi P-type ATPase that enhances Mn2+ efflux and protects against toxicity. Proc Natl Acad Sci U S A. 2011;108:858–863. doi: 10.1073/pnas.1013642108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Gitler AD, Chesi A, Geddie ML, Strathearn KE, Hamamichi S, Hill KJ, et al. Alpha-synuclein is part of a diverse and highly conserved interaction network that includes PARK9 and manganese toxicity. Nat Genet. 2009;41:308–315. doi: 10.1038/ng.300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Park JS, Mehta P, Cooper AA, Veivers D, Heimbach A, Stiller B, Kubisch C, Fung VS, Krainc D, Mackay-Sim A. and Sue CM. (2011), Pathogenic effects of novel mutations in the P-type ATPase ATP13A2 (PARK9) causing Kufor-Rakeb syndrome, a form of early-onset parkinsonism. Hum. Mutat., 32: 956– 964. [DOI] [PubMed]
- 72.Di Fonzo A, Chien HF, Socal M, Giraudo S, Tassorelli C, Iliceto G, et al. ATP13A2 missense mutations in juvenile parkinsonism and young onset Parkinson disease. Neurology. 2007;68:1557–62. [DOI] [PubMed]
- 73.Tan J, Zhang T, Jiang L, Chi J, Hu D, Pan Q, et al. Regulation of intracellular manganese homeostasis by Kufor-Rakeb syndrome-associated ATP13A2 protein. J Biol Chem. 2011;286:29654–29662. doi: 10.1074/jbc.M111.233874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Kumar KK, Lowe JEW, Aboud AA, Neely MD, Redha R, Bauer JA, et al. Cellular manganese content is developmentally regulated in human dopaminergic neurons. Sci Rep. 2014;4:6801. doi: 10.1038/srep06801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Guilarte TR, Burton NC, Mcglothan JL, Verina T, Zhou Y, Alexander M, et al. Impairment of nigrostriatal dopamine neurotransmission by manganese is mediated by pre-synaptic mechanism (s): implications to manganese-induced parkinsonism. J Neurochem. 2008;107:1236–1247. doi: 10.1111/j.1471-4159.2008.05695.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Fordahl SC, Erikson KM. Manganese accumulation in membrane fractions of primary astrocytes is associated with decreased γ-aminobutyric acid (GABA) uptake, and is exacerbated by oleic acid and palmitate. Environ Toxicol Pharmacol. 2014;37:1148–1156. doi: 10.1016/j.etap.2014.03.016. [DOI] [PubMed] [Google Scholar]
- 77.Lee E-SY, Sidoryk M, Jiang H, Yin Z, Aschner M. Estrogen and tamoxifen reverse manganese-induced glutamate transporter impairment in astrocytes. J Neurochem. 2009;110:530–544. doi: 10.1111/j.1471-4159.2009.06105.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Erikson KM, Suber RL, Aschner M. Glutamate/Aspartate Transporter (GLAST), Taurine Transporter and Metallothionein mRNA Levels are Differentially Altered in Astrocytes Exposed to Manganese Chloride, Manganese Phosphate or Manganese Sulfate. Neuro Toxicol. 2002;23:281–288. doi: 10.1016/s0161-813x(02)00041-4. [DOI] [PubMed] [Google Scholar]
- 79.Sidoryk-Wegrzynowicz M, Aschner M. Manganese toxicity in the central nervous system: the glutamine/glutamate-γ-aminobutyric acid cycle. J Intern Med. 2013;273:466–477. doi: 10.1111/joim.12040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Sidoryk-Węgrzynowicz M, Lee E, Albrecht J, Aschner M. Manganese disrupts astrocyte glutamine transporter expression and function. J Neurochem. 2009;110:822–830. doi: 10.1111/j.1471-4159.2009.06172.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.da Silva Santos V, Bisen-Hersh E, Yu Y, Cabral IS, Nardini V, Culbreth M, et al. Anthocyanin-rich acai (Euterpe oleracea Mart.) extract attenuates manganese-induced oxidative stress in rat primary astrocyte cultures. J Toxicol Environ Health A. 2014;77:390–404. doi: 10.1080/15287394.2014.880392. [DOI] [PubMed] [Google Scholar]
- 82.Karki P, Smith K, Johnson J, Jr, Aschner M, Lee E. Role of transcription factor yin yang 1 in manganese-induced reduction of astrocytic glutamate transporters: Putative mechanism for manganese-induced neurotoxicity. Neurochem Int. 2015;88:53–59. doi: 10.1016/j.neuint.2014.08.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Finkelstein Y, Milatovic D, Aschner M. Modulation of cholinergic systems by manganese. Neuro Toxicol. 2007;28:1003–1014. doi: 10.1016/j.neuro.2007.08.006. [DOI] [PubMed] [Google Scholar]
- 84.Santos D, Milatovic D, Andrade V, Batoreu MC, Aschner M, Marreilha Dos Santos AP. The inhibitory effect of manganese on acetylcholinesterase activity enhances oxidative stress and neuroinflammation in the rat brain. Toxicology. 2012;292:90–98. doi: 10.1016/j.tox.2011.11.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Ye Q, Kim J. Loss of hfe function reverses impaired recognition memory caused by olfactory manganese exposure in mice. Toxicol res. 2015;31:17–23. doi: 10.5487/TR.2015.31.1.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Sarter M, Bruno JP. Cognitive functions of cortical acetylcholine: toward a unifying hypothesis. Brain Res Rev. 1997;23:28–46. doi: 10.1016/S0165-0173(96)00009-4. [DOI] [PubMed] [Google Scholar]
- 87.Calabresi P, Ammassari-Teule M, Gubellini P, Sancesario G, Morello M, Centonze D, et al. A Synaptic Mechanism Underlying the Behavioral Abnormalities Induced by Manganese Intoxication. Neurobiol Dis. 2001;8:419–432. doi: 10.1006/nbdi.2000.0379. [DOI] [PubMed] [Google Scholar]
- 88.Silman I, Sussman JL. Acetylcholinesterase: ‘classical’ and ‘non-classical’ functions and pharmacology. Curr Opin Pharmacol. 2005;5:293–302. doi: 10.1016/j.coph.2005.01.014. [DOI] [PubMed] [Google Scholar]
- 89.Massoulié J, Pezzementi L, Bon S, Krejci E, Vallette F-M. Molecular and cellular biology of cholinesterases. Prog Neurobiol. 1993;41:31–91. doi: 10.1016/0301-0082(93)90040-Y. [DOI] [PubMed] [Google Scholar]
- 90.Tõugu V, Kesvatera T: Role of ionic interactions in cholinesterase catalysis. Biochimica et Biophysica Acta (BBA) - Protein Structure and Molecular Enzymology 1996, 1298:12–30. [DOI] [PubMed]
- 91.Johnson G, Moore SW. The Adhesion Function on Acetylcholinesterase Is Located at the Peripheral Anionic Site. Biochem Biophys Res Commun. 1999;258:758–762. doi: 10.1006/bbrc.1999.0705. [DOI] [PubMed] [Google Scholar]
- 92.Talesa VN. Acetylcholinesterase in Alzheimer’s disease. Mech Ageing Dev. 2001;122:1961–1969. doi: 10.1016/S0047-6374(01)00309-8. [DOI] [PubMed] [Google Scholar]
- 93.Mesulam MM, Guillozet A, Shaw P, Levey A, Duysen EG, Lockridge O. Acetylcholinesterase knockouts establish central cholinergic pathways and can use butyrylcholinesterase to hydrolyze acetylcholine. Neuroscience. 2002;110:627–639. doi: 10.1016/S0306-4522(01)00613-3. [DOI] [PubMed] [Google Scholar]
- 94.Fernsebner K, Zorn J, Kanawati B, Walker A, Michalke B. Manganese leads to an increase in markers of oxidative stress as well as to a shift in the ratio of Fe (ii)/(iii) in rat brain tissue. Metallomics. 2014;6:921–931. doi: 10.1039/c4mt00022f. [DOI] [PubMed] [Google Scholar]
- 95.Kaizer RR, Corrêa MC, Spanevello RM, Morsch VM, Mazzanti CM, Gonçalves JF, et al. Acetylcholinesterase activation and enhanced lipid peroxidation after long-term exposure to low levels of aluminum on different mouse brain regions. J Inorg Biochem. 2005;99:1865–1870. doi: 10.1016/j.jinorgbio.2005.06.015. [DOI] [PubMed] [Google Scholar]
- 96.Thome GR, Spanevello RM, Mazzanti A, Fiorenza AM, Duarte MM, da Luz SC, et al. Vitamin E decreased the activity of acetylcholinesterase and level of lipid peroxidation in brain of rats exposed to aged and diluted sidestream smoke. Nicotine tob res : off j Soc Res Nicotine Tob. 2011;13:1210–1219. doi: 10.1093/ntr/ntr154. [DOI] [PubMed] [Google Scholar]
- 97.Lai JC, Chan AW, Leung TK, Minski MJ, Lim L. Neurochemical changes in rats chronically treated with a high concentration of manganese chloride. Neurochem Res. 1992;17:841–847. doi: 10.1007/BF00993259. [DOI] [PubMed] [Google Scholar]
- 98.Lai JCK, Leung TKC, Lim L. The Ontogeny of Acetylcholinesterase Activities in Rat Brain Regions and the Effect of Chronic Treatment with Manganese Chloride. J Neurochem. 1982;39:1767–1769. doi: 10.1111/j.1471-4159.1982.tb08019.x. [DOI] [PubMed] [Google Scholar]
- 99.Liapi C, Zarros A, Galanopoulou P, Theocharis S, Skandali N, Al-Humadi H, et al. Effects of short-term exposure to manganese on the adult rat brain antioxidant status and the activities of acetylcholinesterase, (Na, K)-ATPase and Mg-ATPase: modulation by L-cysteine. Basic Clin Pharmacol Toxicol. 2008;103:171–175. doi: 10.1111/j.1742-7843.2008.00281.x. [DOI] [PubMed] [Google Scholar]
- 100.Yousefi Babadi V, Sadeghi L, Shirani K, Malekirad AA, Rezaei M: The toxic effect of manganese on the acetylcholinesterase activity in rat brains. Journal of toxicology 2014, 2014:946372. [DOI] [PMC free article] [PubMed]
- 101.Felder CC, Bymaster FP, Ward J, Delapp N. Therapeutic opportunities for muscarinic receptors in the central nervous system. J Med Chem. 2000;43:4333–4353. doi: 10.1021/jm990607u. [DOI] [PubMed] [Google Scholar]
- 102.Unwin N. Structure and action of the nicotinic acetylcholine receptor explored by electron microscopy. FEBS Lett. 2003;555:91–95. doi: 10.1016/S0014-5793(03)01084-6. [DOI] [PubMed] [Google Scholar]
- 103.Baldwin JM. Structure and function of receptors coupled to G proteins. Curr Opin Cell Biol. 1994;6:180–190. doi: 10.1016/0955-0674(94)90134-1. [DOI] [PubMed] [Google Scholar]
- 104.Cano G, Bonilla E, Castro F, Alburges M, Wamsley JK. [Chronic manganese poisoning: autoradiographic quantification of cholinergic muscarinic receptors in mouse brain] Invest Clin. 1992;33:69–79. [PubMed] [Google Scholar]
- 105.Villalobos V, Castro F, Bonilla E, Estevez J, Davila JO. Manganese toxicity: muscarinic receptor binding in the mouse brain. J Toxicol Environ Health. 1994;42:185–191. doi: 10.1080/15287399409531872. [DOI] [PubMed] [Google Scholar]
- 106.Eriksson H, Gillberg PG, Aquilonius SM, Hedstrom KG, Heilbronn E. Receptor alterations in manganese intoxicated monkeys. Arch Toxicol. 1992;66:359–364. doi: 10.1007/BF01973632. [DOI] [PubMed] [Google Scholar]
- 107.Wang H, Liao H, Ochani M, Justiniani M, Lin X, Yang L, et al. Cholinergic agonists inhibit HMGB1 release and improve survival in experimental sepsis. Nat Med. 2004;10:1216–1221. doi: 10.1038/nm1124. [DOI] [PubMed] [Google Scholar]
- 108.Wang H, Yu M, Ochani M, Amella CA, Tanovic M, Susarla S, et al. Nicotinic acetylcholine receptor [alpha] 7 subunit is an essential regulator of inflammation. Nature. 2003;421:384–388. doi: 10.1038/nature01339. [DOI] [PubMed] [Google Scholar]
- 109.Borovikova LV, Ivanova S, Zhang M, Yang H, Botchkina GI, Watkins LR, et al. Vagus nerve stimulation attenuates the systemic inflammatory response to endotoxin. Nature. 2000;405:458–462. doi: 10.1038/35013070. [DOI] [PubMed] [Google Scholar]
- 110.Tracey KJ. The inflammatory reflex. Nature. 2002;420:853–859. doi: 10.1038/nature01321. [DOI] [PubMed] [Google Scholar]
- 111.Dinarello CA. Therapeutic strategies to reduce IL-1 activity in treating local and systemic inflammation. Curr Opin Pharmacol. 2004;4:378–385. doi: 10.1016/j.coph.2004.03.010. [DOI] [PubMed] [Google Scholar]
- 112.Pavlov VA, Wang H, Czura CJ, Friedman SG, Tracey KJ. The cholinergic anti-inflammatory pathway: a missing link in neuroimmunomodulation. Mol Med. 2003;9:125–134. [PMC free article] [PubMed] [Google Scholar]
- 113.Das UN: Acetylcholinesterase and butyrylcholinesterase as possible markers of low-grade systemic inflammation. Med Sci Monitor 2007, 13:Ra214-Ra221. [PubMed]
- 114.Kimura R, Ushiyama N, Fujii T, Kawashima K. Nicotine-induced Ca2+ signaling and down-regulation of nicotinic acetylcholine receptor subunit expression in the CEM human leukemic T-cell line. Life Sci. 2003;72:2155–2158. doi: 10.1016/S0024-3205(03)00077-8. [DOI] [PubMed] [Google Scholar]
- 115.Li B, Stribley JA, Ticu A, Xie W, Schopfer LM, Hammond P, et al. Abundant tissue butyrylcholinesterase and its possible function in the acetylcholinesterase knockout mouse. J Neurochem. 2000;75:1320–1331. doi: 10.1046/j.1471-4159.2000.751320.x. [DOI] [PubMed] [Google Scholar]
- 116.Meeker JD, Susi P, Flynn MR. Manganese and welding fume exposure and control in construction. J Occup Environ Hyg. 2007;4:943–951. doi: 10.1080/15459620701718867. [DOI] [PubMed] [Google Scholar]
- 117.Gianutsos G, Morrow GR, Morris JB. Accumulation of manganese in rat brain following intranasal administration. Fundam Appl Toxicol. 1997;37:102–105. doi: 10.1006/faat.1997.2306. [DOI] [PubMed] [Google Scholar]
- 118.Henriksson J, Tallkvist J, Tjälve H. Transport of Manganese via the Olfactory Pathway in Rats: Dosage Dependency of the Uptake and Subcellular Distribution of the Metal in the Olfactory Epithelium and the Brain. Toxicol Appl Pharmacol. 1999;156:119–128. doi: 10.1006/taap.1999.8639. [DOI] [PubMed] [Google Scholar]
- 119.Brenneman KA, Wong BA, Buccellato MA, Costa ER, Gross EA, Dorman DC. Direct Olfactory Transport of Inhaled Manganese (54MnCl2) to the Rat Brain: Toxicokinetic Investigations in a Unilateral Nasal Occlusion Model. Toxicol Appl Pharmacol. 2000;169:238–248. doi: 10.1006/taap.2000.9073. [DOI] [PubMed] [Google Scholar]
- 120.Kim J, Buckett PD, Wessling-Resnick M: Absorption of Manganese and Iron in a Mouse Model of Hemochromatosis. PLoS One 2013, 8. [DOI] [PMC free article] [PubMed]
- 121.Thompson K, Molina RM, Donaghey T, Schwob JE, Brain JD, Wessling-Resnick M. Olfactory uptake of manganese requires DMT1 and is enhanced by anemia. FASEB J. 2007;21:223–230. doi: 10.1096/fj.06-6710com. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Heilig E, Molina R, Donaghey T, Brain JD, Wessling-Resnick M. Pharmacokinetics of pulmonary manganese absorption: evidence for increased susceptibility to manganese loading in iron-deficient rats. Am J Physiol Lung C. 2005;288:L887–L893. doi: 10.1152/ajplung.00382.2004. [DOI] [PubMed] [Google Scholar]
- 123.Gorell JM, Johnson CC, Rybicki BA, Peterson EL, Kortsha GX, Brown GG, et al. Occupational exposure to manganese, copper, lead, iron, mercury and zinc and the risk of Parkinson’s disease. Neurotoxicology. 1999;20:239–247. [PubMed] [Google Scholar]
- 124.Racette BA, Mcgee-Minnich L, Moerlein SM, Mink JW, Videen TO, Perlmutter JS. Welding-related parkinsonism: clinical features, treatment, and pathophysiology. Neurology. 2001;56:8–13. doi: 10.1212/WNL.56.1.8. [DOI] [PubMed] [Google Scholar]
- 125.Harischandra DS, Jin H, Anantharam V, Kanthasamy A, Kanthasamy AG. α-Synuclein Protects Against Manganese Neurotoxic Insult During the Early Stages of Exposure in a Dopaminergic Cell Model of Parkinson’s Disease. Toxicol Sci. 2015;143:454–468. doi: 10.1093/toxsci/kfu247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Bornhorst J, Chakraborty S, Meyer S, Lohren H, Gro, Knight AL, Caldwell KA, Caldwell GA, Karst U, Schwerdtle T, et al.: The effects of pdr1, djr1.1 and pink1 loss in manganese-induced toxicity and the role of alpha-synuclein in C. elegans. Metallomics 2014, 6:476–490. [DOI] [PMC free article] [PubMed]
- 127.Dučić T, Carboni E, Lai B, Chen S, Michalke B, Lázaro DF, et al. Alpha-Synuclein Regulates Neuronal Levels of Manganese and Calcium. ACS Chem Nerosci. 2015;6:1769–1779. doi: 10.1021/acschemneuro.5b00093. [DOI] [PubMed] [Google Scholar]
- 128.Mendonca-Dias MH, Gaggelli E, Lauterbur PC. Paramagnetic contrast agents in nuclear magnetic resonance medical imaging. Semin Nucl Med. 1983;13:364–376. doi: 10.1016/S0001-2998(83)80048-8. [DOI] [PubMed] [Google Scholar]
- 129.Criswell SR, Perlmutter JS, Huang JL, Golchin N, Flores HP, Hobson A, et al. Basal ganglia intensity indices and diffusion weighted imaging in manganese-exposed welders. Occup Environ Med. 2012;69:437–443. doi: 10.1136/oemed-2011-100119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Dietz MC, Ihrig A, Wrazidlo W, Bader M, Jansen O, Triebig G. Results of magnetic resonance imaging in long-term manganese dioxide-exposed workers. Environ Res. 2001;85:37–40. doi: 10.1006/enrs.2000.4068. [DOI] [PubMed] [Google Scholar]
- 131.Baker MG, Criswell SR, Racette BA, Simpson CD, Sheppard L, Checkoway H, et al. Neurological outcomes associated with low-level manganese exposure in an inception cohort of asymptomatic welding trainees. Scand J Work Environ Health. 2015;41:94–101. doi: 10.5271/sjweh.3466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Criswell SR, Nelson G, Gonzalez-Cuyar LF, Huang J, Shimony JS, Checkoway H, et al. Ex vivo magnetic resonance imaging in South African manganese mine workers. Neurotoxicology. 2015;49:8–14. doi: 10.1016/j.neuro.2015.04.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Gonzalez-Cuyar LF, Nelson G, Criswell SR, Ho P, Lonzanida JA, Checkoway H, et al. Quantitative neuropathology associated with chronic manganese exposure in South African mine workers. Neuro Toxicol. 2014;45:260–266. doi: 10.1016/j.neuro.2013.12.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Filipov NM, Dodd CA. Role of glial cells in manganese neurotoxicity. J Appl Toxicol. 2012;32:310–317. doi: 10.1002/jat.1762. [DOI] [PubMed] [Google Scholar]
- 135.Aschner M, Gannon M, Kimelberg HK. Manganese uptake and efflux in cultured rat astrocytes. J Neurochem. 1992;58:730–735. doi: 10.1111/j.1471-4159.1992.tb09778.x. [DOI] [PubMed] [Google Scholar]
- 136.Wedler FC, Ley BW, Grippo AA. Manganese (II) dynamics and distribution in glial cells cultured from chick cerebral cortex. Neurochem Res. 1989;14:1129–1135. doi: 10.1007/BF00965619. [DOI] [PubMed] [Google Scholar]
- 137.Gonzalez LE, Juknat AA, Venosa AJ, Verrengia N, Kotler ML. Manganese activates the mitochondrial apoptotic pathway in rat astrocytes by modulating the expression of proteins of the Bcl-2 family. Neurochem Int. 2008;53:408–415. doi: 10.1016/j.neuint.2008.09.008. [DOI] [PubMed] [Google Scholar]
- 138.Henriksson J, Tjalve H. Manganese taken up into the CNS via the olfactory pathway in rats affects astrocytes. Toxicol Sci. 2000;55:392–398. doi: 10.1093/toxsci/55.2.392. [DOI] [PubMed] [Google Scholar]
- 139.Peres TV, Eyng H, Lopes SC, Colle D, Goncalves FM, Venske DK, et al. Developmental exposure to manganese induces lasting motor and cognitive impairment in rats. Neurotoxicology. 2015;50:28–37. doi: 10.1016/j.neuro.2015.07.005. [DOI] [PubMed] [Google Scholar]
- 140.Kern CH, Smith DR. Preweaning Mn exposure leads to prolonged astrocyte activation and lasting effects on the dopaminergic system in adult male rats. Synapse. 2011;65:532–544. doi: 10.1002/syn.20873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Chen CJ, Ou YC, Lin SY, Liao SL, Chen SY, Chen JH. Manganese modulates pro-inflammatory gene expression in activated glia. Neurochem Int. 2006;49:62–71. doi: 10.1016/j.neuint.2005.12.020. [DOI] [PubMed] [Google Scholar]
- 142.Lucchini RG, Albini E, Benedetti L, Borghesi S, Coccaglio R, Malara EC, et al. High prevalence of parkinsonian disorders associated to manganese exposure in the vicinities of ferroalloy industries. Am J Ind Med. 2007;50:788–800. doi: 10.1002/ajim.20494. [DOI] [PubMed] [Google Scholar]
- 143.Bowler RM, Kornblith ES, Gocheva VV, Colledge MA, Bollweg G, Kim Y, et al. Environmental exposure to manganese in air: Associations with cognitive functions. Neurotoxicology. 2015;49:139–148. doi: 10.1016/j.neuro.2015.06.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Haynes EN, Sucharew H, Kuhnell P, Alden J, Barnas M, Wright RO, Parsons PJ, Aldous KM, Praamsma ML, Beidler C, Dietrich KN: Manganese Exposure and Neurocognitive Outcomes in Rural School-Age Children: The Communities Actively Researching Exposure Study (Ohio, USA). Environ Health Perspect 2015. [DOI] [PMC free article] [PubMed]
- 145.Rugless F, Bhattacharya A, Succop P, Dietrich KN, Cox C, Alden J, et al. Childhood exposure to manganese and postural instability in children living near a ferromanganese refinery in Southeastern Ohio. Neurotoxicol Teratol. 2014;41:71–79. doi: 10.1016/j.ntt.2013.12.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Grandjean P, Landrigan PJ. Neurobehavioural effects of developmental toxicity. Lancet Neurol. 2014;13:330–338. doi: 10.1016/S1474-4422(13)70278-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Zoni S, Lucchini RG. Manganese exposure: cognitive, motor and behavioral effects on children: a review of recent findings. Curr Opin Pediatr. 2013;25:255–260. doi: 10.1097/MOP.0b013e32835e906b. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Moreno JA, Yeomans EC, Streifel KM, Brattin BL, Taylor RJ, Tjalkens RB. Age-Dependent Susceptibility to Manganese-Induced Neurological Dysfunction. Toxicol Sci. 2009;112:394–404. doi: 10.1093/toxsci/kfp220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Kern CH, Stanwood GD, Smith DR. Preweaning manganese exposure causes hyperactivity, disinhibition, and spatial learning and memory deficits associated with altered dopamine receptor and transporter levels. Synapse. 2010;64:363–378. doi: 10.1002/syn.20736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Beaudin SA, Nisam S, Smith DR. Early life versus lifelong oral manganese exposure differently impairs skilled forelimb performance in adult rats. Neurotoxicol Teratol. 2013;38:36–45. doi: 10.1016/j.ntt.2013.04.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Bouchard M. Hair manganese and hyperactive behaviors: Pilot study of school-age children exposed through tap water. Epidemiology. 2007;18:S164–S165. doi: 10.1097/01.ede.0000276837.62405.bb. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Wasserman GA, Liu X, Parvez F, Ahsan H, Levy D, Factor-Litvak P, et al. Water manganese exposure and children’s intellectual function in Araihazar, Bangladesh. Environ Health Perspect. 2006;114:124–129. doi: 10.1289/ehp.8030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Menezes-Filho JA, Novaes CO, Moreira JC, Sarcinelli PN, Mergler D. Elevated manganese and cognitive performance in school-aged children and their mothers. Environ Res. 2011;111:156–163. doi: 10.1016/j.envres.2010.09.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Carvalho CF, Menezes-Filho JA, Matos VP, Bessa JR, Coelho-Santos J, Viana GF, Argollo N, Abreu N: Elevated airborne manganese and low executive function in school-aged children in Brazil. Neurotoxicology 2013. [DOI] [PubMed]
- 155.Alves G, Thiebot J, Tracqui A, Delangre T, Guedon C, Lerebours E. Neurologic disorders due to brain manganese deposition in a jaundiced patient receiving long-term parenteral nutrition. J Parenter Enteral Nutr. 1997;21:41–45. doi: 10.1177/014860719702100141. [DOI] [PubMed] [Google Scholar]
- 156.Siepler JK, Nishikawa RA, Diamantidis T, Okamoto R. Asymptomatic hypermanganesemia in long-term home parenteral nutrition patients. Nutr clin pract off pub Am Soc Parenter Enter Nutr. 2003;18:370–373. doi: 10.1177/0115426503018005370. [DOI] [PubMed] [Google Scholar]
- 157.Iinuma Y, Kubota M, Uchiyama M, Yagi M, Kanada S, Yamazaki S, et al. Whole-blood manganese levels and brain manganese accumulation in children receiving long-term home parenteral nutrition. Pediatr Surg Int. 2003;19:268–272. doi: 10.1007/s00383-002-0929-6. [DOI] [PubMed] [Google Scholar]
- 158.Horning KJ, Caito SW, Tipps KG, Bowman AB, Aschner M. Manganese Is Essential for Neuronal Health. Annu Rev Nutr. 2015;35:71–108. doi: 10.1146/annurev-nutr-071714-034419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Betharia S, Maher TJ. Neurobehavioral effects of lead and manganese individually and in combination in developmentally exposed rats. Neurotoxicology. 2012;33:1117–1127. doi: 10.1016/j.neuro.2012.06.002. [DOI] [PubMed] [Google Scholar]
- 160.Blecharz-Klin K, Piechal A, Joniec-Maciejak I, Pyrzanowska J, Widy-Tyszkiewicz E. Effect of intranasal manganese administration on neurotransmission and spatial learning in rats. Toxicol Appl Pharmacol. 2012;265:1–9. doi: 10.1016/j.taap.2012.09.015. [DOI] [PubMed] [Google Scholar]
- 161.Chen P, Dewitt MR, Bornhorst J, Soares FA, Mukhopadhyay S, Bowman AB, et al. Age- and manganese-dependent modulation of dopaminergic phenotypes in a C. elegans DJ-1 genetic model of Parkinson’s disease. Metallomics. 2015;7:289–298. doi: 10.1039/C4MT00292J. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Li SJ, Meng HY, Deng XF, Fu X, Chen JW, Huang S, et al. Protective effects of sodium p-aminosalicylic acid on learning and memory via increasing the number of basal forebrain choline acetyltransferase neurons in manganese-exposed rats. Hum Exp Toxicol. 2015;34:240–248. doi: 10.1177/0960327114529454. [DOI] [PubMed] [Google Scholar]
- 163.Liang G, Qin H, Zhang L, Ma S, Huang X, Lv Y, et al. Effects of chronic manganese exposure on the learning and memory of rats by observing the changes in the hippocampal cAMP signaling pathway. Food Chem Toxicol. 2015;83:261–267. doi: 10.1016/j.fct.2015.07.005. [DOI] [PubMed] [Google Scholar]
- 164.Lu CL, Tang S, Meng ZJ, He YY, Song LY, Liu YP, et al. Taurine improves the spatial learning and memory ability impaired by sub-chronic manganese exposure. J Biomed Sci. 2014;21:51. doi: 10.1186/1423-0127-21-51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Schneider JS, Decamp E, Clark K, Bouquio C, Syversen T, Guilarte TR: Effects of chronic manganese exposure on working memory in non-human primates. Brain Res 2009, 1258:86–95. [DOI] [PMC free article] [PubMed]
- 166.Schneider JS, Decamp E, Koser AJ, Fritz S, Gonczi H, Syversen T, Guilarte TR: Effects of chronic manganese exposure on cognitive and motor functioning in non-human primates. Brain Res 2006, 1118:222–231. [DOI] [PMC free article] [PubMed]
- 167.Schneider JS, Williams C, Ault M, Guilarte TR. Effects of chronic manganese exposure on attention and working memory in non-human primates. Neurotoxicology. 2015;48:217–222. doi: 10.1016/j.neuro.2015.04.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Shukakidze A, Lazriev I, Mitagvariya N. Behavioral impairments in acute and chronic manganese poisoning in white rats. Neurosci Behav Physiol. 2003;33:263–267. doi: 10.1023/A:1022103415424. [DOI] [PubMed] [Google Scholar]
- 169.Su C, Chen K, Zou Y, Shen Y, Xia B, Liang G, Lv Y, Wang F, Huang D, Yang X: Chronic exposure to manganese sulfate leads to adverse dose-dependent effects on the neurobehavioral ability of rats. Environ Toxicol 2015. [DOI] [PubMed]
- 170.Vezer T, Papp A, Hoyk Z, Varga C, Naray M, Nagymajtenyi L. Behavioral and neurotoxicological effects of subchronic manganese exposure in rats. Environ Toxicol Pharmacol. 2005;19:797–810. doi: 10.1016/j.etap.2004.12.046. [DOI] [PubMed] [Google Scholar]
- 171.Bromley-Brits K, Deng Y, Song W: Morris water maze test for learning and memory deficits in Alzheimer’s disease model mice. Journal of visualized experiments : JoVE 2011. [DOI] [PMC free article] [PubMed]
- 172.Penley SC, Gaudet CM, Threlkeld SW: Use of an Eight-arm Radial Water Maze to Assess Working and Reference Memory Following Neonatal Brain Injury. 2013:e50940. [DOI] [PMC free article] [PubMed]
- 173.Zovkic IB, Guzman-Karlsson MC, Sweatt JD. Epigenetic regulation of memory formation and maintenance. Learn Mem. 2013;20:61–74. doi: 10.1101/lm.026575.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Benedetto A, Au C, Avila DS, Milatovic D, Aschner M. Extracellular Dopamine Potentiates Mn-Induced Oxidative Stress, Lifespan Reduction, and Dopaminergic Neurodegeneration in a BLI-3–Dependent Manner in Caenorhabditis elegans PLoS Genet. 2010;6:e1001084. [DOI] [PMC free article] [PubMed]
- 175.Chen P, Martinez-Finley EJ, Bornhorst J, Chakraborty S, Aschner M: Metal-Induced Neurodegeneration in C. elegans. Frontiers in aging neuroscience 2013, 5. [DOI] [PMC free article] [PubMed]
- 176.Chen P, DeWitt MR, Bornhorst J, Soares FA, Mukhopadhyay S, Bowman AB, Aschner M: Age- and manganese-dependent modulation of dopaminergic phenotypes in a C. elegans DJ-1 genetic model of Parkinson’s disease. Metallomics 2015. [DOI] [PMC free article] [PubMed]
- 177.Sawin ER, Ranganathan R, Horvitz HR. C. elegans Locomotory Rate Is Modulated by the Environment through a Dopaminergic Pathway and by Experience through a Serotonergic Pathway. Neuron. 2000;26:619–631. doi: 10.1016/S0896-6273(00)81199-X. [DOI] [PubMed] [Google Scholar]
- 178.Chakraborty S, Chen P, Bornhorst J, Schwerdtle T, Schumacher F, Kleuser B, et al. Loss of pdr-1/parkin influences Mn homeostasis through altered ferroportin expression in C. elegans. Metallomics. 2015;7:847–856. doi: 10.1039/C5MT00052A. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Aschner M, Guilarte TR, Schneider JS, Zheng W. Manganese: Recent advances in understanding its transport and neurotoxicity. Toxicol Appl Pharmacol. 2007;221:131–147. doi: 10.1016/j.taap.2007.03.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Santos D, Batoreu CM, Tavares De Almeida I, Davis Randall L, Mateus LM, Andrade V, et al. Evaluation of neurobehavioral and neuroinflammatory end-points in the post-exposure period in rats sub-acutely exposed to manganese. Toxicology. 2013;314:95–99. doi: 10.1016/j.tox.2013.09.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Koller WC, Lyons KE, Truly W. Effect of levodopa treatment for parkinsonism in welders: A double-blind study. Neurology. 2004;62:730–733. doi: 10.1212/01.WNL.0000113726.34734.15. [DOI] [PubMed] [Google Scholar]
- 182.Sadek AH, Rauch R, Schulz PE. Parkinsonism due to Manganism in a Welder. Int J Toxicol. 2003;22:393–401. doi: 10.1177/109158180302200511. [DOI] [PubMed] [Google Scholar]
- 183.Cordova F, Aguiar A, Jr., Peres T, Lopes M, Gonçalves F, Pedro D, Lopes S, Pilati C, Prediger RS, Farina M, et al.: Manganese-exposed developing rats display motor deficits and striatal oxidative stress that are reversed by Trolox. Arch Toxicol 2013:1–14. [DOI] [PMC free article] [PubMed]
- 184.Milatovic D, Gupta RC, Yu Y, Zaja-Milatovic S, Aschner M. Protective effects of antioxidants and anti-inflammatory agents against manganese-induced oxidative damage and neuronal injury. Toxicol Appl Pharmacol. 2011;256:219–226. doi: 10.1016/j.taap.2011.06.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Marreilha dos Santos AP, Santos D, Au C, Milatovic D, Aschner M, Batoreu MC: Antioxidants prevent the cytotoxicity of manganese in RBE4 cells. Brain Res 2008, 1236:200–205. [DOI] [PubMed]
- 186.Stephenson AP, Schneider JA, Nelson BC, Atha DH, Jain A, Soliman KF, et al. Manganese-induced oxidative DNA damage in neuronal SH-SY5Y cells: attenuation of thymine base lesions by glutathione and N-acetylcysteine. Toxicol Lett. 2013;218:299–307. doi: 10.1016/j.toxlet.2012.12.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Martins EN, Pessano NTC, Leal L, Roos DH, Folmer V, Puntel GO, et al. Protective effect of Melissa officinalis aqueous extract against Mn-induced oxidative stress in chronically exposed mice. Brain Res Bull. 2012;87:74–79. doi: 10.1016/j.brainresbull.2011.10.003. [DOI] [PubMed] [Google Scholar]
- 188.Chtourou Y, Trabelsi K, Fetoui H, Mkannez G, Kallel H, Zeghal N. Manganese induces oxidative stress, redox state unbalance and disrupts membrane bound ATPases on murine neuroblastoma cells in vitro: protective role of silymarin. Neurochem Res. 2011;36:1546–1557. doi: 10.1007/s11064-011-0483-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Chtourou Y, Fetoui H, Garoui El M, Boudawara T, Zeghal N. Improvement of cerebellum redox states and cholinergic functions contribute to the beneficial effects of silymarin against manganese-induced neurotoxicity. Neurochem Res. 2012;37:469–479. doi: 10.1007/s11064-011-0632-x. [DOI] [PubMed] [Google Scholar]
- 190.Chtourou Y, Garoui El M, Boudawara T, Zeghal N. Protective role of silymarin against manganese-induced nephrotoxicity and oxidative stress in rat. Environ Toxicol. 2014;29:1147–1154. doi: 10.1002/tox.21845. [DOI] [PubMed] [Google Scholar]
- 191.Chtourou Y, Fetoui H, Sefi M, Trabelsi K, Barkallah M, Boudawara T, et al. Silymarin, a natural antioxidant, protects cerebral cortex against manganese-induced neurotoxicity in adult rats. Bio Metals. 2010;23:985–996. doi: 10.1007/s10534-010-9345-x. [DOI] [PubMed] [Google Scholar]
- 192.Lebda MA, El-Neweshy MS, El-Sayed YS. Neurohepatic toxicity of subacute manganese chloride exposure and potential chemoprotective effects of lycopene. Neurotoxicology. 2012;33:98–104. doi: 10.1016/j.neuro.2011.12.008. [DOI] [PubMed] [Google Scholar]
- 193.Farina M, Avila DS, da Rocha JB, Aschner M. Metals, oxidative stress and neurodegeneration: a focus on iron, manganese and mercury. Neurochem Int. 2013;62:575–594. doi: 10.1016/j.neuint.2012.12.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Oboh G, Rocha JB. Hot Pepper (Capsicum spp.) protects brain from sodium nitroprusside- and quinolinic acid-induced oxidative stress in vitro. J Med Food. 2008;11:349–355. doi: 10.1089/jmf.2007.341. [DOI] [PubMed] [Google Scholar]
- 195.Pereira RP, Fachinetto R, de Souza PA, Puntel RL, Santos Da Silva GN, Heinzmann BM, et al. Antioxidant effects of different extracts from Melissa officinalis, Matricaria recutita and Cymbopogon citratus. Neurochem Res. 2009;34:973–983. doi: 10.1007/s11064-008-9861-z. [DOI] [PubMed] [Google Scholar]
- 196.Hanasaki Y, Ogawa S, Fukui S. The Correlation between Active Oxygens Scavenging and Antioxidative Effects of Flavonoids. Free Radic Biol Med. 1994;16:845–850. doi: 10.1016/0891-5849(94)90202-X. [DOI] [PubMed] [Google Scholar]
- 197.Nachtman JP, Delor S, Brennan CE. Manganese neurotoxicity: effects of varying oxygen tension and EDTA on dopamine auto-oxidation. Neurotoxicology. 1987;8:249–253. [PubMed] [Google Scholar]
- 198.De Paris P, Caroldi S. In vivo inhibition of serum dopamine-beta-hydroxylase by CaNa2 EDTA injection. Hum Exp Toxicol. 1994;13:253–256. doi: 10.1177/096032719401300405. [DOI] [PubMed] [Google Scholar]
- 199.Kosal MF, Boyle AJ. Ethylenediaminetetraacetic acid in manganese poisoning of rats; a preliminary study. Ind Med Surg. 1956;25:1–3. [PubMed] [Google Scholar]
- 200.Crossgrove J, Zheng W. Manganese toxicity upon overexposure. NMR Biomed. 2004;17:544–553. doi: 10.1002/nbm.931. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201.Huang CC, Chu NS, Lu CS, Wang JD, Tsai JL, Tzeng JL, et al. Chronic manganese intoxication. Arch Neurol. 1989;46:1104–1106. doi: 10.1001/archneur.1989.00520460090018. [DOI] [PubMed] [Google Scholar]
- 202.Nogueira CW, Zeni G, Rocha JB. Organoselenium and organotellurium compounds: toxicology and pharmacology. Chem Rev. 2004;104:6255–6285. doi: 10.1021/cr0406559. [DOI] [PubMed] [Google Scholar]
- 203.Salgueiro WG, Xavier MCDF, Duarte LFB, Câmara DF, Fagundez DA, Soares ATG, et al. Direct synthesis of 4-organylsulfenyl-7-chloro quinolines and their toxicological and pharmacological activities in Caenorhabditis elegans. Eur J Med Chem. 2014;75:448–459. doi: 10.1016/j.ejmech.2014.01.037. [DOI] [PubMed] [Google Scholar]
- 204.Wollenhaupt SGN, Soares AT, Salgueiro WG, Noremberg S, Reis G, Viana C, et al. Seleno- and Telluro-xylofuranosides attenuate Mn-induced toxicity in C. elegans via the DAF-16/FOXO pathway. Food Chem Toxicol. 2014;64:192–199. doi: 10.1016/j.fct.2013.11.030. [DOI] [PubMed] [Google Scholar]
- 205.Santos D, Batoreu MC, Aschner M, Marreilha Dos Santos A. Comparison Between 5-Aminosalicylic Acid (5-ASA) and Para-Aminosalicylic Acid (4-PAS) as Potential Protectors Against Mn-Induced Neurotoxicity. Biol Trace Elem Res. 2013;152:113–116. doi: 10.1007/s12011-012-9597-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 206.Erikson K, Aschner M. Manganese Causes Differential Regulation of Glutamate Transporter (GLAST) Taurine Transporter and Metallothionein in Cultured Rat Astrocytes. NeuroToxicol. 2002;23:595–602. doi: 10.1016/S0161-813X(02)00012-8. [DOI] [PubMed] [Google Scholar]
- 207.Sidoryk-Wegrzynowicz M, Lee E, Mingwei N, Aschner M. Disruption of astrocytic glutamine turnover by manganese is mediated by the protein kinase C pathway. Glia. 2011;59:1732–1743. doi: 10.1002/glia.21219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 208.Lee E, Sidoryk-Wegrzynowicz M, Farina M, Rocha JB, Aschner M. Estrogen attenuates manganese-induced glutamate transporter impairment in rat primary astrocytes. Neurotox Res. 2013;23:124–130. doi: 10.1007/s12640-012-9347-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 209.Lee E, Sidoryk-Wegrzynowicz M, Yin Z, Webb A, Son DS, Aschner M. Transforming growth factor-alpha mediates estrogen-induced upregulation of glutamate transporter GLT-1 in rat primary astrocytes. Glia. 2012;60:1024–1036. doi: 10.1002/glia.22329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 210.Pawlak J, Brito V, Kuppers E, Beyer C. Regulation of glutamate transporter GLAST and GLT-1 expression in astrocytes by estrogen. Brain Res Mol Brain Res. 2005;138:1–7. doi: 10.1016/j.molbrainres.2004.10.043. [DOI] [PubMed] [Google Scholar]
- 211.Karki P, Webb A, Zerguine A, Choi J, Son DS, Lee E. Mechanism of raloxifene-induced upregulation of glutamate transporters in rat primary astrocytes. Glia. 2014;62:1270–1283. doi: 10.1002/glia.22679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 212.Ramona Cristina HM, Gabriel HM, Petru N, Radu S, Adina N, Ducu S. The monitoring of mineral elements content in fruit purchased in supermarkets and food markets from Timisoara, Romania. Ann Agric Environ Med AAEM. 2014;21:98–105. [PubMed] [Google Scholar]
- 213.Williams AB, Ayejuyo OO, Ogunyale AF. Trace metal levels in fruit juices and carbonated beverages in Nigeria. Environ Monit Assess. 2009;156:303–306. doi: 10.1007/s10661-008-0485-1. [DOI] [PubMed] [Google Scholar]
- 214.Lei B, Chen L, Hao Y, Cao T, Zhang X, Yu Y, et al. Trace elements in animal-based food from Shanghai markets and associated human daily intake and uptake estimation considering bioaccessibility. Ecotoxicol Environ Saf. 2013;96:160–167. doi: 10.1016/j.ecoenv.2013.06.032. [DOI] [PubMed] [Google Scholar]
- 215.Gouda AA. Cloud point extraction, preconcentration and spectrophotometric determination of trace amount of manganese (II) in water and food samples. Spectrochim Acta A Mol Biomol Spectrosc. 2014;131:138–144. doi: 10.1016/j.saa.2014.04.075. [DOI] [PubMed] [Google Scholar]
- 216.Bae YJ, Kim MH, Lee JH, Choi MK. Analysis of six elements (Ca, Mg, Fe, Zn, Cu, and Mn) in several wild vegetables and evaluation of their intakes based on Korea National Health and Nutrition Examination Survey 2010–2011. Biol Trace Elem Res. 2015;164:114–121. doi: 10.1007/s12011-014-0203-5. [DOI] [PubMed] [Google Scholar]
- 217.Nedzarek A, Torz A, Karakiewicz B, Clark JS, Laszczynska M, Kaleta A, et al. Concentrations of heavy metals (Mn, Co, Ni, Cr, Ag, Pb) in coffee. Acta Biochim Pol. 2013;60:623–627. [PubMed] [Google Scholar]
- 218.Micic R, Mitic S, Arsic B, Jokic A, Mitic M, Kostic D, et al. Statistical characteristics of selected elements in vegetables from Kosovo. Environ Monit Assess. 2015;187:389. doi: 10.1007/s10661-015-4606-3. [DOI] [PubMed] [Google Scholar]
- 219.Lonnerdal B. Nutritional Aspects of Soy Formula. Acta Paediatr. 1994;83:105–108. doi: 10.1111/j.1651-2227.1994.tb13371.x. [DOI] [PubMed] [Google Scholar]
- 220.Liu F, Wang WX. Linking trace element variations with macronutrients and major cations in marine mussels Mytilus edulis and Perna viridis. Environ Toxicol Chem. 2015;34:2041–2050. doi: 10.1002/etc.3027. [DOI] [PubMed] [Google Scholar]
- 221.Lutfullah G, Khan AA, Amjad AY, Perveen S: Comparative study of heavy metals in dried and fluid milk in Peshawar by atomic absorption spectrophotometry. ScientificWorldJournal 2014, 2014:715845. [DOI] [PMC free article] [PubMed]
- 222.Luis G, Rubio C, Gutierrez AJ, Gonzalez-Weller D, Revert C, Hardisson A. Evaluation of metals in several varieties of sweet potatoes (Ipomoea batatas L.): comparative study. Environ Monit Assess. 2014;186:433–440. doi: 10.1007/s10661-013-3388-8. [DOI] [PubMed] [Google Scholar]
- 223.Rahman MA, Rahman MM, Reichman SM, Lim RP, Naidu R. Heavy metals in Australian grown and imported rice and vegetables on sale in Australia: health hazard. Ecotoxicol Environ Saf. 2014;100:53–60. doi: 10.1016/j.ecoenv.2013.11.024. [DOI] [PubMed] [Google Scholar]
- 224.Barbosa JT, Santos CM, Peralva VN, Flores EM, Korn M, Nobrega JA, et al. Microwave-assisted diluted acid digestion for trace elements analysis of edible soybean products. Food Chem. 2015;175:212–217. doi: 10.1016/j.foodchem.2014.11.092. [DOI] [PubMed] [Google Scholar]
- 225.Otaka A, Hokura A, Nakai I. Determination of trace elements in soybean by X-ray fluorescence analysis and its application to identification of their production areas. Food Chem. 2014;147:318–326. doi: 10.1016/j.foodchem.2013.09.142. [DOI] [PubMed] [Google Scholar]
- 226.Cockell KA, Bonacci G, Belonje B. Manganese content of soy or rice beverages is high in comparison to infant formulas. J Am Coll Nutr. 2004;23:124–130. doi: 10.1080/07315724.2004.10719352. [DOI] [PubMed] [Google Scholar]
- 227.Rodriguez-Iruretagoiena A, Trebolazabala J, Martinez-Arkarazo I, de Diego A, Madariaga JM. Metals and metalloids in fruits of tomatoes (Solanum lycopersicum) and their cultivation soils in the Basque Country: concentrations and accumulation trends. Food Chem. 2015;173:1083–1089. doi: 10.1016/j.foodchem.2014.10.133. [DOI] [PubMed] [Google Scholar]
- 228.Galani-Nikolakaki S, Kallithrakas-Kontos N, Katsanos AA. Trace element analysis of Cretan wines and wine products. Sci Total Environ. 2002;285:155–163. doi: 10.1016/S0048-9697(01)00912-3. [DOI] [PubMed] [Google Scholar]
- 229.He L, Girijashanker K, Dalton TP, Reed J, Li H, Soleimani M, et al. ZIP8, Member of the Solute-Carrier-39 (SLC39) Metal-Transporter Family: Characterization of Transporter Properties. Mol Pharmacol. 2006;70:171–180. doi: 10.1124/mol.106.024521. [DOI] [PubMed] [Google Scholar]
- 230.Himeno S, Yanagiya T, Fujishiro H. The role of zinc transporters in cadmium and manganese transport in mammalian cells. Biochimie. 2009;91:1218–1222. doi: 10.1016/j.biochi.2009.04.002. [DOI] [PubMed] [Google Scholar]
- 231.Girijashanker K, He L, Soleimani M, Reed JM, Li H, Liu Z, et al. Slc39a14 Gene Encodes ZIP14, A Metal/Bicarbonate Symporter: Similarities to the ZIP8 Transporter. Mol Pharmacol. 2008;73:1413–1423. doi: 10.1124/mol.107.043588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 232.Benedetto A, Au C, Avila DS, Milatovic D, Aschner M: Extracellular dopamine potentiates Mn-induced oxidative stress, lifespan reduction, and dopaminergic neurodegeneration in a BLI-3-dependent manner in Caenorhabditis elegans. . PLoS Genet 2010, 6 (8). [DOI] [PMC free article] [PubMed]
- 233.Hübner CA, Jentsch TJ. Ion channel diseases. Hum Mol Genet. 2002;11:2435–2445. doi: 10.1093/hmg/11.20.2435. [DOI] [PubMed] [Google Scholar]
- 234.Crossgrove JS, Yokel RA. Manganese distribution across the blood–brain barrier: IV. Evidence for brain influx through store-operated calcium channels. Neuro Toxicol. 2005;26:297–307. doi: 10.1016/j.neuro.2004.09.004. [DOI] [PubMed] [Google Scholar]
- 235.Lockman PR, Roder KE, Allen DD. Inhibition of the rat blood–brain barrier choline transporter by manganese chloride. J Neurochem. 2001;79:588–594. doi: 10.1046/j.1471-4159.2001.00589.x. [DOI] [PubMed] [Google Scholar]
- 236.Mellott TJ, Kowall NW, Lopez-Couiella I, Blusztajn JK: Prenatal choline deficiency increases choline transporter expression in the septum and hippocampus during postnatal development and in adulthood in rats. Brain Res 2007, 1151:1–11. [DOI] [PMC free article] [PubMed]
- 237.Crossgrove JS, Allen DD, Bukaveckas BL, Rhineheimer SS, Yokel RA. Manganese Distribution Across the Blood–Brain Barrier: I. Evidence for Carrier-Mediated Influx of Manganese Citrate as Well as Manganese and Manganese Transferrin. Neuro Toxicol. 2003;24:3–13. doi: 10.1016/s0161-813x(02)00089-x. [DOI] [PubMed] [Google Scholar]
- 238.Nota B, Struys EA, Pop A, Jansen EE, Ojeda MRF, Kanhai WA, et al. Deficiency in SLC25A1, Encoding the Mitochondrial Citrate Carrier, Causes Combined D-2-and L-2-Hydroxyglutaric Aciduria. Am J Hum Genet. 2013;92:627–631. doi: 10.1016/j.ajhg.2013.03.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 239.Wysokinski D, Danisz K, Pawlowska E, Dorecka M, Romaniuk D, Robaszkiewicz J, et al. Transferrin receptor levels and polymorphism of its gene in age-related macular degeneration. Acta Biochim Pol. 2015;62:177–184. doi: 10.18388/abp.2014_843. [DOI] [PubMed] [Google Scholar]
- 240.Ponka P, Lok CN. The transferrin receptor: role in health and disease. Int J Biochem Cell Biol. 1999;31:1111–1137. doi: 10.1016/S1357-2725(99)00070-9. [DOI] [PubMed] [Google Scholar]
- 241.Leitch S, Feng M, Muend S, Braiterman LT, Hubbard AL and Rao R. (2011). Vesicular distribution of Secretory Pathway Ca2+-ATPase isoform 1 and a role in manganese detoxification in liver-derived polarized cells. BioMetals 24(1), 159-170. doi:10.1007/s10534-010-9384-3. [DOI] [PMC free article] [PubMed]
- 242.Podhajska A, Musso A, Trancikova A, Stafa K, Moser R, et al. (2012) Common Pathogenic Effects of Missense Mutations in the P-Type ATPase ATP13A2 (PARK9) Associated with Early-Onset Parkinsonism. PLoS ONE 7(6): e39942. doi:10.1371/journal.pone.0039942 [DOI] [PMC free article] [PubMed]
- 243.Behrens MI, Bruggemann N, Chana P, Venegas P, Kagi M, Parrao T, et al. Clinical Spectrum of Kufor-Rakeb Syndrome in the Chilean Kindred with ATP13A2 Mutations. Mov Disord. 2010;25:1929–1937. doi: 10.1002/mds.22996. [DOI] [PubMed] [Google Scholar]
- 244.van Veen S, Sørensen DM, Holemans T, Holen HW, Palmgren MG and Vangheluwe P. (2014). Cellular function and pathological role of ATP13A2 and related P-type transport ATPases in Parkinson's disease and other neurological disorders. Frontiers in Molecular Neuroscience 7(48). doi:10.3389/fnmol.2014.00048. [DOI] [PMC free article] [PubMed]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Not applicable.