

Abstract
Endothelial dysfunction has been implicated in the pathophysiology of multiple cardiovascular diseases and involves components of both innate and acquired immune mechanisms. Identifying signature patterns and targets associated with endothelial dysfunction can help in the development of novel nanotherapeutic platforms for treatment of vascular diseases. This review discusses nucleic acid-based regulation of endothelial function and the different nucleic acid-based nanotherapeutic approaches designed to target endothelial dysfunction in cardiovascular disorders.
Keywords: endothelium, nucleic acids, nanoparticle delivery, cardiovascular diseases, CVD, inflammation, oxidative stress, DNA damage, miRNA, oligonucleotide-based therapeutics
Introduction
Cardiovascular diseases (CVDs) are the leading cause of mortality in the United States and account for one of every three deaths, more than all forms of cancer combined.1 It is well established that endothelial dysfunction is a major determinant of the initiation and/or progression of several CVDs, including atherosclerosis, hypertension, and heart failure.2,3 A healthy and metabolically active endothelium has several pivotal autocrine, paracrine, and endocrine functions that help to maintain vascular homeostasis.3 Thus, restoring impaired vascular homeostasis by improving endothelial function can serve as a promising therapeutic target in the treatment of several cardiovascular disorders. There are many classes of drugs under investigation or already approved for the treatment of CVDs, including: (A) antihypertensive drug classes such as angiotensin-converting enzyme inhibitors, angiotensin AT1 receptors blockers, beta-blockers, and calcium channel blockers;4 (B) nitric oxide (NO) donors and endothelial NO synthase enhancers (substrates, cofactors, and transcription enhancers);5 (C) anti-inflammatory drugs;6 (D) statins with their pleiotropic properties;7 (E) phosphodiesterase 5 inhibitors;8 and (F) other cardiovascular drugs reviewed elsewhere.4,9,10 Whereas these drugs may improve endothelial function and are useful in reducing the progression of CVD, it is likely that novel endothelial-targeted therapies can further improve cardiovascular care.
Nucleic acid-based therapies are well suited for targeting endothelial dysfunction in CVD given their ability to reprogram and/or restore gene function. However, several biological obstacles limit both the delivery and stability of genetic material.11 Barriers to efficacious delivery of nucleic acids include circulating nucleases, the cytoplasmic membrane, endosomal sequestration, and degradation in the enzyme-rich and highly acidic lysosomal compartment, cytoplasmic nucleases, and, in the case of plasmid DNA, penetration through the nuclear membrane. Viral vectors have major disadvantages for clinical use, including the actual or potential integration into the host genome, immunogenicity, biological variability, and manufacturing complexities for clinical products. Accordingly, substantial research has focused on the development of non-viral nanoplatforms based on lipids and polymers, inorganic iron oxide, and gold nanoparticles.12–15 Nanotherapeutics for delivery of nucleic acids prove promising given the ability of the carriers to efficiently package the material; in addition, the multi-functionality of the platforms allow them to incorporate targeting ligands and chemistries that enable rapid endosomal escape and other useful attributes. In this review, we discuss the role of nucleic acids in vascular regulation and dysfunction and potential nanotherapeutics that may restore endothelial function.
Pathophysiology of Endothelial Dysfunction
The endothelial monolayer lining the systemic vascular system maintains vascular homeostasis. Its heterogeneity across multiple vascular beds allows dynamic and organ-specific regulation of blood flow, vascular structure, permeability, and interaction with circulating blood elements.16 In the arterial circulation, this interface usually promotes an anti-inflammatory, antithrombotic, and quiescent state of the vessel wall that is critical for cardiovascular health.10 The endothelium releases vasorelaxant molecules such as nitric oxide (NO), prostacyclin, adrenomedullin, and endothelium-derived hyperpolarizing factors (EDHFs) as well as endothelium-derived constricting factors (EDCFs) such as endothelin.10 In most conduit arteries, the vasodilators predominate, releasing in response to the tractive force of blood flow and maintaining the vascular smooth muscle in a relaxed state. Notably, these vasodilators also suppress proliferation of vascular smooth muscle cells and prevent the adherence of immune cells and platelets.10
All known cardiovascular risk factors (e.g., hypercholesterolemia, diabetes mellitus) are associated with impaired endothelial function characterized by a shift in the balance between endothelial vasodilators and constrictors. While decreased NO bioavailability is considered a hallmark of endothelial dysfunction (also known as endothelial activation), there are also other pathological consequences, including increased expression of adhesion molecules and cytokines; altered vascular permeability through a decrease in gap junction proteins, which facilitates the transendothelial migration of immune cells (monocytes, macrophages, and platelets) and macromolecules (lipoproteins) across the vessel wall; altered vessel reactivity and vasospasm capable of inducing angina and myocardial infarction; increased production of reactive oxygen and nitrosative species; increased production of vascular growth factors that stimulate the proliferation of underlying smooth muscle cells; and an altered thrombotic/fibrinolytic balance that leads to increased deposition of thrombin, fibrin, and extracellular matrix remodeling and increased platelet aggregation.17–20 Any alteration in the functions of the strategically located endothelial monolayer disrupts vascular homeostasis and contributes to the development and progression of various cardiovascular disorders such as atherosclerosis.21
Role of Nucleic Acid Construct in Regulating Endothelial Functions
Endothelial dysfunction/activation involves elements of both innate and adaptive immunity, which may occur as an acute or chronic inflammatory response. The innate immunity-derived surveillance mechanisms play a vital role in triggering inflammation in CVD.22 These mechanisms involve specialized pattern recognition receptors (PRRs) and their response to pathogen-associated molecular patterns (PAMPs) and endogenous damage-associated molecular patterns (DAMPs). The precise nucleic acid-sensing pathways involve PRRs belonging to four major receptor families that include the membrane-based toll-like receptors (TLRs).23 These TLRs work synergistically with specific receptors that identify the DAMPs and PAMPs within vasculature and elicit an activated, prothrombotic, and proinflammatory response. This in turn leads to further endothelial damage and may induce epigenetic changes that can cause phenotypic alterations.22–25 Since exogenous nucleic acids can stimulate the PRRs and trigger endothelial activation and inflammation, nucleic acid therapies must be cloaked by nanoparticle delivery systems as described below.
DAMPs, PAMPs, and Endothelial Dysfunction
The endothelium-based recognition of different DAMPs and PAMPs (Figure 1) and subsequent cellular responses are an integral component of the innate immune response in endothelial dysfunction.22 An activated endothelium produces a variety of substances, including proinflammatory cytokines from the interleukin (IL) family; immune cell-attracting chemokines; vascular endothelial (VEGF), platelet-derived, and fibroblast growth factors (FGF); and adhesion molecules such as intercellular adhesion molecule-1 (ICAM-1) and P-selectin. All of these substances contribute to endothelium-related vascular injury.22,26 In addition, as a response to various stimuli, endothelial cells (ECs) act as semiprofessional antigen-presenting cells by expressing the major histocompatibility complex class II and costimulatory molecules, which induce migration of lymphocytes across endothelium to underlying tissue at the site of vascular injury.22 Several research groups have reported that oxidative injury-induced DAMPs primarily drive the innate-immune response associated with a dysfunctional endothelium.25 Recent studies have reported altered mitochondrial dynamics and endoplasmic reticulum-induced stress as factors initiating EC activation.27,28 Alterations in the function of these organelles may modulate endothelial nitric oxide synthase (eNOS) activity, alter barrier function, and prompt endothelial cell migration and angiogenesis.28 Thus, impaired organelles act as potential triggers (presenting endothelial DAMPs) that generate excessive reactive oxygen species (ROS), which can increase the expression of endothelial adhesion molecules and inflammatory cytokines.29 To varying degrees, the activation of the endothelium may be offset by the endothelial elaboration of anti-inflammatory cytokines such as IL-10.30 It is the shift in balance between the secreted proinflammatory and anti-inflammatory cytokines that modulates the inflammatory cascade associated with dysfunctional endothelium.30
Figure 1.
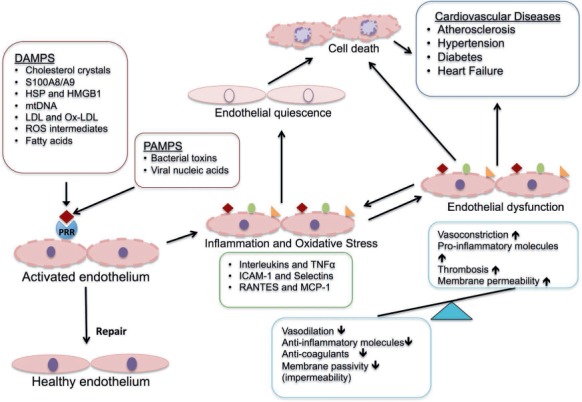
Pathophysiological role of PAMPs and DAMPS in endothelial dysfunction. PAMPs and DAMPS activate the endothelial cells through a pattern recognition receptor (PRR)-based interaction that triggers the initial inflammatory response. In the absence of endothelial repair, the inflammation leads to a proinflammatory and prothrombotic response manifested by several cardiovascular diseases. PAMPS: pathogen associated molecular patterns; DAMPS: damage associated molecular patterns; S100A8/A9: S100 calcium-binding protein A8 or A9; HSP: heat shock proteins; HMGB1: high mobility group box 1 protein; mtDNA: mitochondrial DNA; LDL: low-density lipoprotein; Ox-LDL: oxidized low-density lipoprotein; ROS: reactive oxygen species
DNA Damage and Repair in Vascular Disease
Oxidative stress-induced damage to nuclear and mitochondrial DNA also contributes to endothelial dysfunction.31 Various forms of DNA damage have been reported in vascular diseases, including single-strand and double-strand breaks, telomere dysfunction, and base-pair modifications such as oxidation, deletion, and mispairing.32 Based on the type of DNA damage and stage of the cell cycle, vascular cells employ the different DNA repair pathways such as base excision repair, nucleotide excision repair, nonhomologous end joining, and homologous recombination in an attempt to correct the damaged DNA.32 While there is evidence highlighting the presence of both DNA damage and repair in vascular disorders, it is the ineffective repair that leads to growth arrest, cell senescence, and apoptosis, all of which promote further disease progression by activating the inflammatory cascade.32 Interestingly, mitochondrial DNA (mtDNA), in circulation as well as in the cellular cytosol, induces genomic DNA damage and subsequent endothelial dysfunction, which is implicated in a host of vascular diseases like diabetes, hypertension, and atherosclerosis.29 Endothelial mitochondria are endowed with significant DNA repair capacity. However, mitochondrial proximity with the oxidative respiratory chain and the absence of histone or chromatin structures around the mitochondria render it susceptible to oxidative damage, making it a potential presenter of DAMPs and a source of endothelial dysfunction.29 Thus, therapeutic strategies designed to prevent DNA damage or induce/enhance DNA repair mechanisms, particularly focused on inducers like mtDNA, can serve as potential targets for treating endothelial dysfunction-mediated vascular disorders.
MicroRNA and Their Role in Vascular Homeostasis
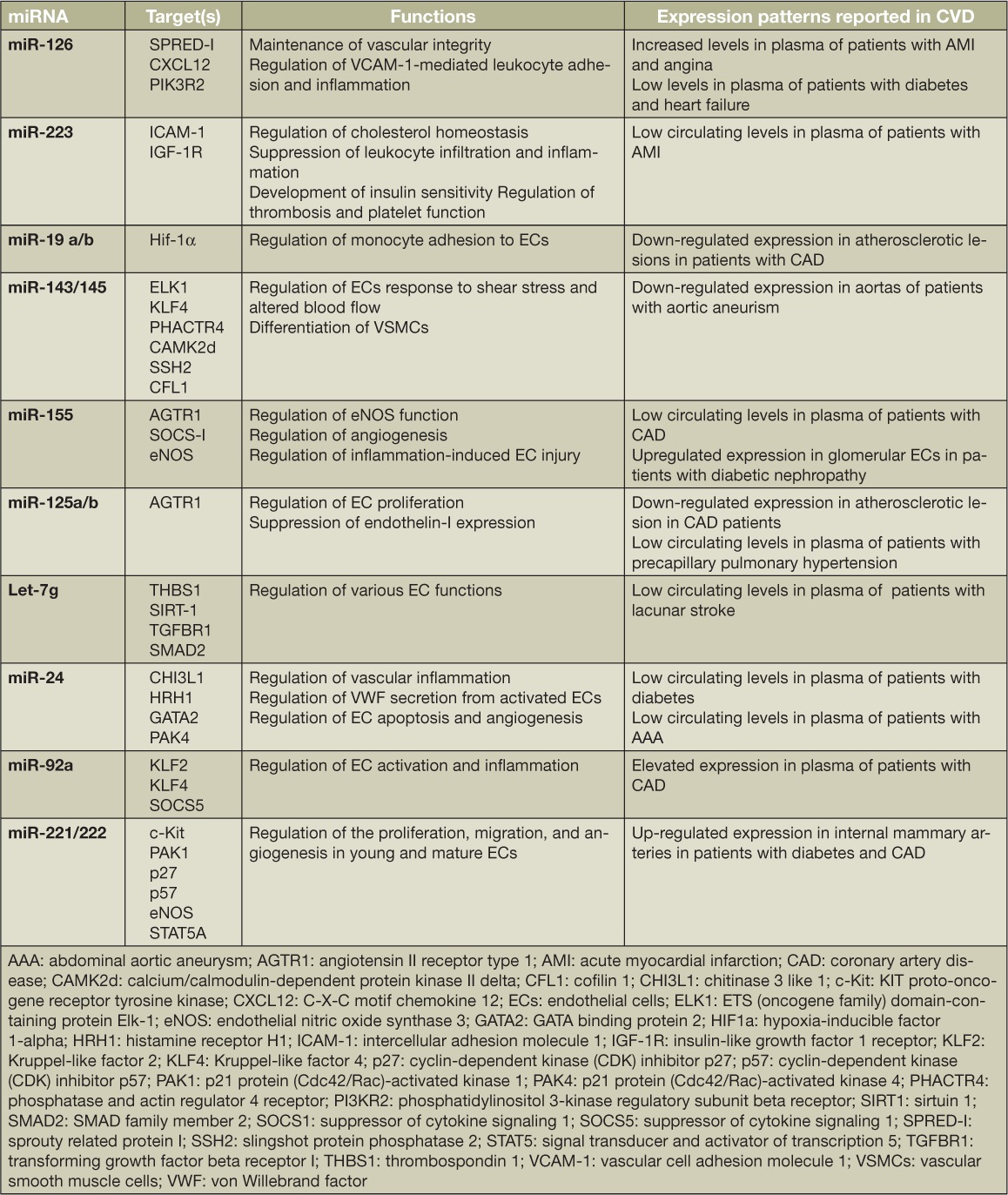
MicroRNAs (miRNAs) play a pivotal role in vascular development and function by regulating the expression of genes through post-transcriptional modification or translational repression. Dicer, a key enzyme involved in the biosynthesis and processing of mi-RNAs, also plays a vital role in the regulatory functions of ECs.33 Endothelium-specific inactivation of this enzyme is associated with altered expression of vascular endothelial growth factor (VEGF), eNOS, IL-8, and the angiopoietin cell surface receptors Tie-I and Tie-2, compromising endothelial function.34 In addition, miRNAs also serve as paracrine ligands of the toll-like receptors, an exosome-mediated activity that is independent of post-transcriptional regulatory function.35 Several miRNAs have been identified to play an important role in regulating endothelial function in normal as well as pathological conditions (Table 1).36 Of the miRNAs studied to date, miR-126 is considered the key regulator of vascular function.33 With low expression during senescence and overexpression during re-endothelialization and angiogenesis, miR-126 specifically targets regulators of proangiogenic VEGF and the angiopoietin pathway that helps to maintain vascular integrity and inhibit endothelial activation.33 Variable expression patterns of circulating miR-126 have been reported in several vascular disorders such as myocardial infarction, angina (miR-126 up-regulation), diabetes, and heart failure (miR-126 down-regulation), among others.33 Similarly, a recent study reported miR-19a as a functional link between endothelial dysfunction, risk factors such as hyperlipidemia and inflammation, and the ensuing atherosclerosis.37 On the whole, endothelial miRNAs play a major role in regulating several key endothelial functions, including monitoring immune cell trafficking and inflammation, regulating EC senescence, regulating endothelial progenitor cell biology, maintaining endothelial barrier function, and regulating the vascular tone of blood vessels.36 Therefore, targeting miRNAs and their expression patterns paves the way for a novel platform for treating vascular disorders.
Table 1.
Examples of microRNAs regulating endothelial function and their expression pattern in various cardiovascular diseases.
Nucleic Acid-Based Therapeutic Delivery Strategies for Vascular Diseases
The advent of nanotechnology has led to the development of several nanoformulations capable of site-specific drug delivery. These can serve as robust platforms for treating endothelial dysfunction in cardiovascular diseases. One formulation by our own group used an omega-3-polyunsaturated fatty acid (PUFA)-rich oil-in-water nanoemulsion system selectively targeting fibrin clots within the plaque region to deliver 17-β-estradiol (17-βE) in a murine model of atherosclerosis.38 This formulation combined the pleiotropic cardio- and vasoprotective properties of PUFAs and 17-βE with the targeting efficiency of the fibrin-binding CREKA peptide, leading to a relatively high targeting efficiency and maximum exposure/accumulation of 17-βE within plasma and tissues.39 In vitro delivery to cultured ECs led to an increased production of nitric oxide, while in vivo delivery in apolipoprotein-E knock out (ApoE−/−) mice fed a high-fat diet decreased the levels of circulating lipids, reduced gene expression of inflammatory markers associated with atherosclerosis, and reduced vascular lesions.38
It is important to note that the success of conventional and investigational therapeutic molecules is limited by their palliative nature and may not restore a healthy vascular endothelium. Endothelial dysfunction is characterized by several overlapping pathological components, each of which is associated with a unique nucleic acid expression pattern, opening several avenues for genetic therapies. As nucleic acids have limited in vivo stability and poor permeability across cellular membranes, adequate delivery of these molecules to disease sites is a major hurdle limiting their therapeutic applications. In addition, the exogenous nucleic acids must be delivered in a manner that reduces their activation of nucleic acid-sensing PRRs. Thus, several nucleic acid-based nanotherapeutics are being developed to treat vascular disorders, with the endothelium serving as one of the most prominent targets. The following section describes advances in nanoparticle-mediated delivery of genetic material targeting endothelial dysfunction in CVD as well as emerging strategies (e.g., ASOs, aptamers) in which nanoparticles may potentially result in efficacious clinical translation.
DNA-Based Therapeutics
DNA-based therapeutics that deliver angiogenic factors are one of the most promising strategies in the area of cardiovascular gene therapy. Among the several angiogenic factors studied, the initial gene therapy trials evaluated the effect of angiopoietins, FGF, HIF-α, and VEGF-A in the treatment of either ischemia-induced vascular conditions or peripheral occlusive disorders.40 The VEGF family of growth factors is crucial in regulating several endothelial functions including vasodilation and nitric oxide release, recruitment of circulating progenitor cells, regulation of vascular permeability, and endothelial proliferation and migration,41 making it particularly well-suited for gene therapy targeting endothelial dysfunction.
In terms of gene delivery platforms, adenoviral vectors, adeno-associated viral vectors, and lentiviral vectors have been used to translate DNA-based therapeutic strategies, with lentiviral vectors capable of specifically targeting endothelial dysfunction.40 An example of DNA-based gene therapy using proangiogenic VEGF is the ongoing KAT301 clinical trial.40 This trial is evaluating the safety and efficacy of the adenoviral vector-mediated delivery of VEGF-D, a mature form of VEGF, delivered using the NOGA-mediated transendocardial injection system in patients with severe coronary heart disease (NOGA® XP Cardiac Navigation System, Biologics Delivery Systems Group, Cordis Corporation, Miami Lakes, FL).42 Preliminary results at 3 months demonstrated increased myocardial perfusion with intramyocardial delivery of VEGF-D in segments with the lowest perfusion reserve, thus justifying a phase II/III evaluation.42 Rather than targeting the vasculature, another approach is to modulate the immune system. Oral administration of a DNA vaccine against either CD99 or VEG-FR2-induced antigen-specific CD8+T cells reduces vascular lesions in murine models of atherosclerosis.43
A major disadvantage is that viral vectors can activate PRRs to aggravate endothelial activation and inflammation. Accordingly, such DNA-based therapeutics could potentially be facilitated by a nanoparticle delivery system. These include liposome-DNA complexes, polymer-based DNA complexes such as dendrimers, and lipid-polymer complexes in which condensed DNA is encapsulated within lipid-based or polymeric delivery systems.44 Paul and coworkers developed hydrogel-containing nanocomplexes of graphene oxide (GO) and VEGF-165 DNA for direct injection into infarcted areas in the myocardium.45 The hydrogel was capable of modulating the release kinetics of the DNA nanocomplexes, resulting in decreases in both the scar area fraction and the loss of ejection fraction after an acute myocardial infarction (MI) in a rodent model.
RNA-Based Therapeutics
Over the past two decades, several small interfering RNA (si-RNA) and miRNA-based therapeutics have been developed for the treatment of CVD. RNA interference (RNAi) strategies via delivery of siRNA prove especially efficient at specific silencing of gene expression.46 Endothelial adhesion molecules and other receptors mediating inflammatory response serve as excellent targets for developing endothelium-targeted siRNA-based therapeutics.47 An example of a reasonable target is CD40, a glycoprotein expressed on the surface of endothelial and immune cells that mediates endothelial activation. In one study, RNAi against CD40 was delivered using a viral vector to induce the regression of atherosclerotic lesions in apolipoprotein E-deficient (ApoE−/−) mice. In this case, a recombinant CD40-RNAi-lentivirus system was used for knockdown of CD40 signaling, leading to reduced expression of proinflammatory cytokines and chemokines.48
Similar to DNA-based therapeutics, siRNA-based nanoparticle platforms, such as liposome-RNA complexes or polymer-based RNA complexes,49 have been successful in preclinical studies of various cardiovascular disorders. Liposomal nanoparticles (cationic amphiphiles) invested with antibodies against either VCAM-1 or E-selectin can deliver siRNA to activated primary ECs in vitro, which might be an approach for preferentially targeting activated endothelium.50 Polymeric constructs have been widely used to conjugate siRNA. Kim and coworkers successfully reduced ischemia-reperfusion injury by knockdown of a receptor for advanced glycation end-products (RAGE) following direct injection into infarcted areas in a rat MI model.51 In this study, siRNA targeting the RAGE mRNA was conjugated with deoxycholic acid-modified polyethylenimine, successfully silencing RAGE expression and leading to reduced levels of inflammatory cytokines and apoptosis in vivo as well as reduced ventricular remodeling.
Given their role in regulating EC function, miRNAs and their manipulation provide a well-suited strategy for treating endothelial dysfunction. miRNA-based therapeutics focus on reinstating dysregulated miRNA expression associated with the disease pathology using either miRNA mimics or antagomirs.52 Individual miRNAs are capable of modulating multifactorial disease pathways, targeting more than one molecular target, thereby increasing the efficacy of miRNA-based therapies.53 Transfection of activated ECs with miR-17-3p mimic and miR-31 mimic reduces the expression of adhesion molecules (ICAM-1 and E-selectin), thereby decreasing neutrophil adhesion to ECs.54 Another study demonstrated that hepatic overexpression of miR-30c reduced hyperlipidemia and attenuated vascular lesions in the hypercholesterolemic mouse, thus highlighting the potential of miR-30c mimics in treating atherosclerosis.55
Oligonucleotide-Based Therapeutics
Oligonucleotide-based therapeutics such as antisense oligonucleotides (ASOs), anti-microRNA oligonucleotides (AMOs), and aptamers have emerged as powerful tools in the treatment of various disorders including vascular diseases.56 ASOs are highly selective therapeutics that bind to their complementary RNA substrates via Watson-Crick hybridization. Furthermore, ASOs inactivate RNA by blocking translation or, more commonly, by destabilizing the RNA-DNA complex, which is then destroyed by ribonuclease H1, a ubiquitously present cellular endonuclease.57 ASO-mediated mRNA termination allows precise targeting of disease-associated protein expression, making these therapeutics more specific than most conventional drugs.57 The gapmer design was used to develop second-generation ASOs to improve the stability against nuclease-based degradation and the proinflammatory characteristics of ASOs.57 Mipomersen sodium, the first antisense drug approved by the U.S. Food and Drug Administration, is a second-generation gapmer ASO that targets the apolipoprotein B (apoB) mRNA and is used as an adjunct to lipid-lowering agents prescribed to patients with familial hypercholesterolemia.57 Other examples of clinically investigated ASOs include SPC5001, APoC-III, and BMS-844421 that primarily target apoB, apoC-III, and PCSK9 protein expression, respectively.57
ASOs that target miRNA function are termed anti-miRNA oligonucleotides (AMOs). The application of AMOs in treating CVDs is currently limited to preclinical evaluation in small animals and non-human primates. A recent study demonstrated that AMOs targeting miR-33a and miR-33b increased plasma high-density lipoprotein (HDL) cholesterol levels by modulating the expression of the ATP-binding cassette transporter ABCA1.58 Since ABCA1 is associated with the biosynthesis and transport of HDLs, targeting the miR-33 family with AMOs has the potential of reversing hyper-lipidemia and the subsequent development of atherosclerosis.58 For improved delivery and reduced non-specific effects, the ASOs and AMOs undergo various modifications; for example, the addition of locked nucleic acids, conjugation with cholesterol or GalNAc (N-acetylgalactosamine) sugar, 2′-O-methylation of the antisense oligonucleotide, chemical modification using the non-nucleotide naphthyl-azo group (ZEN) chemical modifier or steric blockers, and lastly, insertion of either pre-miRNA sequences or tandem miRNA target sites into the expression vectors to create decoy binding sites.59 Jo and coworkers successfully inhibited atherosclerosis in apoE−/− mice by delivering an AMO (anti-miR-712) via a selective multifunctional lipoparticle conjugated to a peptide to target vascular cell adhesion molecule 1 (VCAM1).60 The resulting nanoparticle inhibited miR-712 in only the affected organs with no off-target effects.
Another oligonucleotide-based therapeutic approach involves the use of aptamers for treatment of vascular disorders. These molecules are generated through systematic evolution of ligands by exponential enrichment (SELEX) and can be classified as DNA/RNA aptamers and peptide aptamers.61 Their ease of synthesis and scale-up, thermal stability, and lack of immunogenicity makes them a powerful class of therapeutic ligands.61 Aptamers are designed to alter the expression of disease-associated molecular targets involved in the pathology of various disorders. Currently, aptamer-based therapeutics targeting the von Willebrand factor (vWF), thrombin, and factor IX are undergoing clinical trials and being developed as antithrombotics or anticoagulants to treat patients with acute coronary syndromes.61 vWF is synthesized by ECs and endothelial connective tissue and plays an active role in vascular homeostasis, thrombus formation, and platelet activation.61 Circulating levels of vWF represent predictive biomarkers of endothelial injury. An aptamer developed by Archemix Corp. (Cambridge, MA), named ARC1779, inhibits vWF and platelet interaction and binding, preventing thrombus formation associated with endothelial activation.61 Similarly, nucleic acid-based aptamers are also being developed for other cardiovascular molecular targets, such as platelet-derived growth factor, integrins, P-selectin, and CXCL10, all of which are expressed during endothelial dysfunction.61
Concluding Remarks
Endothelial dysfunction has emerged as the underlying cause of several cardiovascular disorders. Targeting endothelial dysfunction at the cellular level can lead to innovative strategies aimed at reducing or inhibiting the progression of cardiovascular disease. Our review illustrates how manipulating the vascular endothelium through nucleic acid-based therapeutics involving nanoparticle-based platforms can serve as a promising strategy for the treatment of atherosclerosis, hypertension, myocardial infarction, and other cardiovascular disorders. Research with large, randomized, and adequately powered clinical trials will help determine the success of these therapeutics in targeting endothelial dysfunction.
Acknowledgments
We are deeply grateful to Houston Methodist Research Institute and Northeastern University for establishing collaborations and financial support. Additionally, this work was generously supported by funding from the George and Angelina Kostas Research Center for Cardiovascular Nanomedicine at Houston Methodist.
Key Points
Endothelial dysfunction is a major contributor to the pathogenesis of cardiovascular diseases.
Nucleic acid constructs such as genes, siRNA, and microRNA can reprogram the vascular endothelium to restore active functions.
Nanotechnology-based targeted delivery systems can be engineered to efficiently deliver nucleic acid cargo to the vascular endothelial cells.
Footnotes
Conflict of Interest Disclosure: This work was supported by funding from the George and Angelina Kostas Research Center for Cardiovascular Nanomedicine at Houston Methodist.
References
- 1. Mozaffarian D, Benjamin EJ, Go AS, . et al. Heart Disease and Stroke Statistics-2016 Update: A Report From the American Heart Association. Circulation. 2016. January 26; 133( 4): e38– e360. [DOI] [PubMed] [Google Scholar]
- 2. Lam CS, Brutsaert DL. Endothelial dysfunction: a pathophysiologic factor in heart failure with preserved ejection fraction. J Am Coll Cardiol. 2012. October 30; 60( 18): 1787– 9. [DOI] [PubMed] [Google Scholar]
- 3. Sena CM, Pereira AM, Seica R.. Endothelial dysfunction - a major mediator of diabetic vascular disease. Biochim Biophys Acta. 2013. December; 1832( 12): 2216– 31. [DOI] [PubMed] [Google Scholar]
- 4. Su JB. Vascular endothelial dysfunction and pharmacological treatment. World J Cardiol. 2015. November 26; 7( 11): 719– 41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Zhang Y, Janssens SP, Wingler K, Schmidt HH, Moens AL.. Modulating endothelial nitric oxide synthase: a new cardiovascular therapeutic strategy. Am J Physiol Heart Circ Physiol. 2011. September; 301( 3): H634– 46. [DOI] [PubMed] [Google Scholar]
- 6. Bäck M, Hansson GK. Anti-inflammatory therapies for atherosclerosis. Nat Rev Cardiol. 2015. April; 12( 4): 199– 211. [DOI] [PubMed] [Google Scholar]
- 7. Beckman JA, Creager MA. The nonlipid effects of statins on endothelial function. Trends Cardiovasc Med. 2006. July; 16( 5): 156– 62. [DOI] [PubMed] [Google Scholar]
- 8. Schwartz BG, Jackson G, Stecher VJ, Campoli-Richards DM, Kloner RA.. Phosphodiesterase type 5 inhibitors improve endothelial function and may benefit cardiovascular conditions. Am J Med. 2013. March; 126( 3): 192– 9. [DOI] [PubMed] [Google Scholar]
- 9. Deshpande DD, Janero DR, Amiji MM.. Therapeutic strategies for endothelial dysfunction. Expert Opin Biol Ther. 2011. December; 11( 12): 1637– 54. [DOI] [PubMed] [Google Scholar]
- 10. Cooke JP. Flow, NO, and atherogenesis. Proc Natl Acad Sci U S A. 2003. February 4; 100( 3): 768– 70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Blanco E, Shen H, Ferrari M.. Principles of nanoparticle design for overcoming biological barriers to drug delivery. Nat Biotechnol. 2015. September; 33( 9): 941– 51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Dobson J. Gene therapy progress and prospects: magnetic nanoparticle-based gene delivery. Gene Ther. 2006. February; 13( 4): 283– 7. [DOI] [PubMed] [Google Scholar]
- 13. Pack DW, Hoffman AS, Pun S, Stayton PS.. Design and development of polymers for gene delivery. Nat Rev Drug Discov. 2005. July; 4( 7): 581– 93. [DOI] [PubMed] [Google Scholar]
- 14. Pissuwan D, Niidome T, Cortie MB.. The forthcoming applications of gold nanoparticles in drug and gene delivery systems. J Control Release. 2011. January 5; 149( 1): 65– 71. [DOI] [PubMed] [Google Scholar]
- 15. Torchilin VP. Recent advances with liposomes as pharmaceutical carriers. Nat Rev Drug Discov. 2005. February; 4( 2): 145– 60. [DOI] [PubMed] [Google Scholar]
- 16. Aird WC. Endothelial cell heterogeneity. Cold Spring Harb Perspect Med. 2012. January; 2( 1): a006429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Chiu JJ, Chien S. Effects of disturbed flow on vascular endothelium: pathophysiological basis and clinical perspectives. Physiol Rev. 2011. January; 91( 1): 327– 87. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Higashi Y, Noma K, Yoshizumi M, Kihara Y.. Endothelial function and oxidative stress in cardiovascular diseases. Circ J. 2009. March; 73( 3): 411– 8. [DOI] [PubMed] [Google Scholar]
- 19. Deanfield JE, Halcox JP, Rabelink TJ.. Endothelial function and dysfunction: testing and clinical relevance. Circulation. 2007. March 13; 115( 10): 1285– 95. [DOI] [PubMed] [Google Scholar]
- 20. Vita JA. Endothelial function. Circulation. 2011. December 20; 124( 25): e906– 12. [DOI] [PubMed] [Google Scholar]
- 21. Rajendran P, Rengarajan T, Thangavel J, . et al. The vascular endothelium and human diseases. Int J Biol Sci. 2013. November 9; 9( 10): 1057– 69. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Yang XF, Yin Y, Wang H.. Vascular inflammation and atherogenesis are activated via receptors for PAMPs and suppressed by regulatory T cells. Drug Discov Today Ther Strateg. 2008; 5( 2): 125– 142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Zimmer S, Steinmetz M, Asdonk T, . et al. Activation of endothelial toll-like receptor 3 impairs endothelial function. Circ Res. 2011. May 27; 108( 11): 1358– 66. [DOI] [PubMed] [Google Scholar]
- 24. Lee J, Sayed N, Hunter A, . et al. Activation of innate immunity is required for efficient nuclear reprogramming. Cell. 2012. October 26; 151( 3): 547– 58 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Witztum JL, Lichtman AH. The influence of innate and adaptive immune responses on atherosclerosis. Annu Rev Pathol. 2014; 9: 73– 102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Davignon J, Ganz P. Role of endothelial dysfunction in atherosclerosis. Circulation. June 15 2004; 109( 23 Suppl 1): III27– 32. [DOI] [PubMed] [Google Scholar]
- 27. Galán M, Kassan M, Kadowitz PJ, Trebak M, Belmadani S, Matrougui K.. Mechanism of endoplasmic reticulum stress-induced vascular endothelial dysfunction. Biochim Biophys Acta. 2014. June; 1843( 6): 1063– 75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Kluge MA, Fetterman JL, Vita JA.. Mitochondria and endothelial function. Circ Res. 2013. April 12; 112( 8): 1171– 88. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Wenceslau CF1, McCarthy CG, Szasz T, Spitler K, Goulopoulou S, Webb RC; Working Group on DAMPs in Cardiovascular Disease. . Mitochondrial damage-associated molecular patterns and vascular function. Eur Heart J. 2014. May; 35( 18): 1172– 7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Shao Y, Cheng Z, Li X, Chernaya V, Wang H, Yang XF.. Immuno-suppressive/anti-inflammatory cytokines directly and indirectly inhibit endothelial dysfunction--a novel mechanism for maintaining vascular function. J Hematol Oncol. 2014. October 31; 7: 80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Mikhed Y, Daiber A, Steven S.. Mitochondrial Oxidative Stress, Mitochondrial DNA Damage and Their Role in Age-Related Vascular Dysfunction. Int J Mol Sci. 2015. July 13; 16( 7): 15918– 53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Uryga A, Gray K, Bennett M.. DNA Damage and Repair in Vascular Disease. Annu Rev Physiol. 2016. February 10; 78: 45– 66. [DOI] [PubMed] [Google Scholar]
- 33. Santulli G. MicroRNAs and Endothelial (Dys) Function. J Cell Physiol. 2016. August; 231( 8): 1638– 44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Wronska A, Kurkowska-Jastrzebska I, Santulli G.. Application of microRNAs in diagnosis and treatment of cardiovascular disease. Acta Physiol (Oxf). 2015. January; 213( 1): 60– 83. [DOI] [PubMed] [Google Scholar]
- 35. Chen X, Liang H, Zhang J, Zen K, Zhang CY.. MicroRNAs are ligands of Toll-like receptors. RNA. 2013. June; 19( 6): 737– 9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Chamorro-Jorganes A, Araldi E, Suárez Y.. MicroRNAs as pharmacological targets in endothelial cell function and dysfunction. Pharmacol Res. 2013. September; 75: 15– 27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Akhtar S, Hartmann P, Karshovska E, . et al. Endothelial Hypoxia-Inducible Factor-1α Promotes Atherosclerosis and Monocyte Recruitment by Upregulating MicroRNA-19a. Hypertension. 2015. December; 66( 6): 1220– 6. [DOI] [PubMed] [Google Scholar]
- 38. Deshpande D, Kethireddy S, Janero DR, Amiji MM.. Therapeutic Efficacy of an omega-3-Fatty Acid-Containing 17-beta Estradiol Nano-Delivery System against Experimental Atherosclerosis. PLoS One. 2016. February 3; 11( 2): e0147337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Deshpande D, Kethireddy S, Gattacceca F, Amiji M.. Comparative pharmacokinetics and tissue distribution analysis of systemically administered 17-β-estradiol and its metabolites in vivo delivered using a cationic nanoemulsion or a peptide-modified nanoemulsion system for targeting atherosclerosis. J Control Release. 2014. April 28; 180: 117– 24. [DOI] [PubMed] [Google Scholar]
- 40. Rincon MY, VandenDriessche T, Chuah MK.. Gene therapy for cardiovascular disease: advances in vector development, targeting, and delivery for clinical translation. Cardiovasc Res. 2015. October 1; 108( 1): 4– 20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Taimeh Z, Loughran J, Birks EJ, Bolli R.. Vascular endothelial growth factor in heart failure. Nat Rev Cardiol. 2013. September; 10( 9): 519– 30. [DOI] [PubMed] [Google Scholar]
- 42. Hassinen I, Hartikainen J, Hedman A, . et al. Abstract 11987: Adenoviral intramyocardial VEGF-D gene transfer increases myocardial perfusion in refractory angina patients. Circulation. 2015. November 10; 132( Suppl 3): A11987. [Google Scholar]
- 43. van Wanrooij EJ, de Vos P, Bixel MG, Vestweber D, van Berkel TJ, Kuiper J.. Vaccination against CD99 inhibits atherogenesis in low-density lipoprotein receptor-deficient mice. Cardiovasc Res. 2008. June 1; 78( 3): 590– 6. [DOI] [PubMed] [Google Scholar]
- 44. Nayerossadat N, Maedeh T, Ali PA.. Viral and nonviral delivery systems for gene delivery. Adv Biomed Res. 2012; 1: 27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Paul A, Hasan A, Kindi HA, . et al. Injectable graphene oxide/hydrogel-based angiogenic gene delivery system for vasculogenesis and cardiac repair. ACS Nano. 2014. August 26; 8( 8): 8050– 62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Poller W, Tank J, Skurk C, Gast M.. Cardiovascular RNA interference therapy: the broadening tool and target spectrum. Circ Res. 2013. August 16; 113( 5): 588– 602. [DOI] [PubMed] [Google Scholar]
- 47. Pradhan-Nabzdyk L, Huang C, LoGerfo FW, Nabzdyk CS.. Current siRNA targets in atherosclerosis and aortic aneurysm. Discov Med. 2014. May; 17( 95): 233– 46. [PMC free article] [PubMed] [Google Scholar]
- 48. Wang B, Qian H, Yang H, Xu L, Xu W, Yan J.. Regression of atherosclerosis plaques in apolipoprotein E−/− mice after lentivirus-mediated RNA interference of CD40. Int J Cardiol. 2013. February 10; 163( 1): 34– 9. [DOI] [PubMed] [Google Scholar]
- 49. Koenig O, Walker T, Perle N, . et al. New aspects of gene-silencing for the treatment of cardiovascular diseases. Pharmaceuticals (Basel). 2013. July 19; 6( 7): 881– 914. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Kowalski PS, Lintermans LL, Morselt HW, . et al. Anti-VCAM-1 and anti-E-selectin SAINT-O-Somes for selective delivery of siRNA into inflammation-activated primary endothelial cells. Mol Pharm. 2013. August 5; 10( 8): 3033– 44. [DOI] [PubMed] [Google Scholar]
- 51. Hong J, Ku SH, Lee MS, . et al. Cardiac RNAi therapy using RAGE siRNA/deoxycholic acid-modified polyethylenimine complexes for myocardial infarction. Biomaterials. 2014. August; 35( 26): 7562– 73. [DOI] [PubMed] [Google Scholar]
- 52. Nouraee N, Mowla SJ. miRNA therapeutics in cardiovascular diseases: promises and problems. Front Genet. 2015. June 30; 6: 232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Small EM, Olson EN. Pervasive roles of microRNAs in cardiovascular biology. Nature. 2011. January 20; 469( 7330): 336– 42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Suárez Y, Wang C, Manes TD, Pober JS.. Cutting edge: TNF-induced microRNAs regulate TNF-induced expression of E-selectin and intercellular adhesion molecule-1 on human endothelial cells: feedback control of inflammation. J Immunol. 2010. January 1; 184( 1): 21– 5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Soh J, Iqbal J, Queiroz J, Fernandez-Hernando C, Hussain MM.. MicroRNA-30c reduces hyperlipidemia and atherosclerosis in mice by decreasing lipid synthesis and lipoprotein secretion. Nat Med. 2013. July; 19( 7): 892– 900. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. van Rooij E, Kauppinen S. Development of microRNA therapeutics is coming of age. EMBO Mol Med. 2014. June 16; 6( 7): 851– 64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Lee RG, Crosby J, Baker BF, Graham MJ, Crooke RM.. Antisense technology: an emerging platform for cardiovascular disease therapeutics. J Cardiovasc Transl Res. 2013. December; 6( 6): 969– 80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Rayner KJ, Esau CC, Hussain FN, . et al. Inhibition of miR-33a/b in non-human primates raises plasma HDL and lowers VLDL triglycerides. Nature. 2011. October 19; 478( 7369): 404– 7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Philippen LE, Dirkx E, Wit JB, Burggraaf K, de Windt LJ, da Costa Martins PA.. Antisense MicroRNA Therapeutics in Cardiovascular Disease: Quo Vadis? Mol Ther. 2015. December; 23( 12): 1810– 8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Kheirolomoom A, Kim CW, Seo JW, . et al. Multifunctional Nanoparticles Facilitate Molecular Targeting and miRNA Delivery to Inhibit Atherosclerosis in ApoE(−/−) Mice. ACS Nano. 2015. September 22; 9( 9): 8885– 97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Wang P, Yang Y, Hong H, Zhang Y, Cai W, Fang D.. Aptamers as therapeutics in cardiovascular diseases. Curr Med Chem. 2011; 18( 27): 4169– 74. [DOI] [PMC free article] [PubMed] [Google Scholar]