

The transformation of wild animals into domestic ones was a key prerequisite for modern human societies (1). A variety of species have been domesticated, but no species has had such an impact on warfare, transportation, and communication capability as the horse. Despite its importance for human history, the time and place of horse domestication have remained enigmatic for a long time (2, 3). Recently, evidence was found for horse domestication having occurred in Kazakhstan about 5500 years before the present (yr B.P.) (4). Genetic studies have shown that coat color variation arose rapidly during domestication, resulting from human selection (5). To further investigate the dynamics and place of horse domestication, we investigated DNA sequence polymorphisms responsible for coat color variation in fossil horses.
We successfully typed eight mutations in six genes (6) responsible for coat color variation for 89 [out of 152 tested; tables S1 to S5 (6)] ancient samples. To assess coat color variation of predomestic horses, we analyzed the bones of wild horses from the Late Pleistocene and Early Holocene found in Siberia, East and Central Europe, and the Iberian Peninsula. We found no variation in the Siberian and European Pleistocene horses, suggesting that these horses were bay or bay-dun in color. There is evidence that the wild type is dun, but, because the nondun mutation has not yet been identified, we cannot distinguish between dun and nondun horses. In contrast, Iberian Early Holocene horses carried both the nonblack allele A (25%) and the black allele a (75%) at the ASIP gene, resulting in four black and four bay horses. An analysis of 21 Neolithic and Copper Age samples, dating to the millennia before archaeological evidence for domestication exists, produced similar allelic patterns. These periods are characterized by an absence of horse bones in the archaeological record for some regions, which prevents the inclusion of Siberian horses from 8000 to 5000 yr B.P. (table S6). East European horses dating to 7000 to 6000 yr B.P. revealed a mixed population (76% bay and 24% black). All these horses are regarded as wild because they clearly predate the earliest evidence for horse husbandry in this region. Because black horses are found neither in Siberia nor in East and Central Europe during the Late Pleistocene, their occurrence in the Early Holocene is probably the result of postglacial immigration or of selection caused by increased forest cover, as recently shown for wolves (7). Human selection is less likely because we observed no other coloration. A binomial test furthermore shows that, given the sample size of our predomestic horses, the chance that we missed any color allele with a frequency larger than 4% is P ≤ 0.05 (6) (table S7).
In contrast, a rapid and substantial increase in the number of coat colorations is found in both Siberia and East Europe beginning in the fifth millennium B.P. (Fig. 1 and figs. S1 and S2). Although the earliest chestnut allele (MC1R gene) was identified in a Romanian sample from the late seventh millennium B.P., chestnut horses were first observed in Siberia (fifth millenium B.P.). Their prevalence increased rapidly, reaching 28% during the Bronze Age.
Fig. 1.
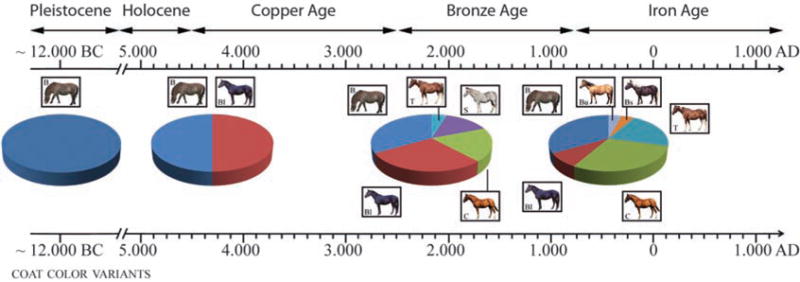
Timeline showing the rapid increase in coat color variation in horses around the Bronze Age. The size of the pie charts corresponds to the frequency of the respective phenotype. B indicates bay color; Bl, black; C, chestnut; T, tobiano; S, sabino; Bu, buckskin; and Bs, black silver. Sample sizes from left to right are 16, 18, 26, and 29 specimens.
Mutations responsible for coat color dilutions or spottings seem to appear later. Cream (buckskin) and (black) silver dilutions (2800 to 2600 yr B.P.) were first observed in Siberia. Sabino is the first spotting phenotype, appearing during the fifth millennium B.P. in Siberia, and present in Armenia and Moldavia during the middle Bronze Age. The Tobiano spotting was first found in a single Eastern European sample (3500 to 3000 yr B.P.) and later also in Asia. Unlike in samples from Siberia and Eastern Europe, we observed no color change in Spanish samples until medieval times.
Although we cannot statistically reject neutrality for four out of six loci tested, the selection coefficient for ASIP and MC1R was found to be significantly different from zero (6, 8); (table S8, and fig. S3). Our results are therefore best explained by selective breeding, which supports domestication as the cause for the observed changes in coloration. Given the rapid change, we conclude that horse domestication started in the Eurasian steppe region around at least 5000 yr B.P.
Supplementary Material
Acknowledgments
We thank D. Lieckfeldt, S. Aksu, M. Meixner, and N. Hensel for technical assistance; K. Finstermeier and D. Blum for help with the figure design; and numerous scientists, horse breeders, and curators, especially A. Morales, who provided material for DNA analyses. This work was funded by the Deutsche Forschungsgemeinschaft (A.L., M.P., M.C., and N.B.), the Max Planck Gesellschaft (S.L. and M.H.), and the Janggen Pöhn Foundation (A.-S.M.). A.-S.M. thanks J. Jensen and O. Malaspinas for very helpful discussions.
Footnotes
Supporting Online Material
www.sciencemag.org/cgi/content/full/324/5926/485/DC1
Materials and Methods
References
References and Notes
- 1.Diamond J. Nature. 2002;418:700. doi: 10.1038/nature01019. [DOI] [PubMed] [Google Scholar]
- 2.Jansen T, et al. Proc Natl Acad Sci USA. 2002;99:10905. doi: 10.1073/pnas.152330099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Vila C, et al. Science. 2001;291:474. doi: 10.1126/science.291.5503.474. [DOI] [PubMed] [Google Scholar]
- 4.Outram AK, et al. Science. 2009;323:1332. doi: 10.1126/science.1168594. [DOI] [PubMed] [Google Scholar]
- 5.Fang M, et al. PLoS Genet. 2009;5:e1000341. doi: 10.1371/journal.pgen.1000341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Materials and methods are available as supporting material on Science Online.
- 7.Anderson TM, et al. Science. 2009;323:1339. doi: 10.1126/science.1165448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Bollback JP, York TL, Nielsen R. Genetics. 2008;179:497. doi: 10.1534/genetics.107.085019. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.